
Effects of a Short-Term Lipopolysaccharides Challenge on Mouse Brain and Liver Peroxisomal Antioxidant and β-oxidative Functions: Protective Action of Argan Oil

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
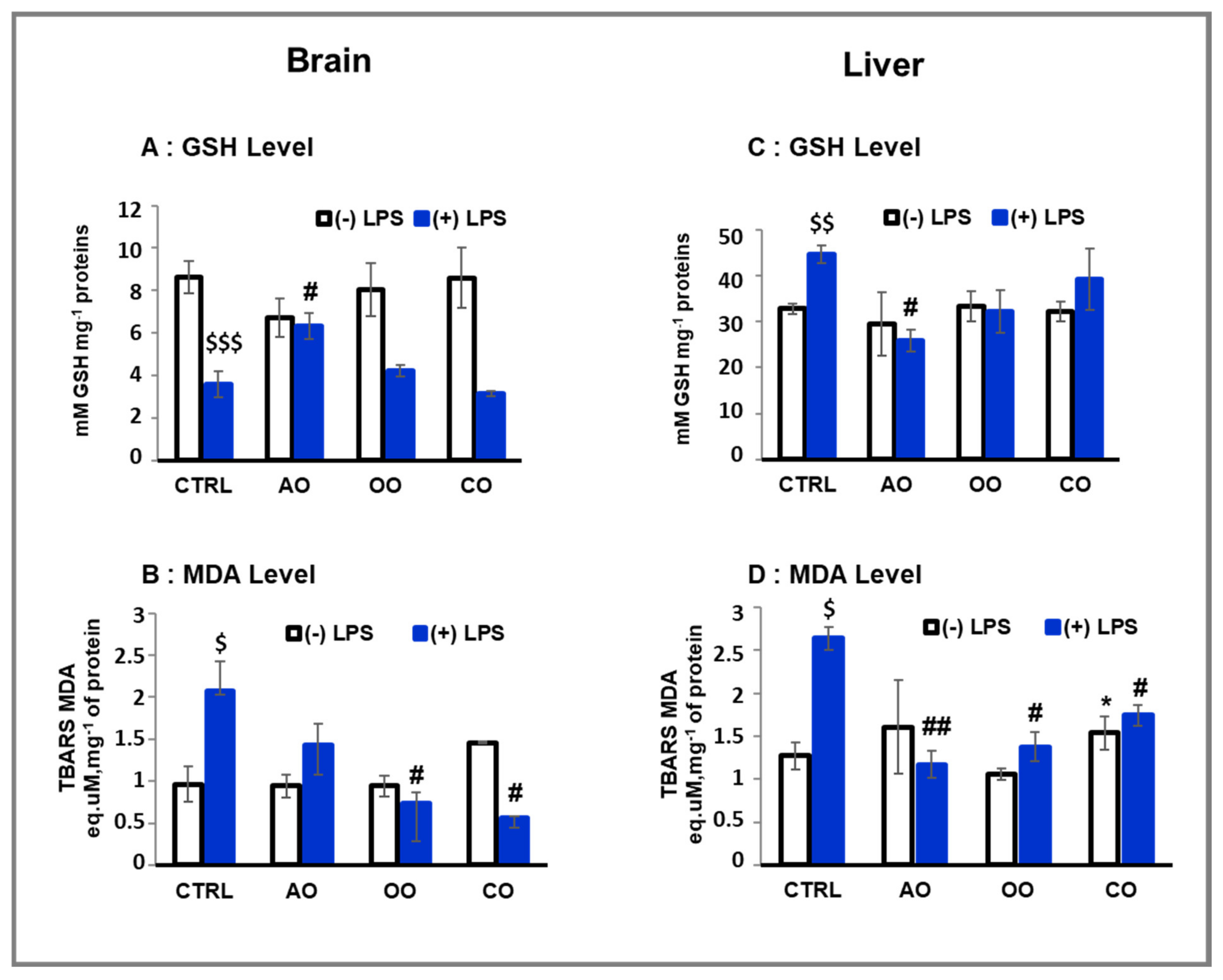
2.1. Oxidative Stress and Inflammatory Biomarkers
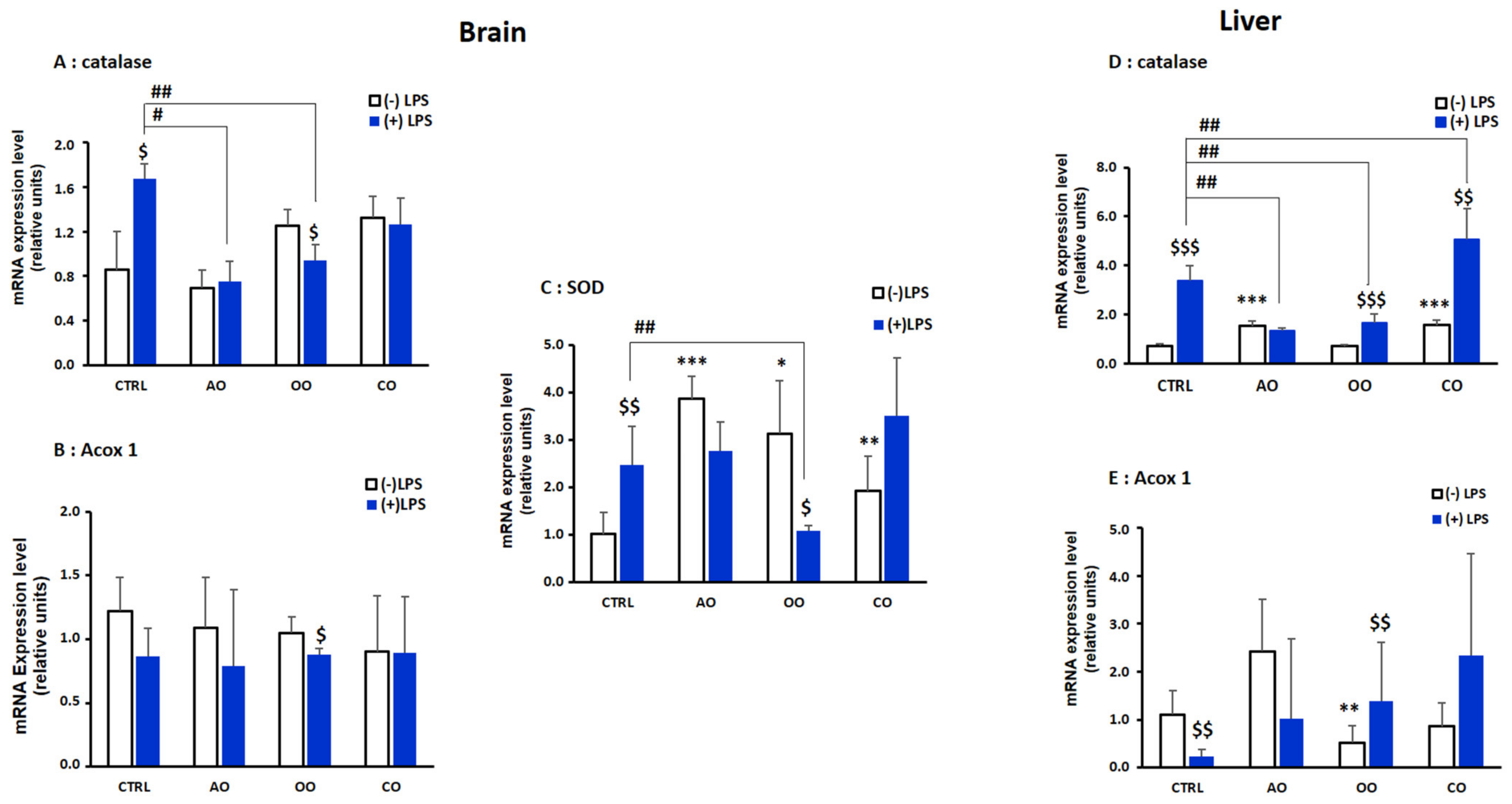
2.2. Brain and Liver Gene Expression of Peroxisomal Protein-Encoding Genes
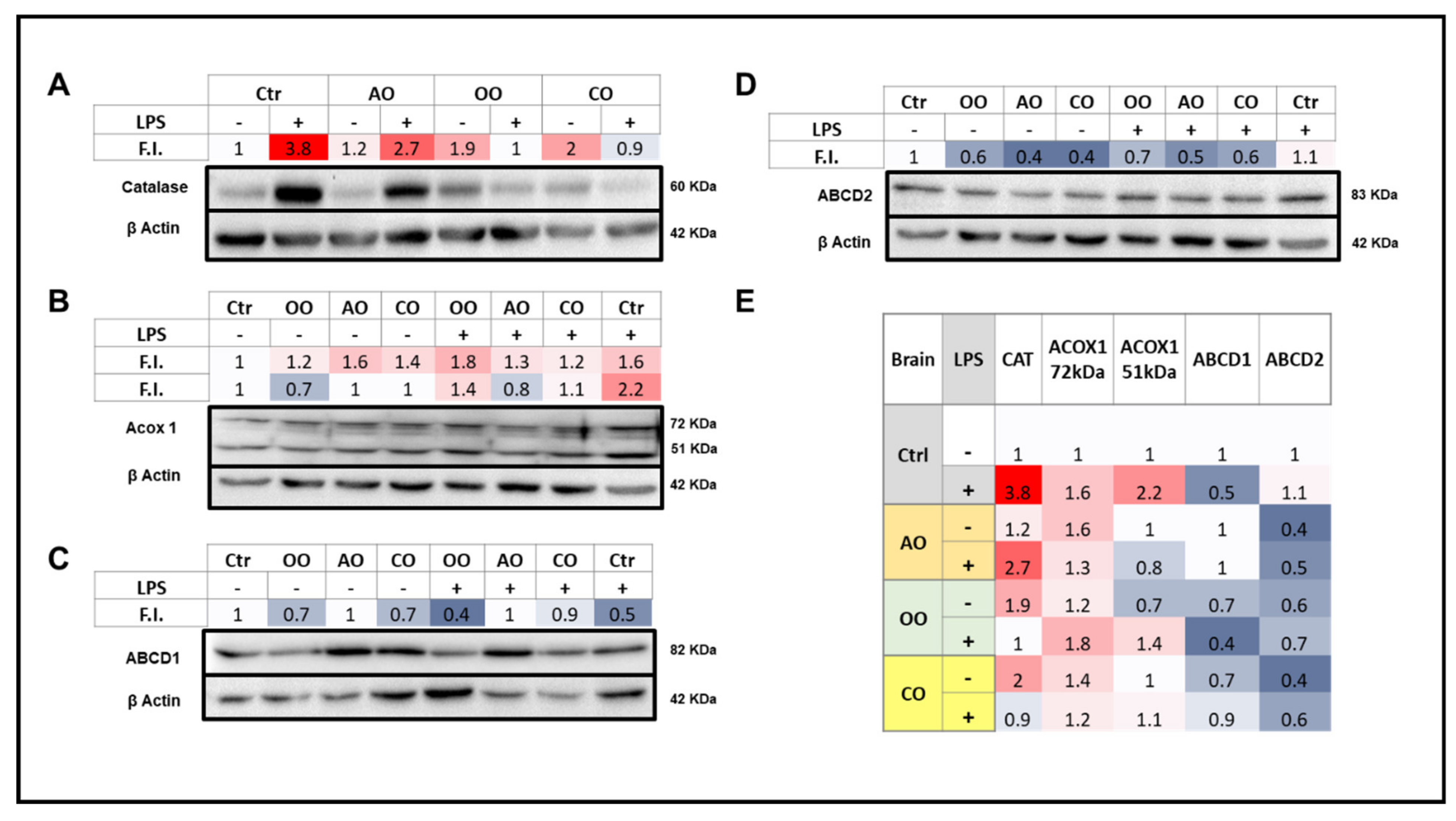
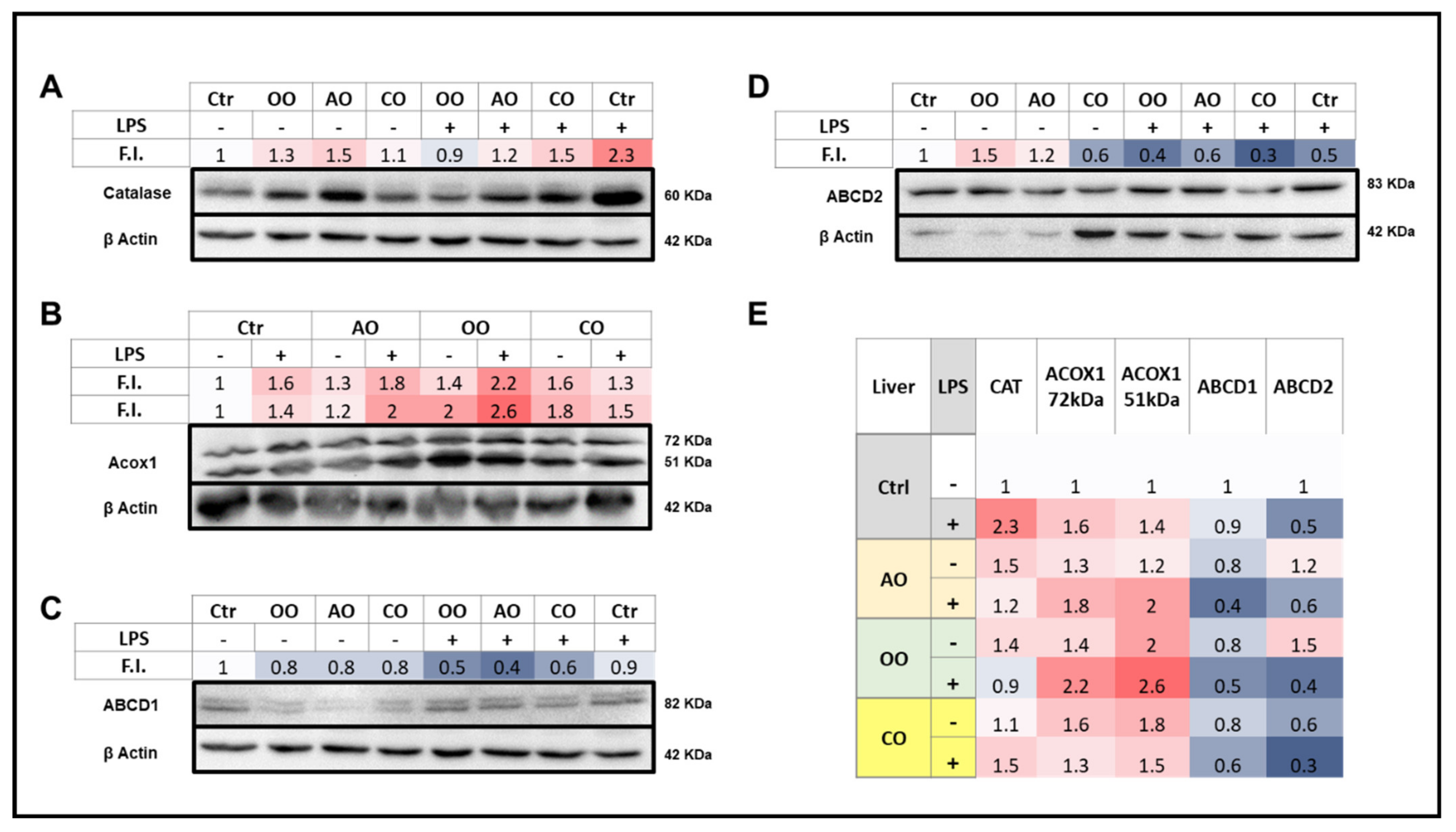
2.3. Brain and Liver Expressions of Peroxisomal Proteins
2.3.1. Brain
2.3.2. Liver
2.4. Brain and Liver Peroxisomal Antioxidant Enzymes Activities
3. Discussion
4. Material and Methods
4.1. Chemicals and Reagents
4.2. Mice Treatments
4.3. Origin, Extraction and Composition of Oils
4.4. Tissue Homogenates Preparation for Enzymatic Activity Assays
4.5. Measurement of Enzymatic Activities
4.5.1. Determination of Catalase Activity
4.5.2. Determination of Superoxide Dismutase Activity
4.5.3. Determination of Glutathione Peroxidase Activity
4.6. Determination of Reduced Glutathione Level
4.7. MDA Level Evaluation
4.8. Quantification of Gene Expression by RT-qPCR
4.9. Immunoblotting
4.10. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharshar, T.; Annane, D.; de la Grandmaison, G.L.; Brouland, J.P.; Hopkinson, N.S.; Françoise, G. The Neuropathology of Septic Shock. Brain Pathol. Zurich Switz. 2004, 14, 21–33. [Google Scholar] [CrossRef]
- Mantzarlis, K.; Tsolaki, V.; Zakynthinos, E. Role of Oxidative Stress and Mitochondrial Dysfunction in Sepsis and Potential Therapies. Oxid. Med. Cell. Longev. 2017, 2017, 5985209. [Google Scholar] [CrossRef] [PubMed]
- Sriskandan, S.; Altmann, D.M. The Immunology of Sepsis. J. Pathol. 2008, 214, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Spolarics, Z. Endotoxemia, Pentose Cycle, and the Oxidant/Antioxidant Balance in the Hepatic Sinusoid. J. Leukoc. Biol. 1998, 63, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Fransen, M.; Lismont, C. Peroxisomes and Cellular Oxidant/Antioxidant Balance: Protein Redox Modifications and Impact on Inter-Organelle Communication. Subcell. Biochem. 2018, 89, 435–461. [Google Scholar] [CrossRef]
- Vamecq, J.; Andreoletti, P.; El Kebbaj, R.; Saih, F.E.; Latruffe, N.; El Kebbaj, M.H.S.; Lizard, G.; Nasser, B.; Cherkaoui-Malki, M. Peroxisomal Acyl-CoA Oxidase Type 1: Anti-Inflammatory and Anti-Aging Properties with a Special Emphasis on Studies with Lps and Argan Oil as a Model Transposable to Aging. Oxid. Med. Cell. Longev. 2018, 2018, 6986984. [Google Scholar] [CrossRef] [Green Version]
- Schrader, M.; Fahimi, H.D. Peroxisomes and Oxidative Stress. Biochim. Biophys. Acta 2006, 1763, 1755–1766. [Google Scholar] [CrossRef] [Green Version]
- Tawbeh, A.; Gondcaille, C.; Trompier, D.; Savary, S. Peroxisomal ABC Transporters: An Update. Int. J. Mol. Sci. 2021, 22, 6093. [Google Scholar] [CrossRef]
- Khan, M.; Contreras, M.; Singh, I. Endotoxin-Induced Alterations of Lipid and Fatty Acid Compositions in Rat Liver Peroxisomes. J. Endotoxin Res. 2000, 6, 41–50. [Google Scholar] [CrossRef]
- El Kamouni, S.; El Kebbaj, R.; Andreoletti, P.; El Ktaibi, A.; Rharrassi, I.; Essamadi, A.; El Kebbaj, M.S.; Mandard, S.; Latruffe, N.; Vamecq, J.; et al. Protective Effect of Argan and Olive Oils against Lps-Induced Oxidative Stress and Inflammation in Mice Livers. Int. J. Mol. Sci. 2017, 18, 2181. [Google Scholar] [CrossRef] [Green Version]
- Charrouf, Z.; Guillaume, D. Ethnoeconomical, Ethnomedical, and Phytochemical Study of Argania spinosa (L.) Skeels. J. Ethnopharmacol. 1999, 67, 7–14. [Google Scholar] [CrossRef]
- Khallouki, F.; Younos, C.; Soulimani, R.; Oster, T.; Charrouf, Z.; Spiegelhalder, B.; Bartsch, H.; Owen, R.W. Consumption of Argan Oil (Morocco) with Its Unique Profile of Fatty Acids, Tocopherols, Squalene, Sterols and Phenolic Compounds Should Confer Valuable Cancer Chemopreventive Effects. Eur. J. Cancer Prev. 2003, 12, 67–75. [Google Scholar] [CrossRef]
- Drissi, A.; Girona, J.; Cherki, M.; Godàs, G.; Derouiche, A.; El Messal, M.; Saile, R.; Kettani, A.; Solà, R.; Masana, L.; et al. Evidence of Hypolipemiant and Antioxidant Properties of Argan Oil Derived from the Argan Tree (Argania spinosa). Clin. Nutr. 2004, 23, 1159–1166. [Google Scholar] [CrossRef]
- Cayuela, J.A.; Rada, M.; Pérez-Camino, M.d.C.; Benaissa, M.; Abdelaziz, E.; Guinda, Á. Characterization of Artisanally and Semiautomatically Extracted Argan Oils from Morocco. Eur. J. Lipid Sci. Technol. 2008, 110, 1159–1166. [Google Scholar] [CrossRef]
- Charrouf, Z.; Guillaume, D. Argan Oil: Occurrence, Composition and Impact on Human Health. Eur. J. Lipid Sci. Technol. 2008, 110, 632–636. [Google Scholar] [CrossRef]
- El Monfalouti, H.; Guillaume, D.; Denhez, C.; Charrouf, Z. Therapeutic Potential of Argan Oil: A Review. J. Pharm. Pharmacol. 2010, 62, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Trichopoulou, A.; Lagiou, P.; Kuper, H.; Trichopoulos, D. Cancer and Mediterranean Dietary Traditions. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2000, 9, 869–873. [Google Scholar]
- El Kebbaj, R.; Andreoletti, P.; El Hajj, H.I.; El Kharrassi, Y.; Vamecq, J.; Mandard, S.; Saih, F.E.; Latruffe, N.; El Kebbaj, M.S.; Lizard, G.; et al. Argan Oil Prevents Down-Regulation Induced by Endotoxin on Liver Fatty Acid Oxidation and Gluconeogenesis and on Peroxisome Proliferator-Activated Receptor Gamma Coactivator-1α, (PGC-1α), Peroxisome Proliferator-Activated Receptor α (PPARα) and Estrogen Receptor α (ERRα). Biochim. Open 2015, 1, 51–59. [Google Scholar] [CrossRef]
- Elmostafi, H.; Bahbiti, Y.; Elhessni, A.; Bousalham, R.; Doumar, H.; Ouichou, A.; Benmhammed, H.; Touil, T.; Mesfioui, A. Neuroprotective Potential of Argan Oil in Neuropsychiatric Disorders in Rats: A Review. J. Funct. Foods 2020, 75, 104233. [Google Scholar] [CrossRef]
- Saber, T.M.; Mansour, M.F.; Abdelaziz, A.S.; Mohamed, R.M.S.; Fouad, R.A.; Arisha, A.H. Argan Oil Ameliorates Sodium Fluoride–Induced Renal Damage via Inhibiting Oxidative Damage, Inflammation, and Intermediate Filament Protein Expression in Male Rats. Environ. Sci. Pollut. Res. 2020, 27, 30426–30436. [Google Scholar] [CrossRef]
- Manfredini, A.; Constantino, L.; Pinto, M.C.; Michels, M.; Burger, H.; Kist, L.W.; Silva, M.C.; Gomes, L.M.; Dominguini, D.; Steckert, A.; et al. Mitochondrial Dysfunction Is Associated with Long-Term Cognitive Impairment in an Animal Sepsis Model. Clin. Sci. 2019, 133, 1993–2004. [Google Scholar] [CrossRef] [PubMed]
- Paintlia, M.K.; Paintlia, A.S.; Contreras, M.A.; Singh, I.; Singh, A.K. Lipopolysaccharide-Induced Peroxisomal Dysfunction Exacerbates Cerebral White Matter Injury: Attenuation by N-Acetyl Cysteine. Exp. Neurol. 2008, 210, 560–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purba, J.H.V. The Analysis of European Union’s Vegetable Oil Consumption: “Will The European Parliament Resolution Halt the Consumption of Crude Palm Oil in the European Union in the Future?”. Int. J. Appl. Bus. Econ. Res. 2017, 15, 27–39. [Google Scholar]
- Chen, Z.; Wu, A.; Jin, H.; Liu, F. β-Sitosterol Attenuates Liver Injury in a Rat Model of Chronic Alcohol Intake. Arch. Pharm. Res. 2020, 43, 1197–1206. [Google Scholar] [CrossRef]
- Oaxaca-Castillo, D.; Andreoletti, P.; Vluggens, A.; Yu, S.; van Veldhoven, P.P.; Reddy, J.K.; Cherkaoui-Malki, M. Biochemical Characterization of Two Functional Human Liver Acyl-CoA Oxidase Isoforms 1a and 1b Encoded by a Single Gene. Biochem. Biophys. Res. Commun. 2007, 360, 314–319. [Google Scholar] [CrossRef] [Green Version]
- Meister, A. Glutathione, Ascorbate, and Cellular Protection. Cancer Res. 1994, 54, 1969s–1975s. [Google Scholar]
- Jaeschke, H. Enhanced Sinusoidal Glutathione Efflux during Endotoxin-Induced Oxidant Stress in Vivo. Am. J. Physiol. 1992, 263, G60–G68. [Google Scholar] [CrossRef]
- Miura, K.; Ishii, T.; Sugita, Y.; Bannai, S. Cystine Uptake and Glutathione Level in Endothelial Cells Exposed to Oxidative Stress. Am. J. Physiol.—Cell Physiol. 1992, 262, C50–C58. [Google Scholar] [CrossRef] [PubMed]
- Rouzer, C.A.; Scott, W.A.; Griffith, O.W.; Hamill, A.L.; Cohn, Z.A. Glutathione Metabolism in Resting and Phagocytizing Peritoneal Macrophages. J. Biol. Chem. 1982, 257, 2002–2008. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.E.; Youness, E.R.; Mohammed, N.A.; Morsy, S.M.Y.; Omara, E.A.; Sleem, A.A. Citric Acid Effects on Brain and Liver Oxidative Stress in Lipopolysaccharide-Treated Mice. J. Med. Food 2014, 17, 588–598. [Google Scholar] [CrossRef] [Green Version]
- Vilhardt, F.; Haslund-Vinding, J.; Jaquet, V.; McBean, G. Microglia Antioxidant Systems and Redox Signalling. Br. J. Pharmacol. 2017, 174, 1719–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinert, K.R.S.; Umphlet, C.D.; Quattlebaum, A.; Boger, H.A. Short-Term Effects of an Endotoxin on Substantia Nigra Dopamine Neurons. Brain Res. 2014, 1557, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, S.; Noack, H.; Possel, H.; Wolf, G. Induction of Nitric Oxide Synthesis Lowers Intracellular Glutathione in Microglia of Primary Glial Cultures. Glia 2000, 29, 98–101. [Google Scholar] [CrossRef]
- López, L.C.; Cabrera-Vique, C.; Venegas, C.; García-Corzo, L.; Luna-Sánchez, M.; Acuña-Castroviejo, D.; Escames, G. Argan Oil-Contained Antioxidants for Human Mitochondria. Nat. Prod. Commun. 2013, 8, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Urata, Y.; Honma, S.; Goto, S.; Todoroki, S.; Iida, T.; Cho, S.; Honma, K.; Kondo, T. Melatonin Induces Gamma-Glutamylcysteine Synthetase Mediated by Activator Protein-1 in Human Vascular Endothelial Cells. Free Radic. Biol. Med. 1999, 27, 838–847. [Google Scholar] [CrossRef]
- Gustaw-Rothenberg, K.; Kowalczuk, K.; Stryjecka-Zimmer, M. Lipids’ Peroxidation Markers in Alzheimer’s Disease and Vascular Dementia. Geriatr. Gerontol. Int. 2010, 10, 161–166. [Google Scholar] [CrossRef]
- Subramanyam, D.; Gurunathan, D.; Gaayathri, R.; Vishnu Priya, V. Comparative Evaluation of Salivary Malondialdehyde Levels as a Marker of Lipid Peroxidation in Early Childhood Caries. Eur. J. Dent. 2018, 12, 67–70. [Google Scholar] [CrossRef]
- Ammari, M.; Othman, H.; Hajri, A.; Sakly, M.; Abdelmelek, H. Pistacia Lentiscus Oil Attenuates Memory Dysfunction and Decreases Levels of Biomarkers of Oxidative Stress Induced by Lipopolysaccharide in Rats. Brain Res. Bull. 2018, 140, 140–147. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Q.; Zhao, G.; Kan, Z.; Wang, X.; Wang, H.; Huang, J.; Wang, T.; Qian, F.; Ho, C.T.; et al. Protective Effect and Mechanism of Theanine on Lipopolysaccharide-Induced Inflammation and Acute Liver Injury in Mice. J. Agric. Food Chem. 2018, 66, 7674–7683. [Google Scholar] [CrossRef]
- Eljaoudi, R.; Elkabbaj, D.; Bahadi, A.; Ibrahimi, A.; Benyahia, M.; Errasfa, M. Consumption of Argan Oil Improves Anti-Oxidant and Lipid Status in Hemodialysis Patients. Phytother. Res. 2015, 29, 1595–1599. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, L.; Yang, F.; Zhang, N.; Fan, J.; Qin, S.; Shao, T.; Xu, X.; Yan, S.; Guo, H.; et al. Antioxidant Activity Assessment of Yingjisha Sweet Almond Oil. Int. J. Food Sci. Technol. 2022, 57, 1773–1781. [Google Scholar] [CrossRef]
- Badreddine, A.; Zarrouk, A.; Karym, E.M.; Debbabi, M.; Nury, T.; Meddeb, W.; Sghaier, R.; Bezine, M.; Vejux, A.; Martine, L.; et al. Argan Oil-Mediated Attenuation of Organelle Dysfunction, Oxidative Stress and Cell Death Induced by 7-Ketocholesterol in Murine Oligodendrocytes 158N. Int. J. Mol. Sci. 2017, 18, 2220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Kishore, R.; McMullen, M.R.; Nagy, L.E. Lipopolysaccharide Stimulation of ERK1/2 Increases TNF-Alpha Production via Egr-1. Am. J. Physiol. Cell Physiol. 2002, 282, C1205–C1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dopp, J.M.; Sarafian, T.A.; Spinella, F.M.; Kahn, M.A.; Shau, H.; de Vellis, J. Expression of the P75 TNF Receptor Is Linked to TNF-Induced NFkappaB Translocation and Oxyradical Neutralization in Glial Cells. Neurochem. Res. 2002, 27, 1535–1542. [Google Scholar] [CrossRef]
- Persson, M.; Sandberg, M.; Hansson, E.; Rönnbäck, L. Microglial Glutamate Uptake Is Coupled to Glutathione Synthesis and Glutamate Release. Eur. J. Neurosci. 2006, 24, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Pils, M.C.; Pisano, F.; Fasnacht, N.; Heinrich, J.-M.; Groebe, L.; Schippers, A.; Rozell, B.; Jack, R.S.; Müller, W. Monocytes/Macrophages and/or Neutrophils Are the Target of IL-10 in the LPS Endotoxemia Model. Eur. J. Immunol. 2010, 40, 443–448. [Google Scholar] [CrossRef]
- Abdelhamid, A.K.; Arafa, M.M. ORIGINAL Propolis Derivatives Inhibit the Systemic Inflammatory Response and Protect Hepatic and Neuronal Cells in Acute Septic Shock. Braz. J. Infect. Dis. 2011, 15, 332–338. [Google Scholar] [CrossRef] [Green Version]
- Raas, Q.; Saih, F.; Gondcaille, C.; Trompier, D.; Hamon, Y.; Leoni, V.; Caccia, C.; Nasser, B.; Jadot, M.; Ménétrier, F.; et al. A Microglial Cell Model for Acyl-CoA Oxidase 1 Deficiency. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 567–576. [Google Scholar] [CrossRef]
- Ivashchenko, O.; Van Veldhoven, P.P.; Brees, C.; Ho, Y.-S.; Terlecky, S.R.; Fransen, M. Intraperoxisomal Redox Balance in Mammalian Cells: Oxidative Stress and Interorganellar Cross-Talk. Mol. Biol. Cell 2011, 22, 1440–1451. [Google Scholar] [CrossRef]
- López-Erauskin, J.; Galino, J.; Bianchi, P.; Fourcade, S.; Andreu, A.L.; Ferrer, I.; Muñoz-Pinedo, C.; Pujol, A. Oxidative Stress Modulates Mitochondrial Failure and Cyclophilin D Function in X-Linked Adrenoleukodystrophy. Brain J. Neurol. 2012, 135, 3584–3598. [Google Scholar] [CrossRef] [Green Version]
- Walton, P.A.; Pizzitelli, M. Effects of Peroxisomal Catalase Inhibition on Mitochondrial Function. Front. Physiol. 2012, 3, 108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Viswakarma, N.; Yu, S.; Jia, Y.; Bai, L.; Vluggens, A.; Cherkaoui-Malki, M.; Khan, M.; Singh, I.; Yang, G.; et al. Progressive Endoplasmic Reticulum Stress Contributes to Hepatocarcinogenesis in Fatty Acyl-CoA Oxidase 1-Deficient Mice. Am. J. Pathol. 2011, 179, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Fessel, J.P.; Sherrill, T.; Kocurek, E.G.; Yull, F.E.; Blackwell, T.S. Enhanced Expression of Catalase in Mitochondria Modulates NF-ΚB-Dependent Lung Inflammation through Alteration of Metabolic Activity in Macrophages. J. Immunol. Baltim. Md 1950 2020, 205, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Tsan, M.F.; Clark, R.N.; Goyert, S.M.; White, J.E. Induction of TNF-Alpha and MnSOD by Endotoxin: Role of Membrane CD14 and Toll-like Receptor-4. Am. J. Physiol. Cell Physiol. 2001, 280, C1422–C1430. [Google Scholar] [CrossRef] [Green Version]
- Krautbauer, S.; Eisinger, K.; Neumeier, M.; Hader, Y.; Buettner, R.; Schmid, P.M.; Aslanidis, C.; Buechler, C. Free Fatty Acids, Lipopolysaccharide and IL-1α Induce Adipocyte Manganese Superoxide Dismutase Which Is Increased in Visceral Adipose Tissues of Obese Rodents. PLoS ONE 2014, 9, e86866. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, E.; Bernard, A.-S.; Ching, H.Y.V.; Somogyi, A.; Medjoubi, K.; Fores, J.R.; Bertrand, H.C.; Vincent, A.; Trépout, S.; Guerquin-Kern, J.-L.; et al. Anti-Inflammatory Activity of Superoxide Dismutase Mimics Functionalized with Cell-Penetrating Peptides. Dalton Trans. Camb. Engl. 2003 2020, 49, 2323–2330. [Google Scholar] [CrossRef] [Green Version]
- Schrader, M.; Krieglstein, K.; Fahimi, H.D. Tubular Peroxisomes in HepG2 Cells: Selective Induction by Growth Factors and Arachidonic Acid. Eur. J. Cell Biol. 1998, 75, 87–96. [Google Scholar] [CrossRef]
- Girnun, G.D.; Domann, F.E.; Moore, S.A.; Robbins, M.E.C. Identification of a Functional Peroxisome Proliferator-Activated Receptor Response Element in the Rat Catalase Promoter. Mol. Endocrinol. 2002, 16, 2793–2801. [Google Scholar] [CrossRef] [Green Version]
- Mattace Raso, G.; Simeoli, R.; Russo, R.; Santoro, A.; Pirozzi, C.; d’Emmanuele di Villa Bianca, R.; Mitidieri, E.; Paciello, O.; Pagano, T.B.; Orefice, N.S.; et al. N-Palmitoylethanolamide Protects the Kidney from Hypertensive Injury in Spontaneously Hypertensive Rats via Inhibition of Oxidative Stress. Pharmacol. Res. 2013, 76, 67–76. [Google Scholar] [CrossRef]
- Balkan, J.; Parldar, F.H.; Doğru-Abbasoğlu, S.; Aykaç-Toker, G.; Uysal, M. The Effect of Taurine or Betaine Pretreatment on Hepatotoxicity and Prooxidant Status Induced by Lipopolysaccharide Treatment in the Liver of Rats. Eur. J. Gastroenterol. Hepatol. 2005, 17, 917–921. [Google Scholar] [CrossRef]
- Hsu, D.-Z.; Chiang, P.-J.; Chien, S.-P.; Huang, B.-M.; Liu, M.-Y. Parenteral Sesame Oil Attenuates Oxidative Stress after Endotoxin Intoxication in Rats. Toxicology 2004, 196, 147–153. [Google Scholar] [CrossRef]
- Zhu, J.-H.; Lei, X.G. Lipopolysaccharide-Induced Hepatic Oxidative Injury Is Not Potentiated by Knockout of GPX1 and SOD1 in Mice. Biochem. Biophys. Res. Commun. 2011, 404, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.; Jacobson, A. The Intimate Relationships of MRNA Decay and Translation. Trends Genet. TIG 2013, 29, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Guan, B.-J.; Krokowski, D.; Majumder, M.; Schmotzer, C.L.; Kimball, S.R.; Merrick, W.C.; Koromilas, A.E.; Hatzoglou, M. Translational Control during Endoplasmic Reticulum Stress beyond Phosphorylation of the Translation Initiation Factor EIF2α. J. Biol. Chem. 2014, 289, 12593–12611. [Google Scholar] [CrossRef] [Green Version]
- Rabie, K.; Mourad, K.; Imig, J.D.; Meryem, E.; Zouhra, D.; Nacieri Mrabti, H.; Elmsellem, H.; Abdelaziz, B.; Yahia, C.; Katim, A. Antioxidant Activities, Total Polyphenolic Compounds And Hplc/Dad/Ms Phenolic Profile of Argan Oil Derived from Two Differents Methods of Extractions. J. Chem. Pharm. Res. 2017, 8, 1320–1327. [Google Scholar]
- Seiquer, I.; Rueda, A.; Olalla, M.; Cabrera-Vique, C. Assessing the Bioavailability of Polyphenols and Antioxidant Properties of Extra Virgin Argan Oil by Simulated Digestion and Caco-2 Cell Assays. Comparative Study with Extra Virgin Olive Oil. Food Chem. 2015, 188, 496–503. [Google Scholar] [CrossRef]
- Martínez-Huélamo, M.; Rodríguez-Morató, J.; Boronat, A.; De la Torre, R. Modulation of Nrf2 by Olive Oil and Wine Polyphenols and Neuroprotection. Antioxidants 2017, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Cherkaoui-Malki, M.; Meyer, K.; Cao, W.Q.; Latruffe, N.; Yeldandi, A.V.; Rao, M.S.; Bradfield, C.A.; Reddy, J.K. Identification of Novel Peroxisome Proliferator-Activated Receptor Alpha (PPARalpha) Target Genes in Mouse Liver Using CDNA Microarray Analysis. Gene Expr. 2001, 9, 291–304. [Google Scholar] [CrossRef]
- Feingold, K.R.; Wang, Y.; Moser, A.; Shigenaga, J.K.; Grunfeld, C. LPS Decreases Fatty Acid Oxidation and Nuclear Hormone Receptors in the Kidney. J. Lipid Res. 2008, 49, 2179–2187. [Google Scholar] [CrossRef] [Green Version]
- Cherki, M.; Derouiche, A.; Drissi, A.; El Messal, M.; Bamou, Y.; Idrissi-ouadghiri, A.; Khalil, A.; Adlouni, A.; Msik, B. Consumption of Argan Oil May Have an Antiatherogenic Effect by Improving Paraoxonase Activities and Antioxidant Status: Intervention Study in Healthy Men. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 352–360. [Google Scholar] [CrossRef]
- Wiesinger, C.; Kunze, M.; Regelsberger, G.; Forss-Petter, S.; Berger, J. Impaired Very Long-Chain Acyl-CoA β-Oxidation in Human X-Linked Adrenoleukodystrophy Fibroblasts Is a Direct Consequence of ABCD1 Transporter Dysfunction. J. Biol. Chem. 2013, 288, 19269–19279. [Google Scholar] [CrossRef] [Green Version]
- Baarine, M.; Andréoletti, P.; Athias, A.; Nury, T.; Zarrouk, A.; Ragot, K.; Vejux, A.; Riedinger, J.M.; Kattan, Z.; Bessede, G.; et al. Evidence of Oxidative Stress in Very Long Chain Fatty Acid—Treated Oligodendrocytes and Potentialization of ROS Production Using RNA Interference-Directed Knockdown of ABCD1 and ACOX1 Peroxisomal Proteins. Neuroscience 2012, 213, 1–18. [Google Scholar] [CrossRef]
- Kemp, S.; Berger, J.; Aubourg, P. X-Linked Adrenoleukodystrophy: Clinical, Metabolic, Genetic and Pathophysiological Aspects. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 1465–1474. [Google Scholar] [CrossRef] [Green Version]
- El Kharrassi, Y.; Maata, N.; Mazri, M.A.; El Kamouni, S.; Talbi, M.; El Kebbaj, R.; Moustaid, K.; Essamadi, A.K.; Andreoletti, P.; El Mzouri, E.H.; et al. Chemical and Phytochemical Characterizations of Argan Oil (Argania spinosa L. Skeels), Olive Oil (Olea europaea L. Cv. Moroccan Picholine), Cactus Pear (Opuntia megacantha Salm-Dyck) Seed Oil and Cactus Cladode Essential Oil. J. Food Meas. Charact. 2018, 12, 747–754. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Ni, J.; Sasaki, Y.; Tokuyama, S.; Sogabe, A.; Tahara, Y. Conversion of a Typical Catalase from Bacillus Sp. TE124 to a Catalase-Peroxidase by Directed Evolution. J. Biosci. Bioeng. 2002, 93, 31–36. [Google Scholar] [CrossRef]
- Beyer, W.F.J.; Fridovich, I. Assaying for Superoxide Dismutase Activity: Some Large Consequences of Minor Changes in Conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assays of Glutathione Peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar] [CrossRef]
- ELLMAN, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 358, 351–358. [Google Scholar] [CrossRef]
- Raas, Q.; Gondcaille, C.; Hamon, Y.; Leoni, V.; Caccia, C.; Menetrier, F.; Lizard, G.; Trompier, D.; Savary, S. CRISPR/Cas9-Mediated Knockout of Abcd1 and Abcd2 Genes in BV-2 Cells: Novel Microglial Models for X-Linked Adrenoleukodystrophy. Biochim Biophys Acta Mol. Cell Biol. Lipids 2019, 1864, 704–714. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids (g/100 g Oil) | Argan Oil | Olive Oil | Colza Oil |
|---|---|---|---|
| Myristic acid C14:0 | 0.1 | - | - |
| Palmitic acid C16:0 | 13.4 | 9.09 | 4.5 |
| Palmitoleic acid C16:1 n-7 | 0.1 | 0.73 | 0.3 |
| Stearic acid C18:0 | 6.5 | 2.26 | 1.6 |
| Oleic acid C18:1n-9 | 46.4 | 76.35 | 63 |
| Linoleic acid C18:2n-6 | 32.2 | 9.95 | 19 |
| Linolenic acid C18:3 n-3 | 0.1 | 0.86 | 9 |
| Arachidic acid C20:0 | 0.4 | 0.31 | 0.5 |
| Eicosenoic acid C20:1 | 0.4 | 0.35 | 1.3 |
| Phytosterol (mg/100 g oil) | |||
| Cholesterol | - | 0.06 | 0.3 |
| Brassicasterol | - | - | 10.1 |
| Campesterol | 0.2 | 3.21 | 34.4 |
| Campestanol | - | 0.04 | - |
| Stigmasterol | - | 1.59 | 0.2 |
| b-Sitosterol | - | 84.77 | 44.5 |
| Δ-5-Avenasterol | - | 5.23 | 3.1 |
| Δ- 7-Stigmasterol | - | 0.24 | 0.1 |
| Δ-7-Avenasterol | 4.2 | 0.336 | 0.1 |
| Spinasterol | 35.3 | - | - |
| Schottenol | 43.8 | - | - |
| Tocopherol (mg/Kg oil) | |||
| α-Tocopherol | 8.3 | 87.92 | 24.03 |
| γ-Tocopherol | 88.8 | 4.62 | 69.01 |
| δ-Tocopherol | 1.2 | 7.46 | 6.96 |
| Gene Name | Primer Sequences |
|---|---|
| Il10-F Il10-R | 5′ GCTGGACAACATACTGCTAACC 3′ |
| 5′ CCCAAGTAACCCTTAAAGTCCTG 3′ | |
| Acox1-F Acox1-R | 5′ TCGAAGCCAGCGTTACGAG3′ 5′ GGTCTGCGATGCCAAATTCC3′ |
| Tnf α-F Tnf α-R | 5′ GACGTGGAAGTGGCAGAAGAG3′ 5′ TGCCACAAGCAGGAATGAGA3′ |
| Catalase-F Catalase-R | 5′ AGCGACCAGATGAAGCAGTG3′ 5′TCCGCTCTCTGTCAAAGTGTG3′ |
| Sod1-F Sod1-R | 5′AACCAGTTGTGTTGTCAGGAC3′ 5′CCACCATGTTTCTTAGAGTGAGG3′ |
| 36b4-F 36b4-R | 5′CGACCTGGAAGTCCAACTAC3′ 5′ATCTGCTGCATCTGCTTG3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Essadek, S.; Bouchab, H.; El Kebbaj, R.; Gondcaille, C.; El Kamouni, S.; Savary, S.; Vamecq, J.; Essamadi, A.; Cherkaoui-Malki, M.; Nasser, B.; et al. Effects of a Short-Term Lipopolysaccharides Challenge on Mouse Brain and Liver Peroxisomal Antioxidant and β-oxidative Functions: Protective Action of Argan Oil. Pharmaceuticals 2022, 15, 465. https://doi.org/10.3390/ph15040465
Essadek S, Bouchab H, El Kebbaj R, Gondcaille C, El Kamouni S, Savary S, Vamecq J, Essamadi A, Cherkaoui-Malki M, Nasser B, et al. Effects of a Short-Term Lipopolysaccharides Challenge on Mouse Brain and Liver Peroxisomal Antioxidant and β-oxidative Functions: Protective Action of Argan Oil. Pharmaceuticals. 2022; 15(4):465. https://doi.org/10.3390/ph15040465
Chicago/Turabian StyleEssadek, Soukaina, Habiba Bouchab, Riad El Kebbaj, Catherine Gondcaille, Soufiane El Kamouni, Stéphane Savary, Joseph Vamecq, Abdelkhalid Essamadi, Mustapha Cherkaoui-Malki, Boubker Nasser, and et al. 2022. "Effects of a Short-Term Lipopolysaccharides Challenge on Mouse Brain and Liver Peroxisomal Antioxidant and β-oxidative Functions: Protective Action of Argan Oil" Pharmaceuticals 15, no. 4: 465. https://doi.org/10.3390/ph15040465
APA StyleEssadek, S., Bouchab, H., El Kebbaj, R., Gondcaille, C., El Kamouni, S., Savary, S., Vamecq, J., Essamadi, A., Cherkaoui-Malki, M., Nasser, B., & Andreoletti, P. (2022). Effects of a Short-Term Lipopolysaccharides Challenge on Mouse Brain and Liver Peroxisomal Antioxidant and β-oxidative Functions: Protective Action of Argan Oil. Pharmaceuticals, 15(4), 465. https://doi.org/10.3390/ph15040465