Abstract
Dopamine neurotransmission plays critical roles in regulating complex cognitive and behavioral processes including reward, motivation, reinforcement learning, and movement. Dopamine receptors are classified into five subtypes, widely distributed across the brain, including regions responsible for motor functions and specific areas related to cognitive and emotional functions. Dopamine also acts on astrocytes, which express dopamine receptors as well. The discovery of direct receptor–receptor interactions, leading to the formation of multimeric receptor complexes at the cell membrane and providing the cell decoding apparatus with flexible dynamics in terms of recognition and signal transduction, has expanded the knowledge of the G-protein-coupled receptor-mediated signaling processes. The purpose of this review article is to provide an overview of currently identified receptor complexes containing dopamine receptors and of their modulatory action on dopamine-mediated signaling between neurons and between neurons and astrocytes. Pharmacological possibilities offered by targeting receptor complexes in terms of addressing neuropsychiatric disorders associated with altered dopamine signaling will also be briefly discussed.
1. Introduction
Dopamine (DA) is a catecholamine, that is, an ethylamine with an attached catechol group (a phenyl group with two hydroxyl groups in meta- and para positions). DA-producing neurons were first identified and mapped in animals by Dahlström and Fuxe in 1964 [1,2], indicating the existence of neuronal circuits using DA as a neurotransmitter. In the years that followed, the characterization of the circuits which utilize DA, their organization, molecular signature, and cellular and functional features represented one of the most fertile fields of research in neuroscience (see [3]). Over the past decades, technological advances have also helped to expand the knowledge about the anatomical organization of the DA systems in the human brain. Dopamine neuronal populations have, indeed, been identified and characterized in the human brain from the level of gene transcription to the level of the distribution of related proteins by using post-mortem immunohistochemistry and in vitro autoradiography methods, as well as through in vivo neuroimaging techniques such as positron emission tomography and single photon emission tomography (see [4]). As summarized in Table 1, four major dopamine pathways and two additional ones can be described in the human brain [5,6]. They are involved in the regulation of both physiological and behavioral processes, including movement, endocrine control, cognition, reward, and motivation.

Table 1.
Dopaminergic pathways [3,6].
The release of the catecholamine from nerve endings upon axonal stimulation certainly represents the main process of dopamine-mediated interneural communication. Released DA acts on postsynaptic and presynaptic receptors at the synapse and is mostly taken up back into nerve endings by the dopamine transporter protein, which belongs to the solute carrier transporter family [6]. In some regions of the central nervous system, however, dopamine signaling also occurs through processes of “volume transmission”, based on the diffusion of the molecule in the extra-cellular space to reach more distant targets (see [7,8,9]). Examples include globus pallidus [10], substantia nigra [11], ventral tegmental area [12], ventral subiculum [13], pedunculopontine nucleus [14], and retina [15]. In this respect, of significant interest is evidence indicating that DA, interacting with DA receptors expressed by astrocytes [16,17,18], may also act on these cells, leading to a modulation of neuron–astrocyte crosstalk (see [19]).
Dopamine receptors belong to the superfamily of G-protein-coupled receptors (GPCRs). A first indication of their existence was reported in 1972 [20]; they were identified in 1975 [21,22], and five different subtypes have been described so far. In view of the strong implication of DA signaling in a variety of neurological, psychiatric, and drug addiction disorders, with a relevant impact not only on afflicted individuals, but also on society, DA receptors have been the focus of intense research efforts and a variety of drugs have been designed to treat these illnesses by targeting DA receptors directly or indirectly (see [6]).
In recent decades, experimental evidence demonstrating that structural receptor–receptor interactions (RRIs) may occur between receptor proteins has been of interest [23,24,25,26,27,28,29,30,31,32,33]. The term RRI indicates a type of interaction needing a direct physical contact between the partner proteins, with the formation of oligomeric complexes at the cell membrane (see [34] for a recent review). Available studies indicated the formation of receptor complexes as a quite common process in the different receptor families, where the ion channel receptors are at one end of the spectrum (being assembled by multimerization) and GPCRs at the other. Thus, as pointed out by Changeux and Christopoulos in a detailed review [35], RRIs emerge as an efficient mechanism for modulating the functional properties of receptor proteins, including GPCRs that are able to signal as monomers. This mechanism, indeed, allows a sophisticated regulation of the intercellular communication already at the membrane level [9] and opens the possibility of new pharmacological strategies to modulate receptor signaling. In this context, several groups (including our group), have focused their attention on the detection of receptor complexes containing DA receptors in nervous tissues and on the role they can play in DA-mediated signaling in neurons and astrocytes. In the present review, published data concerning this modulatory process will be presented and discussed. Since the subject is quite broad, review articles focused on specific aspects of the topic will also be suggested for further information.
2. Dopamine Receptors
Five different subtypes of DA receptors (D1, D2, D3, D4, and D5) have been identified in brain tissue (see [6] for a recent review), and based on their structure and pharmacological properties, they can be classified into two major groups [36]: D1-like receptors (including D1 and D5) and D2-like receptors (comprising D2, D3, and D4). Binding studies have demonstrated some differences between the two groups in terms of affinity to DA, with D2-like receptors exhibiting a 10- to 100-fold greater affinity to DA than D1-like receptors [37,38,39]. D1- and D2-like receptors also differ in their genetic structure. D2-like receptor genes, indeed, have introns in their coding regions, while D1-like receptor genes do not exhibit this feature [40]. This genetic organization, therefore, enables the generation of D2-like receptor splice variants, and alternative splicing is particularly important for the D2 receptor, leading to the generation of two distinct receptor isoforms: D2-short and D2-long [41,42], differing because of the insertion of 29 amino acids in the D2-long intracellular domain, which may play a role in determining second messenger specificity [36,42].
Concerning signal transduction, it is commonly accepted that the receptors of the D1-like group mainly mediate the stimulation of the second messenger adenylyl cyclase (cAMP) by coupling to the Gs protein, whereas receptors of the D2-like group mainly exert inhibitory effects on this enzyme by coupling to Gi/0 protein [6,43]. In addition to the just mentioned main pathway, D1-like receptors may also couple to the Gq protein [44,45,46] and modulate phospholipase C [44,46,47], leading to an increase in intracellular calcium levels and activation of protein kinase C. In this respect, the regulation of intracellular calcium levels is a well-documented action of dopamine on astrocytes [48]. DA receptors are expressed by astrocytes [49], and D2 receptor activation was reported to decrease intracellular Ca2+ levels in hippocampal [50] and ventral midbrain astrocytes [51], while D1 receptor activation elevated astrocytic Ca2+ levels in the hippocampus [50], nucleus accumbens [52], and cerebellum [53].
DA signaling cascade, however, may also be modulated by the significant network of molecular interactions that DA receptors can establish in their environment [6,43], which interfere with the GPCRs activity. A first example [54,55] is provided by G-protein-coupled receptor kinases (GRKs). GRKs phosphorylate receptors in response to persistent stimulation [56]. Consequently, the receptor becomes a target for a scaffolding protein, named arrestin, blocking further activation of the GPCR [57] and allowing the GPCR–arrestin complex to engage a variety of G-protein-independent signaling pathways [58]. A second example [59,60] is represented by the regulators of G protein signaling (RGS). RGS are a family of more than 35 intracellular proteins (see [6]) that induce inhibitory effects on GPCRs. Concerning DA receptors, they mainly regulate the D2-like class [61] and are important in order to stop signaling in the slow synaptic transmission elicited by D2 receptors [38,62].
In this context, of particular interest is the possibility of direct RRI involving DA receptors with the formation of receptor complexes at the cell membrane [63,64]. In receptor complexes, indeed, the chain of events linking the recognition of a ligand by the single protomers to the signal transduction also depends on the neighboring receptors. This specific mechanism modulating DA signaling will be the focus of the next sections.
3. Structural Receptor–Receptor Interactions
Functional interactions between receptors, by mechanisms of transactivation or by sharing signaling pathways, are well-known processes that do not need a physical contact between the involved proteins [65]. In the 1980s, however, Agnati, Fuxe, and collaborators [23,66], through in vitro and in vivo experiments, provided indirect evidence that GPCR monomers can establish structural interactions (see [63] for historical details). These findings led to the hypothesis that neuron activity could be modulated by receptor complexes present at the cell membrane and formed by different types of GPCRs [64], a mechanism allowing (already at the membrane level) some integration of synaptic (wiring transmission) and extra-synaptic (volume transmission) signals [64]. The term RRI was subsequently proposed to emphasize the concept of an interaction between receptors requiring a direct physical contact between the molecules and leading to the formation of dimers or high-order molecular complexes at the cell membrane [67]. In the years that followed, several groups [23,24,25,26,27,28,29,30,31,32,33] provided direct evidence of the existence of this structural organization, and the amount of data supporting the existence of GPCR complexes further increased with the advent of biophysical techniques capable of detecting the spatial proximity of protein molecules [68,69,70,71]. The obtained results demonstrated that GPCRs can signal not only as monomers, but also as part of receptor complexes [72] and indicated that receptor complexes represent a quite common molecular organization in the different families of receptors [34].
The basic molecular mechanism underlying the formation and the dynamics of these receptor assemblies are allosteric interactions (see [73]). Allostery (see [35,74,75,76] for extensive reviews) is a mode of communication between distant sites in a protein, in which the energy associated with dynamic or conformational changes at one site can be transferred (along specific pathways within the protein structure) to other sites, that, in turn, will change their conformational or dynamic features. Thus, when a quaternary structure is established via direct RRIs between protomers, energy perturbations at some site of one protomer can propagate into the nearby protomers and change their conformational and functional properties, leading to a cooperative behavior of the whole complex [34,77]. In current research on receptor oligomerization, therefore, the identification of the residues forming the interface between protomers is of significant interest. They, indeed, influence the overall architecture that the receptor complex can assume. In this respect, to predict the interfaces available for RRIs, several bioinformatics methods have been developed (see [78,79,80] for reviews on this topic). As a matter of fact, the number of ways GPCRs interact in the membrane to form complexes is probably limited. The vast majority of experimentally identified receptor complexes, indeed, are dimers. And some interfaces have been observed to be more exploited than others for RRIs [81]. Nevertheless, oligomeric heteroreceptors have been detected (see [81,82,83,84]).
The signaling outcomes from a receptor complex, therefore, depend on several factors (including the composition and the topological organization of the complex and the effects of ligands on its stability and trafficking), which may strongly influence the cascade of events linking the recognition of a ligand by single protomers to the signal transduction (see [43,80,85]). Some of the possible modulations that allosteric RRIs may induce on signaling when a receptor complex forms are summarized in Figure 1. They include changes in ligand recognition, G-protein activation, receptor desensitization [86], and switching to β-arrestin signaling [87]. In this context, a relevant aspect of receptor complex formation is also the possible appearance in the formed quaternary structure of novel specific allosteric sites allowing the binding of some modulator. Thus, ligands specific to the receptor complex as such may also exist (see [88]).
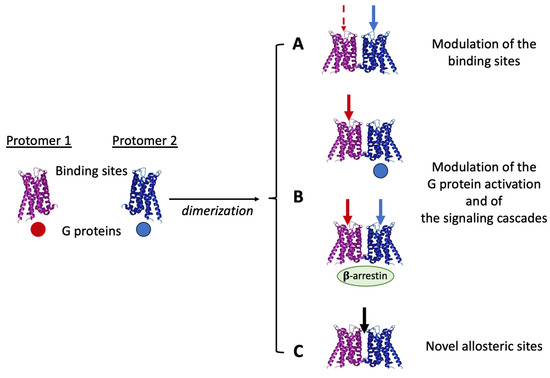
Figure 1.
As a result of allosteric RRIs, receptor complexes appear to be endowed with pharmacological features that cannot be fully derived from the characteristics of the single participating protomers (see text).
A final aspect deserving consideration (see [34] for a discussion) concerns the cell environment in which receptor complexes are located. In fact, the network of molecular interactions they can establish at the cell membrane with other biochemical components (the so-called “horizontal molecular networks” [89]) may influence their signaling. In this context, a specific aspect of interest is the lipid environment, since it was shown to influence receptor function [90]. In particular, changes in the membrane composition altering receptor signaling were associated with several health disorders during aging [90].
4. Receptor Complexes Involving Dopamine Receptors
DA receptors belong to class A GPCRs [91], well known for being able to signal as monomers [92]. In addition, however, the overall available evidence (obtained through multiple approaches with consistent results) strongly supports the presence of class A GPCR complexes in native systems [72]. In this respect, studies concerning the kinetics of complex formation and its dependence on the involved interaction energy [93] are of substantial interest. The observed half-lives of dimers indicate that they are often transient (lasting few hours) and may undergo recombination (“kiss-and-run” encounters [80]). These processes may lead to a dynamic equilibrium between monomers and receptor complexes for class A GPCRs, as suggested by studies on the corticotropin-releasing factor receptor type 1 in the endoplasmic reticulum [94], indicating that the ratio of monomers/receptor complexes was maintained at an almost constant level in the plasma membrane, even in spite of agonist activation of the receptors. Receptor complexes including DA receptors (see also [95]) are shown in Table 2.
Table 2.
Receptor complexes involving dopamine receptors.
4.1. Receptor Complexes Involving D2-like Dopamine Receptors
A first aspect emerging from the available data is that D2 appears to be a hub receptor which interacts with many other GPCRs.
Probably, the most studied interaction is between dopamine D2 and adenosine A2A receptors, leading to the formation of A2A-D2 heterodimers (see [95,120] for reviews). By using pull-down and mass spectrometry techniques, it has been demonstrated [121,122] that the heteromerization between A2A and D2 receptors significantly depends on charged residues located at the intracellular part of the transmembrane helix 5 (TM5) of the D2 receptor. The role of TM helix interactions within the A2A-D2 heteroreceptor complex interface has also been explored by using synthetic TM α-helix peptides of the D2 receptor [123], and the results allowed for the identification of a TM4/5 interface between the two monomers. The A2A-D2 heterodimer is also representative of many aspects concerning the signaling outcome from a receptor complex. Experimental evidence has shown that the receptor complex formation modifies the signaling from the single protomers. In particular, early in vitro experiments on membrane preparations showed a reduction in the affinity of the high-affinity D2-agonist-binding site after incubation with the A2A agonist CGS21680 [124,125], demonstrating that antagonistic interactions occur in the A2A-D2 heterodimer. By using receptor autoradiography, this finding was subsequently confirmed by studies on brain tissue from rats and humans [126]. They showed a strong reduction in D2 receptor affinity for dopamine in the nucleus accumbens core and shell after the A2A receptor agonist treatment. By using functional, biochemical, and biophysical techniques (such as co-immunoprecipitation and proximity ligation assay), antagonistic interactions between A2A and D2 receptors were also recently demonstrated in astrocytes [96,127]. In this context, observations indicating that agonist activation of the A2A protomer in the A2A-D2 heteroreceptor complex inhibits D2 Gi/o-mediated signaling but increases the D2 β-arrestin2-mediated signaling are of interest. This marks a difference compared with the action of D2 receptor antagonists, which block all the D2 signaling pathways. Thus, through the allosteric receptor–receptor interaction, an A2A agonist becomes a biased inhibitory modulator of the Gi/o-mediated D2 signaling [128]. The possible formation, as a consequence of the formation of a receptor complex, of new allosteric sites allowing the binding of some ligand is a further modulatory mechanism that the A2A-D2 heteromer illustrates. Homocysteine can, indeed, bind to the heterodimer without interfering with the RRI between A2A and D2 and acts as an allosteric antagonist of the D2 receptor [129]. Thus, the inhibitory effect of A2A agonists is amplified by homocysteine. These modulatory actions were demonstrated in striatal neurons [129], as well as in astrocytes [130], where homocysteine reduces the D2-mediated inhibition of glutamate release. An intriguing process involving A2A and D2 receptors was highlighted by studies on cell lines [19] that demonstrated intercellular transfer of these GPCRs by exosomes, resulting in the incorporation of functional receptors into acceptor cells. As shown by photo-bleaching fluorescence resonance energy transfer, the transferred receptors may also undergo A2A-D2 receptor heteromerization in the target cell. Thus, the release of extracellular micro vesicles (the so-called “roamer type” of volume transmission [19]) may represent a significant mechanism for the modulation of neuron-neuron and astrocyte–neuron intercellular signaling.
Evidence has been provided indicating that the adenosine A2A receptor can establish antagonistic RRIs with the other D2-like receptors as well, namely, D3 [97] and D4 [98], leading to a reduction in the affinity of their binding site for DA. Antagonistic RRIs also characterize other receptor complexes involving the D2 receptor, as, for instance, the heterodimers it can form with the glutamate NMDA [99] and mGluR5 [84] receptors, the neurotensin NTS1 [100] receptor, and the cannabinoid CB1 [102,131] receptor. Higher-order heteroreceptor complexes, involving both A2A and D2, have also been identified. Examples include the heterotrimers formed by A2A and D2 receptors with the metabotropic glutamate receptor 5 (A2A-D2-mGluR5 [84]), the sigma1 receptor (A2A-D2-sigma1 [113]), and the cannabinoid CB1 receptor (A2A-D2-CB1 [82]). In these receptor complexes, the pattern of allosteric interactions on the D2 protomer also inhibits the recognition and signaling of the DA receptor.
Synergistic RRIs involving the D2 receptor, however, were also identified. A first example is provided by the receptor complex between the D2 receptor and the serotonin 5-HT2A receptor [104], where the activation of the 5-HT2A protomer by 5-HT2A agonists produced an enhancement of D2 signaling. In astrocytes, receptor complexes between the dopamine D2 receptor and the serotonin 5-HT1A receptor have been observed [103]. However, the functional consequences of the signaling pathways mediated by D2-5-HT1 heteromers in these cells are still not known in detail [132]. A further example is represented by the D2-OTR heterodimer, involving D2 and the oxytocin receptor. In neurons [105], oxytocin, via the allosteric RRI established in the heterocomplex, markedly increased D2 receptor recognition (increased affinity of the high-affinity state) and increased the coupling of Gi/o to the receptor. The D2-OTR heterodimer was recently identified in astrocytes as well [106], and the activation of OTR was shown to have a facilitatory effect on the response of D2 receptors, causing them to be activated by subthreshold D2 agonist concentrations and leading to an inhibition of glutamate release by the cells.
Synergistic RRIs are also in operation in the heterodimer involving the dopamine D4 receptor and μ-opioid receptor (MOR) [110], since D4 activation causes a substantial increase in the affinity of the MOR agonist binding sites. Evidence was also obtained that the D4 and β2-adrenergic receptor may form a D4-β2 receptor complex that integrates Gs- and Gi-mediated regulation of adenylyl cyclase [109]. In this context, of particular interest are also studies (see [108]) focused on the dopamine D4 receptor polymorphic variants D4.4 (four repeats in exon 3) and D4.7 (seven repeats in exon 3), both able to heterodimerize with the norepinephrine α2A receptor. However, only heteromerization with D4.7, but not with D4.4, increases the potency of norepinephrine in terms of activating the α2A receptor, indicating the possible polymorphic variants of a D2-like receptor as a factor conferring significantly different pharmacological properties onto the receptor complexes it may form.
4.2. Receptor Complexes Involving D1-like Dopamine Receptors
The potentiation of immediate early gene expression and of arachidonic acid release have been described as functional interactions between activated dopamine D1 and D2 receptors (see [45]). However, it was also demonstrated that stably co-expressed D1 and D2 receptors may form heteromeric units [114]. It is of substantial interest that the two receptors, when coactivated in the same cell, produce a phospholipase C-mediated calcium signal that is not seen when the receptors are activated alone. The pharmacological analysis of this receptor complex indicated a specific coupling to the Gq/11 pathway to produce such a response. Activation of Gq/11, however, could not be elicited through activation of either receptor when activated alone. Thus, the recruitment of G proteins other than those expected for the monomers has been observed after D1-D2 dimerization, a further mechanism of signal transduction modulation associated with receptor complex formation.
Antagonistic interactions between D1 and the adenosine A1 receptor, associated with the formation of A1-D1 heterodimers [116,117], were also characterized. A1 agonists, indeed, were found to reduce the number of D1 agonist binding sites in the high-affinity state, and with receptor autoradiography, A1 agonists were found to antagonistically modulate D1 binding sites, causing a reduction in their affinity (see [133] for details).
Receptor complexes between dopamine D1 and D3 have been demonstrated using several techniques, giving evidence for synergistic intramembrane D1-D3 interactions at the level of D1 recognition, since D3 activation was able to increase the affinity of the D1 agonist binding sites [115]. Synergistic RRIs also exist in the D1-NMDA heterodimer [118], by which NMDA receptor activation can recruit D1 receptors to the plasma membrane, thereby leading to an increase in D1 signaling and cAMP accumulation.
Recent interesting findings on prefrontal cortex astrocytes indicated a significant functional interaction between α1-adrenergic and DA receptors, driving downstream Ca2+ signaling [112]. Also, in light of the abovementioned data showing that DA receptors may form receptor complexes with adrenergic receptors [109,134], and of neuroanatomical data showing that D1 and α1-adrenergic receptors colocalize on prefrontal cortex dendrites and may undergo co-trafficking [119], the hypothesis has been put forward that in cortical astrocytes as well, heterodimers involving DA receptors and adrenergic receptors could be present [112]. A direct experimental demonstration, however, is still lacking.
GABAA and dopamine D5 heteromerization, demonstrated by Liu and collaborators [135], was the first identification of a receptor complex involving a GPCR and an ion-channel receptor. The results indicated that co-activation of the monomers was required for the formation of the complex, which allowed for a bidirectional crosstalk, leading to a reduction in GABAA signaling and a reduced coupling between D5 and Gs proteins.
4.3. Possible Differences in Receptor Complex Dynamics in Neurons and Astrocytes
As briefly illustrated before, a number of receptor complexes (such as, for instance, the A2A-D2 heterodimer) are expressed both in neurons and astrocytes. In this respect, it is reasonable to assume that the conformation of a receptor complex in the two cases may exhibit some difference because of differences in the membrane microenvironment. Differences in the energy landscape, indeed, modulate the pattern of allosteric interaction between monomers and may lead to changes in the signaling features of the complex that they can form [80].
Differences in membrane potential between the two cell types, for instance, have been documented [136]. Unlike neurons, astrocytes do not generate action potentials, but they are electrically dynamic cells. Indeed, in contrast to most non-excitable cells that have relatively depolarized membrane potentials, astrocytes have a hyperpolarized membrane (at a level that typically rests significantly below that of neurons) and a low membrane resistance. For the present discussion, membrane composition is another factor deserving consideration. This aspect was the focus of an extensive lipidome analysis by Fitzner and collaborators [137], showing that each cell type was characterized by a unique lipid composition: neurons, for instance, exhibited quite high levels of cholesterol, while astrocytes were enriched in phosphatidylinositol.
All these features of the membrane microenvironment, therefore, have the potential to modulate the pharmacological properties of a given receptor complex. To illustrate this concept, the results of a simulation based on molecular modeling methods and focused on the A2A-D2 heterodimer in two different membrane environments (neuron-like and astrocyte-like) are shown in Figure 2.
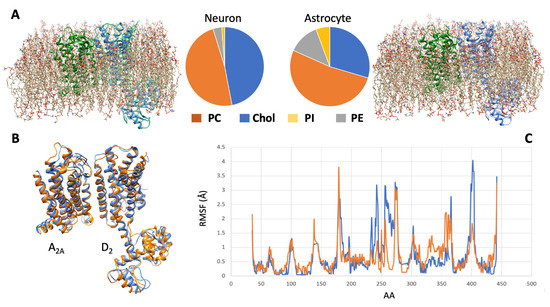
Figure 2.
Molecular dynamics simulation of the A2A-D2 receptor complex in different cell membranes. (A) By using the CHARMM-GUI membrane builder web server (http://www.charmm-gui.org/?doc=input [138], accessed on 5 July 2023), four phospholipids, namely, phosphatidylcholine (PC), cholesterol (Chol), phosphatidylinositol (PI), and phosphatidylethanolamine (PE), were used to model two different membrane bilayers around the molecular model [120,123] of the heterodimer. The first (left panel) approximated the neuronal membrane composition, the second one (right panel) the astrocytic one (see [137]). A molecular dynamics procedure, based on the CABSflex method [139] and available as a web server (https://biocomp.chem.uw.edu.pl/CABSflex2, accessed on 6 July 2023), was then used to evaluate the conformations that the receptor complex may acquire in the two environments. (B) Configurations of minimal energy of the A2A-D2 heterodimer in neuronal (orange) and astrocytic (blue) membrane. (C) Root mean square fluctuations (RMSF) diagrams, per amino acid position, of the D2 monomer chain when in neuronal (orange) and astrocytic (blue) membrane. The estimated differences in configuration and dynamical behavior of the heterodimer suggest that different membrane environments could represent a factor modulating the pharmacological properties of the receptor complex.
5. Complexes Involving Dopamine Receptors in the Main Dopaminergic Pathways: Impact on Neuropharmacology
The intermingling of findings from functional neuroanatomy (linking dopaminergic pathways to specific functions and diseases) with evidence emerging from chemical neuroanatomy (describing the distribution of receptor complexes involving DA receptors in brain cells and regions) may help to better appreciate the function that receptor complexes containing DA receptors can fulfill, and may contribute to the development of new pharmacological approaches with a potentially major impact on molecular medicine. In this respect, presently available information is limited to ascending dopaminergic pathways (nigro-striatal, mesolimbic, and mesocortical) and neuron–astrocyte crosstalk, being descending pathways (mentioned in Table 1) almost uninvestigated in terms of receptor complexes containing DA receptors. Thus, in the sections that follow, only the abovementioned signaling pathways will be considered (see also [108,133,140,141,142] for reviews). These are, however, of significant interest, being associated with an impact on neuropsychiatric diseases. Reported findings are summarized in Table 3.
Table 3.
Complexes involving dopamine receptors in the ascending dopamine pathways and in neuron–astrocyte crosstalk [108,133,140,141,142].
5.1. Nigro-Striatal Dopamine Pathway
The nigro-striatal pathway starts from dopamine-containing cells in the substantia nigra pars compacta (SNc) of the midbrain to establish multiple synaptic contacts with medium spiny neurons (MSNs) of the ipsilateral dorsal striatum [143]. MSNs also receive cortico-striatal glutamatergic afferents and are GABAergic projection neurons classified into three populations [3]. Island (patch) MSNs are localized in the so-called striatosomes [144] and send a feedback signal to neurons of the SNc, striato-nigral/entopenducular MSNs project to the substantia nigra pars reticulata (SNr) and the entopeduncular nucleus (EPN) (nuclei from which the so-called direct pathway of motor control starts), and striato-pallidal MSNs project to the external globus pallidus (GPe) (nucleus from which the indirect pathway of motor control starts), which in turn modulates the subthalamic nucleus (STh). The direct pathway triggers a disinhibition of the target regions, whereas the indirect pathway triggers their inhibition, leading to activation and suppression of motor behavior, respectively. In terms of the dopaminergic modulation of these pathways, the direct pathway is dominated by D1 receptors, expressed at a high level by striato-nigral/entopeduncular MSNs, while the indirect pathway is mainly regulated by D2 receptors, well expressed by striato-pallidal MSNs [3].
As an endogenous neuroprotectant agent, adenosine is extensively distributed in the central nervous system, where it acts trough specific receptors [145], and in the dorsal striatum, A1 and A2A adenosine receptors are widely expressed in both MSNs [133] and glutamatergic terminals [134]. It is not surprising, therefore, that receptor complexes involving adenosine and dopamine receptors were identified in the dorsal striatum. In striato-nigral/entopeduncular MSNs, for instance, the presence of the A1-D1 heterodimer has been reported [116,146], while receptor complexes involving the adenosine A2A and the D2 receptors (namely, the A2A-D2 heterodimer and the heterotrimers A2A-D2-mGluR5 and A2A-D2-CB1) were found in striato-pallidal MSNs and their glutamate inputs [82,84,95]. STh is also innervated by collaterals of the nigro-striatal bundle [143], and co-localization of A2A and D2 receptors has been recently documented in this nucleus [147], opening the possibility of the presence (yet to be substantiated) of A2A-D2 heterodimers within the dorsal and medial aspects of the structure.
Parkinson’s disease (PD) is a common disease, associated with neurodegeneration of the nigro-striatal pathway, leading to imbalance or loss of dopaminergic signaling to the dorsal striatum with the emergence of altered motor features, such as bradykinesias, tremor, and rigidity. The introduction of L-DOPA [148] revolutionized the management of this disease, leading to an effective symptomatic treatment. However, it soon became apparent that the drug offered only symptomatic relief and did not affect the underlying pathology. Moreover, chronic use of the drug was associated with a range of adverse effects, such as dyskinesias, toxicity, or loss of efficacy [149]. Current therapeutic protocols, therefore, seek to delay long-term complications of treatment for as long as possible. In this context, the antagonistic allosteric RRIs described earlier, which characterize the receptor complexes involving adenosine and dopamine receptors, led to the hypothesis (schematically illustrated in Figure 3A) that by targeting these heteromers with antagonists of the adenosine receptors, antiparkinsonian effects could be obtained (see [150] for a specific review on this topic). This research effort mainly focused on A2A-D2 receptor complexes. Animal models of PD gave support to the hypothesis and clinical evidence was also obtained (see [120] for references). In this respect, it is of interest to mention the very recent approval in the United States of an A2A antagonist (istradefylline) as an adjunctive treatment to L-DOPA [151] in PD. Following the same logic, D1 signaling in the A1-D1 heterodimer could be modulated by targeting the adenosine A1 receptor to obtain antiparkinsonian effects [133].
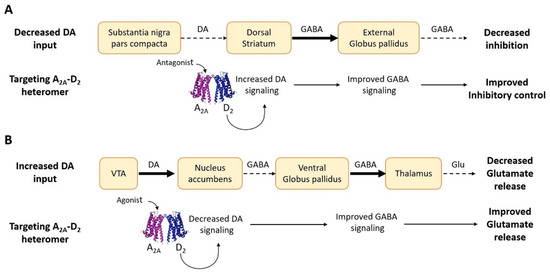
Figure 3.
Schematic representation of pharmacological strategies to address imbalance of DA signaling by targeting the A2A-D2 receptor complex [120]. (A) Decreased DA signaling in the nigro-striatal pathway (as in Parkinson’s disease) leads to a reduced D2 activity and to a decreased inhibitory output from the external globus pallidus to the downstream structures, resulting in unbalanced motor control. Targeting A2A-D2 heteromers in the striatum with antagonists of the A2A receptors may improve D2-mediated dopaminergic signaling and motor control. (B) Overactivity of the mesolimbic dopamine neurons increases the D2-mediated dopamine transmission to the ventral striatum, leading to a reduced glutamate drive from the mediodorsal thalamic nucleus. A2A agonists targeting the antagonistic interactions between A2A and D2 receptors in the complex may improve this condition. Dashed arrows and thick arrows indicate decreased and increased signaling, respectively.
Other receptor complexes in the dorsal striatum, however, deserve a mention as possible pharmacological targets in PD. CB1 antagonists targeting the CB1-D2 heterodimer, for instance, may represent possible antiparkinsonian drugs, since the antagonistic RRIs, characterizing this receptor complex, can enhance D2 signaling [152]. Behavioral correlates to the antagonistic receptor interactions in CB1-D2 heterodimers have also been obtained using the CB1 receptor agonist HU-210, which has been found to reduce L-DOPA-induced rotations in 6-hydroxydopamine-lesioned rats [153]. In cortico-striatal glutamate terminals, the D2-NMDA receptor complex (with antagonistic RRI) is constitutively present [99] and inspired the possibility that a dual approach in PD with low doses of selective D2 agonists and NMDA antagonists could lead to antiparkinsonian actions with reduced development of dyskinesias [133].
5.2. Mesolimbic Dopamine Pathway
The mesolimbic pathway connects the ventral tegmental area (VTA), a dopaminergic nucleus of the midbrain, with the ventral striatum (occupying about 20% of the striatum), including the nucleus accumbens (NAc) and the olfactory tubercle, which are striatal regions receiving their major telencephalic input from the hippocampal formation and amygdala, and projecting to the ventral pallidum (VP) and SNr. From there, information is transferred to the anterior cingulate cortex and the orbitofrontal cortex [154]. Concerning the NAc, two main subterritories have been identified, namely, the shell and the core, the shell region being more closely associated with the limbic system than the other regions of the ventral striatum [3].
Ventral striatum neurons are MSNs, similar to those of the dorsal striatum, and their dopaminergic input are mainly regulated by D2 receptors [155]. A2A-D2 heteroreceptor complexes with antagonistic RRIs were demonstrated in the ventral striatum [125], as were high-order receptor complexes including adenosine A2A and dopamine D2 receptors, such as, for instance, the A2A-D2-mGluR5 and A2A-D2-sigma1 heterotrimers [156]. Of interest is also the presence in ventral MSNs of cortico-accumbens terminals of receptor complexes involving dopamine D2, glutamate NMDA [99], neurotensin NTS1 [100], serotonin 5-HT2A [104], and oxytocin [105] receptors.
The mesolimbic pathway is a key element of the so-called reward circuit (see [154]), because the release of dopamine through this pathway regulates motivation and desire for rewarding stimuli (i.e., incentive salience), facilitates reinforcement- and reward-related motor function learning, and may also play a role in the subjective perception of pleasure. Thus, the dysregulation of the mesolimbic pathway and its downstream neurons plays a significant role in the development of significant neuropsychiatric diseases, including addiction and schizophrenia (see [156,157] for specific reviews).
A study [158], for instance, showed that chronic cocaine self-administration increased behavioral responses mediated by D2 receptors, indicating the relevance of D2 for cocaine use disorder. Furthermore, chronic cocaine self-administration persistently evoked more than 100% elevations of D2 binding sites of the high-affinity type [159], and D2 activation produced a strong relapse of cocaine seeking in animals [160]. In this respect, studies focused on the antagonistic RRIs in the A2A-D2 receptor complex as a possible pharmacological target indicated that A2A agonists exhibited an inhibitory effect on cocaine reward [160], and A2A activation, leading to D2-like receptor blockade, counteracted cocaine relapse. It is also of interest that cocaine induces a selective increase in sigma1 receptors in the ventral striatum [161]. Thus, the A2A-D2 antagonistic interaction may become more present thanks to a higher presence of A2A-D2-sigma1 receptor complexes. In this context, also results suggesting the existence of D4-MOR heterodimers [110] in the striatosomes and SNr, in which D4-MOR interactions are in operation, are also of interest. They may play a critical role in at least the early stages of the expression of the morphine effects. In view of the limbic-prefrontal–striatosome-nigral circuitry and its function (see [162]), this interaction may participate in reward-based motor learning and play a significant role in habit acquisition in drug addiction [133].
In schizophrenia, salience becomes exaggerated due, inter alia, to an increased D2 recognition and signaling in the ventral striatum (mainly nucleus accumbens) [163]. Thus, the classic treatment [164] in schizophrenia is the use of DA receptor antagonists, typically haloperidol and chlorpromazine. Through the blockade of excessive D2-mediated DA transmission in the mesolimbic dopaminergic pathway, they allow an improvement of mental symptoms, but induce motor side effects due to the parallel block of the nigrostriatal pathway. Thus, based on the presence in the ventral GABAergic MSNs, in astrocytes, and in glutamatergic terminals of A2A-D2 containing heteroreceptor complexes with antagonistic A2A-D2 interactions, the use of A2A receptor agonists (see Figure 3B) as a strategy for the treatment of schizophrenia has been proposed [133] and promising results in animal models have been found [165]. It is worth noting that A2A agonist treatment, especially in combination with low doses of typical and/or atypical D2 antagonists, could also represent a possible strategy for reducing the development of extrapyramidal side effects [133]. Facilitatory RRIs in the 5-HT2A-D2 receptor complex may represent a further target for treatments based on antagonists of the serotonin receptor (see [157]), and a reduction in the inhibitory D2 signaling at the cortico-accumbens glutamatergic terminal level could be obtained by targeting NTS1-D2 receptor complexes with agonists of the neurotensin receptor [101]. The D2-OTR heterodimer also deserves interest as a possible target in schizophrenia. Indeed, evidence was obtained that the molecular mechanism mediating the social salience was the formation of D2-OTR heteroreceptor complexes in the nucleus accumbens core [105]. In fact, being located to a special component of the ventral GABAergic MSNs involved in regulating a brain circuit reaching into the prefrontal cortex, the result of the activation of the D2-OTR heteroreceptor complex may produce social attachment and trust and the negative symptoms of schizophrenia may become markedly reduced [140]. Consistent with this hypothesis are data showing that oxytocin can induce antipsychotic actions [166], which appears to be true after being given to schizophrenic patients intranasally [167].
5.3. Mesocortical Dopamine Pathway
The mesocortical pathway connects the VTA to the prefrontal cortex, but dopaminergic axons branch within the cortex to reach multiple cortical areas [3]. By applying a modified Falck–Hillarp technique, Hökfelt and coworkers [168] identified a plexus of dopaminergic fibers in the limbic cortex with an uneven innervation of the entorhinal cortex. DA-containing varicosities preferentially establish synaptic contacts on pyramidal neurons [169].
This pathway is essential to the normal cognitive function of the dorsolateral prefrontal cortex (part of the frontal lobe) and is thought to be involved in cognitive control, motivation, and emotional response [170]. In this respect, it is closely associated with the mesolimbic pathway.
As recently discussed by Ferré and collaborators [108], an interesting aspect of this innervation pattern is the high expression of dopamine D4 receptors in the cortex of mammals: most glutamatergic pyramidal neurons and about half of the GABAergic interneurons express D4. Considering the Gi-coupled D4 as mostly inhibitory, the D4 localized in neurons should be expected to exhibit an inhibitory effect on dopamine, while those localized in GABAergic interneurons should be expected to produce disinhibition. Several studies, however, indicate a more complex picture, associated with evidence indicating that D4 receptors can form receptor complexes with adrenergic receptors [171]. As briefly discussed in Section 4.1, these receptor complexes may have significantly different pharmacological properties depending on the polymorphic variant of the D4 receptor involved.
In this respect, available evidence associating D4 polymorphisms with individual differences in impulse control-related neuropsychiatric disorders is of interest, with the most consistent associations found between the gene encoding D4.7 and attention-deficit hyperactivity disorder (ADHD) [172]. On this basis, it has been proposed that receptor complexes involving the D4 receptor should be investigated as possible therapeutic targets for ADHD, as well as for restless legs syndrome [108].
5.4. Neuron–Astrocyte Crosstalk
Increasing evidence (see [141] for a specific review) indicates that astrocytes are directly involved in the regulation of neuronal excitability and action potential propagation. According to this view, a bidirectional relationship exists between astrocytes and neurons, where neural activity influences astrocytic activation, which in turn modulates the activity of neurons [173].
Astrocytes, indeed, monitor the extracellular environment through specific receptors, including many neurotransmitter receptors (such as those for DA). Single astrocytes integrate this information through the elevation of intracellular Ca2+ [141] and can propagate this information over large distances by communicating with each other through calcium waves [174]. Such calcium dynamics are considered a key step leading to the release of gliotransmitters (D-serine, ATP, and glutamate) that regulate ongoing neural activity [175]. As indicated by several experimental studies (see [173] for a review), this intercellular crosstalk significantly influences synaptic plasticity and, consequently, higher CNS functions such as, for instance, learning and memory.
In this context, extensive available data indicate that RRIs may play a significant role. Relevant examples include the heterodimers A2A-D2 and D2-OTR [96,106], formed by the association of the dopamine D2 receptor with the adenosine A2A or the oxytocin receptor, respectively. These receptor complexes are present in astrocytes and regulate the release of glutamate from these cells [106,127], a process relevant for the control of glutamatergic transmission in striatum and with potential roles in the dysregulation of glutamatergic transmission in various neuropsychiatric diseases (see [176] for a specific review on this topic).
The results of a study [177], showing that knocking down the striatal astrocytic glutamate transporter GLT-1 induces PD-like changes in rodents, illustrate the importance of the regulation of the striatal extracellular glutamate level by astrocytes in this pathology. Furthermore, dopamine-mediated glutamate release from striatal astrocyte processes can modulate the activation of NMDA and metabotropic glutamate receptors on striatal MSN [178], suggesting the abovementioned receptor complexes as potential targets to counteract striatal glutamatergic transmission disfunctions and circuit derangement in PD [176]. In this respect, an interesting possibility was suggested by findings showing that homocysteine (an allosteric modulator of the A2A-D2 heterodimer, see Section 4.1) was able to counteract the DA-mediated inhibition of glutamate release by astrocytes [130]. The relevance of this finding from a physio-pathological standpoint can be appreciated when considering that L-DOPA treatment can trigger synthesis of homocysteine in astrocytes and their release into the extracellular space [179].
Evidence indicating astrocyte involvement in schizophrenia has also been collected [180,181], where glial abnormalities were proposed to contribute to glutamatergic and dopaminergic neurotransmission dysfunctions [182]. In a mouse model of astrocytic A2A receptor knockout, for instance, impaired glutamate homeostasis associated with enhanced behavioral sensitization to psychoactive drugs and reduced working memory (two behavioral symptoms of the pathology) was reported [180]. Thus, the astrocytic A2A-D2 heteromers may represent a possible target for A2A agonist or other drugs (see [183,184]) in order to ameliorate the impaired glutamate homeostasis in schizophrenia.
Regulation of astrocytic RRIs involving the D2 receptor can also be of importance for the pathophysiology and treatment of drug addiction. Accumulating evidence, indeed, indicates that drugs of abuse can trigger glutamatergic dysregulation through astroglial mechanisms (see [185]). On this background, D2-containing heteromers in astrocytes may provide new perspectives in the search for drug addiction therapies.
6. Concluding Remarks
Since the discovery of DA as a neurotransmitter, the relationship between the dopaminergic signaling network and essential physiological and pathological processes in the nervous systems has become clear. The dopaminergic system is a complex system, organized in parallel and segregated functional streams consisting of motor, reward (limbic), and associative (cognitive) control pathways [186]. However, evidence also exists that the system also exploits integrative mechanisms by which information is transferred between these functional circuits (see [3]). Furthermore, it extensively interacts with other critical signaling pathways [6]. Such a complex intercellular communication occurs through both synaptic and volume transmission (see [64]) and is mediated by a set of GPCRs.
In this respect, extensive evidence has been provided showing that DA receptors can also establish direct allosteric RRIs with other receptor proteins, leading to the formation of receptor complexes and allowing a modulation of signal decoding already at the membrane level and characterized by specific pharmacological profiles; these are potentially of interest to devise new strategies to address relevant disorders. As briefly discussed here, in recent decades, an increasing number of receptor complexes involving DA receptors have been identified and studied. Several aspects, however, remain to be addressed to better understand their function and the possibilities that their targeting may offer.
As previously suggested [157], a first point (of a neuroanatomical nature) we would like to emphasize concerns the need for a more detailed mapping of the different DA-receptor-containing receptor complexes to better understand their distribution in the dopaminergic pathways and to better characterize their location at the cellular level. In this regard, of particular interest would be the study of the descending dopamine pathways, since almost no data concerning the distribution of receptor complexes containing DA receptors in these districts have been obtained so far. A second point (of a pharmacological nature) involves a more detailed assessment of how typical and atypical neuropsychiatric drugs may act on the different receptor complexes in order to optimize existing pharmacological treatments or to develop completely new pharmacological strategies. In this respect, however, the development of receptor-complex-specific ligands appears another very promising strategy. Indeed, the possibility to develop bivalent ligands [187] or to exploit allosteric modulators that are selective for structural domains in the heteroreceptor complexes [129,130] has been demonstrated.
Finally, it should be noted that the research effort to identify and characterize RRIs and receptor complexes has been mainly focused on neurons, given that available data on RRIs and on receptor complexes in astrocytes are more limited. However, a more intense effort in pharmacological research applied to receptor complexes in astrocytes may represent a topic of particular interest, not only to reach a better understanding of the role of neuron–astrocyte crosstalk in dopaminergic systems, but also from a therapeutical standpoint. Such a research effort, indeed, may open the possibility of exploring novel, glia-mediated strategies to address neurodegenerative and functional DA-related disorders (see [141]).
Author Contributions
Conceptualization, D.G. and L.F.A.; data collection, C.T., M.M. and C.C.; writing—original draft preparation, D.G.; writing—review and editing, G.M., R.D.C. and L.F.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dahlström, A.; Fuxe, K. A method for the demonstration of monoamine-containing nerve fibres in the central nervous system. Acta Physiol. 1964, 60, 293–294. [Google Scholar] [CrossRef] [PubMed]
- Andén, N.-E.; Carlsson, A.; Dahlström, A.; Fuxe, K.; Hillarp, N.A.; Larsson, K. Demonstration and mapping out of nigro-neostriatal dopamine neurons. Life Sci. 1964, 3, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Bentivoglio, M.; Morelli, M. The organization and circuits of mesencephalic dopaminergic neurons and the distribution of dopamine receptors in the brain. In Dopamine; Dunnet, S.B., Bentivoglio, M., Björklund, A., Hökfelt, T., Eds.; Handbook of Chemical Neuroanatomy; Elsevier: Amsterdam, The Netherlands, 2005; Volume 21, pp. 1–107. [Google Scholar]
- Hurd, Y.L.; Hall, H. Human forebrain dopamine systems: Characterization of the normal brain and in relation to psychiatric disorders. In Dopamine; Dunnet, S.B., Bentivoglio, M., Björklund, A., Hökfelt, T., Eds.; Handbook of Chemical Neuroanatomy; Elsevier: Amsterdam, The Netherlands, 2005; Volume 21, pp. 525–571. [Google Scholar]
- Albanese, A.; Altavista, M.C.; Rossi, P. Organization of central nervous system dopaminergic pathways. J. Neural Transm. 1986, 22, 3–17. [Google Scholar]
- Klein, M.O.; Battagello, D.S.; Cardoso, A.R.; Hauser, D.N.; Bittencourt, J.C.; Correa, R.G. Dopamine: Function, signaling, and association with neurological diseases. Cell. Mol. Neurobiol. 2019, 39, 31–59. [Google Scholar]
- Fuxe, K.; Rivera, A.; Jacobsen, K.X.; Höistad, M.; Leo, G.; Horvath, T.L.; Staines, W.; De la Calle, A.; Agnati, L.F. Dynamics of volume transmission in the brain. Focus on catecholamine and opioid peptide communication and the role of uncoupling protein 2. J. Neural Transm. 2005, 112, 65–76. [Google Scholar] [CrossRef]
- Agnati, L.F.; Guidolin, D.; Guescini, M.; Genedani, S.; Fuxe, K. Understanding wiring and volume transmission. Brain Res. Rev. 2010, 64, 137–159. [Google Scholar]
- Guidolin, D.; Marcoli, M.; Maura, G.; Agnati, L.F. New dimensions of connectomics and network plasticity in the central nervous system. Rev. Neurosci. 2017, 28, 113–132. [Google Scholar]
- Eid, L.; Parent, M. Chemical anatomy of pallidal afferents in primates. Brain Struct. Funct. 2016, 221, 4291–4317. [Google Scholar]
- Rice, M.E.; Cragg, S.J. Dopamine spillover after quantal release: Rethinking dopamine transmission in the nigrostriatal pathway. Brain Res. Rev. 2008, 58, 303–313. [Google Scholar] [CrossRef]
- Rice, M.E.; Patel, J.C. Somatodendritic dopamine release: Recent mechanistic insights. Phil. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140185. [Google Scholar] [CrossRef]
- Goto, Y.; Otani, S.; Grace, A.A. The yin and yang of dopamine release: A new perspective. Neuropharmacology 2007, 53, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Floresco, S.B. Dopaminergic regulation of limbic-striatal interplay. J. Psychiatry Neurosci. 2007, 32, 400–411. [Google Scholar] [PubMed]
- Hirasawa, H.; Contini, M.; Raviola, E. Extrasynaptic release of GABA and dopamine by retinal dopaminergic neurons. Phylos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140186. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, I.; Asanuma, M.; Diaz-Corrales, F.J.; Miyoshi, K.; Ogawa, N. Direct evidence for expression of dopamine receptors in astrocytes from basal ganglia. Brain Res. 2004, 1029, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Mladinov, M.; Mayer, D.; Brčic, L.; Wolstencroft, E.; thi Man, N.; Holt, I.; Hof, P.R.; Morris, G.E.; Šimic, G. Astrocyte expression of D2-like dopamine receptors in the prefrontal cortex. Transl. Neurosci. 2010, 1, 238–243. [Google Scholar] [CrossRef]
- Montoya, A.; Elgueta, D.; Campos, J.; Chovar, O.; Falcòn, P.; Matus, S.; Alfaro, I.; Bono, M.R.; Pacheco, R. Dopamine receptor D3 signalling in astrocytes promotes neuroinflammation. J. Neuroinflamm. 2019, 16, 258. [Google Scholar] [CrossRef]
- Guidolin, D.; Tortorella, C.; Marcoli, M.; Cervetto, C.; Maura, G.; Agnati, L.F. Receptor-receptor interactions and microvesicle exchange as mechanisms modulating signaling between neurons and astrocytes. Neuropharmacology 2023, 231, 109509. [Google Scholar]
- Kebabian, J.W.; Petzold, G.L.; Greengard, P. Dopamine-sensitive adenylate cyclase in caudate nucleus of rat brain, and its similarity to the “dopamine receptor”. Proc. Natl. Acad. Sci. USA 1972, 69, 2145–2149. [Google Scholar] [CrossRef]
- Burt, D.R.; Enna, S.J.; Creese, I.; Snyder, S.H. Dopamine receptor binding in the corpus striatum of mammalian brain. Proc. Natl. Acad. Sci. USA 1975, 72, 4655–4659. [Google Scholar] [CrossRef]
- Seeman, P.; Chau-Wong, M.; Tedesco, J.; Wong, K. Brain receptors for antipsychotic drugs and dopamine: Direct binding assays. Proc. Natl. Acad. Sci. USA 1975, 72, 4376–4380. [Google Scholar] [CrossRef]
- Fuxe, K.; Agnati, L.F.; Benfenati, F.; Celani, M.; Zini, I.; Zoli, M.; Mutt, V. Evidence for the existence of receptor-receptor interactions in the central nervous system. Studies on the regulation of monoamine receptors by neuropeptides. In Basic Aspects of Receptor Biochemistry; Springer: Vienna, Austria, 1983; Volume 18, pp. 165–179. [Google Scholar]
- Bockaert, J.; Pin, J.P. Molecular tinkering of G protein coupled receptors: An evolutionary success. EMBO J. 1999, 18, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Marshall, F.H.; White, J.; Main, M.; Green, A.; Wise, A. GABA(B) receptors function as heterodimers. Biochem. Soc. Trans. 1999, 27, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Lee, S.P.; O’Dowd, B.F.; George, S.R. Serotonin 5-HT1B and 5-HT1D receptors form homodimers when expressed alone and heterodimers when co-expressed. FEBS Lett. 1999, 456, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.P.; Xie, Z.; Varghese, G.; Nguyen, T.; O’Dowd, B.F.; George, S.R. Oligomerization of dopamine and serotonin receptors. Neuropsychopharmacology 2000, 23, S32–S40. [Google Scholar] [CrossRef]
- Overton, M.C.; Blumer, K.J. G protein-coupled receptors function as oligomers in vivo. Curr. Biol. 2000, 10, 341–344. [Google Scholar] [CrossRef]
- Zeng, F.; Wess, J. Molecular aspects of muscarinic receptor dimerization. Neuropsychopharmacology 2000, 23, S19–S31. [Google Scholar] [CrossRef]
- Angers, S.; Salahpour, A.; Bouvier, M. Biochemical and biophysical demonstration of GPCR oligomerization in mammalian cells. Life Sci. 2001, 68, 2243–2250. [Google Scholar] [CrossRef]
- Dean, M.K.; Higgs, C.; Smith, R.E.; Bywater, R.P.; Snell, C.R.; Scott, P.D.; Upton, G.J.; Howe, T.J.; Reynolds, C.A. Dimerization of G protein-coupled receptors. J. Med. Chem. 2001, 44, 4595–4614. [Google Scholar]
- Kenakin, T. Drug efficacy at G protein-coupled receptors. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 349–379. [Google Scholar] [CrossRef]
- Waldhoer, M.; Fong, J.; Jones, R.M.; Lunzer, M.M.; Sharma, S.K.; Kostenis, E.; Portoghese, P.S.; Whistler, J.L. A heterodimer-selective agonist shows in vivo relevance of G protein-coupled receptor dimers. Proc. Natl. Acad. Sci. USA 2005, 102, 9050–9055. [Google Scholar] [CrossRef]
- Guidolin, D.; Marcoli, M.; Tortorella, C.; Maura, G.; Agnati, L.F. Receptor-receptor interactions as a widespread phenomenon: Novel targets for drug development? Front. Endocrinol. 2019, 10, 53. [Google Scholar]
- Changeux, J.P.; Christopoulos, A. Allosteric modulation as a unifying mechanism for receptor function and regulation. Diabetes Obes. Metabol. 2017, 19, 4–21. [Google Scholar] [CrossRef] [PubMed]
- Baik, J.-H. Dopamine Signaling in reward-related behaviors. Front. Neural Circuits 2013, 7, 152. [Google Scholar] [CrossRef] [PubMed]
- Missale, C.; Nash, S.R.; Robinson, S.W.; Jaber, M.; Caron, M.G.; Wishart, D.S.; Anselmi, L.; Toti, L.; Bove, C.; Prakash, Y.S.; et al. Dopamine receptors: From structure to function. Physiol. Rev. 1998, 78, 189–225. [Google Scholar]
- Beaulieu, J.-M.; Gainetdinov, R.R. The physiology, signaling, and pharmacology of dopamine receptors. Pharmacol. Rev. 2011, 63, 182–217. [Google Scholar]
- Tritsch, N.X.; Sabatini, B.L. Dopaminergic modulation of synaptic transmission in cortex and striatum. Neuron 2012, 76, 33–50. [Google Scholar]
- Gingrich, J.A.; Caron, M.G. Recent advances in the molecular biology of dopamine receptors. Annu. Rev. Neurosci. 1993, 16, 299–321. [Google Scholar] [CrossRef]
- Dal Toso, R.; Sommer, B.; Ewert, M.; Herb, A.; Pritchett, D.B.; Bach, A.; Shivers, B.D.; Seeburg, P.H. The dopamine D2 receptor: Two molecular forms generated by alternative splicing. EMBO J. 1989, 8, 4025–4034. [Google Scholar] [CrossRef]
- Giros, B.; Sokoloff, P.; Martres, M.P.; Riou, J.-F.; Emorine, L.J.; Schwartz, J.-C. Alternative splicing directs the expression of two D2 dopamine receptor isoforms. Nature 1989, 342, 923–926. [Google Scholar] [CrossRef]
- Kim, K.-M. Unveiling the differences in signaling and regulatory mechanisms between dopamine D2 and D3 receptors and their impact on behavioral sensitization. Int. J. Mol. Sci. 2023, 24, 6742. [Google Scholar]
- Jose, P.A.; Yu, P.-Y.; Yamapchi, I.; Eisner, G.M.; Mouradian, M.M.; Felder, C.C.; Felder, R.A. Dopamine D1 receptor regulation of phospholipase C. Hypertens. Res. 1995, 18 (Suppl. 1), S39–S42. [Google Scholar] [CrossRef]
- Rashid, A.; O’Dowd, B.F.; Verma, V.; George, S.R. Neuromal Gq/11-coupled dopamine receptors: An uncharted role for dopamine. Trends Pharmacol. Sci. 2007, 28, 551–555. [Google Scholar] [CrossRef]
- Sahu, A.; Tyeryar, K.R.; Vongtau, H.O.; Sibley, D.R.; Undieh, A.S. D5 dopamine receptors are required for dopaminergic activation of phospholipase C. Mol. Pharmacol. 2009, 75, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Felder, C.C.; Jose, P.A.; Axelrod, J. The dopamine-1 agonist, SKF 82526, stimulates phospholipase-C activity independent of adenylate cyclase. J. Pharmacol. Exp. Ther. 1989, 248, 171–175. [Google Scholar] [PubMed]
- Kofuji, P.; Araque, A. G-protein-coupled receptors in astrocyte-neuron communication. Neuroscience 2021, 456, 71–84. [Google Scholar] [PubMed]
- Oda, S.; Funato, H. D1- and D2-type dopamine receptors are immunolocalized in pial and layer I astrocytes in the rat cerebral cortex. Front. Neuroanat. 2023, 17, 1111008. [Google Scholar] [CrossRef] [PubMed]
- Jennings, A.; Tyurikova, O.; Bard, L.; Zheng, K.; Semyanov, A.; Henneberger, C.; Rusakov, D.A. Dopamine elevates and lowers astroglial Ca(2+) through distinct pathways depending on local synaptic circuitry. Glia 2017, 65, 447–459. [Google Scholar] [CrossRef]
- Xin, W.; Schuebel, K.E.; Jair, K.W.; Cimbro, R.; De Biase, L.M.; Goldman, D.; Bonci, A. Ventral midbrain astrocytes display unique physiological features and sensitivity to dopamine D2 receptor signaling. Neuropsychopharmacology 2019, 44, 344–355. [Google Scholar]
- Corkrum, M.; Covelo, A.; Lines, J.; Bellocchio, L.; Pisansky, M.; Loke, K.; Quintana, R.; Rothwell, P.E.; Lujan, R.; Marsicano, G.V.; et al. Dopamine-evoked synaptic regulation in the nucleus accumbens requires astrocyte activity. Neuron 2020, 105, 1036–1047. [Google Scholar]
- Li, C.; Saliba, N.B.; Martin, H.; Losurdo, N.A.; Kolahdouzan, K.; Siddiqui, R.; Medeiros, D.; Li, W. Purkinje cell dopaminergic inputs to astrocytes regulate cerebellar-dependent behavior. Nat. Commun. 2023, 14, 1613. [Google Scholar]
- Gurevich, V.V.; Gurevich, E.V. The structural basis of arrestin-mediated regulation of G-protein-coupled receptors. Pharmacol. Ther. 2006, 110, 465–502. [Google Scholar] [PubMed]
- Komolov, K.E.; Benovic, J.L. G protein-coupled receptor kinases: Past, present and future. Cell. Signal. 2018, 41, 17–24. [Google Scholar] [PubMed]
- Pitcher, J.A.; Freedman, N.J.; Lefkowitz, R.J. G protein-coupled receptor kinases. Annu. Rev. Biochem. 1998, 67, 653–692. [Google Scholar]
- Lohse, M.J.; Benovic, J.L.; Codina, J.; Caron, M.G.; Lefkowitz, R.J. beta-Arrestin: A protein that regulates beta-adrenergic receptor function. Science 1990, 248, 1547–1550. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, L.M.; Lefkowitz, R.J. The role of beta-arrestins in the termination and transduction of G-protein-coupled receptor signals. J. Cell Sci. 2002, 115, 455–465. [Google Scholar] [CrossRef]
- Hollinger, S.; Hepler, J.R. Cellular regulation of RGS proteins: Modulators and integrators of G protein signaling. Pharmacol. Rev. 2002, 34, 527–559. [Google Scholar] [CrossRef]
- Woodard, G.E.; Jardin, I.; Berna-Erro, A.; Salido, G.M.; Rosado, J.A. Regulators of G-protein-signaling proteins: Negative modulators of G-protein-coupled receptor signaling. Int. Rev. Cell Mol. Biol. 2015, 317, 97–183. [Google Scholar]
- Kovoor, A.; Seyffarth, P.; Ebert, J.; Barghshoon, S.; Chen, C.-K.; Schwarz, S.; Axelrod, J.D.; Cheyette, B.N.R.; Simon, M.I.; Lester, H.A.; et al. D2 dopamine receptors colocalize regulator of G-protein signaling 9-2 (RGS9-2) via the RGS9 DEP domain, and RGS9 knock-out mice develop dyskinesias associated with dopamine pathways. J. Neurosci. 2005, 25, 2157–2165. [Google Scholar] [CrossRef]
- Celver, J.; Sharma, M.; Kovoor, A. RGS9-2 mediates specific inhibition of agonist-induced internalization of D2-dopamine receptors. J. Neurochem. 2010, 114, 739–749. [Google Scholar] [CrossRef]
- Fuxe, K.; Canals, M.; Torvinen, M.; Marcellino, D.; Terasmaa, A.; Genedani, S.; Leo, G.; Guidolin, D.; Diaz-Cabiale, Z.; Rivera, A.; et al. Intramembrane receptor-receptor interactions: A novel principle in molecular medicine. J. Neural Transm. 2007, 114, 49–75. [Google Scholar]
- Guidolin, D.; Tortorella, C.; Marcoli, M.; Maura, G.; Agnati, L.F. Intercellular communication in the central nervous system as deduced by chemical neuroanatomy and quantitative analysis of images: Impact on neuropharmacology. Int. J. Mol. Sci. 2022, 23, 5805. [Google Scholar]
- Prezeau, L.; Rives, M.L.; Comps-Agrar, L.; Maurel, D.; Knlazeff, J.; Pin, J.P. Functional crosstalk between GPCRs: With or without oligomerization. Curr. Opin. Pharm. 2010, 10, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Agnati, L.F.; Fuxe, K.; Giardino, L.; Calzà, L.; Zoli, M.; Battistini, N.; Benfenati, F.; Vanderhaeghen, J.J.; Guidolin, D.; Ruggeri, M. Evidence for cholecystokinin-dopamine receptor interactions in the central nervous system of the adult and old rat. Studies on their functional meaning. Ann. N. Y. Acad. Sci. 1985, 448, 315–333. [Google Scholar] [CrossRef] [PubMed]
- Kenakin, T.; Agnati, L.F.; Caron, M.; Fredholm, B.; Guidolin, D.; Kobilka, B.; Lefkowitz, R.W.; Lohse, M.; Woods, A.; Fuxe, K. International workshop at the Nobel Forum, Karolinska Institutet, on G protein-coupled receptors: Finding the words to describe monomers, oligomers, and their molecular mechanisms and defining their meaning. Can a consensus be reached? J. Recept. Signal Transduct. Res. 2010, 30, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Trifilieff, P.; Rives, M.L.; Urizar, E.; Piskorowski, R.A.; Vishwasrao, H.D.; Castrillon, J.; Schmauss, C.; Stätmann, M.; Gullberg, M.; Javitch, J.A. Detection of antigen interactions ex vivo by proximity ligation assay: Endogenous dopamine D2-adenosine A2A receptor complexes in the striatum. Biotechniques 2011, 51, 111–118. [Google Scholar] [CrossRef]
- Fernández-Dueñas, V.; Gòmez-Soler, M.; Valle-Leòn, M.; Watanabe, M.; Ferrer, I.; Ciruela, F. Revealing adenosine A2A-dopamine D2 receptor heteromers in Parkinson’s disease post-mortem brain through a new AlphaScreen-based approach. Int. J. Mol. Sci. 2019, 20, 3600. [Google Scholar] [CrossRef]
- Petazzi, R.A.; Aji, A.K.; Chiantia, S. Fluorescence microscopy methods for the study of protein oligomerization. In Progress in Molecular Biology and Translational Science; Giraldo, J., Ciruela, F., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 169, pp. 1–42. [Google Scholar]
- De Oliveira, P.; Moreno, E.; Casajuana-Martin, N.; Casadò-Anguera, V.; Cai, N.-S.; Camacho-Hernandez, G.A.; Zhu, H.; Bonifazi, A.; Hall, M.D.; Weinshenker, D.; et al. Preferential Gs protein coupling of the galanin Gal1 receptor in the μ-opioid-Gal1 receptor heterotetramer. Pharmacol. Res. 2022, 182, 106322. [Google Scholar] [CrossRef]
- Franco, R.; Martinez-Pinilla, E.; Lanciego, J.L.; Navarro, G. Basic pharmacological and structural evidence for class A G-protein-coupled receptor heteromerization. Front. Pharmacol. 2016, 7, 76. [Google Scholar]
- Changeux, J.P. The origins of allostery: From personal memories to material for the future. J. Mol. Biol. 2013, 425, 1396–1406. [Google Scholar]
- Kenakin, T.; Miller, I.J. Seven transmembrane receptors as shape shifting proteins: The impact of allosteric modulation and functional selectivity on new drug discovery. Pharm. Rev. 2010, 62, 265–304. [Google Scholar]
- Smith, N.J.; Milligan, G. Allostery of G protein-coupled receptors homo- and heteromers: Uncharted pharmacological landscapes. Pharm. Rev. 2010, 62, 701–725. [Google Scholar] [PubMed]
- Liu, J.; Nussinov, R. Allostery: An overview of its history, concepts, methods and applications. PLoS Comput. Biol. 2016, 12, e1004966. [Google Scholar] [CrossRef] [PubMed]
- Ferré, S.; Ciruela, F.; Woods, A.S.; Lluis, C.; Franco, R. Functional relevance of neurotransmitter receptor heteromers in the central nervous system. Trends Neurosci. 2007, 30, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Filizola, M.; Weinstein, H. The study of G-protein coupled receptor oligomerization with computational modeling and bioinformatics. FEBS J. 2005, 272, 2926–2938. [Google Scholar] [PubMed]
- Simpson, L.M.; Taddese, B.; Wall, I.D.; Reynolds, C.A. Bioinformatics and molecular modelling approaches to GPCR oligomerization. Curr. Opin. Pharm. 2010, 10, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Guidolin, D.; Ciruela, F.; Genedani, S.; Guescini, M.; Tortorella, C.; Albertin, G.; Fuxe, K.; Agnati, L.F. Bioinformatics and mathematical modeling in the study of receptor-receptor interactions and receptor oligomerization. Focus on adenosine receptors. Biochim. Biophys. Acta 2011, 1808, 1267–1283. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Tarakanov, A.O.; Brito, I.; Fuxe, K. Glutamate heteroreceptor complexes in the brain. Pharmacol. Rep. 2018, 70, 936–950. [Google Scholar]
- Pinna, A.; Bonaventura, J.; Farré, D.; Sànchez, M.; Simola, N.; Mallol, J.; Lluís, C.; Costa, G.; Baqi, Y.; Müller, C.E.; et al. L-DOPA disrupts adenosine A(2A)-cannabinoid CB(1)-dopamine D(2) receptor heteromer cross-talk in the striatum of hemiparkinsonian rats: Biochemical and behavioral studies. Exp. Neurol. 2014, 253, 180–191. [Google Scholar] [CrossRef]
- Cabello, N.; Gandia, J.; Bertarelli, D.C.; Watanabe, M.; Lluis, C.; Franco, R.; Ferré, S.; Luján, R.; Ciruela, F. Metabotropic glutamate type 5, dopamine D2 and adenosine A2a receptors form higher-order oligomers in living cells. J. Neurochem. 2009, 109, 1497–1507. [Google Scholar] [CrossRef]
- Beggiato, S.; Tomasini, M.C.; Borelli, A.C.; Borroto-Escuela, D.O.; Fuxe, K.; Antonelli, T.; Tanganelli, S.; Ferraro, L. Functional role of striatal A2A, D2, and mGlu5 receptor interactions in regulating striatopallidal GABA neuronal transmission. J. Neurochem. 2016, 138, 254–264. [Google Scholar] [CrossRef]
- Farran, B. An update on the physiological and therapeutic relevance of GPCR oligomers. Pharmacol. Res. 2017, 117, 303–327. [Google Scholar] [PubMed]
- Gainetdinov, R.R.; Premont, R.T.; Bohn, L.M.; Lefkowitz, R.J.; Caron, M.G. Desensitization of G protein-coupled receptors and neural functions. Annu. Rev. Neurosci. 2004, 27, 107–144. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.S.; Rajagopal, S. The β-arrestin: Multifunctional regulators of G protein-coupled receptors. J. Biol. Chem. 2016, 291, 8969–8977. [Google Scholar] [PubMed]
- Agnati, L.F.; Leo, G.; Genedani, S.; Andreoli, N.; Marcellino, D.; Woods, A.; Piron, L.; Guidolin, D.; Fuxe, K. Structural plasticity in G-protein coupled receptors as demonstrated by the allosteric actions of homocysteine and computer-assisted analysis of disordered domains. Brain Res. Rev. 2008, 58, 459–474. [Google Scholar] [PubMed]
- Agnati, L.F.; Guidolin, D.; Vilardaga, J.P.; Ciruela, F.; Fuxe, K. On the expanding terminology in the GPCR field: The meaning of receptor mosaics and receptor heteromers. J. Recept. Signal Transduct. Res. 2010, 30, 287–303. [Google Scholar] [PubMed]
- Alemany, R.; Perona, J.S.; Sánchez-Dominguez, J.M.; Montero, E.; Cañizares, J.; Bressani, R.; Escribà, P.V.; Ruiz-Gutierrez, V.G. protein-coupled receptor systems and their lipid environment in health disorders during aging. Biochim. Biophys. Acta 2007, 1768, 964–975. [Google Scholar]
- Foord, S.M.; Jupe, S.; Holbrook, J. Bioinformatics and type II G-protein-coupled receptors. Biochem. Soc. Trans. 2002, 30, 473–479. [Google Scholar] [CrossRef]
- Whorton, M.R.; Bokoch, M.P.; Rasmussen, S.G.F.; Huang, B.; Zare, R.N.; Kobilka, B.; Sunahara, R.K. A monomeric G protein-coupled receptor isolated in a high-density lipoprotein particle efficiently activates its G protein. Proc. Natl. Acad. Sci. USA 2007, 104, 7682–7687. [Google Scholar]
- Gurevich, V.V.; Gurevich, E.V. How and why do GPCRs dimerize? Trends Pharmacol. Sci. 2008, 29, 234–240. [Google Scholar]
- Teichmann, A.; Gibert, A.; Lampe, A.; Grzesik, P.; Rutz, C.; Furkert, J.; Schmoranzer, J.; Krause, G.; Wiesner, B.; Schülein, R. The specific monomer/dimer equilibrium of the corticotropin-releasing factor receptor type 1 is established in the endoplasmic reticulum. J Biol. Chem. 2014, 289, 24250–24262. [Google Scholar] [CrossRef]
- Fuxe, K.; Marcellino, D.; Guidolin, D.; Woods, A.; Agnati, L.F. Dopamine Receptor Oligomerization. In The Dopamine Receptors; Neve, K.A., Ed.; Humana Press: Totowa, NJ, USA; Springer: Berlin/Heidelberg, Germany, 2010; pp. 255–280. [Google Scholar]
- Pelassa, S.; Guidolin, D.; Venturini, A.; Averna, M.; Frumento, G.; Campanini, L.; Bernardi, R.; Cortelli, P.; Buonaura, G.C.; Maura, G.; et al. A2A-D2 heteromers on striatal astrocytes: Biochemical and biophysical evidence. Int. J. Mol. Sci. 2019, 20, 2457. [Google Scholar] [CrossRef] [PubMed]
- Torvinen, M.; Marcellino, D.; Canals, M.; Agnati, L.F.; Lluis, C.; Franco, R.; Fuxe, K. Adenosine a2a receptor and dopamine d3 receptor interactions: Evidence of functional a2a/d3 heteromeric complexes. Mol. Pharmacol. 2005, 67, 400–407. [Google Scholar] [PubMed]
- Fuxe, K.; Borroto-Escuela, D.O. Receptor-Receptor Interactions in the Central Nervous System; Humana Press: New York, NY, USA, 2018; Volume 140, p. 346. [Google Scholar]
- Liu, X.Y.; Chu, X.P.; Mao, L.M.; Wang, M.; Lan, H.X.; Li, M.H.; Zhang, G.C.; Parelkar, N.K.; Fibuch, E.E.; Haines, M.; et al. Modulation of D2R–NR2B interactions in response to cocaine. Neuron 2006, 52, 897–909. [Google Scholar] [PubMed]
- Koschatzky, S.; Tschammer, N.; Gmeiner, P. Cross-receptor interactions between dopamine D2L and neurotensin NTS1 receptors modulate binding affinities of dopaminergics. ACS Chem. Neurosci. 2011, 2, 308–316. [Google Scholar]
- Plach, M.; Schäfer, T.; Borroto-Escuela, D.O.; Weickert, D.; Gmeiner, P.; Fuxe, K.; Friedland, K. Differential allosteric modulation within dopamine D2R-neurotensin NTS1R and D2R-serotonin 5-HT2AR receptor complexes gives bias to intracellular calcium signaling. Sci. Rep. 2019, 9, 16312. [Google Scholar]
- Przybyla, J.A.; Watts, V.J. Ligand-induced regulation and localization of cannabinoid CB1 and dopamine D2L receptor heterodimers. J. Pharmacol. Exp. Ther. 2010, 332, 710–719. [Google Scholar] [CrossRef]
- Kolasa, M.; Solich, J.; Faron-Górecka, A.; Żurawek, D.; Pabian, P.; Łukasiewicz, S.; Kuśmider, M.; Szafran-Pilch, K.; Szlachta, M.; Dziedzicka-Wasylewska, M. Paroxetine and Low-dose Risperidone Induce Serotonin 5-HT1A and Dopamine D2 Receptor Heteromerization in the Mouse Prefrontal Cortex. Neuroscience 2018, 377, 184–196. [Google Scholar]
- Borroto-Escuela, D.O.; Romero-Fernandez, W.; Tarakanov, A.O.; Marcellino, D.; Ciruela, F.; Agnati, L.F.; Fuxe, K. Dopamine D2 and 5-hydroxytryptamine 5-HT2A receptors assemble into functionally interacting heteromers. Biochem. Biophys. Res. Commun. 2010, 401, 605–610. [Google Scholar] [CrossRef]
- Romero-Fernandez, W.; Borroto-Escuela, D.O.; Agnati, L.F.; Fuxe, K. Evidence for the existence of dopamine D2-oxytocin receptor heteromers in the ventral and dorsal striatum with facilitatory receptor–receptor interactions. Mol. Psychiatry 2013, 18, 849–850. [Google Scholar] [CrossRef]
- Amato, S.; Averna, M.; Guidolin, D.; Ceccoli, C.; Gatta, E.; Candiani, S.; Pedrazzi, M.; Capraro, M.; Maura, G.; Agnati, L.F.; et al. Heteromerization of Dopamine D2 and Oxytocin Receptor in Adult Striatal Astrocytes. Int. J. Mol. Sci. 2023, 24, 4677. [Google Scholar]
- Kern, A.; Albarran-Zeckler, R.; Walsh, H.E.; Smith, R.G. Apo-grelin receptor forms heteromers with DRD2 in hypothalamic neurons and is essential for anorexigenic effects of DRD2 agonism. Neuron 2012, 73, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Ferré, S.; Becher, A.M.; Bonaventura, J.; Quiroz, C.; Sanchez-Soto, M.; Casadò-Anguera, V.; Cai, N.-S.; Moreno, E.; Boateng, C.A.; Keck, T.M.; et al. Functional and pharmacological role of the dopamine D4 receptor and its polymorphic variants. Front. Endocrinol. 2022, 13, 1014678. [Google Scholar]
- Rebois, R.V.; Maki, K.; Meeks, J.A.; Fishman, P.H.; Hébert, T.E.; Northup, J.K. D2-like dopamine and β-adrenergic receptors form a signaling complex that integrates Gs- and Gi-mediated regulation of adenylyl cyclase. Cell. Signal. 2012, 24, 2051–2060. [Google Scholar]
- Gago, B.; Fuxe, K.; Agnati, L.; Penafiel, A.; De La Calle, A.; Rivera, A. Dopamine D(4) receptor activation decreases the expression of mu-opioid receptors in the rat striatum. J. Comp. Neurol. 2007, 502, 358–366. [Google Scholar] [PubMed]
- Petkova-Kirova, P.; Giovannini, M.G.; Kalfin, R.; Rakovska, A. Modulation of acetylcholine release by cholecystokinin in striatum: Receptor specificity; role of dopaminergic neuronal activity. Brain Res. Bull. 2012, 89, 177–184. [Google Scholar]
- Pittolo, S.; Yokoyama, S.; Willoughby, D.D.; Taylor, C.R.; Reitman, M.E.; Tse, V.; Wu, Z.; Etchenique, R.; Li, Y.; Poskanzer, K.E. Dopamine activates astrocytes in prefrontal cortex via α1-adrenergic receptors. Cell Rep. 2022, 40, 111426. [Google Scholar]
- Pinton, L.; Borroto-Escuela, D.O.; Narváez, M.; Oflijan, J.; Agnati, L.F.; Fuxe, K. Evidence for the existence of dopamine D2R and Sigma 1 allosteric receptor–receptor interaction in the rat brain: Role in brain plasticity and cocaine action. Springerplus 2015, 4, P37. [Google Scholar] [CrossRef]
- Rashid, A.J.; So, C.H.; Kong, M.M.C.; Furtak, T.; El-Ghundi, M.; Cheng, R.; O’Dowd, B.F.; George, S.R. D1-D2 dopamine receptor heterooligomers with unique pharmacology are coupled to rapid activation of Gq/11 in the striatum. Proc. Natl. Acad. Sci. USA 2007, 104, 654–659. [Google Scholar] [CrossRef]
- Marcellino, D.; Ferré, S.; Casado, V.; Cortés, A.; Le Foll, B.; Mazzola, C.; Drago, F.; Saur, O.; Stark, H.; Soriano, A.; et al. Identification of dopamine D1-D3 receptor heteromers: Indications for a role of synergistic D1-D3 receptor interactions in the striatum. J. Biol. Chem. 2008, 283, 26016–26025. [Google Scholar]
- Ginés, S.; Hillion, J.; Torvinen, M.; Le Crom, S.; Casadó, V.; Canela, E.I.; Rondin, S.; Lew, J.Y.; Watson, S.; Zoli, M.; et al. Dopamine D1 and adenosine A1 receptors form functionally interacting heteromeric complexes. Proc. Natl. Acad. Sci. USA 2000, 97, 8606–8611. [Google Scholar] [CrossRef]
- Franco, R.; Lluis, C.; Canela, E.I.; Mallol, J.; Agnati, L.; Casadó, V.; Ciruela, F.; Ferré, S.; Fuxe, K. Receptor-receptor interactions involving adenosine A1 or dopamine D1 receptors and accessory proteins. J. Neural Transm. 2007, 114, 93–104. [Google Scholar] [PubMed]
- Lee, F.J.; Xue, S.; Pei, L.; Vukusic, B.; Chéry, N.; Wang, Y.; Wang, Y.T.; Niznik, H.B.; Yu, X.-M.; Liu, F. Dual regulation of NMDA receptor functions by direct protein-protein interactions with the dopamine D1 receptor. Cell 2002, 111, 219–230. [Google Scholar] [PubMed]
- Mitrano, D.A.; Pare, J.F.; Smith, Y.; Weinshenker, D. D1-dopamine and a1-adrenergic receptors co-localize in dendrites of the rat prefrontal cortex. Neuroscience 2014, 258, 90–100. [Google Scholar]
- Guidolin, D.; Marcoli, M.; Tortorella, C.; Maura, G.; Agnati, L.F. Adenosine A2A-Dopamine D2 Receptor-Receptor Interaction in Neurons and Astrocytes: Evidence and Perspectives. Prog. Mol. Biol. Transl. Sci. 2020, 169, 247–277. [Google Scholar] [PubMed]
- Ciruela, F.; Burgueno, J.; Casado, V.; Canals, M.; Marcellino, D.; Goldberg, S.R.; Bader, M.; Fuxe, K.; Agnati, L.F.; Lluis, C.; et al. Combining mass spectrometry and pull-down techniques for the study of receptor heteromerization. Direct epitope-epitope electro- static interactions between adenosine A2A and dopamine D2 receptors. Anal. Chem. 2004, 76, 5354–5363. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.S.; Ciruela, F.; Fuxe, K.; Agnati, L.F.; Lluis, C.; Franco, R.; Ferré, S. Role of electrostatic interaction in receptor–receptor heteromerization. J. Mol. Neurosci. 2005, 26, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.O.; Rodriguez, D.; Romero-Fernandez, W.; Kapla, J.; Jaiteh, M.; Ranganathan, A.; Lazarova, T.; Fuxe, K.; Carlsson, J. Mapping the interface of a GPCR dimer: A structural model of the A2A adenosine and D2 dopamine receptor heteromer. Front. Pharmacol. 2018, 9, 829. [Google Scholar]
- Ferré, S.; von Euler, G.; Johansson, B.; Fredholm, B.B.; Fuxe, K. Stimulation of high-affinity adenosine A2 receptors decreases the affinity of dopamine D2 receptors in rat striatal membranes. Proc. Natl. Acad. Sci. USA 1991, 88, 7238–7241. [Google Scholar] [CrossRef]
- Fuxe, K.; Ferré, S.; Zoli, M.; Agnati, L.F. Integrated events in central dopamine transmission as analyzed at multiple levels. Evidence for intramembrane adenosine A2A/dopamine D2 and adenosine A1/dopamine D1 receptor interactions in the basal ganglia. Brain Res. Rev. 1998, 26, 258–273. [Google Scholar] [CrossRef]
- Diaz-Cabiale, Z.; Hurd, Y.; Guidolin, D.; Finnman, U.B.; Zoli, M.; Agnati, L.F.; Vanderhaeghen, J.J.; Fuxe, K.; Ferré, S. Adenosine A2A agonist CGS 21680 decreases the affinity of dopamine D2 receptors for dopamine in human striatum. Neuroreport 2001, 12, 1831–1834. [Google Scholar] [CrossRef]
- Cervetto, C.; Venturini, A.; Passalacqua, M.; Guidolin, D.; Genedani, S.; Fuxe, K.; Borroto- Escuela, D.O.; Cortelli, P.; Woods, A.; Maura, G.; et al. A2A-D2 receptor- receptor interaction modulates gliotransmitter release from striatal astrocyte processes. J. Neurochem. 2016, 140, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Borroto-Escuela, D.; Romero-Fernandez, W.; Tarakanov, A.; Ciruela, F.; Agnati, L.; Fuxe, K. On the existence of a possible A2A-D2-beta- arrestin2 complex: A2A agonist modulation of D2 agonist-induced beta-arrestin2 recruitment. J. Mol. Biol. 2011, 406, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Agnati, L.F.; Ferré, S.; Genedani, S.; Leo, G.; Guidolin, D.; Filaferro, M.; Carriba, P.; Casado, V.; Lluis, C.; Franco, R.; et al. Allosteric modulation of dopamine D2 receptors by homocysteine. J. Proteome Res. 2006, 5, 3077–3083. [Google Scholar] [CrossRef] [PubMed]
- Cervetto, C.; Venturini, A.; Guidolin, D.; Maura, G.; Passalacqua, M.; Tacchetti, C.; Cortelli, P.; Genedani, S.; Candiani, S.; Ramoino, P.; et al. Homocysteine and A2A-D2 receptor-receptor interaction at striatal astrocyte processes. J. Mol. Neurosci. 2018, 65, 456–466. [Google Scholar] [PubMed]
- Borgkvist, A.; Marcellino, D.; Fuxe, K.; Greengard, P.; Fisone, G. Regulation of DARPP-32 phosphorylation by Delta(9)-tetrahydrocannabinol. Neuropharmacology 2008, 54, 31–35. [Google Scholar] [CrossRef]
- Łukasiewicz, S.; Błasiak, E.; Szafran-Pilch, K.; Dziedzicka-Wasylewska, M. Dopamine D2 and serotonin 5-HT1A receptor interaction in the context of the effects of antipsychotics—In Vitro studies. J Neurochem. 2016, 137, 549–560. [Google Scholar] [CrossRef]
- Fuxe, K.; Marcellino, D.; Rivera, A.; Diaz-cabiale, Z.; Filip, M.; Gago, B.; Roberts, D.C.S.; Lange, U.; Genedani, S.; Ferraro, L.; et al. Receptor-receptor interactions within receptor mosaics. Impact on neuropharmacology. Brain Res. Rev. 2008, 58, 415–452. [Google Scholar]
- Ferré, S.; Sarasola, L.I.; Quiroz, C.; Ciruela, F. Presynaptic adenosine receptor heteromers as key modulators of glutamatergic and dopaminergic neurotransmission in the striatum. Neuropharmacology 2023, 223, 109329. [Google Scholar]
- Liu, F.; Wan, Q.; Pristupa, Z.B.; Yu, X.M.; Wang, Y.T.; Niznik, H.B. Direct protein-protein coupling enables cross-talk between dopamine D5 and gamma-aminobutyric acid A receptors. Nature 2000, 403, 274–280. [Google Scholar] [CrossRef]
- McNeill, J.; Rudyk, C.; Hildebrand, M.E.; Salmaso, N. Ion channels and electrophysiological properties of astrocytes: Implications for emergent stimulation technologies. Front. Cell. Neurosci. 2021, 15, 644126. [Google Scholar]
- Fitzner, D.; Bader, J.M.; Penkert, H.; Bergner, C.G.; Su, M.; Weil, M.-T.; Surma, M.A.; Mann, M.; Klose, C.; Simons, M. Cell-type- and brain-region-resolved mouse brain lipidome. Cell Rep. 2020, 32, 108132. [Google Scholar] [PubMed]
- Lee, J.; Patel, D.S.; Ståhle, J.; Park, S.-J.; Kern, N.R.; Kim, S.; Lee, J.; Cheng, X.; Valvano, M.A.; Holst, O.; et al. CHARMM-GUI Membrane Builder for Complex Biological Membrane Simulations with glycolipids and lipoglycans. Chem. Comput. 2019, 15, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Kuriata, A.; Gierut, A.M.; Oleniecki, T.; Ciemny, M.P.; Kolinski, A.; Kurcinski, M.; Kmiecik, S. CABS-Flex 2.0: A Web Server for fast simulations of flexibility of protein structures. Nucleic Acids Res. 2018, 46, W338–W343. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Carlsson, J.; Ambrogini, P.; Narvàez, M.; Wydra, K.; Tarakanov, A.D.; Li, X.; Millòn, C.; Ferraro, L.; Cuppini, R.; et al. Understanding the role of GPCR heteroreceptor complexes in modulating the brain networks in health and disease. Front. Cell. Neurosci. 2017, 11, 37. [Google Scholar] [PubMed]
- Guidolin, D.; Tortorella, C.; Marcoli, M.; Cervetto, C.; Maura, G.; Agnati, L.F. Receptor–Receptor Interactions and Glial Cell Functions with a Special Focus on G Protein-Coupled Receptors. Int. J. Mol. Sci. 2021, 22, 8656. [Google Scholar] [CrossRef] [PubMed]
- Perreault, M.; Hasbi, A.; O’Dowd, B.F.; George, S.R. heteromeric dopamine receptor signaling complexes: Emerging neurobiology and disease relevance. Neuropsychopharmacology 2014, 39, 156–168. [Google Scholar]
- Lindvall, O.; Björklund, A. The organization of the ascending catecholamine neuron systems in the rat brain as revealed by the glyoxylic acid fluorescence method. Acta Physiol. Scand. 1974, 412, 1–48. [Google Scholar]
- Gerfen, C.R. The neostriatal mosaic: Multiple levels of compartmental organization. Trends Neurosci. 1992, 15, 133–139. [Google Scholar]
- Liu, Y.-J.; Chen, J.; Li, X.; Zhou, X.; Hu, Y.-M.; Chu, S.-F.; Peng, Y.; Chen, N.-H. Research progress on adenosine in central nervous system diseases. CNS Neurosci. Ther. 2019, 25, 899–910. [Google Scholar]
- Ferré, S.; Popoli, P.; Gimenez-Llort, L.; Finnman, U.B.; Martinez, E.; Scotti de Carolis, A.; Fuxe, K. Postsynaptic antagonistic interaction between adenosine A1 and dopamine D1 receptors. Neuroreport 1994, 6, 73–76. [Google Scholar] [CrossRef]
- Emmi, A.; Antonini, A.; Sandre, M.; Baldo, A.; Contran, M.; Macchi, V.; Guidolin, D.; Porzionato, A.; De Caro, R. Topography and distribution of adenosine A2A and dopamine D2 receptors in the human subthalamic nucleus. Front. Neurosci. 2022, 16, 945574. [Google Scholar] [CrossRef] [PubMed]
- Cotzias, G.C. Levodopa in the treatment of Parkinsonism. JAMA 1971, 218, 1903–1908. [Google Scholar] [CrossRef] [PubMed]
- Marsden, C.D. Problems with long-term levodopa therapy for Parkinson’s disease. Clin. Neuropharmacol. 1994, 17, S32–S544. [Google Scholar] [CrossRef] [PubMed]
- Fuxe, K.; Guidolin, D.; Agnati, L.F.; Borroto-Escuela, D.O. Dopamine heteroreceptor complexes as therapeutic targets in Parkinson’s disease. Exp. Opin. Ther. Targets 2015, 19, 377–398. [Google Scholar]
- Chen, J.F.; Cunha, R.A. The belated US FDA approval of the adenosine A2A receptor antagonist istradefylline for treatment of Parkinson’s disease. Purinergic Signal. 2020, 16, 167–174. [Google Scholar]
- Marcellino, D.; Carriba, P.; Filip, M.; Borgkvist, A.; Frankowska, M.; Bellido, I.; Tanganelli, S.; Muller, C.E.; Roberts, D.C.; Fisone, G.; et al. Antagonistic cannabinoid CB1/dopamine D2 receptor interactions in striatal CB1/D2 heteromers. A combined neurochemical and behavioural analysis. Neuropharmacology 2008, 54, 815–828. [Google Scholar] [CrossRef]
- Gilgun-Sherki, Y.; Melamed, E.; Mechoulam, R.; Offen, D. The CB1 cannabinoid receptor agonist, HU-210, reduces levodopa-induced rotations in 6-hydroxydopamine-lesioned rats. Pharmacol. Toxicol. 2003, 93, 66–70. [Google Scholar] [CrossRef]
- Haber, S.N. Neuroanatomy of reward: A view from the ventral striatum. In Neurobiology of Sensation and Reward; Gottfried, J.A., Ed.; CRC Press/Taylor and Francis: Boca Raton, FL, USA, 2011. [Google Scholar]
- Fuxe, K.; Dahlstrom, A.; Jonsson, G.; Marcellino, D.; Guescini, M.; Dam, M.; Manger, P.; Agnati, L. The discovery of central monoamine neurons gave volume transmission to the wired brain. Prog. Neurobiol. 2010, 90, 82–100. [Google Scholar]
- Borroto-Escuela, D.O.; Ferraro, L.; Narvaez, M.; Tanganelli, S.; Beggiato, S.; Liu, F.; Rvera, A.; Fuxe, K. Multiple adenosine-dopamine (A2A-D2 like) heteroreceptor complexes in the brain and their role in schizophrenia. Cells 2020, 9, 1077. [Google Scholar] [CrossRef]
- Borroto-Escuela, D.O.; Pintsuk, J.; Schäfer, T.; Friedland, K.; Ferraro, L.; Tanganelli, S.; Liu, F.; Fuxe, K. Multiple D2 heteroreceptor complexes: New targets for treatment of schizophrenia. Ther. Adv. Psychopharmacol. 2016, 6, 77–94. [Google Scholar] [CrossRef]
- Edwards, S.; Whisler, K.N.; Fuller, D.C.; Orsulak, P.J.; Self, D.W. Addiction-related alterations in D1 and D2 dopamine receptor behavioral responses following chronic cocaine self-administration. Neuropsychopharmacology 2007, 32, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Briand, L.A.; Flagel, S.B.; Garcia-Fuster, M.J.; Watson, S.J.; Akil, H.; Sarter, M.; Robinson, T.E. Persistent alterations in cognitive function and prefrontal dopamine D2 receptors following extended, but not limited, access to self-administered cocaine. Neuropsychopharmacology 2008, 33, 2969–2980. [Google Scholar] [PubMed]
- Wydra, K.; Suder, A.; Borroto-Escuela, D.O.; Filip, M.; Fuxe, K. On the role of A(2)A and D(2) receptors in control of cocaine and food-seeking behaviors in rats. Psychopharmacology 2015, 232, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Romieu, P.; Phan, V.L.; Martin-Fardon, R.; Maurice, T. Involvement of the sigma1 receptor in cocaine-induced conditioned place preference: Possible dependence on dopamine uptake blockade. Neuropsychopharmacology 2002, 26, 444–455. [Google Scholar] [CrossRef]
- Canales, J.J. Stimulant-induced adaptations in neostriatal matrix and striosome systems: Transiting from instrumental responding to habitual behavior in drug addiction. Neurobiol. Learn. Mem. 2005, 83, 93–103. [Google Scholar] [CrossRef]
- Seeman, P. Targeting the dopamine D2 receptor in schizophrenia. Expert Opin. Ther. Targets 2006, 10, 515–531. [Google Scholar] [CrossRef]
- Carlsson, A. The current status of the dopamine hypothesis of schizophrenia. Neuropsychopharmacology 1988, 1, 179–186. [Google Scholar] [CrossRef]
- Andersen, M.B.; Fuxe, K.; Werge, T.; Gerlach, J. The adenosine A2A receptor agonist CGS 21680 exhibits antipsychotic-like activity in Cebus apella monkeys. Behav. Pharmacol. 2022, 13, 639–644. [Google Scholar] [CrossRef]
- Caldwell, H.K.; Stephens, S.L.; Young, W.S. III. Oxytocin as a natural antipsychotic: A study using oxytocin knockout mice. Mol. Psychiatry 2009, 14, 190–196. [Google Scholar] [CrossRef]
- Feifel, D. Oxytocin as a potential therapeutic target for schizophrenia and other neuropsychiatric conditions. Neuropsychopharmacology 2012, 37, 304–305. [Google Scholar] [CrossRef]
- Hökfelt, T.; Fuxe, K.; Johansson, O.; Ljungdahl, Å. Pharmaco-histochemical evidence of the existence of dopamine nerve terminals in the limbic cortex. Eur. J. Pharmacol. 1974, 25, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Berger, B.; Gaspar, P.; Verney, C. Dopaminergic innervation of the cerebral cortex: Unexpected differences between rodents and primates. Trends Neurosci. 1991, 14, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Malenka, E.J.; Nestler, S.E.; Hyman, R.C. Molecular Neuropharmacology: A Foundation for Clinical Neuroscience, 2nd ed.; McGraw-Hill Medical: New York, NY, USA, 2009; Chapter 13; p. 318. [Google Scholar]
- Casadó-Anguera, V.; Moreno, E.; Sánchez-Soto, M.; Cai, N.S.; Bonaventura, J.; Homar-Ruano, P.; Rubinstein, M.; Cortés, A.; Canela, E.I.; Ferré, S.; et al. Heteromerization between a2A adrenoceptors and different polymorphic variants of the dopamine D4 receptor determines pharmacological and functional differences. implications for impulsive-control disorders. Pharmacol. Res. 2021, 170, 105745. [Google Scholar] [CrossRef] [PubMed]
- LaHoste, G.J.; Swanson, J.M.; Wigal, S.B.; Glabe, C.; Wigal, T.; King, N.; Kennedy, J. Dopamine D4 receptor gene polymorphism is associated with attention deficit hyperactivity disorder. Mol. Psychiatry 1996, 1, 121–124. [Google Scholar]
- Sancho, L.; Contreras, M.; Allen, N.J. Glia as sculptors of synaptic plasticity. Neurosci. Res. 2021, 167, 17–29. [Google Scholar]
- Allen, N.J.; Barres, B.A. Signaling between glia and neurons: Focus on synaptic plasticity. Curr. Opin. Neurobiol. 2005, 15, 542–548. [Google Scholar] [CrossRef]
- Fellin, T.; Carmignoto, G. Neurone-to-astrocyte signalling in the brain represents a distinct multifunctional unit. J. Physiol. 2004, 559, 3–15. [Google Scholar]
- Cervetto, C.; Maura, G.; Guidolin, D.; Amato, S.; Ceccoli, C.; Agnati, L.F.; Marcoli, M. Striatal astrocytic A2A-D2 receptor-receptor interactions and their role in neuropsychiatric disorders. Neuropharmacology 2023, 237, 109636. [Google Scholar]
- Ren, C.; He, K.J.; Hu, H.; Zhang, J.B.; Dong, L.G.; Li, D.; Chen, J.; Mao, C.J.; Wang, F.; Liu, C.F. Induction of parkinsonian-like changes via targeted downregulation of astrocytic glutamate transporter GLT-1 in the striatum. J. Park. Dis. 2022, 12, 295–314. [Google Scholar]
- Martín, R.; Bajo-Grañeras, R.; Moratalla, R.; Perea, G.; Araque, A. Circuit-specific signaling in astrocyte-neuron networks in basal ganglia pathways. Science 2015, 349, 730–734. [Google Scholar] [CrossRef]
- Huang, G.; Dragan, M.; Freeman, D.; Wilson, J.X. Activation of catechol-O-methyltransferase in astrocytes stimulates homocysteine synthesis and export to neurons. Glia 2005, 51, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.; Shen, H.Y.; Augusto, E.; Wang, Y.; Wei, C.J.; Wang, Y.T.; Agostinho, P.; Boison, D.; Cunha, R.A.; Chen, J.F. Deletion of adenosine A2A receptors from astrocytes disrupts glutamate homeostasis leading to psychomotor and cognitive impairment: Relevance to schizophrenia. Biol. Psychiatr. 2015, 78, 763–774. [Google Scholar]
- Kruyer, A.; Kalivas, P.W.; Scofield, M.D. Astrocyte regulation of synaptic signaling in psychiatric disorders. Neuropsychopharmacology 2023, 48, 21–36. [Google Scholar] [PubMed]
- Berretta, S. Extracellular matrix abnormalities in schizophrenia. Neuropharmacology 2012, 62, 1584–1597. [Google Scholar] [PubMed]
- Wardas, J. Potential role of adenosine A2A receptors in the treatment of schizophrenia. Front. Biosci. 2008, 13, 4071–4096. [Google Scholar] [CrossRef]
- Valle-Léon, M.; Casajuana-Martin, N.; del Torrent, C.L.; Argerich, J.; Gòmez-Acero, L.; Sahlholm, K.; Ferré, S.; Pardo, L.; Ciruela, F. Unique effect of clozapine on adenosine A2A-dopamine D2 receptor heteromerization. Biomed. Pharmacother. 2023, 160, 114327. [Google Scholar]
- Kruyer, A.; Kalivas, P.W. Astrocytes as cellular mediators of cue reactivity in addiction. Curr. Opin. Pharmacol. 2021, 56, 1–6. [Google Scholar] [CrossRef]
- Alexander, G.E.; Crutcher, M.D. Functional architecture of basal ganglia circuits: Neural substrates of parallel processing. Trends Neurosci. 1990, 13, 266–271. [Google Scholar]
- Daniels, D.J.; Lenard, N.R.; Etienne, C.L.; Law, P.-Y.; Roerig, S.C.; Portoghese, P.S. Opioid-induced tolerance and dependence in mice is modulated by the distance between pharmacophores in a bivalent ligand series. Proc. Natl. Acad. Sci. USA 2005, 102, 19208–19213. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).