
Cannabidiol and Minor Phytocannabinoids: A Preliminary Study to Assess Their Anti-Melanoma, Anti-Melanogenic, and Anti-Tyrosinase Properties




Abstract
1. Introduction
2. Results and Discussion
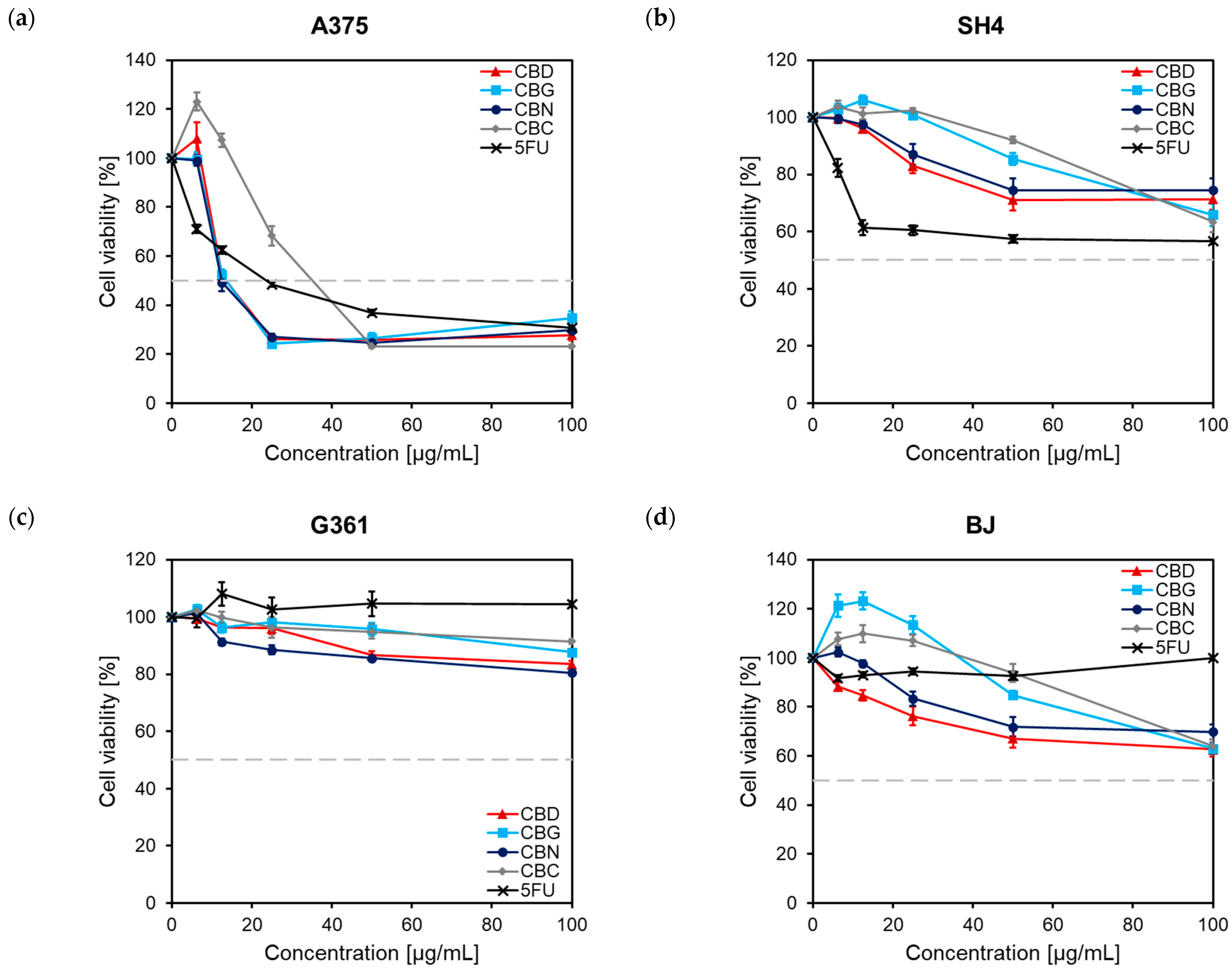
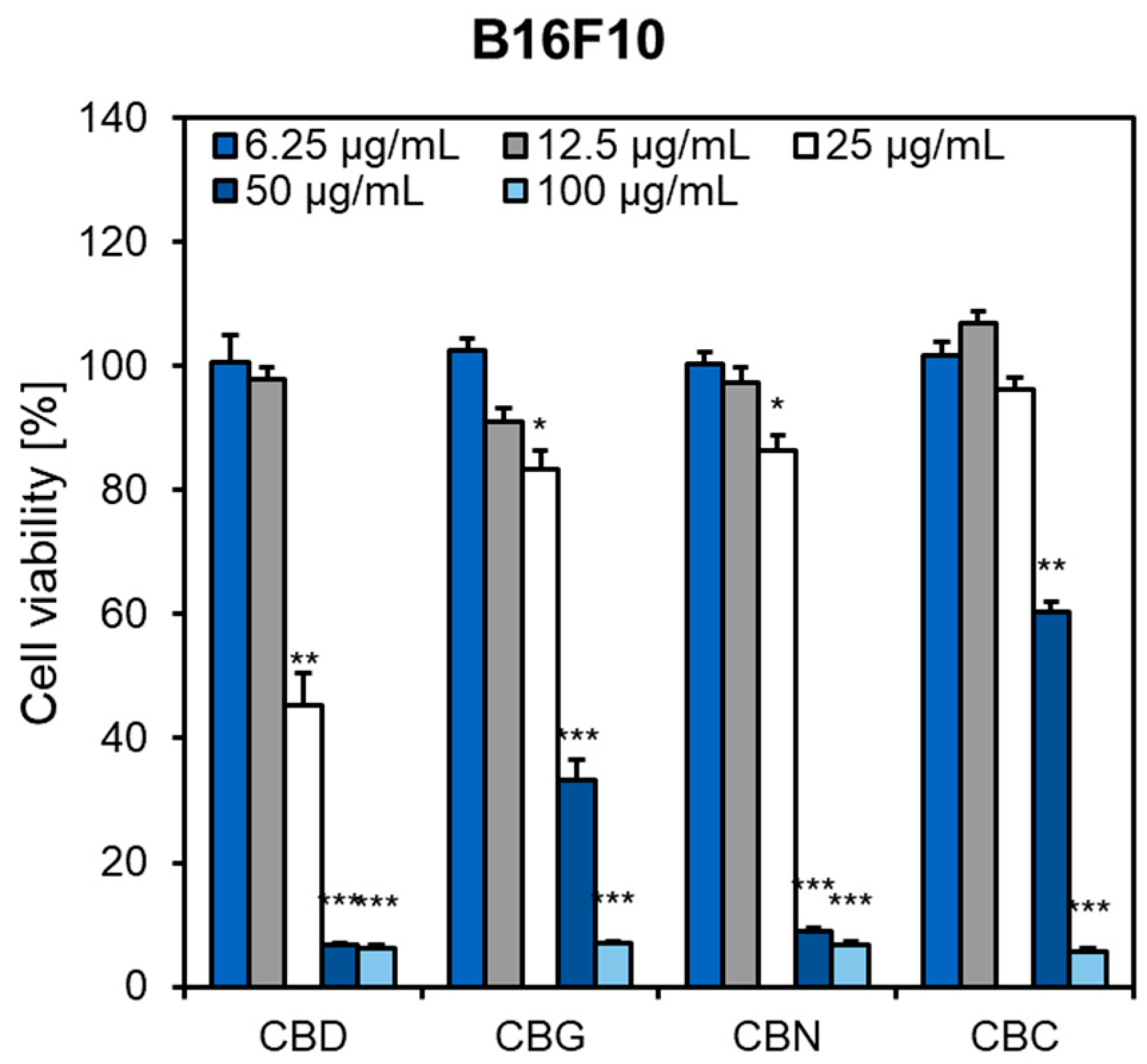
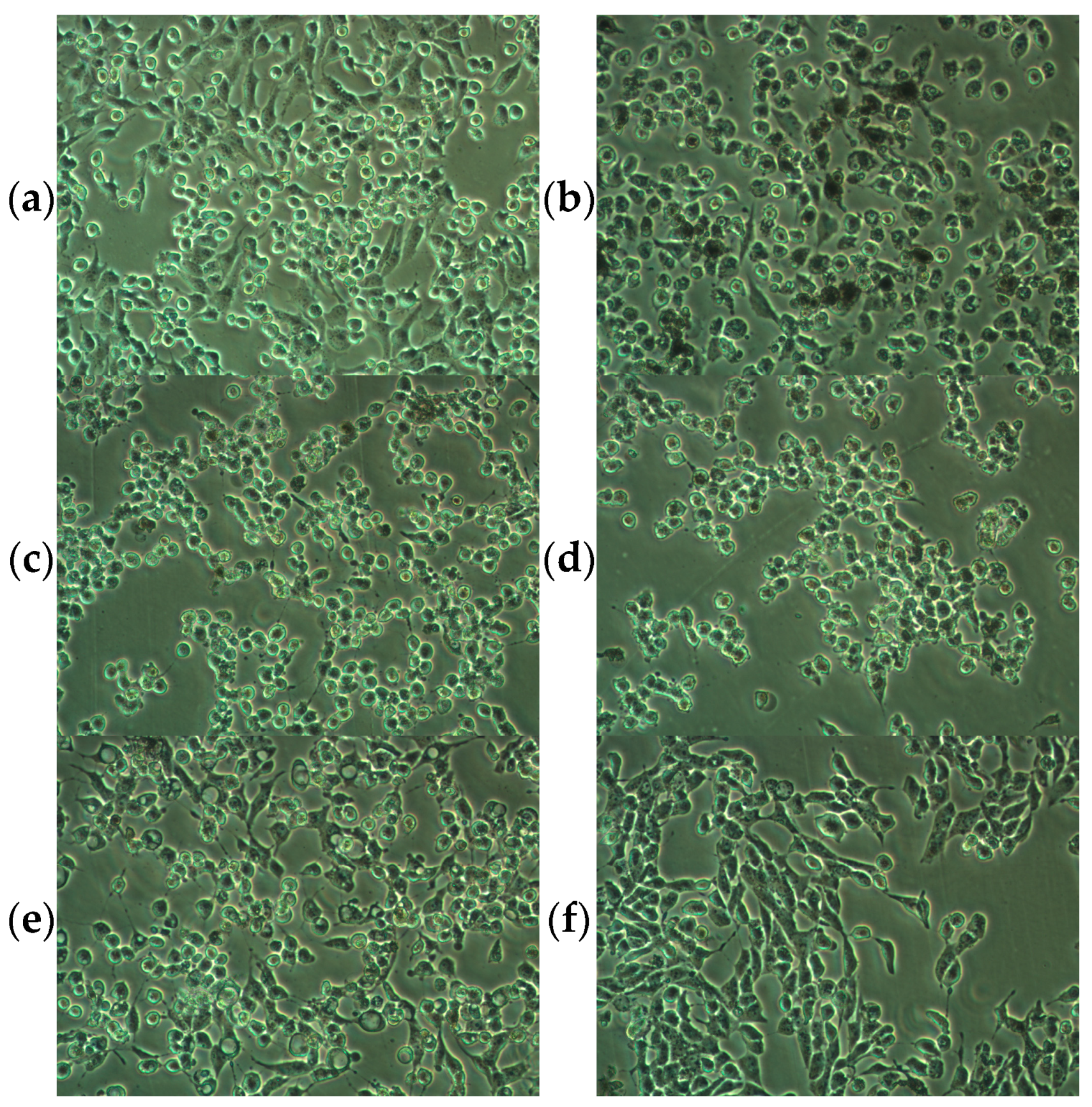
2.1. Influence of Phytocannabinoids on Malignant Melanoma Cell Viability
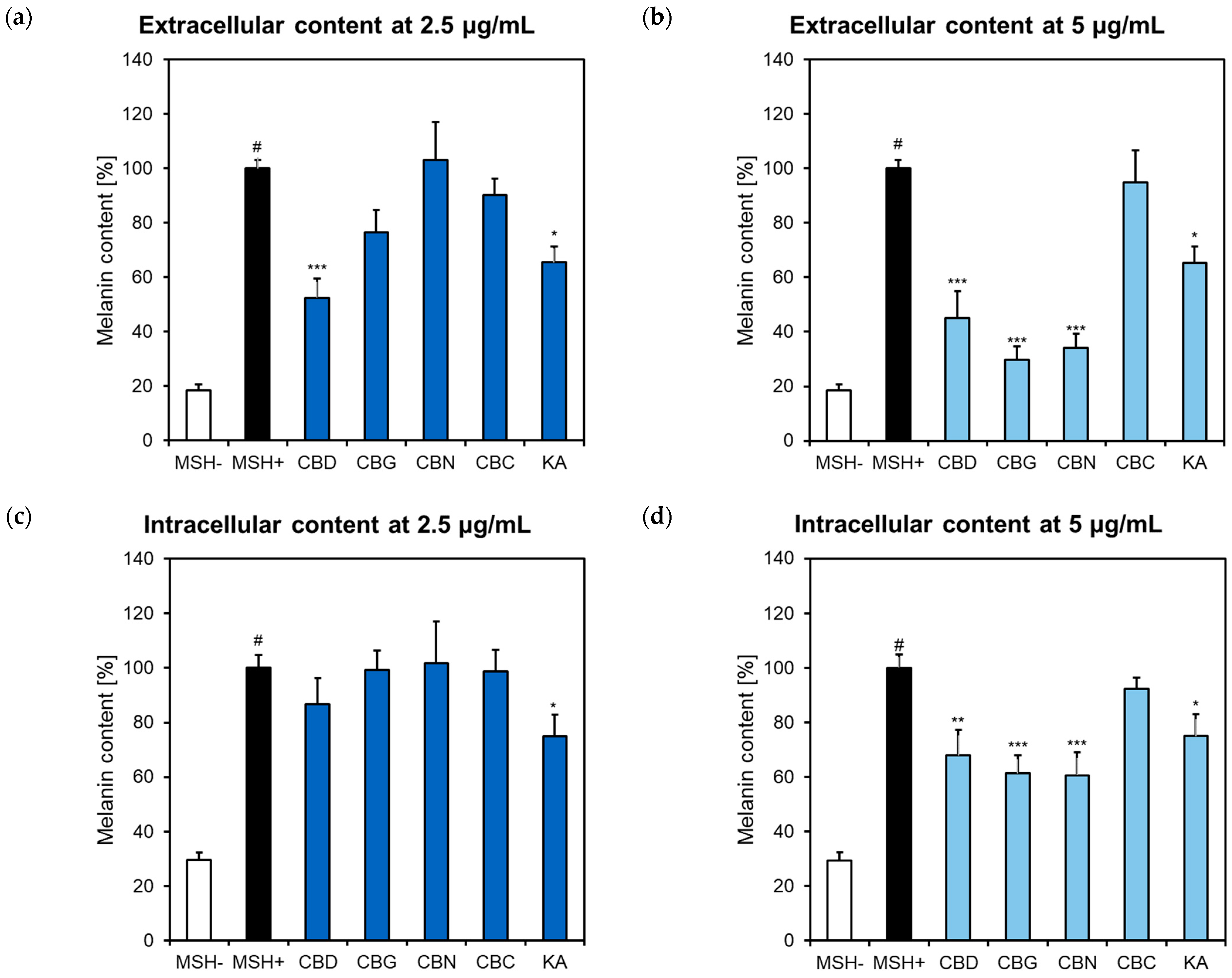
2.2. Influence of Phytocannabinoids on Melanin Synthesis
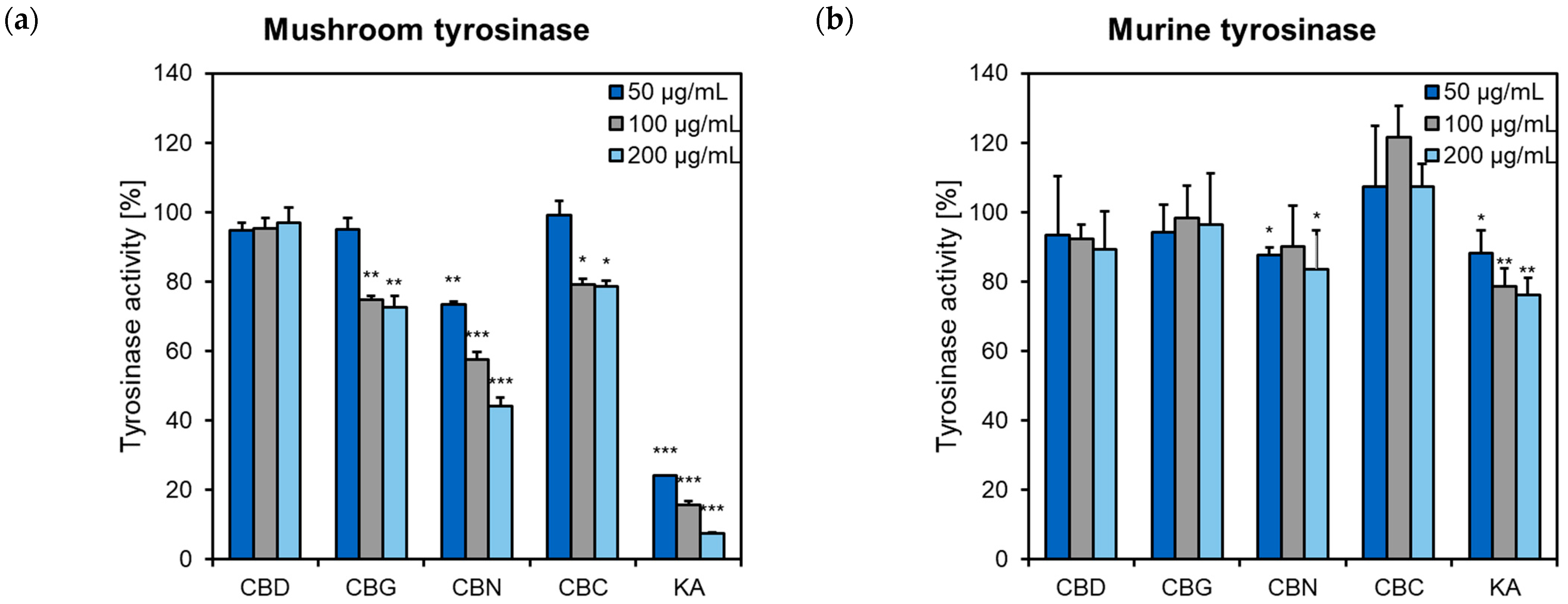
2.3. Influence of Phytocannabinoids on Tyrosinase Activity
3. Materials and Methods
3.1. Chemicals
3.2. Cell Lines
3.3. Cell Viability Assay (Neutral Red Uptake)
3.4. Melanin Assay
3.5. Tyrosinase Activity Assays
3.5.1. Mushroom Tyrosinase Activity Assay
3.5.2. Murine Tyrosinase Activity Assay
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baswan, S.M.; Klosner, A.E.; Glynn, K.; Rajgopal, A.; Malik, K.; Yim, S.; Stern, N. Therapeutic potential of cannabidiol (CBD) for skin health and disorders. Clin. Cosmet. Investig. Dermatol. 2020, 13, 927–942. [Google Scholar] [CrossRef] [PubMed]
- Shao, K.; Stewart, C.; Grant-Kels, J.M. Cannabis and the skin. Clin. Dermatol. 2021, 39, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Szepietowski, J.C.; Szepietowski, T.; Reich, A. Efficacy and tolerance of the cream containing structured physiological lipids with endocannabinoids in the treatment of uremic pruritus: A preliminary study. Acta Dermatovenerol. Croat. 2005, 13, 97–103. [Google Scholar]
- Visse, K.; Blome, C.; Phan, N.Q.; Augustin, M.; Ständer, S. Efficacy of body lotion containing N-palmitoylethanolamine in subjects with chronic pruritus due to dry skin: A dermatocosmetic study. Acta Derm. Venereol. 2017, 97, 639–641. [Google Scholar] [CrossRef] [PubMed]
- Avila, C.; Massick, S.; Kaffenberger, B.H.; Kwatra, S.G.; Bechtel, M. Cannabinoids for the treatment of chronic pruritus: A review. J. Am. Acad. Dermatol. 2020, 82, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.M.; Gomes, A.L.; Vilas Boas, I.; Marto, J.; Ribeiro, H.M. Cannabis-based products for the treatment of skin inflammatory diseases: A timely review. Pharmaceuticals 2022, 15, 210. [Google Scholar] [CrossRef]
- Wroński, A.; Jarocka-Karpowicz, I.; Stasiewicz, A.; Skrzydlewska, E. Phytocannabinoids in the pharmacotherapy of psoriasis. Molecules 2023, 28, 1192. [Google Scholar] [CrossRef] [PubMed]
- Friedman, A.J.; Momeni, K.; Kogan, M. Topical cannabinoids for the management of psoriasis vulgaris: Report of a case and review of the literature. J. Drugs Dermatol. 2020, 19, 795. [Google Scholar] [CrossRef]
- Ali, A.; Akhtar, N. The safety and efficacy of 3% Cannabis seeds extract cream for reduction of human cheek skin sebum and erythema content. Pak. J. Pharm. Sci. 2015, 28, 1389–1395. [Google Scholar]
- Jiang, Z.; Jin, S.; Fan, X.; Cao, K.; Liu, Y.; Wang, X.; Ma, Y.; Xiang, L. Cannabidiol inhibits inflammation induced by Cutibacterium acnes-derived extracellular vesicles via activation of CB2 receptor in keratinocytes. J. Inflamm. Res. 2022, 15, 4573–4583. [Google Scholar] [CrossRef]
- Eberlein, B.; Eicke, C.; Reinhardt, H.W.; Ring, J. Adjuvant treatment of atopic eczema: Assessment of an emollient containing N-palmitoylethanolamine (ATOPA study). J. Eur. Acad. Dermatol. Venereol. 2008, 22, 73–82. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, X.-M.; Guichard, A.; Tan, Y.-M.; Qian, C.-Y.; Yang, L.-J.; Humbert, P. N-palmitoylethanolamine and N-acetylethanolamine are effective in asteatotic eczema: Results of a randomized, double-blind, controlled study in 60 patients. Clin. Interv. Aging 2014, 9, 1163–1169. [Google Scholar] [CrossRef]
- Gupta, A.K.; Talukder, M. Cannabinoids for skin diseases and hair regrowth. J. Cosm. Dermatol. 2021, 20, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Tijani, A.O.; Thakur, D.; Mishra, D.; Frempong, D.; Chukwunyere, U.I.; Puri, A. Delivering therapeutic cannabinoids via skin: Current state and future perspectives. J. Contr. Rel. 2021, 334, 427–451. [Google Scholar] [CrossRef] [PubMed]
- Ramer, R.; Hinz, B. Cannabinoid Compounds as a pharmacotherapeutic option for the treatment of non-cancer skin diseases. Cells 2022, 11, 4102. [Google Scholar] [CrossRef] [PubMed]
- Sheriff, T.; Lin, M.J.; Dubin, D.; Khorasani, H. The potential role of cannabinoids in dermatology. J. Dermatol. Treat. 2020, 31, 839–845. [Google Scholar] [CrossRef]
- Martinelli, G.; Magnavacca, A.; Fumagalli, M.; Dell’Agli, M.; Piazza, S.; Sangiovanni, E. Cannabis sativa and skin health: Dissecting the role of phytocannabinoids. Plant Med. 2022, 88, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Pasquariello, N.; Battista, N.; Di Tommaso, M.; Rapino, C.; Fezza, F.; Zuccolo, M.; Jourdain, R.; Agrò, A.F.; Breton, L. Endocannabinoids stimulate human melanogenesis via type-1 cannabinoid receptor. J. Biol. Chem. 2012, 287, 15466–15478. [Google Scholar] [CrossRef]
- Merhavi-Shoham, E.; Itzhaki, O.; Markel, G.; Schachter, J.; Besser, M.J. Adoptive cell therapy for metastatic melanoma. Cancer J. 2017, 23, 48–53. [Google Scholar] [CrossRef]
- Ribas, A. Anti-CTLA4 antibody clinical trials in melanoma. Update Cancer Ther. 2007, 2, 133–139. [Google Scholar] [CrossRef]
- Davey, R.J.; van der Westhuizen, A.; Bowden, N.A. Metastatic melanoma treatment: Combining old and new therapies. Crit. Rev. Oncol. Hematol. 2016, 98, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Ramer, R.; Wendt, F.; Wittig, F.; Schäfer, M.; Boeckmann, L.; Emmert, S.; Hinz, B. Impact of Cannabinoid compounds on skin cancer. Cancers 2022, 14, 1769. [Google Scholar] [CrossRef] [PubMed]
- Schanknecht, E.; Bachari, A.; Nassar, N.; Piva, T.; Mantri, N. Phytochemical constituents and derivatives of Cannabis sativa; bridging the gap in melanoma treatment. Int. J. Mol. Sci. 2023, 24, 859. [Google Scholar] [CrossRef] [PubMed]
- Bachari, A.; Piva, T.J.; Salami, S.A.; Jamshidi, N.; Mantri, N. Roles of cannabinoids in melanoma: Evidence from in vivo studies. Int. J. Mol. Sci. 2020, 21, 6040. [Google Scholar] [CrossRef]
- Baek, S.-H.; Du Han, S.; Yook, C.N.; Kim, Y.C.; Kwak, J.S. Synthesis and antitumor activity of cannabigerol. Arch. Pharm. Res. 1996, 19, 228–230. [Google Scholar] [CrossRef]
- Orlando, G.; Recinella, L.; Chiavaroli, A.; Brunetti, L.; Leone, S.; Carradori, S.; Di Simone, S.; Ciferri, M.C.; Zengin, G.; Ak, G. Water extract from inflorescences of industrial hemp Futura 75 variety as a source of anti-inflammatory, anti-proliferative and antimycotic agents: Results from in silico, in vitro and ex vivo studies. Antioxidants 2020, 9, 437. [Google Scholar] [CrossRef]
- Corni, G.; Brighenti, V.; Pellati, F.; Morlock, G.E. Effect-directed analysis of bioactive compounds in Cannabis sativa L. by high-performance thin-layer chromatography. J. Chromatogr. A 2020, 1629, 461511. [Google Scholar] [CrossRef]
- di Giacomo, V.; Recinella, L.; Chiavaroli, A.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Politi, M.; Antolini, M.D.; Acquaviva, A. Metabolomic profile and antioxidant/anti-inflammatory effects of industrial hemp water extract in fibroblasts, keratinocytes and isolated mouse skin specimens. Antioxidants 2021, 10, 44. [Google Scholar] [CrossRef]
- Telek, A.; Bíró, T.; Bodó, E.; Tóth, B.I.; Borbíró, I.; Kunos, G.; Paus, R. Inhibition of human hair follicle growth by endo-and exocannabinoids. FASEB J. 2007, 21, 3534–3541. [Google Scholar] [CrossRef]
- Hwang, Y.S.; Kim, Y.-J.; Kim, M.O.; Kang, M.; Oh, S.W.; Nho, Y.H.; Park, S.-H.; Lee, J. Cannabidiol upregulates melanogenesis through CB1 dependent pathway by activating p38 MAPK and p42/44 MAPK. Chem. Biol. Int. 2017, 273, 107–114. [Google Scholar] [CrossRef]
- Goenka, S. Comparative study of Δ9-tetrahydrocannabinol and cannabidiol on melanogenesis in human epidermal melanocytes from different pigmentation phototypes: A pilot study. J. Xenobiot. 2022, 12, 131–144. [Google Scholar] [CrossRef]
- Luca, S.V.; Roehrer, S.; Kleigrewe, K.; Minceva, M. Approach for simultaneous cannabidiol isolation and pesticide removal from hemp extracts with liquid-liquid chromatography. Ind. Crops Prod. 2020, 155, 112726. [Google Scholar] [CrossRef]
- Luca, S.V.; Braumann, L.; Gerigk, M.; Frank, O.; Minceva, M. Separation of minor cannabinoids from hemp extract with trapping multiple dual mode liquid-liquid chromatography. J. Chromatogr. A 2021, 1658, 462608. [Google Scholar] [CrossRef]
- Marzęda, P.; Wróblewska-Łuczka, P.; Drozd, M.; Florek-Łuszczki, M.; Załuska-Ogryzek, K.; Łuszczki, J.J. Cannabidiol interacts antagonistically with cisplatin and additively with mitoxantrone in various melanoma cell lines—An isobolographic analysis. Int. J. Mol. Sci. 2022, 23, 675. [Google Scholar] [CrossRef] [PubMed]
- Vacek, J.; Vostalova, J.; Papouskova, B.; Skarupova, D.; Kos, M.; Kabelac, M.; Storch, J. Antioxidant function of phytocannabinoids: Molecular basis of their stability and cytoprotective properties under UV-irradiation. Free Radic. Biol. Med. 2021, 164, 258–270. [Google Scholar] [CrossRef]
- Burch, R.; Mortuza, A.; Blumenthal, E.; Mustafa, A. Effects of cannabidiol (CBD) on the inhibition of melanoma cells in vitro. J. Immunoass. Immunochem. 2021, 42, 285–291. [Google Scholar] [CrossRef]
- Choi, W.H.; Park, H.D.; Baek, S.H.; Chu, J.P.; Kang, M.H.; Mi, Y.J. Cannabidiol induces cytotoxicity and cell death via apoptotic pathway in cancer cell lines. Biomolec. Ther. 2008, 16, 87–94. [Google Scholar]
- Adinolfi, B.; Romanini, A.; Vanni, A.; Martinotti, E.; Chicca, A.; Fogli, S.; Nieri, P. Anticancer activity of anandamide in human cutaneous melanoma cells. Eur. J. Pharmacol. 2013, 718, 154–159. [Google Scholar] [CrossRef]
- Blázquez, C.; Carracedo, A.; Barrado, L.; Jose Real, P.; Luis Fernández-Luna, J.; Velasco, G.; Malumbres, M.; Guzmán, M.; Blázquez, C.; Carracedo, A. Cannabinoid receptors as novel targets for the treatment of melanoma. FASEB J. 2006, 20, 2633–2635. [Google Scholar] [CrossRef]
- Hamtiaux, L.; Masquelier, J.; Muccioli, G.G.; Bouzin, C.; Feron, O.; Gallez, B.; Lambert, D.M. The association of N-palmitoylethanolamine with the FAAH inhibitor URB597 impairs melanoma growth through a supra-additive action. BMC Cancer 2012, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Su, J.; Wang, R.; Hao, R.; Fu, C.; Chen, J.; Li, J.; Wang, X. Structural optimization of cannabidiol as multifunctional cosmetic raw materials. Antioxidants 2023, 12, 314. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [PubMed]
- Strzępek-Gomółka, M.; Gaweł-Bęben, K.; Angelis, A.; Antosiewicz, B.; Sakipova, Z.; Kozhanova, K.; Głowniak, K.; Kukula-Koch, W. Identification of mushroom and murine tyrosinase inhibitors from Achillea biebersteinii Afan. extract. Molecules 2021, 26, 964. [Google Scholar] [CrossRef] [PubMed]
- Widelski, J.; Gaweł-Bęben, K.; Czech, K.; Paluch, E.; Bortkiewicz, O.; Kozachok, S.; Mroczek, T.; Okińczyc, P. Extracts from European propolises as potent tyrosinase inhibitors. Molecules 2022, 28, 55. [Google Scholar] [CrossRef] [PubMed]
- Yoshimori, A.; Oyama, T.; Takahashi, S.; Abe, H.; Kamiya, T.; Abe, T.; Tanuma, S.I. Structure–activity relationships of the thujaplicins for inhibition of human tyrosinase. Bioorg. Med. Chem. 2014, 22, 6193–6200. [Google Scholar] [CrossRef]
- Liu, T.; Lu, Y.; Tonissen, K.; Di Trapani, G.; Tang, W.; Feng, Y. Application of traditional Chinese medicine as skin depigmentation agents. Heliyon 2022, 8, e12571. [Google Scholar] [CrossRef]
- BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 12 April 2023).
- Brand, S.; Winkler, M.; Götz, M.R.; Symrise, A.G. Synthesis and Purification of Cannabigerol and Its Application 2020. Patent WO2020249184A1, 17 December 2020. Available online: https://patents.google.com/patent/WO2020249184A1/en (accessed on 12 April 2023).
- Repetto, G.; Del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Uchida, R.; Ishikawa, S.; Tomoda, H. Inhibition of tyrosinase activity and melanine pigmentation by 2-hydroxytyrosol. Acta Pharm. Sin. B 2014, 4, 141–145. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | CBD | CBG | CBN | CBC | 5FU |
|---|---|---|---|---|---|
| IC50 [μg/mL] | |||||
| A375 | 12.02 ± 0.60 | 12.14 ± 2.34 | 25.13 ± 1.68 | 23.00 ± 2.60 | 29.96 ± 0.78 |
| SH4 | >100 | >100 | >100 | >100 | >100 |
| G361 | >100 | >100 | >100 | >100 | >100 |
| BJ | >100 | >100 | >100 | >100 | >100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaweł-Bęben, K.; Czech, K.; Luca, S.V. Cannabidiol and Minor Phytocannabinoids: A Preliminary Study to Assess Their Anti-Melanoma, Anti-Melanogenic, and Anti-Tyrosinase Properties. Pharmaceuticals 2023, 16, 648. https://doi.org/10.3390/ph16050648
Gaweł-Bęben K, Czech K, Luca SV. Cannabidiol and Minor Phytocannabinoids: A Preliminary Study to Assess Their Anti-Melanoma, Anti-Melanogenic, and Anti-Tyrosinase Properties. Pharmaceuticals. 2023; 16(5):648. https://doi.org/10.3390/ph16050648
Chicago/Turabian StyleGaweł-Bęben, Katarzyna, Karolina Czech, and Simon Vlad Luca. 2023. "Cannabidiol and Minor Phytocannabinoids: A Preliminary Study to Assess Their Anti-Melanoma, Anti-Melanogenic, and Anti-Tyrosinase Properties" Pharmaceuticals 16, no. 5: 648. https://doi.org/10.3390/ph16050648
APA StyleGaweł-Bęben, K., Czech, K., & Luca, S. V. (2023). Cannabidiol and Minor Phytocannabinoids: A Preliminary Study to Assess Their Anti-Melanoma, Anti-Melanogenic, and Anti-Tyrosinase Properties. Pharmaceuticals, 16(5), 648. https://doi.org/10.3390/ph16050648