

Function and Role of Histamine H1 Receptor in the Mammalian Heart

Abstract
:1. Introduction
- Histamine acts as a neurotransmitter in the central nervous system, but also plays a role as a neurotransmitter in the gut.
- Histamine has many effects on the skin and cutaneous immune system.
- Histamine induces the weal and flare reaction in the skin [1].
- Histamine is involved in contractions of the intestinal, but also vascular smooth muscle, leading to dilatation of capillaries and enhancing acid secretion in the stomach.
- Histamine induces bronchoconstriction, especially in allergic asthma, but also in anaphylactic reactions in the cardiovascular system.
- Other widely known effects of histamine include itching, rhinorrhoea (by stimulating secretory glands), and sneezing.
- Most relevant in this context, histamine contributes to the regulation of cardiac contraction [1].

{kind=link}
| H1R Affinity pKi | H2R Affinity pKi | Off-Target Effect | References | |
|---|---|---|---|---|
| 2-Methylhistamine | 17 | 4 | Hill et al., 1997 [12] | |
| 2-(2-Pyridyl)ethylamine (PEA) | 5.6 | 2.5 | Hill et al., 1997 [12] | |
| 2-(2-Thiazolyl) ethyl amine (ThEA) | 1 5.3, 2 26 | 2 2.2 | 3 Release of noradrenaline in the human heart | 1 Panula et al., 2015 [1] 2 Hill et al., 1990, 1997 [10,12] 3 Own unpublished data |
| 8-S-Lisuride | pKi 7.27 | Dopamine-, serotonin-, adrenergic receptors, | Pertz et al., 2006 [11] | |
| Bromocriptine | pKi 5.72 | Dopamine receptors | Pertz et al., 2006 [11] | |
| Histamine | 1 4.6 | 1 5.6 | 2 Release of noradrenaline from mouse heart | 1 Panula et al., 2015 [1] 2 Gergs et al., 2019 [13] |
| Histaprodifen | 5.7 | Panula et al., 2015 [1] | ||
| Methylhistaprodifen | 27.1 nM | Carman-Krzan et al., 2003 [14] | ||
| Suprahistaprodifen | 1 4.3 nM | 2 Release of noradrenaline in the mouse und human heart | 1 Carman-Krzan et al., 2003 [14] 2 Own unpublished data | |
| trans-PAT | 1.15 nM | Moniri et al., 2004 [15] |
2. Histamine H1 Receptor Structure
| H1-R Affinity IC50 Values If Not Stated Otherwise | H2-R Affinity IC50 Values, If Not Stated Otherwise | Off-Target Effect | References | |
|---|---|---|---|---|
| Amitryptiline | pKi 9.04 | pKB 6.95 | antiserotoninergic | Appl et al., 2011 [9] |
| Astemizol | 8 nM | Not done | anticholinergic | Kubo et al., 1987 [27] |
| (+) Chlorpheniramine | 0.4 nM | 1.2 µM | Hill et al., 1997 [12] | |
| Cetirizine | pKi 7.5 | Not done | Hill et al., 1997 [12] | |
| Chlorpromazine | 1.2 nM | 5.9 µM | anticholinergic | Hill et al., 1997 [12] |
| Clemastine | 0.26 nM | Not done | anticholinergic | Kubo et al., 1987 [27] |
| Clozapine | 8.65 | 6.82 | anticholinergic, antiadrenergic, antiserotinergic | Panula et al., 2015 [1] |
| Diphenhydramine | 7.89 | 5.80 | anticholinergic | Panula et al., 2015 [1] |
| Dimet(h)indene | R: pA2: 9.54 S: pA2: 7.86 | Not done | anticholinergic, antiadrenergic, antiserotinergic | Nicholson et al., 1991 [28], Pfaff et al., 1995 [29] |
| Doxepin | 0.06 nM | Not done | anticholinergic, antiadrenergic, antiserotinergic | Hill et al., 1997 [12] |
| Fluphenazine | pKi 8.25 | pKB 4.78 | antiserotoninergic dopamine receptor | Appl et al., 2011 [9] |
| Haloperidol | pKi 5.71 | pKB 5.94 | dopamine receptor | Appl et al., 2011 [9] |
| Imipramine | pKi 8.12 | pKi 6.26 | anticholinergic | Appl et al., 2011 [9] |
| Ketotifen | 1.02 nM | Not done | anticholinergic, antiserotinergic | Feng et al., 2020 [30] |
| Mepyramine = pyrilamine | 8.80 | 4.63 | anticholinergic | Panula et al., 2015 [1] |
| Mianserin | pKi 8.92 | pKi 6.36 | antiadrenergic, antiserotinergic | Appl et al., 2011 [9] |
| Olanzapine | pKi 8.52 | pKB 6.02 | anticholinergic, antidopaminergic, antiserotinergic | Appl et al., 2011 [9] |
| Perphenazine | pKi 8.59 | pKB 5.55 | anticholinergic, antidopaminergic, antiserotinergic | Appl et al., 2011 [9] |
| Promethazine | 1.2 nM | 3.0 µM | anticholinergic | Hill et al., 1997 [12] |
| Terfenadine | 1 7.92 | Not done | 2 anticholinergic | 1 Panula et al., 2015 [1] 2 Gillard et al., 2003 [31] |
| Triprolidine | 0.2 µM | Not done | anticholinergic, antiadrenergic, antiserotinergic | Carman-Krzan 1986 [32], Nicholson et al., 1991 [28], Pfaff et al., 1995 [29] |
3. Histamine H1 Receptor and Signal Transduction
4. Desensitisation, Downregulation and Upregulation of the Histamine H1 Receptor
5. Histamine H1 Receptor and Endothelial Cells
6. Histamine H1 Receptor and Smooth Muscle Cells
7. Histamine H1 Receptor in Cardiac Fibroblasts, Cardiac Mast Cells and Cardiac Blood Cells
8. Histamine H1 Receptor and Cardiomyocytes
9. Histamine H1 Receptor and Cardiac Electrophysiology
10. Histamine H1 Receptor in Animal Hearts
11. Histamine H1 Receptor in Human Hearts
11.1. Histamine H1 Receptor in the Human Atrium
11.2. Histamine H1 Receptor in the Human Ventricle
12. Histamine H1 Receptor in Disease
12.1. Arrhythmias
12.2. Sepsis and Inflammation
12.3. Others
13. Agonists and Antagonists
14. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Gi: | inhibitory guanosine triphosphate binding protein, |
| Gs: | stimulatory guanosine triphosphate binding protein, |
| Gq: | G protein binding protein q, |
| PLC: | phospholipase C, |
| PLA2: | phospholipase A2. |
| PKC: | protein kinase C, |
| NOS: | nitrogen monoxide synthase, |
| IKr: | rapidly inactivating potassium current, |
| IKs: | slowly inactivating potassium current, |
| cGMP: | cyclic 3′,5′-guanosine monophosphate, |
| DAG: | 1,2-diacylglyerol, |
| PDE: | phosphodiesterase, |
| RYR: | ryanodine receptor, |
| MLC kinase: | myosin light chain kinase, |
| LTCC: | L-type calcium ion channel, |
| SERCA: | sarcoplasmic reticulum Ca2+ adenosine triphosphate (ATP)-ase, |
| PLD: | phospholipase D, |
| Cam kinase II: | Ca2+-calmodulin-dependent protein kinase II, |
| AC: | adenylyl cyclase, NAADP: nicotinic acid adenine dinucleotide phosphate, |
| IP3: | inositol 1,2,4, trisphosphate, |
| PKA: | cAMP-dependent protein kinase, |
| EPAC: | exchange protein directly activated by cAMP, |
| MEK: | mitogen-activated protein kinase kinase, |
| PARP-1: | poly-ADP-ribose polymerase 1, |
| RAF kinase: | rapidly accelerated fibrosarcoma kinase |
| L-NAME: | NG-nitroarginine methyl ester, |
| MAP kinase: | mitogen-activated protein kinase, |
| CHO cells: | Chinese hamster ovary cells. |
References
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.S.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef]
- Neumann, J.; Kirchhefer, U.; Dhein, S.; Hofmann, B.; Gergs, U. The Roles of Cardiovascular H2-Histamine Receptors under Normal and Pathophysiological Conditions. Front. Pharmacol. 2021, 12, 732842. [Google Scholar] [CrossRef]
- Neumann, J.; Kirchhefer, U.; Dhein, S.; Hofmann, B.; Gergs, U. On the Cardiac Role of Histamine. In Horizons in World Cardiovascular Research; Bennington, E.H., Ed.; Nova Science Publishers: New York, NY, USA, 2022; Volume 22, pp. 1–75. ISBN 9781685075682. [Google Scholar]
- Akdis, C.A.; Simons, F.E.R. Histamine receptors are hot in immunopharmacology. Eur. J. Pharmacol. 2006, 533, 69–76. [Google Scholar] [CrossRef]
- Thangam, E.B.; Jemima, E.A.; Singh, H.; Baig, M.S.; Khan, M.; Mathias, C.B.; Church, M.K.; Saluja, R. The Role of Histamine and Histamine Receptors in Mast Cell-Mediated Allergy and Inflammation: The Hunt for New Therapeutic Targets. Front. Immunol. 2018, 9, 1873. [Google Scholar] [CrossRef]
- Hsieh, C.-Y.; Tsai, T.-F. Use of H-1 Antihistamine in Dermatology: More than Itch and Urticaria Control: A Systematic Review. Dermatol. Ther. 2021, 11, 719–732. [Google Scholar] [CrossRef]
- Leurs, R.; Church, M.K.; Taglialatela, M. H1-antihistamines: Inverse agonism, anti-inflammatory actions and cardiac effects. Clin. Exp. Allergy 2002, 32, 489–498. [Google Scholar] [CrossRef]
- Abernathy, A.; Alsina, L.; Greer, J.; Egerman, R. Transient Fetal Tachycardia After Intravenous Diphenhydramine Administration. Obstet. Gynecol. 2017, 130, 374–376. [Google Scholar] [CrossRef]
- Appl, H.; Holzammer, T.; Dove, S.; Haen, E.; Strasser, A.; Seifert, R. Interactions of recombinant human histamine H1R, H2R, H3R, and H4R receptors with 34 antidepressants and antipsychotics. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2012, 385, 145–170. [Google Scholar] [CrossRef]
- Hill, S.J. Distribution, properties, and functional characteristics of three classes of histamine receptor. Pharmacol. Rev. 1990, 42, 45–83. [Google Scholar]
- Pertz, H.H.; Görnemann, T.; Schurad, B.; Seifert, R.; Strasser, A. Striking differences of action of lisuride stereoisomers at histamine H1 receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2006, 374, 215–222. [Google Scholar] [CrossRef]
- Hill, S.J.; Ganellin, C.R.; Timmerman, H.; Schwartz, J.C.; Shankley, N.P.; Young, J.M.; Schunack, W.; Levi, R.; Haas, H.L. International Union of Pharmacology. XIII. Classification of histamine receptors. Pharmacol. Rev. 1997, 49, 253–278. [Google Scholar]
- Gergs, U.; Bernhardt, G.; Buchwalow, I.B.; Edler, H.; Fröba, J.; Keller, M.; Kirchhefer, U.; Köhler, F.; Mißlinger, N.; Wache, H.; et al. Initial Characterization of Transgenic Mice Overexpressing Human Histamine H2 Receptors. J. Pharmacol. Exp. Ther. 2019, 369, 129–141. [Google Scholar] [CrossRef]
- Carman-Krzan, M.; Bavec, A.; Zorko, M.; Schunack, W. Molecular characterization of specific H1-receptor agonists histaprodifen and its Nalpha-substituted analogues on bovine aortic H1-receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2003, 367, 538–546. [Google Scholar] [CrossRef]
- Moniri, N.H.; Covington-Strachan, D.; Booth, R.G. Ligand-directed functional heterogeneity of histamine H1 receptors: Novel dual-function ligands selectively activate and block H1-mediated phospholipase C and adenylyl cyclase signaling. J. Pharmacol. Exp. Ther. 2004, 311, 274–281. [Google Scholar] [CrossRef]
- de Backer, M.D.; Gommeren, W.; Moereels, H.; Nobels, G.; van Gompel, P.; Leysen, J.E.; Luyten, W.H. Genomic cloning, heterologous expression and pharmacological characterization of a human histamine H1 receptor. Biochem. Biophys. Res. Commun. 1993, 197, 1601–1608. [Google Scholar] [CrossRef]
- Moguilevsky, N.; Varsalona, F.; Noyer, M.; Gillard, M.; Guillaume, J.P.; Garcia, L.; Szpirer, C.; Szpirer, J.; Bollen, A. Stable expression of human H1-histamine-receptor cDNA in Chinese hamster ovary cells. Pharmacological characterisation of the protein, tissue distribution of messenger RNA and chromosomal localisation of the gene. Eur. J. Biochem. 1994, 224, 489–495. [Google Scholar] [CrossRef]
- Le Coniat, M.; Traiffort, E.; Ruat, M.; Arrang, J.M.; Berger, R. Chromosomal localization of the human histamine H1-receptor gene. Hum. Genet. 1994, 94, 186–188. [Google Scholar] [CrossRef]
- Wang, Y.X.; Kotlikoff, M.I. Signalling pathway for histamine activation of non-selective cation channels in equine tracheal myocytes. J. Physiol. 2000, 523 Pt 1, 131–138. [Google Scholar] [CrossRef]
- Inoue, I.; Yanai, K.; Kitamura, D.; Taniuchi, I.; Kobayashi, T.; Niimura, K.; Watanabe, T. Impaired locomotor activity and exploratory behavior in mice lacking histamine H1 receptors. Proc. Natl. Acad. Sci. USA 1996, 93, 13316–13320. [Google Scholar] [CrossRef]
- Suzuki, H.; Mobarakeh, J.I.; Nunoki, K.; Sukegawa, J.; Watanabe, H.; Kuramasu, A.; Watanabe, T.; Yanai, K.; Yanagisawa, T. Effects of activation of central nervous histamine receptors in cardiovascular regulation; studies in H(1) and H(2) receptor gene knockout mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2005, 371, 99–106. [Google Scholar] [CrossRef]
- Shimamura, T.; Shiroishi, M.; Weyand, S.; Tsujimoto, H.; Winter, G.; Katritch, V.; Abagyan, R.; Cherezov, V.; Liu, W.; Han, G.W.; et al. Structure of the human histamine H1 receptor complex with doxepin. Nature 2011, 475, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, H.; Terao, T.; Kitai, M.; Ikeda, M.; Yoshimura, Y.; Das, A.K.; Kitamura, Y.; Takeda, N.; Fukui, H. Involvement of protein kinase Cdelta/extracellular signal-regulated kinase/poly(ADP-ribose) polymerase-1 (PARP-1) signaling pathway in histamine-induced up-regulation of histamine H1 receptor gene expression in HeLa cells. J. Biol. Chem. 2011, 286, 30542–30551. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wei, X.; Shi, L.; Chen, B.; Zhao, G.; Yang, H. Integrative genomic analyses of the histamine H1 receptor and its role in cancer prediction. Int. J. Mol. Med. 2014, 33, 1019–1026. [Google Scholar] [CrossRef]
- Wientzek, M.; Allen, B.G.; McDonald-Jones, G.; Katz, S. Characterization of calcium-dependent forms of protein kinase C in adult rat ventricular myocytes. Mol. Cell. Biochem. 1997, 166, 11–23. [Google Scholar] [CrossRef]
- Bruysters, M.; Jongejan, A.; Gillard, M.; van de Manakker, F.; Bakker, R.A.; Chatelain, P.; Leurs, R. Pharmacological differences between human and guinea pig histamine H1 receptors: Asn84 (2.61) as key residue within an additional binding pocket in the H1 receptor. Mol. Pharmacol. 2005, 67, 1045–1052. [Google Scholar] [CrossRef]
- Kubo, N.; Shirakawa, O.; Kuno, T.; Tanaka, C. Antimuscarinic effects of antihistamines: Quantitative evaluation by receptor-binding assay. Jpn. J. Pharmacol. 1987, 43, 277–282. [Google Scholar] [CrossRef]
- Nicholson, A.N.; Pascoe, P.A.; Turner, C.; Ganellin, C.R.; Greengrass, P.M.; Casy, A.F.; Mercer, A.D. Sedation and histamine H1-receptor antagonism: Studies in man with the enantiomers of chlorpheniramine and dimethindene. Br. J. Pharmacol. 1991, 104, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Pfaff, O.; Hildebrandt, C.; Waelbroeck, M.; Hou, X.; Moser, U.; Mutschler, E.; Lambrecht, G. The (S)-(+)-enantiomer of dimethindene: A novel M2-selective muscarinic receptor antagonist. Eur. J. Pharmacol. 1995, 286, 229–240. [Google Scholar] [CrossRef]
- Feng, F.; Fawcett, J.P.; Zhang, H.; Tucker, I.G. Cell-based, animal and H1 receptor binding studies relative to the sedative effects of ketotifen and norketotifen atropisomers. J. Pharm. Pharmacol. 2020, 72, 507–518. [Google Scholar] [CrossRef]
- Gillard, M.; Christophe, B.; Wels, B.; Peck, M.; Massingham, R.; Chatelain, P. H1 antagonists: Receptor affinity versus selectivity. Inflamm. Res. 2003, 52 (Suppl. 1), S49–S50. [Google Scholar] [CrossRef]
- Carman-Krzan, M. Regulation of membrane histamine H1-receptor binding sites by guanine nucleotides, mono- and divalent cations. Agents Actions 1986, 18, 120–123. [Google Scholar] [CrossRef]
- Brighton, P.J.; Rana, S.; Challiss, R.J.; Konje, J.C.; Willets, J.M. Arrestins differentially regulate histamine- and oxytocin-evoked phospholipase C and mitogen-activated protein kinase signalling in myometrial cells. Br. J. Pharmacol. 2011, 162, 1603–1617. [Google Scholar] [CrossRef]
- Qi, X.-Y.; Vahdati Hassani, F.; Hoffmann, D.; Xiao, J.; Xiong, F.; Villeneuve, L.R.; Ljubojevic-Holzer, S.; Kamler, M.; Abu-Taha, I.; Heijman, J.; et al. Inositol Trisphosphate Receptors and Nuclear Calcium in Atrial Fibrillation. Circ. Res. 2021, 128, 619–635. [Google Scholar] [CrossRef]
- Bakker, R.A.; Wieland, K.; Timmerman, H.; Leurs, R. Constitutive activity of the histamine H(1) receptor reveals inverse agonism of histamine H(1) receptor antagonists. Eur. J. Pharmacol. 2000, 387, R5–R7. [Google Scholar] [CrossRef]
- Neumann, J.; Grobe, J.M.; Weisgut, J.; Schwelberger, H.G.; Fogel, W.A.; Marušáková, M.; Wache, H.; Bähre, H.; Buchwalow, I.B.; Dhein, S.; et al. Histamine can be Formed and Degraded in the Human and Mouse Heart. Front. Pharmacol. 2021, 12, 582916. [Google Scholar] [CrossRef]
- Ohuchi, Y.; Yanai, K.; Sakurai, E.; Fukui, H.; Yanagisawa, T.; Watanabe, T. Histamine-induced calcium mobilization in single cultured cells expressing histamine H1 receptors: A relationship between its sensitivity and the density of H1 receptors. Int. J. Mol. Med. 1998, 1, 355–360. [Google Scholar] [CrossRef]
- Valencia, S.; Hernández-Angeles, A.; Soria-Jasso, L.E.; Arias-Montaño, J.A. Histamine H(1) receptor activation inhibits the proliferation of human prostatic adenocarcinoma DU-145 cells. Prostate 2001, 48, 179–187. [Google Scholar] [CrossRef]
- Alonso, N.; Fernandez, N.; Notcovich, C.; Monczor, F.; Simaan, M.; Baldi, A.; Gutkind, J.S.; Davio, C.; Shayo, C. Cross-desensitization and cointernalization of H1 and H2 histamine receptors reveal new insights into histamine signal integration. Mol. Pharmacol. 2013, 83, 1087–1098. [Google Scholar] [CrossRef]
- Jain, R.; Watson, U.; Saini, D.K. ERK activated by Histamine H1 receptor is anti-proliferative through spatial restriction in the cytosol. Eur. J. Cell Biol. 2016, 95, 623–634. [Google Scholar] [CrossRef]
- Seifert, R.; Hagelüken, A.; Höer, A.; Höer, D.; Grünbaum, L.; Offermanns, S.; Schwaner, I.; Zingel, V.; Schunack, W.; Schultz, G. The H1 receptor agonist 2-(3-chlorophenyl)histamine activates Gi proteins in HL-60 cells through a mechanism that is independent of known histamine receptor subtypes. Mol. Pharmacol. 1994, 45, 578–586. [Google Scholar]
- Gutowski, S.; Smrcka, A.; Nowak, L.; Wu, D.G.; Simon, M.; Sternweis, P.C. Antibodies to the alpha q subfamily of guanine nucleotide-binding regulatory protein alpha subunits attenuate activation of phosphatidylinositol 4,5-bisphosphate hydrolysis by hormones. J. Biol. Chem. 1991, 266, 20519–20524. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y.; Ogura, T.; Uemura, H.; Saito, T.; Masuda, Y.; Nakaya, H. Histamine H1-receptor-mediated modulation of the delayed rectifier K+ current in guinea-pig atrial cells: Opposite effects on IKs and IKr. Br. J. Pharmacol. 1999, 128, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Sakuma, I.; Kanno, M. Differential effects of histamine mediated by histamine H1- and H2-receptors on contractility, spontaneous rate and cyclic nucleotides in the rabbit heart. Eur. J. Pharmacol. 1988, 153, 221–229. [Google Scholar] [CrossRef]
- Mittra, S.; Bourreau, J.-P. Gs and Gi coupling of adrenomedullin in adult rat ventricular myocytes. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1842–H1847. [Google Scholar] [CrossRef]
- Okazaki, K.; Ishikawa, T.; Inui, M.; Tada, M.; Goshima, K.; Okamoto, T.; Hidaka, H. KN-62, a specific Ca++/calmodulin-dependent protein kinase inhibitor, reversibly depresses the rate of beating of cultured fetal mouse cardiac myocytes. J. Pharmacol. Exp. Ther. 1994, 270, 1319–1324. [Google Scholar]
- Snider, R.M.; McKinney, M.; Forray, C.; Richelson, E. Neurotransmitter receptors mediate cyclic GMP formation by involvement of arachidonic acid and lipoxygenase. Proc. Natl. Acad. Sci. USA 1984, 81, 3905–3909. [Google Scholar] [CrossRef]
- Díaz Nebreda, A.; Zappia, C.D.; Rodríguez González, A.; Sahores, A.; Sosa, M.; Burghi, V.; Monczor, F.; Davio, C.; Fernández, N.; Shayo, C. Involvement of histamine H1 and H2 receptor inverse agonists in receptor’s crossregulation. Eur. J. Pharmacol. 2019, 847, 42–52. [Google Scholar] [CrossRef]
- Dale, P.; Head, V.; Dowling, M.R.; Taylor, C.W. Selective inhibition of histamine-evoked Ca2+ signals by compartmentalized cAMP in human bronchial airway smooth muscle cells. Cell Calcium 2018, 71, 53–64. [Google Scholar] [CrossRef]
- Leurs, R.; Traiffort, E.; Arrang, J.M.; Tardivel-Lacombe, J.; Ruat, M.; Schwartz, J.C. Guinea pig histamine H1 receptor. II. Stable expression in Chinese hamster ovary cells reveals the interaction with three major signal transduction pathways. J. Neurochem. 1994, 62, 519–527. [Google Scholar] [CrossRef]
- Burch, R.M.; Axelrod, J. Dissociation of bradykinin-induced prostaglandin formation from phosphatidylinositol turnover in Swiss 3T3 fibroblasts: Evidence for G protein regulation of phospholipase A2. Proc. Natl. Acad. Sci. USA 1987, 84, 6374–6378. [Google Scholar] [CrossRef]
- Hermes, C.; König, G.M.; Crüsemann, M. The chromodepsins–chemistry, biology and biosynthesis of a selective Gq inhibitor natural product family. Nat. Prod. Rep. 2021, 38, 2276–2292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Nielsen, A.L.; Strømgaard, K. Recent achievements in developing selective Gq inhibitors. Med. Res. Rev. 2020, 40, 135–157. [Google Scholar] [CrossRef]
- Hattori, Y.; Nakaya, H.; Endou, M.; Kanno, M. Inotropic, electrophysiological and biochemical responses to histamine in rabbit papillary muscles: Evidence for coexistence of H1- and H2-receptors. J. Pharmacol. Exp. Ther. 1990, 253, 250–256. [Google Scholar]
- Ibarra, C.; Estrada, M.; Carrasco, L.; Chiong, M.; Liberona, J.L.; Cardenas, C.; Díaz-Araya, G.; Jaimovich, E.; Lavandero, S. Insulin-like growth factor-1 induces an inositol 1,4,5-trisphosphate-dependent increase in nuclear and cytosolic calcium in cultured rat cardiac myocytes. J. Biol. Chem. 2004, 279, 7554–7565. [Google Scholar] [CrossRef]
- Sankar, N.; deTombe, P.P.; Mignery, G.A. Calcineurin-NFATc regulates type 2 inositol 1,4,5-trisphosphate receptor (InsP3R2) expression during cardiac remodeling. J. Biol. Chem. 2014, 289, 6188–6198. [Google Scholar] [CrossRef] [PubMed]
- Iwata, K.; Luo, J.; Penn, R.B.; Benovic, J.L. Bimodal regulation of the human H1 histamine receptor by G protein-coupled receptor kinase 2. J. Biol. Chem. 2005, 280, 2197–2204. [Google Scholar] [CrossRef]
- Smit, M.J.; Bloemers, S.M.; Leurs, R.; Tertoolen, L.G.; Bast, A.; de Laat, S.W.; Timmerman, H. Short-term desensitization of the histamine H1 receptor in human HeLa cells: Involvement of protein kinase C dependent and independent pathways. Br. J. Pharmacol. 1992, 107, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Mizuguchi, H.; Ono, S.; Hattori, M.; Fukui, H. Inverse agonistic activity of antihistamines and suppression of histamine H1 receptor gene expression. J. Pharmacol. Sci. 2012, 118, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Nariai, Y.; Mizuguchi, H.; Ogasawara, T.; Nagai, H.; Sasaki, Y.; Okamoto, Y.; Yoshimura, Y.; Kitamura, Y.; Nemoto, H.; Takeda, N.; et al. Disruption of Heat Shock Protein 90 (Hsp90)-Protein Kinase Cδ (PKCδ) Interaction by (-)-Maackiain Suppresses Histamine H1 Receptor Gene Transcription in HeLa Cells. J. Biol. Chem. 2015, 290, 27393–27402. [Google Scholar] [CrossRef] [PubMed]
- Birkenkamp, K.U.; Tuyt, L.M.; Lummen, C.; Wierenga, A.T.; Kruijer, W.; Vellenga, E. The p38 MAP kinase inhibitor SB203580 enhances nuclear factor-kappa B transcriptional activity by a non-specific effect upon the ERK pathway. Br. J. Pharmacol. 2000, 131, 99–107. [Google Scholar] [CrossRef]
- Moïse, N.; Dingar, D.; Mamarbachi, A.M.; Villeneuve, L.R.; Farhat, N.; Gaestel, M.; Khairallah, M.; Allen, B.G. Characterization of a novel MK3 splice variant from murine ventricular myocardium. Cell. Signal. 2010, 22, 1502–1512. [Google Scholar] [CrossRef]
- Bil-Lula, I.; Krzywonos-Zawadzka, A.; Sawicka, J.; Bialy, D.; Wawrzynska, M.; Wozniak, M.; Sawicki, G. L-NAME improves doxycycline and ML-7 cardioprotection from oxidative stress. Front. Biosci. 2018, 23, 298–309. [Google Scholar] [CrossRef]
- Sun, Y.; Oami, T.; Liang, Z.; Miniet, A.A.; Burd, E.M.; Ford, M.L.; Coopersmith, C.M. Membrane Permeant Inhibitor of Myosin Light Chain Kinase Worsens Survival in Murine Polymicrobial Sepsis. Shock 2021, 56, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Pustovit, K.B.; Kuzmin, V.S.; Abramochkin, D.V. Diadenosine tetra- and pentaphosphates affect contractility and bioelectrical activity in the rat heart via P2 purinergic receptors. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2016, 389, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S.M.; Foote, K.; Kunuthur, S.; Gosain, R.; Tan, N.; Tyser, R.; Zhao, Y.J.; Graeff, R.; Ganesan, A.; Duchen, M.R.; et al. Inhibition of NAADP signalling on reperfusion protects the heart by preventing lethal calcium oscillations via two-pore channel 1 and opening of the mitochondrial permeability transition pore. Cardiovasc. Res. 2015, 108, 357–366. [Google Scholar] [CrossRef]
- Kostić, M.M.; Jakovljević, V.L. Role of histamine in the regulation of coronary circulation. Physiol. Res. 1996, 45, 297–303. [Google Scholar]
- Stemmer, P.; Akera, T. Concealed positive force-frequency relationships in rat and mouse cardiac muscle revealed by ryanodine. Am. J. Physiol. 1986, 251, H1106–H1110. [Google Scholar] [CrossRef]
- Neumann, J.; Voss, R.; Laufs, U.; Werner, C.; Gergs, U. Phosphodiesterases 2, 3 and 4 can decrease cardiac effects of H2-histamine-receptor activation in isolated atria of transgenic mice. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2021, 394, 1215–1229. [Google Scholar] [CrossRef]
- Castro, L.R.V.; Verde, I.; Cooper, D.M.F.; Fischmeister, R. Cyclic guanosine monophosphate compartmentation in rat cardiac myocytes. Circulation 2006, 113, 2221–2228. [Google Scholar] [CrossRef]
- Brown, L.; Lorenz, B.; Erdmann, E. Reduced positive inotropic effects in diseased human ventricular myocardium. Cardiovasc. Res. 1986, 20, 516–520. [Google Scholar] [CrossRef]
- Ukita, T.; Sugahara, M.; Terakawa, Y.; Kuroda, T.; Wada, K.; Nakata, A.; Ohmachi, Y.; Kikkawa, H.; Ikezawa, K.; Naito, K. Novel, potent, and selective phosphodiesterase-4 inhibitors as antiasthmatic agents: Synthesis and biological activities of a series of 1-pyridylnaphthalene derivatives. J. Med. Chem. 1999, 42, 1088–1099. [Google Scholar] [CrossRef]
- McDermott, M.I.; Wang, Y.; Wakelam, M.J.O.; Bankaitis, V.A. Mammalian phospholipase D: Function, and therapeutics. Prog. Lipid Res. 2020, 78, 101018. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, J.; Kim, S.; Shin, M.K.; Min, D.S.; Shin, T. Differential expression of phospholipases D1 and D2 in mouse tissues. Cell Biol. Int. 2007, 31, 148–155. [Google Scholar] [CrossRef]
- Taylor, E.J.A.; Pantazaka, E.; Shelley, K.L.; Taylor, C.W. Prostaglandin E2 Inhibits Histamine-Evoked Ca2+ Release in Human Aortic Smooth Muscle Cells through Hyperactive cAMP Signaling Junctions and Protein Kinase A. Mol. Pharmacol. 2017, 92, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Esposito, B.; Gambara, G.; Lewis, A.M.; Palombi, F.; D’Alessio, A.; Taylor, L.X.; Genazzani, A.A.; Ziparo, E.; Galione, A.; Churchill, G.C.; et al. NAADP links histamine H1 receptors to secretion of von Willebrand factor in human endothelial cells. Blood 2011, 117, 4968–4977. [Google Scholar] [CrossRef]
- Rogers, T.B.; Inesi, G.; Wade, R.; Lederer, W.J. Use of thapsigargin to study Ca2+ homeostasis in cardiac cells. Biosci. Rep. 1995, 15, 341–349. [Google Scholar] [CrossRef]
- Kemecsei, P.; Miklós, Z.; Bíró, T.; Marincsák, R.; Tóth, B.I.; Komlódi-Pásztor, E.; Barnucz, E.; Mirk, E.; van der Vusse, G.J.; Ligeti, L.; et al. Hearts of surviving MLP-KO mice show transient changes of intracellular calcium handling. Mol. Cell. Biochem. 2010, 342, 251–260. [Google Scholar] [CrossRef]
- Hattori, Y.; Kanno, M. Effect of Ni2+ on the multiphasic positive inotropic responses to histamine mediated by H1-receptors in left atria of guinea pigs. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1985, 329, 188–194. [Google Scholar] [CrossRef]
- Akaishi, Y.; Hattori, Y.; Yoshimoto, K.; Kitabatake, A.; Yasuda, K.; Kanno, M. Involvement of tyrosine phosphorylation in the positive inotropic effect produced by H(1)-receptors with histamine in guinea-pig left atrium. Br. J. Pharmacol. 2000, 130, 907–915. [Google Scholar] [CrossRef]
- Hattori, Y.; Endou, M.; Shirota, M.; Kanno, M. Dissociation of phosphoinositide hydrolysis and positive inotropic effect of histamine mediated by H1-receptors in guinea-pig left atria. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1989, 340, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Gando, S.; Endou, M.; Kanno, M. Characterization of histamine receptors modulating inotropic and biochemical activities in rabbit left atria. Eur. J. Pharmacol. 1991, 196, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Gando, S.; Nagashima, M.; Kanno, M. Histamine receptors mediating a positive inotropic effect in guinea pig and rabbit ventricular myocardium: Distribution of the receptors and their possible intracellular coupling processes. Jpn. J. Pharmacol. 1994, 65, 327–336. [Google Scholar] [CrossRef]
- Chiba, S. Absence of blocking effect of burimamide and metiamide on positive chronotropic and inotropic responses to histamine in isolated dog atrium. Jpn. J. Pharmacol. 1977, 27, 267–273. [Google Scholar] [CrossRef]
- Chiba, S. Blocking effect of tripelennamine on histamine--induced positive chronotropic and inotropic responses of the dog atrium. Tohoku J. Exp. Med. 1976, 120, 299–300. [Google Scholar] [CrossRef] [PubMed]
- McNeill, J.H. Histamine and the heart. Can. J. Physiol. Pharmacol. 1984, 62, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Motomura, S.; Hashimoto, K. Histamine H2-receptor mediated positive dromotropic effect in the canine atrioventricular node. Jpn. J. Pharmacol. 1989, 49, 325–335. [Google Scholar] [CrossRef]
- Zavecz, J.H.; Levi, R. Histamine-induced negative inotropism: Mediation by H1-receptors. J. Pharmacol. Exp. Ther. 1978, 206, 274–280. [Google Scholar]
- Mantelli, L.; Amerini, S.; Filippi, S.; Ledda, F. Positive inotropic effects of CGRP and isoprenaline: Analogies and differences. Neuropeptides 1992, 23, 193–198. [Google Scholar] [CrossRef]
- Reinhardt, D.; Wagner, J.; Schümann, H.J. Differentiation of H1- and H2-receptors mediating positive chrono- and inotropic responses to histamine on atrial preparations of the guinea-pig. Agents Actions 1974, 4, 217–221. [Google Scholar] [CrossRef]
- Steinburg, M.I.; Holland, D.R. Separate receptors mediating the positive inotropic and chronotropic effect of histamine in guinea-pig atria. Eur. J. Pharmacol. 1975, 34, 95–104. [Google Scholar] [CrossRef]
- Reinhardt, D.; Schmidt, U.; Brodde, O.E.; Schümann, H.J. H1—And H2-receptor mediated responses to histamine on contractility and cyclic AMP of atrial and papillary muscles from guinea-pig hearts. Agents Actions 1977, 7, 1–12. [Google Scholar] [CrossRef]
- Verma, S.C.; McNeill, J.H. Cardiac histamine receptors: Differences between left and right atria and right ventricle. J. Pharmacol. Exp. Ther. 1977, 200, 352–362. [Google Scholar]
- Amerini, S.; Franconi, F.; Ledda, F.; Mantelli, L.; Mugelli, A. H1- and H2-receptors in the guinea-pig heart: An electrophysiological study. Agents Actions 1982, 12, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.N.; Broadley, K.J. Analysis of the direct and indirect effects of adenosine on atrial and ventricular cardiac muscle. Can. J. Physiol. Pharmacol. 1989, 67, 294–303. [Google Scholar] [CrossRef]
- Wilson, C.; Broadley, K.J. A comparison of the responses to histamine of left and right atria and papillary muscles of guinea-pig hearts. Gen. Pharmacol. 1981, 12, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Levi, R.; Kuye, J.O. Pharmacological characterization of cardiac histamine receptors: Sensitivity to H1-receptor antagonists. Eur. J. Pharmacol. 1974, 27, 330–338. [Google Scholar] [CrossRef]
- Rayo Abella, L.; Pockes, S.; Keller, M.; Bernhardt, G.; Schindler, L.; Gergs, U.; Kirchhefer, U.; Hofmann, B.; Neumann, J. A novel transgenic mouse to study cardiac effects of H1-histamine receptors. In Proceedings of the 50th Anniversary Meeting of the EHRS, Hannover, Germany, 1–3 September 2022; European Histamine Research Society: Hannover, Germany, 2022; p. 15. [Google Scholar]
- Sanders, L.; Lynham, J.A.; Kaumann, A.J. Chronic beta 1-adrenoceptor blockade sensitises the H1 and H2 receptor systems in human atrium: Rôle of cyclic nucleotides. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1996, 353, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.G.; Levi, R.; Graver, L.M.; Robertson, D.A.; Gay, W.A. Inotropic effects of histamine in human myocardium: Differentiation between positive and negative components. J. Cardiovasc. Pharmacol. 1984, 6, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Genovese, A.; Gross, S.S.; Sakuma, I.; Levi, R. Adenosine promotes histamine H1-mediated negative chronotropic and inotropic effects on human atrial myocardium. J. Pharmacol. Exp. Ther. 1988, 247, 844–849. [Google Scholar]
- Du, X.Y.; Schoemaker, R.G.; Bax, W.A.; Bos, E.; Saxena, P.R. Effects of histamine on porcine isolated myocardium: Differentiation from effects on human tissue. J. Cardiovasc. Pharmacol. 1993, 22, 468–473. [Google Scholar] [CrossRef]
- Coruzzi, G.; Bongrani, S.; Bertaccini, G. Histamine receptors in the heart and coronary vessels of rabbits. Pharmacol. Res. Commun. 1979, 11, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Nakaya, H.; Tohse, N.; Kanno, M. Effects of Ca2+ channel antagonists and ryanodine on H1-receptor mediated electromechanical response to histamine in guinea-pig left atria. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1988, 337, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Laher, I.; McNeill, J.H. Effects of histamine on rat isolated atria. Can. J. Physiol. Pharmacol. 1980, 58, 1114–1116. [Google Scholar] [CrossRef] [PubMed]
- Kiniwa, M.; Tasaka, K. Histamine and its actions on isolated tissues of lower vertebrates. Methods Find. Exp. Clin. Pharmacol. 1989, 11, 87–95. [Google Scholar] [PubMed]
- Eto, M.; Kitazawa, T.; Yazawa, M.; Mukai, H.; Ono, Y.; Brautigan, D.L. Histamine-induced vasoconstriction involves phosphorylation of a specific inhibitor protein for myosin phosphatase by protein kinase C alpha and delta isoforms. J. Biol. Chem. 2001, 276, 29072–29078. [Google Scholar] [CrossRef]
- Kolosova, I.A.; Ma, S.-F.; Adyshev, D.M.; Wang, P.; Ohba, M.; Natarajan, V.; Garcia, J.G.N.; Verin, A.D. Role of CPI-17 in the regulation of endothelial cytoskeleton. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, L970–L980. [Google Scholar] [CrossRef]
- Herzig, S.; Neumann, J. Effects of serine/threonine protein phosphatases on ion channels in excitable membranes. Physiol. Rev. 2000, 80, 173–210. [Google Scholar] [CrossRef]
- Dessauer, C.W.; Watts, V.J.; Ostrom, R.S.; Conti, M.; Dove, S.; Seifert, R. International Union of Basic and Clinical Pharmacology. CI. Structures and Small Molecule Modulators of Mammalian Adenylyl Cyclases. Pharmacol. Rev. 2017, 69, 93–139. [Google Scholar] [CrossRef]
- Marino, A.; Levi, R. Salvaging the Ischemic Heart: Gi-Coupled Receptors in Mast Cells Activate a PKCε/ALDH2 Pathway Providing Anti-RAS Cardioprotection. Curr. Med. Chem. 2018, 25, 4416–4431. [Google Scholar] [CrossRef]
- Noble, E.P.; Bommer, M.; Sincini, E.; Costa, T.; Herz, A. H1-histaminergic activation stimulates inositol-1-phosphate accumulation in chromaffin cells. Biochem. Biophys. Res. Commun. 1986, 135, 566–573. [Google Scholar] [CrossRef]
- Richelson, E. Histamine H1 receptor-mediated guanosine 3′,5′-monophosphate formation by cultured mouse neuroblastoma cells. Science 1978, 201, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Richelson, E. Tricyclic antidepressants block histamine H1 receptors of mouse neuroblastoma cells. Nature 1978, 274, 176–177. [Google Scholar] [CrossRef]
- Vandecasteele, G.; Verde, I.; Rücker-Martin, C.; Donzeau-Gouge, P.; Fischmeister, R. Cyclic GMP regulation of the L-type Ca(2+) channel current in human atrial myocytes. J. Physiol. 2001, 533, 329–340. [Google Scholar] [CrossRef]
- Diks, S.H.; Hardwick, J.C.; Diab, R.M.; van Santen, M.M.; Versteeg, H.H.; van Deventer, S.J.H.; Richel, D.J.; Peppelenbosch, M.P. Activation of the canonical beta-catenin pathway by histamine. J. Biol. Chem. 2003, 278, 52491–52496. [Google Scholar] [CrossRef] [PubMed]
- Burghi, V.; Echeverría, E.B.; Zappia, C.D.; Díaz Nebreda, A.; Ripoll, S.; Gómez, N.; Shayo, C.; Davio, C.A.; Monczor, F.; Fernández, N.C. Biased agonism at histamine H1 receptor: Desensitization, internalization and MAPK activation triggered by antihistamines. Eur. J. Pharmacol. 2021, 896, 173913. [Google Scholar] [CrossRef]
- Notcovich, C.; Diez, F.; Tubio, M.R.; Baldi, A.; Kazanietz, M.G.; Davio, C.; Shayo, C. Histamine acting on H1 receptor promotes inhibition of proliferation via PLC, RAC, and JNK-dependent pathways. Exp. Cell Res. 2010, 316, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Burkhardt, C.; Heinrich, U.-R.; Brausch, I.; Xia, N.; Förstermann, U. Histamine upregulates gene expression of endothelial nitric oxide synthase in human vascular endothelial cells. Circulation 2003, 107, 2348–2354. [Google Scholar] [CrossRef]
- Okamoto, Y.; Aung, N.Y.; Tanaka, M.; Takeda, Y.; Takagi, D.; Igarashi, W.; Ishii, K.; Yamakawa, M.; Ono, K. Preferential Expression of Ca2+-Stimulable Adenylyl Cyclase III in the Supraventricular Area, including Arrhythmogenic Pulmonary Vein of the Rat Heart. Biomolecules 2022, 12, 724. [Google Scholar] [CrossRef]
- Ludwig, M.-G.; Seuwen, K. Characterization of the human adenylyl cyclase gene family: cDNA, gene structure, and tissue distribution of the nine isoforms. J. Recept. Signal Transduct. Res. 2002, 22, 79–110. [Google Scholar] [CrossRef]
- Sunahara, R.K.; Taussig, R. Isoforms of mammalian adenylyl cyclase: Multiplicities of signaling. Mol. Interv. 2002, 2, 168–184. [Google Scholar] [CrossRef]
- Forward, N.A.; Furlong, S.J.; Yang, Y.; Lin, T.-J.; Hoskin, D.W. Mast cells down-regulate CD4+CD25+ T regulatory cell suppressor function via histamine H1 receptor interaction. J. Immunol. 2009, 183, 3014–3022. [Google Scholar] [CrossRef]
- Felix, S.B.; Buschauer, A.; Baumann, G. Therapeutic value of H2-receptor stimulation in congestive heart failure. Hemodynamic effects of BU-E-76, BU-E-75 and arpromidine (BU-E-50) in comparison to impromidine. Agents Actions Suppl. 1991, 33, 257–269. [Google Scholar] [CrossRef]
- Fujimoto, K.; Ohta, K.; Kangawa, K.; Kikkawa, U.; Ogino, S.; Fukui, H. Identification of protein kinase C phosphorylation sites involved in phorbol ester-induced desensitization of the histamine H1 receptor. Mol. Pharmacol. 1999, 55, 735–742. [Google Scholar]
- Hamada, E.; Nakajima, T.; Ota, S.; Terano, A.; Omata, M.; Nakade, S.; Mikoshiba, K.; Kurachi, Y. Activation of Ca(2+)-dependent K+ current by acetylcholine and histamine in a human gastric epithelial cell line. J. Gen. Physiol. 1993, 102, 667–692. [Google Scholar] [CrossRef]
- Herepath, M.L.; Broadley, K.J. Desensitization of beta-adrenoceptor-mediated functional responses of guinea pig atria by in vitro incubation with isoprenaline. J. Cardiovasc. Pharmacol. 1990, 15, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Horio, S.; Kato, T.; Ogawa, M.; Fujimoto, K.; Fukui, H. Two threonine residues and two serine residues in the second and third intracellular loops are both involved in histamine H1 receptor downregulation. FEBS Lett. 2004, 573, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Hishinuma, S.; Komazaki, H.; Fukui, H.; Shoji, M. Ubiquitin/proteasome-dependent down-regulation following clathrin-mediated internalization of histamine H1-receptors in Chinese hamster ovary cells. J. Neurochem. 2010, 113, 990–1001. [Google Scholar] [CrossRef]
- Fernandez, N.; Gottardo, F.L.; Alonso, M.N.; Monczor, F.; Shayo, C.; Davio, C. Roles of phosphorylation-dependent and -independent mechanisms in the regulation of histamine H2 receptor by G protein-coupled receptor kinase 2. J. Biol. Chem. 2011, 286, 28697–28706. [Google Scholar] [CrossRef] [PubMed]
- Self, T.J.; Oakley, S.M.; Hill, S.J. Clathrin-independent internalization of the human histamine H1-receptor in CHO-K1 cells. Br. J. Pharmacol. 2005, 146, 612–624. [Google Scholar] [CrossRef]
- Dinh, Q.T.; Cryer, A.; Dinh, S.; Peiser, C.; Wu, S.; Springer, J.; Hamelmann, E.; Klapp, B.F.; Heppt, W.; Fischer, A. Transcriptional up-regulation of histamine receptor-1 in epithelial, mucus and inflammatory cells in perennial allergic rhinitis. Clin. Exp. Allergy 2005, 35, 1443–1448. [Google Scholar] [CrossRef]
- Mizuguchi, H.; Miyagi, K.; Terao, T.; Sakamoto, N.; Yamawaki, Y.; Adachi, T.; Ono, S.; Sasaki, Y.; Yoshimura, Y.; Kitamura, Y.; et al. PMA-induced dissociation of Ku86 from the promoter causes transcriptional up-regulation of histamine H(1) receptor. Sci. Rep. 2012, 2, 916. [Google Scholar] [CrossRef]
- Das, A.K.; Yoshimura, S.; Mishima, R.; Fujimoto, K.; Mizuguchi, H.; Dev, S.; Wakayama, Y.; Kitamura, Y.; Horio, S.; Takeda, N.; et al. Stimulation of histamine H1 receptor up-regulates histamine H1 receptor itself through activation of receptor gene transcription. J. Pharmacol. Sci. 2007, 103, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Mak, J.C.; Roffel, A.F.; Katsunuma, T.; Elzinga, C.R.; Zaagsma, J.; Barnes, P.J. Up-regulation of airway smooth muscle histamine H(1) receptor mRNA, protein, and function by beta(2)-adrenoceptor activation. Mol. Pharmacol. 2000, 57, 857–864. [Google Scholar] [PubMed]
- Lantoine, F.; Iouzalen, L.; Devynck, M.A.; Millanvoye-Van Brussel, E.; David-Dufilho, M. Nitric oxide production in human endothelial cells stimulated by histamine requires Ca2+ influx. Biochem. J. 1998, 330 Pt 2, 695–699. [Google Scholar] [CrossRef]
- Kishi, F.; Nakaya, Y.; Takahashi, A.; Miyoshi, H.; Nomura, M.; Saito, K. Intracellular and extracellular Ca2+ regulate histamine-induced release of nitric oxide in vascular endothelial cells as shown with sensitive and selective nitric oxide electrodes. Pharmacol. Res. 1996, 33, 123–126. [Google Scholar] [CrossRef]
- Bolz, S.S.; Pohl, U. Indomethacin enhances endothelial NO release—evidence for a role of PGI2 in the autocrine control of calcium-dependent autacoid production. Cardiovasc. Res. 1997, 36, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Carman-Krzan, M.; Krzan, M.; Schunack, W. Pharmacological properties of cardiovascular histamine H1 receptor binding sites: Characterisation with 2-phenylhistamines. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1997, 355, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Schuschke, D.A.; Saari, J.T. Histamine-mediated vasoconstriction and cAMP levels in coronary arteries of the isolated rabbit heart. Pharmacology 1989, 38, 23–33. [Google Scholar] [CrossRef]
- Levick, S.P.; Widiapradja, A. Mast Cells: Key Contributors to Cardiac Fibrosis. Int. J. Mol. Sci. 2018, 19, 231. [Google Scholar] [CrossRef]
- Saheera, S.; Potnuri, A.G.; Guha, A.; Palaniyandi, S.S.; Thandavarayan, R.A. Histamine 2 receptors in cardiovascular biology: A friend for the heart. Drug Discov. Today 2022, 27, 234–245. [Google Scholar] [CrossRef]
- Reid, A.C.; Brazin, J.A.; Morrey, C.; Silver, R.B.; Levi, R. Targeting cardiac mast cells: Pharmacological modulation of the local renin-angiotensin system. Curr. Pharm. Des. 2011, 17, 3744–3752. [Google Scholar] [CrossRef]
- Wu, R.-L.; Anthes, J.C.; Kreutner, W.; Harris, A.G.; West, R.E. Desloratadine inhibits constitutive and histamine-stimulated nuclear factor-kappaB activity consistent with inverse agonism at the histamine H1 Receptor. Int. Arch. Allergy Immunol. 2004, 135, 313–318. [Google Scholar] [CrossRef]
- Levi, R.; Malm, J.R.; Bowman, F.O.; Rosen, M.R. The arrhythmogenic actions of histamine on human atrial fibers. Circ. Res. 1981, 49, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Jutel, M.; Akdis, M.; Akdis, C.A. Histamine, histamine receptors and their role in immune pathology. Clin. Exp. Allergy 2009, 39, 1786–1800. [Google Scholar] [CrossRef] [PubMed]
- Holden, C.A.; Chan, S.C.; Norris, S.; Hanifin, J.M. Histamine induced elevation of cyclic AMP phosphodiesterase activity in human monocytes. Agents Actions 1987, 22, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Yamagata, K.; Nakayamada, S.; Sawamukai, N.; Yamaoka, K.; Sakata, K.; Nakano, K.; Tanaka, Y. Histamine inhibits differentiation of skin fibroblasts into myofibroblasts. Biochem. Biophys. Res. Commun. 2015, 463, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.L.; Johnson, C.G.; Bazan, E.; Garver, D.; Gruenstein, E.; Ahluwalia, M. Histamine receptors in human fibroblasts: Inositol phosphates, Ca2+, and cell growth. Am. J. Physiol. 1990, 258, C533–C543. [Google Scholar] [CrossRef] [PubMed]
- Case, L.K.; Moussawi, M.; Roberts, B.; Noubade, R.; Huber, S.A.; Teuscher, C. Histamine H(1) receptor signaling regulates effector T cell responses and susceptibility to coxsackievirus B3-induced myocarditis. Cell. Immunol. 2012, 272, 269–274. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, X.; Zhu, B.; Ding, S.; Shi, H.; Yang, X. Disruption of histamine/H1R-STAT3-SLC7A11 axis exacerbates doxorubicin-induced cardiac ferroptosis. Free Radic. Biol. Med. 2022, 192, 98–114. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Hattori, Y.; Houzen, H.; Kanno, M.; Yasuda, K. Histamine H1-receptor-mediated increase in the Ca2+ transient without a change in the Ca2+ current in electrically stimulated guinea-pig atrial myocytes. Br. J. Pharmacol. 1998, 124, 1744–1750. [Google Scholar] [CrossRef]
- Matsuda, N.; Jesmin, S.; Takahashi, Y.; Hatta, E.; Kobayashi, M.; Matsuyama, K.; Kawakami, N.; Sakuma, I.; Gando, S.; Fukui, H.; et al. Histamine H1 and H2 receptor gene and protein levels are differentially expressed in the hearts of rodents and humans. J. Pharmacol. Exp. Ther. 2004, 309, 786–795. [Google Scholar] [CrossRef]
- Levi, R.; Capurro, N.; Lee, C.H. Pharmacological characterization of cardiac histamine receptors: Sensitivity to H1-and H2-receptor agonists and antagonists. Eur. J. Pharmacol. 1975, 30, 328–335. [Google Scholar] [CrossRef]
- Levi, R.; Ganellin, C.R.; Allan, G.; Willens, H.J. Selective impairment of atrioventricular conduction by 2-(2-pyridyl)-ethylamine and 2-(2-thiazolyl)-ethylamine, two histamine H1-receptor agonists. Eur. J. Pharmacol. 1975, 34, 237–240. [Google Scholar] [CrossRef]
- Borchard, U.; Hafner, D. Electrophysiological characterization of histamine receptor subtypes in mammalian heart preparations. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1986, 334, 294–302. [Google Scholar] [CrossRef]
- Borchard, U.; Hafner, D.; Hirth, C. Electrophysiological actions of histamine and H1-, H2-receptor antagonists in cardiac tissue. Agents Actions 1986, 18, 186–190. [Google Scholar] [CrossRef]
- Satoh, H. Modulation of the automaticity by histamine and cimetidine in rabbit sino-atrial node cells. Gen. Pharmacol. 1993, 24, 1213–1222. [Google Scholar] [CrossRef]
- Laher, I.; McNeill, J.H. Effects of 2-(2-pyridyl)ethylamine (PEA) on the isolated guinea-pig heart. Agents Actions 1980, 10, 417–421. [Google Scholar] [CrossRef]
- Salata, J.J.; Jurkiewicz, N.K.; Wallace, A.A.; Stupienski, R.F.; Guinosso, P.J.; Lynch, J.J. Cardiac electrophysiological actions of the histamine H1-receptor antagonists astemizole and terfenadine compared with chlorpheniramine and pyrilamine. Circ. Res. 1995, 76, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, R.; Bristow, M.R.; Stinson, E.B.; Harrison, D.C. Histamine receptors in the human heart. Life Sci. 1980, 26, 2245–2249. [Google Scholar] [CrossRef] [PubMed]
- Rappen-Cremer, E.; Borchard, U.; Hafner, D.; Berger, F. H1-receptor reserves in guinea-pig left atria, trachea and pig coronary artery as identified by phenoxybenzamine. Agents Actions 1989, 28, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, R.D.; Brennan, S.; Jackson, R.; Beech, A.J.; Bengreed, A.; Waldschmidt, H.V.; Tesmer, J.J.G.; Challiss, R.A.J.; Willets, J.M. Small-Molecule G Protein-Coupled Receptor Kinase Inhibitors Attenuate G Protein-Coupled Receptor Kinase 2-Mediated Desensitization of Vasoconstrictor-Induced Arterial Contractions. Mol. Pharmacol. 2018, 94, 1079–1091. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, I.; Gross, S.S.; Levi, R. Positive inotropic effect of histamine on guinea pig left atrium: H1-receptor-induced stimulation of phosphoinositide turnover. J. Pharmacol. Exp. Ther. 1988, 247, 466–472. [Google Scholar] [PubMed]
- Matsuda, N.; Hattori, Y.; Zhang, X.-H.; Fukui, H.; Kemmotsu, O.; Gando, S. Contractions to histamine in pulmonary and mesenteric arteries from endotoxemic rabbits: Modulation by vascular expressions of inducible nitric-oxide synthase and histamine H1-receptors. J. Pharmacol. Exp. Ther. 2003, 307, 175–181. [Google Scholar] [CrossRef]
- Hattori, Y.; Endou, M.; Gando, S.; Kanno, M. Identification and characterization of histamine H1- and H2-receptors in guinea-pig left atrial membranes by 3H-mepyramine and 3H-tiotidine binding. Br. J. Pharmacol. 1991, 103, 1573–1579. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, N.; Hattori, Y.; Jesmin, S.; Gando, S. Nuclear factor-kappaB decoy oligodeoxynucleotides prevent acute lung injury in mice with cecal ligation and puncture-induced sepsis. Mol. Pharmacol. 2005, 67, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimons, C.P.; Lazar-Molnar, E.; Tomoskozi, Z.; Buzás, E.; Rivera, E.S.; Falus, A. Histamine deficiency induces tissue-specific down-regulation of histamine H2 receptor expression in histidine decarboxylase knockout mice. FEBS Lett. 2001, 508, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Neumann, J.; Vahlensieck, U.; Boknik, P.; Linck, B.; Lüss, H.; Müller, F.U.; Matherne, G.P.; Schmitz, W. Functional studies in atrium overexpressing A1-adenosine receptors. Br. J. Pharmacol. 1999, 128, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- Isidori, A.M.; Cornacchione, M.; Barbagallo, F.; Di Grazia, A.; Barrios, F.; Fassina, L.; Monaco, L.; Giannetta, E.; Gianfrilli, D.; Garofalo, S.; et al. Inhibition of type 5 phosphodiesterase counteracts β2-adrenergic signalling in beating cardiomyocytes. Cardiovasc. Res. 2015, 106, 408–420. [Google Scholar] [CrossRef]
- Gergs, U.; Kirchhefer, U.; Bergmann, F.; Künstler, B.; Mißlinger, N.; Au, B.; Mahnkopf, M.; Wache, H.; Neumann, J. Characterization of Stressed Transgenic Mice Overexpressing H2-Histamine Receptors in the Heart. J. Pharmacol. Exp. Ther. 2020, 374, 479–488. [Google Scholar] [CrossRef]
- Gergs, U.; Weisgut, J.; Griethe, K.; Mißlinger, N.; Kirchhefer, U.; Neumann, J. Human histamine H2 receptors can initiate cardiac arrhythmias in a transgenic mouse. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2021, 394, 1963–1973. [Google Scholar] [CrossRef]
- Gergs, U.; Büxel, M.L.; Bresinsky, M.; Kirchhefer, U.; Fehse, C.; Höring, C.; Hofmann, B.; Marušáková, M.; Čináková, A.; Schwarz, R.; et al. Cardiac Effects of Novel Histamine H2 Receptor Agonists. J. Pharmacol. Exp. Ther. 2021, 379, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.J.; Thompson, C.R.; Walley, K.R.; Gillis, R.P.; Wolinski-Walley, P.E.; Schellenberg, R.R. Histamine decreases left ventricular contractility in normal human subjects. J. Appl. Physiol. (1985) 1992, 73, 2530–2537. [Google Scholar] [CrossRef]
- Zerkowski, H.R.; Broede, A.; Kunde, K.; Hillemann, S.; Schäfer, E.; Vogelsang, M.; Michel, M.C.; Brodde, O.E. Comparison of the positive inotropic effects of serotonin, histamine, angiotensin II, endothelin and isoprenaline in the isolated human right atrium. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1993, 347, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Kühn, B.; Schmid, A.; Harteneck, C.; Gudermann, T.; Schultz, G. G proteins of the Gq family couple the H2 histamine receptor to phospholipase C. Mol. Endocrinol. 1996, 10, 1697–1707. [Google Scholar] [CrossRef]
- Ferreira-Martins, J.; Rondon-Clavo, C.; Tugal, D.; Korn, J.A.; Rizzi, R.; Padin-Iruegas, M.E.; Ottolenghi, S.; de Angelis, A.; Urbanek, K.; Ide-Iwata, N.; et al. Spontaneous calcium oscillations regulate human cardiac progenitor cell growth. Circ. Res. 2009, 105, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Apáti, Á.; Pászty, K.; Hegedűs, L.; Kolacsek, O.; Orbán, T.I.; Erdei, Z.; Szebényi, K.; Péntek, A.; Enyedi, Á.; Sarkadi, B. Characterization of calcium signals in human embryonic stem cells and in their differentiated offspring by a stably integrated calcium indicator protein. Cell. Signal. 2013, 25, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.A.; Levi, R. Histamine and cardiac arrhythmias. Circ. Res. 1986, 58, 1–16. [Google Scholar] [CrossRef]
- Ginsburg, R.; Bristow, M.R.; Davis, K. Receptor mechanisms in the human epicardial coronary artery. Heterogeneous pharmacological response to histamine and carbachol. Circ. Res. 1984, 55, 416–421. [Google Scholar] [CrossRef]
- Ginsburg, R.; Bristow, M.R.; Davis, K.; Dibiase, A.; Billingham, M.E. Quantitative pharmacologic responses of normal and atherosclerotic isolated human epicardial coronary arteries. Circulation 1984, 69, 430–440. [Google Scholar] [CrossRef]
- Ginsburg, R.; Bristow, M.R.; Kantrowitz, N.; Baim, D.S.; Harrison, D.C. Histamine provocation of clinical coronary artery spasm: Implications concerning pathogenesis of variant angina pectoris. Am. Heart J. 1981, 102, 819–822. [Google Scholar] [CrossRef]
- Shimokawa, H.; Tomoike, H.; Nabeyama, S.; Yamamoto, H.; Araki, H.; Nakamura, M.; Ishii, Y.; Tanaka, K. Coronary artery spasm induced in atherosclerotic miniature swine. Science 1983, 221, 560–562. [Google Scholar] [CrossRef]
- Gokina, N.I.; Bevan, J.A. Histamine-induced depolarization: Ionic mechanisms and role in sustained contraction of rabbit cerebral arteries. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H2094–H2104. [Google Scholar] [CrossRef]
- Trzeciakowski, J.P.; Levi, R. Reduction of ventricular fibrillation threshold by histamine: Resolution into separate H1- and H2-mediated components. J. Pharmacol. Exp. Ther. 1982, 223, 774–783. [Google Scholar]
- Gaide, M.S.; Altman, C.B.; Cameron, J.S.; Kaiser, C.J.; Myerburg, R.J.; Bassett, A.L. Histamine modification of spontaneous rate and rhythm in infarcted canine ventricle. Agents Actions 1984, 15, 488–493. [Google Scholar] [CrossRef]
- Mugelli, A.; Mantelli, L.; Manzini, S.; Ledda, F. Induction by histamine of oscillatory activity in sheep Purkinje fibers and suppression by verapamil or lidocaine. J. Cardiovasc. Pharmacol. 1980, 2, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Thome, U.; Berger, F.; Borchard, U.; Hafner, D. Electrophysiological characterization of histamine receptor subtypes in sheep cardiac Purkinje fibers. Agents Actions 1992, 37, 30–38. [Google Scholar] [CrossRef]
- Matsuda, N.; Hattori, Y.; Sakuraya, F.; Kobayashi, M.; Zhang, X.-H.; Kemmotsu, O.; Gando, S. Hemodynamic significance of histamine synthesis and histamine H1- and H2-receptor gene expression during endotoxemia. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2002, 366, 513–521. [Google Scholar] [CrossRef]
- Grange, C.; Gurrieri, M.; Verta, R.; Fantozzi, R.; Pini, A.; Rosa, A.C. Histamine in the kidneys: What is its role in renal pathophysiology? Br. J. Pharmacol. 2020, 177, 503–515. [Google Scholar] [CrossRef]
- Leeper-Woodford, S.K.; Carey, D.; Byrne, K.; Walsh, C.; Fisher, B.; Sugerman, H.J.; Fowler, A.A. Histamine receptor antagonists, cyclooxygenase blockade, and tumor necrosis factor during acute septic insult. Shock 1998, 9, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Dimmeler, S.; Lechleuthner, A.; Auweiler, M.; Troost, C.; Nagelschmidt, M.; Neugebauer, E. Effect of H1-antagonism on cardiovascular, pulmonary, and immunological dysfunction in porcine endotoxic shock. Shock 1995, 3, 416–421. [Google Scholar] [PubMed]
- Stasiak, A.; Gola, J.; Kraszewska, K.; Mussur, M.; Kobos, J.; Mazurek, U.; Stark, H.; Fogel, W.A. Experimental autoimmune myocarditis in rats and therapeutic histamine H1–H4 receptor inhibition. J. Physiol. Pharmacol. 2018, 69, 889–900. [Google Scholar] [CrossRef]
- Ichikawa, A.; Sugimoto, Y.; Tanaka, S. Molecular biology of histidine decarboxylase and prostaglandin receptors. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 848–866. [Google Scholar] [CrossRef] [PubMed]
- Kaliner, M.; Sigler, R.; Summers, R.; Shelhamer, J.H. Effects of infused histamine: Analysis of the effects of H-1 and H-2 histamine receptor antagonists on cardiovascular and pulmonary responses. J. Allergy Clin. Immunol. 1981, 68, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Bani, D.; Nistri, S.; Mannaioni, P.F.; Masini, E. Cardiac anaphylaxis: Pathophysiology and therapeutic perspectives. Curr. Allergy Asthma Rep. 2006, 6, 14–19. [Google Scholar] [CrossRef]
- Hirasawa, N. Expression of Histidine Decarboxylase and Its Roles in Inflammation. Int. J. Mol. Sci. 2019, 20, 376. [Google Scholar] [CrossRef] [PubMed]
- Masini, E.; Vannacci, A.; Marzocca, C.; Mannaioni, P.F.; Befani, O.; Federico, R.; Toma, A.; Mondovì, B. A plant histaminase modulates cardiac anaphylactic response in guinea pig. Biochem. Biophys. Res. Commun. 2002, 296, 840–846. [Google Scholar] [CrossRef]
- Cottineau, C.; Drouet, M.; Costerousse, F.; Dussaussoy, C.; Sabbah, A. Intérêt des médiateurs plasmatiques (histamine et tryptase) et urinaire (méthylhistamine) lors des réactions anaphylactiques et/ou anaphylactoïdes peranesthésiques. Allerg. Immunol. 1996, 28, 270, 273–276. [Google Scholar]
- Renz, C.L.; Laroche, D.; Thurn, J.D.; Finn, H.A.; Lynch, J.P.; Thisted, R.; Moss, J. Tryptase levels are not increased during vancomycin-induced anaphylactoid reactions. Anesthesiology 1998, 89, 620–625. [Google Scholar] [CrossRef]
- Zdravkovic, V.; Pantovic, S.; Rosic, G.; Tomic-Lucic, A.; Zdravkovic, N.; Colic, M.; Obradovic, Z.; Rosic, M. Histamine blood concentration in ischemic heart disease patients. J. Biomed. Biotechnol. 2011, 2011, 315709. [Google Scholar] [CrossRef]
- Chen, J.; Hong, T.; Ding, S.; Deng, L.; Abudupataer, M.; Zhang, W.; Tong, M.; Jia, J.; Gong, H.; Zou, Y.; et al. Aggravated myocardial infarction-induced cardiac remodeling and heart failure in histamine-deficient mice. Sci. Rep. 2017, 7, 44007. [Google Scholar] [CrossRef]
- Dy, M.; Lebel, B.; Kamoun, P.; Hamburger, J. Histamine production during the anti-allograft response. Demonstration of a new lymphokine enhancing histamine synthesis. J. Exp. Med. 1981, 153, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Chen, B.; Zhao, Z.; He, N.; Zeng, Z.; Wu, B.; Fukushima, Y.; Dai, M.; Huang, Q.; Xu, D.; et al. Histamine H2 receptor activation exacerbates myocardial ischemia/reperfusion injury by disturbing mitochondrial and endothelial function. Basic Res. Cardiol. 2013, 108, 342. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.C.; Chang, J.K. Urinary histamine excretion in the rat following skin homografting and autografting. Ann. Surg. 1968, 167, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Ding, S.; Li, H.; Zhang, Z.; Xu, L.; Wu, J.; Wang, X.; Zou, Y.; Yang, X.; Ge, J. Disruption of histamine/H1R signaling pathway represses cardiac differentiation and maturation of human induced pluripotent stem cells. Stem Cell Res. Ther. 2020, 11, 27. [Google Scholar] [CrossRef]
- Hattori, M.; Mizuguchi, H.; Baba, Y.; Ono, S.; Nakano, T.; Zhang, Q.; Sasaki, Y.; Kobayashi, M.; Kitamura, Y.; Takeda, N.; et al. Quercetin inhibits transcriptional up-regulation of histamine H1 receptor via suppressing protein kinase C-δ/extracellular signal-regulated kinase/poly(ADP-ribose) polymerase-1 signaling pathway in HeLa cells. Int. Immunopharmacol. 2013, 15, 232–239. [Google Scholar] [CrossRef]
- Fukuda, S.; Midoro, K.; Yamasaki, M.; Gyoten, M.; Kawano, Y.; Fukui, H.; Ashida, Y.; Nagaya, H. Characteristics of the antihistamine effect of TAK-427, a novel imidazopyridazine derivative. Inflamm. Res. 2003, 52, 206–214. [Google Scholar] [CrossRef]
- Govoni, M.; Bakker, R.A.; van de Wetering, I.; Smit, M.J.; Menge, W.M.B.P.; Timmerman, H.; Elz, S.; Schunack, W.; Leurs, R. Synthesis and pharmacological identification of neutral histamine H1-receptor antagonists. J. Med. Chem. 2003, 46, 5812–5824. [Google Scholar] [CrossRef]
- Wittmann, H.-J.; Elz, S.; Seifert, R.; Straber, A. N (α)-Methylated phenylhistamines exhibit affinity to the hH(4)R-a pharmacological and molecular modelling study. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2011, 384, 287–299. [Google Scholar] [CrossRef]
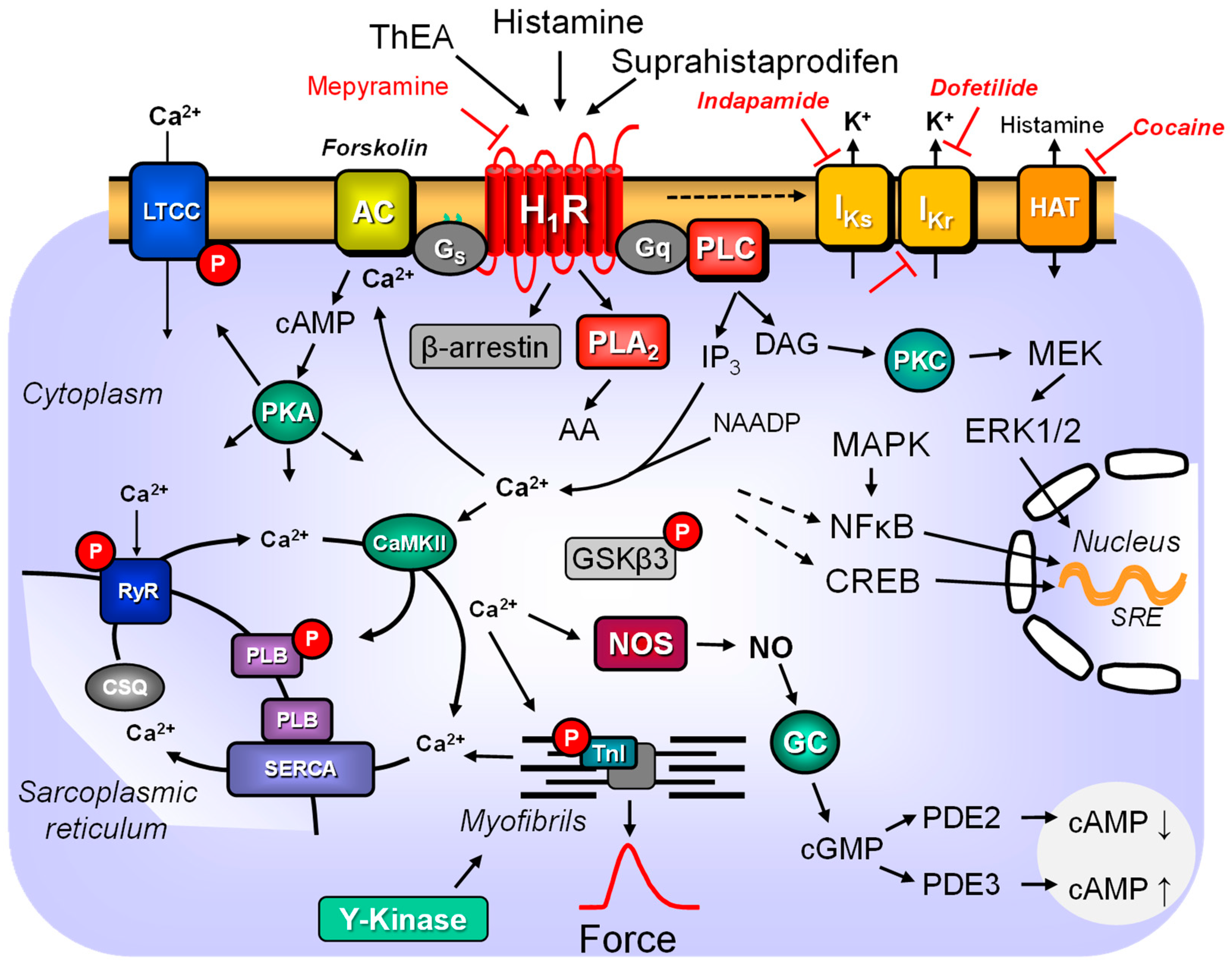
| Pathway | Inhibitor | Cell Type Studied | References |
|---|---|---|---|
| AC | SQ 22,536 | rat cardiomyocytes | Mittra and Bourreau 2006 [45] |
| Cam kinase II | KN-62 | mouse fetal cardiomyocytes | Okazaki et al., 1994 [46] |
| Cyclooxigenase | indomethacine | murine neuroblastoma cells | Snider et al., 1984 [47] |
| cGMP | R-cGMP-S | rabbit heart | Hattori et al., 1988 [44] |
| DG | R59022: increases DAG level | rat cardiomyocytes | Wientzek et al., 1997 [25] |
| Dynamin | dynasore | transfected U937 cells | Diaz Nebreda et al., 2019 [48] |
| EPAC | ESI-05 | human bronchial smooth muscle cells | Dale et al., 2018 [49] |
| Gi | 1 pertussis toxin, 2 guanosine 5′-O-(2-thiodiphosphate) | 1 CHO cells 2 fibroblasts | 1 Leurs et al., 1994 [50] 2 Burch and Axelrod 1987 [51] |
| Gq | 1 chromodepsin, 2 BIM-46187 | 1,2 various | 1 Hermes et al., 2021 [52] 2 Zhang et al., 2020 [53] |
| Guanylyl cyclases | H-(1,2,4)-oxadiazolo-(4,3-a)-quinoxalin-1-one (ODQ, 10 μM), methylene blue | rabbit atrium | Hattori et al., 1990 [54] |
| Lipoxigenase | nordihydroguaiaretic acid | murine neuroblastoma cells | Snider et al., 1984 [47] |
| Ikr | E-4031 | guinea pig left atrial cardiomyocytes | Matsumoto et al., 1999 [43] |
| Iks | indapamide | guinea pig left atrial cardiomyocytes | Matsumoto et al., 1999 [43] |
| IP3 | 1 2-aminoethoxy-diphenyl borate (2-APB), xestospongin B 5 µM | 1,2 rat cardiomyocytes | 1 Ibarra et al., 2004 [55] 2 Sankar et al., 2014 [56] |
| LTCC | nifedipine | guinea pig atrial cells | Matsumoto et al., 1999 [43] |
| PKC | 1 bisindolylmaleimide, 2 PMA, 3 rottlerin, 4 Ro-31-8220 | 1 T-cells 2,3,4 HeLa cells | 1 Iwata et al., 2005 [57] 2 Smit et al., 1992 [58] 3 Mizuguchi et al., 2012 [59] 4 Nariai et al., 2015 [60] |
| PLC | 1 U73112, 2 staurosporin, 2 calphostatin, 3 neomycin,3 2-nitro-4-carboxyphenyl-N,N-diphenylcarbamate, | 1 U937, 1 CHO cells 2 guinea pig atrial cells 3 Rabbit atrium | 1 Alonso et al., 2013 [39] 2 Matsumoto et al., 1999 [43] 3 Hattori et al., 1988 [44] |
| PLA2 | quinacrin | murine neuroblastoma cells | Snider et al., 1984 [47] |
| MAP kinases | SB203580 | 1 leukemic cell line 2 mouse cardiomyocytes | 1 Birkenkamp et al., 2000 [61] 2 Moise et al., 2010 [62] |
| MEK | U0126 | HeLa cells | Mizuguchi et al., 2011 [23] |
| MLC kinase | 1 ML-7, PIK | 1 rat heart 2 mouse heart | 1 Bil-Lula et al., 2018 [63] 2 Sun et al., 2021 [64] |
| NAADP | 1 PPADS 2 Ned 19 | 1 rat heart 2 mouse heart | 1 Pustovit et al., 2016 [65] 2 Davidson et al., 2015 [66] |
| NOS | L-NAME | rat heart | Kostić and Jakovljević 1996 [67] |
| RAF kinase | RAF kinase Inhibitor 1 | HeLa cells | Mizuguchi et al., 2011 [23] |
| RYR | ryanodine | 1 guinea pig atrium 2 mouse atrium | 1 Hattori et al., 1988 [44] 2 Stemmer and Akera 1986 [68] |
| PARP-1 | DPG | HeLa cells | Mizuguchi et al., 2011 [23] |
| PDE 2 | 1 EHNA, Bay 607550 | 1 mouse heart 2 rat cardiomyocytes | 1 Neumann et al., 2021 [69] 2 Castro et al., 2006 [70] |
| PDE 3 | 1 cilostamide, 2 milrinone | 1 mouse heart 2 human ventricle | 1 Neumann et al., 2021 [69], 2 Brown et al., 1986 [71] |
| PDE 4 | 1rolipram | 1 guinea pig heart 2 mouse heart | 1 Ukita et al., 1999 [72] 2 Neumann et al., 2021 [69] |
| PLD | 1 FIPI 10 nM | 2 mouse heart | 1 McDermott et al., 2020 [73] 2 Kim et al., 2007 [74] |
| PKA | 1 KT5720 2 Rp-8-CPT-cAMPS) 2 H89 2 PKI-myr | 1 U937, 1 CHO cells 2 human bronchial smooth muscle cells | 1 Alonso et al., 2013 [39] 2 Dale et al., 2018 [49] |
| PKG | 8-bromo cGMP | human aortic smooth muscle cells | Taylor et al., 2017 [75] |
| Protein kinase G | GF109203 20 μM | U937, CHO cells | Alonso et al., 2013 [39] |
| SERCA | 1,2 tharpsigargin, 1,3 cyclopiazonic acid | 1 human endothelial cells 2 rat cardiomyocytes 3 mouse cardiomyocytes | 1 Esposito et al., 2011 [76] 2 Rogers et al., 1995 [77] 3 Kemecsei et al., 2010 [78] |
| Store operated calcium ion channels | Ni2+ | guinea pig left atrium | Hattori and Kanno 1985 [79] |
| Tyrosine kinases | genistein | guinea pig left atrium | Akaishi et al., 2000 [80] |
| Species | Right Atrium | Left Atrium | Ventricle | References |
|---|---|---|---|---|
| Dog | 1,2 PCE, 1,2 PIE | 3 No effect 4 AV: negative dromotropic | 1 Chiba 1977 [84] 2 Chiba 1976 [85] 3 McNeill 1984 [86] 4 Motomura and Hashimoto 1989 [87] | |
| Guinea pig | No inotropic effect, 3 NCE, 9 NIE | 3,4,5,6,7 PIE, 6,7 PEA | 1 NIE: 1 Langendorff, muscle strips 2,6,8 PIE: muscle strips,2,6 PEA 10 AV: negative dromotropic | 1 Zavecz and Levi 1978 [88] 2 Mantelli et al., 1992 [89] 3 Reinhardt et al., 1974 [90] 4 Steinburg and Holland 1975 [91] 5 Reinhardt et al., 1977 [92] 6 Verma and McNeill 1977 [93] 7 Amerini et al., 1982 [94] 8 Hattori et al., 1994 [83] 9 Wilson and Broadley 1981, 1989 [95,96] 10 Levi and Kuye 1974 [97] |
| H1-TG | NCE | NIE, then PIE | Rayo-Abella et al., 2022 [98] | |
| Man | 1 PIE or 2 NIE, 4 NCE | not done | 3,4 NIE: muscle strips 4 AV: negative dromotropic | 1 Sanders et al., 1996 [99] 2 Guo et al., 1984 [100] 4 Genovese et al., 1988 [101] 3 Du et al., 1993 [102] |
| Mouse (wild type) | No effect | No effect | No effect | Gergs et al., 2019 [13] |
| Pig | No effect | No effect | No effect | Du et al., 1993 [102] |
| Rabbit | 2,5 No effect 7 PCE | 5 PIE 6 No effect | 1 NIE: Langendorff 2,3,4,5 PIE: muscle strips | 1 Coruzzi et al., 1979 [103] 2 Hattori et al., 1988 [104], 3 Hattori et al., 1990 [54], 4 Hattori et al., 1994 [83] 5 Verma and McNeill 1977 [93] 6 Hattori et al., 1991 [82] 7 McNeill 1984 [86] |
| Rat | No effect | No effect | No effect | Laher and McNeill 1980 [105] |
| Turtle | PIE | Kinawa and Tasaka 1989 [106] |
| References | Tissue | Measured Parameters | Agonists and Antagonists Used | Age, Gender | Disease | Medication |
|---|---|---|---|---|---|---|
| Du et al., 1993 [102] | human atrial preparations | isometric force in atrial and ventricular preparations paced at 1 Hz | histamine (1–1000 µM), mepyramine (1 µM), cimetidine (10 µM), propranolol (1 µM), norepinephrine (1–10 µM) | 5 males, 2 females, age 11–42 years | healthy organ donors, death from polytrauma | no drugs, only organ preserving buffer |
| Genovese et al., 1988 [101] | human right atrial appendage, human ventricular papillary muscles | isometric force in spontaneously beating atrial preparations or paced at 1 Hz, papillary muscle strips paced at 1 Hz, Tyrode solution | histamine (1 µM, 100 µM), pyrilamine (1 µM), adenosine 0.1 µM–100 µM), N6-cyclo-pentyladenosine (1 nM–10 µM), carbachol (20 nM) | not reported | corrective cardiac surgery, no heart failure | no cardiotonic drug, no anti-arrhythmic, no diuretics |
| Guo et al., 1984 [100] | human right atrial appendage | isometric force in spontaneous-ly beating atrial preparations or paced at 1 Hz, Tyrode solution | histamine (0.1–100 µM, pyrilamine (1 µM), cimetidine (3 µM), ThEA (0.1–300 µM), impromidine (0.1–100 µM), pindolol (1 nM), norepinephrine (0.1–10 µM) | not reported | bypass surgery, no heart failure | No cardiotonic drugs, no anti-arrhythmics, no diuretics |
| Sanders et al., 1996 [99] | human right atrial appendage | Isometric force in atrial preparations paced at 1 Hz or 0.5 Hz or 0.2 Hz, Krebs solution with fumarate, pyruvate, glutamate, glucose | histamine (0.1 µM–1 mM), famotidine (0.1 µM, 30 µM), sodium nitroprusside 10 µM, mepyramine (1 µM), CGP 20712A (300 nM), | 71 males, 18 females, mean age: 60 years | coronary artery disease, aortic mitral valve disease, mitral valve disease, no terminal heart failure | β-adrenoceptor antagonists (57 patients), no cimetidine or ranitidine, L-type calcium channel blockers, diuretics, nitrates, ACE-inhibitors, antibiotics, allopurinol, aminophylline, amiodarone, aspirin, amitryptiline, corticoids, bezafibrate, carbimazole, diazepam, analgetics, antidiabetics, pravastatin, simvastatin, prazosin, salbutamol, triazolam, warfarin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neumann, J.; Hofmann, B.; Kirchhefer, U.; Dhein, S.; Gergs, U. Function and Role of Histamine H1 Receptor in the Mammalian Heart. Pharmaceuticals 2023, 16, 734. https://doi.org/10.3390/ph16050734
Neumann J, Hofmann B, Kirchhefer U, Dhein S, Gergs U. Function and Role of Histamine H1 Receptor in the Mammalian Heart. Pharmaceuticals. 2023; 16(5):734. https://doi.org/10.3390/ph16050734
Chicago/Turabian StyleNeumann, Joachim, Britt Hofmann, Uwe Kirchhefer, Stefan Dhein, and Ulrich Gergs. 2023. "Function and Role of Histamine H1 Receptor in the Mammalian Heart" Pharmaceuticals 16, no. 5: 734. https://doi.org/10.3390/ph16050734
APA StyleNeumann, J., Hofmann, B., Kirchhefer, U., Dhein, S., & Gergs, U. (2023). Function and Role of Histamine H1 Receptor in the Mammalian Heart. Pharmaceuticals, 16(5), 734. https://doi.org/10.3390/ph16050734