
NMDA Receptor Glycine Binding Site Modulators for Prevention and Treatment of Ketamine Use Disorder

Abstract
1. Introduction
2. Results
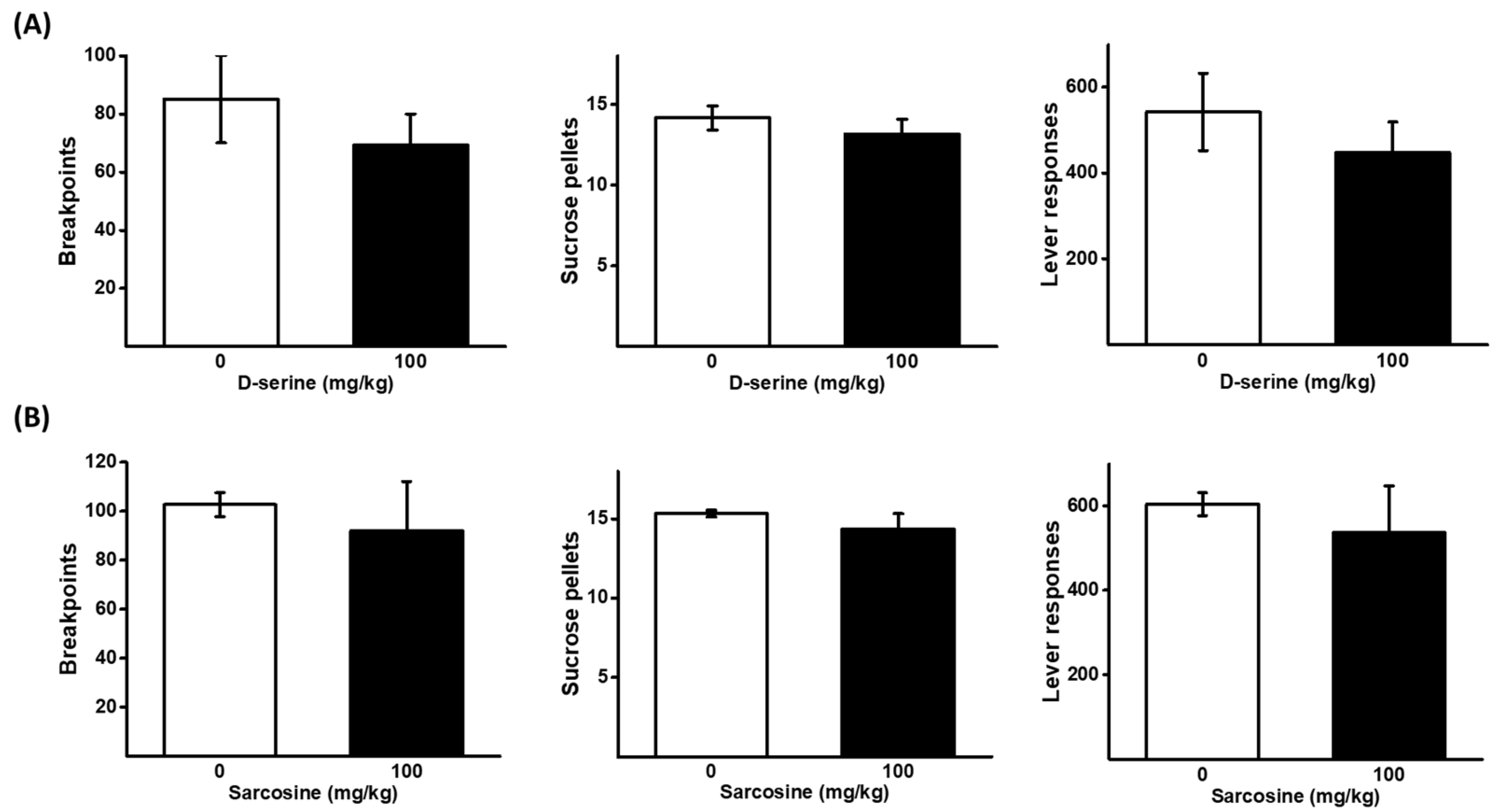
2.1. Effects of D-Serine on Ketamine Self-Administration Using a Progressive Ratio (PR) Schedule
2.2. Effects of Sarcosine on Ketamine Self-Administration Using a Progressive Ratio (PR) Schedule
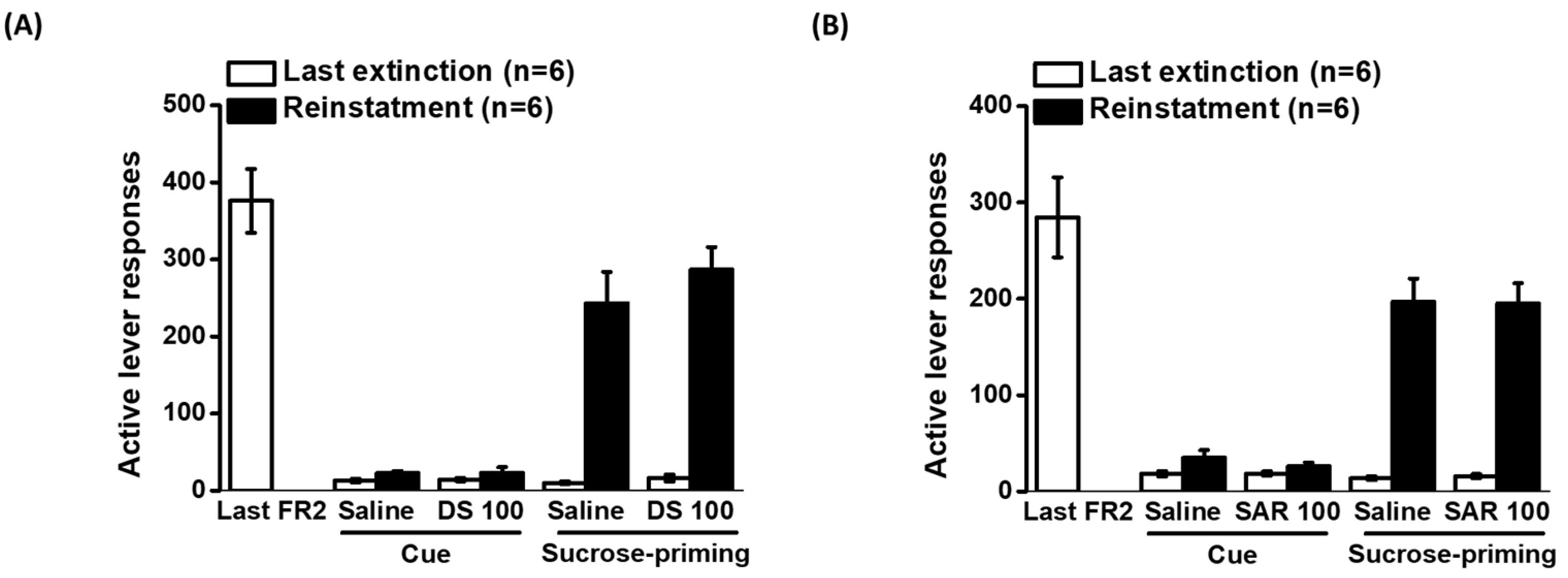
2.3. Effects of D-Serine on Reinstatement of Ketamine Seeking in Response to Cues and Drug Exposure
2.4. Effects of Sarcosine on Reinstatement of Ketamine Seeking in Response to Cues and Drug Exposure
2.5. Effects of D-Serine and Sarcosine on Sucrose Self-Administration under a PR Schedule
2.6. Effects of D-Serine and Sarcosine on Cue- and Sucrose-Induced Reinstatement of Sucrose Seeking
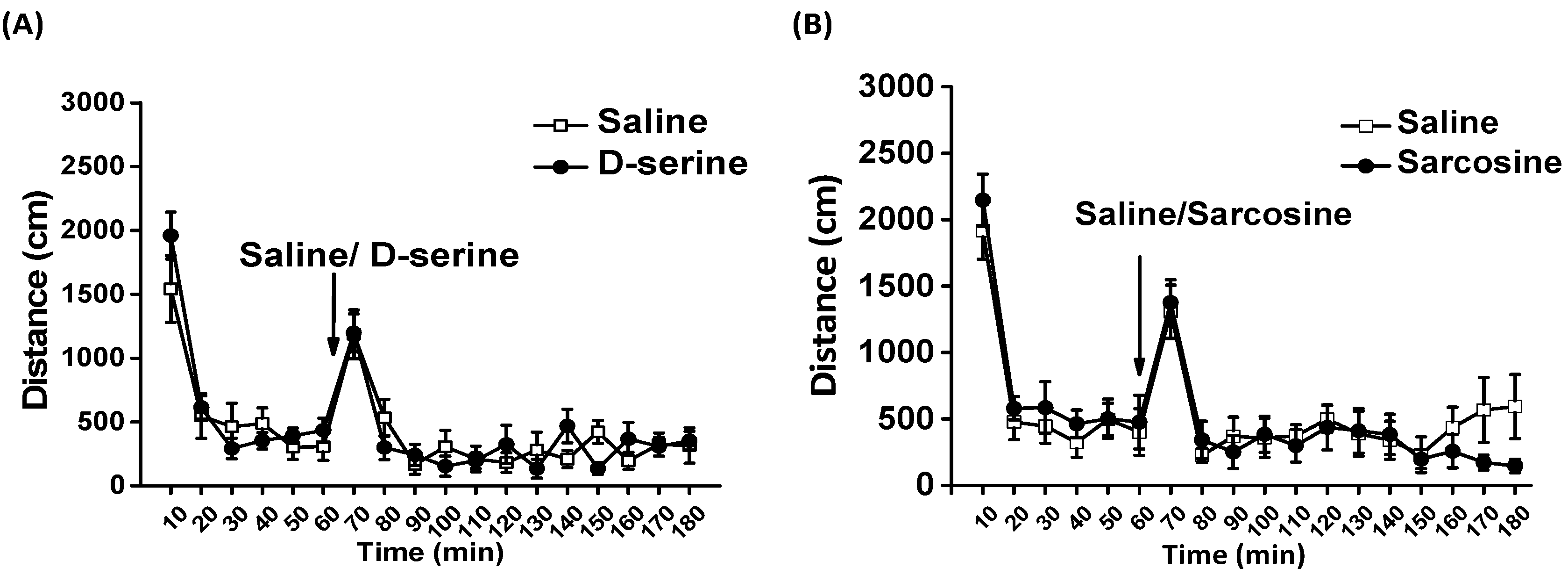
2.7. Effects of D-Serine and Sarcosine on Locomotor Activity
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Ketamine Self-Administration Procedures
4.4. A Progressive Ratio (PR) Schedule of Reinforcement
4.5. Cue- and Drug Priming-Induced Reinstatement of Ketamine Seeking
4.6. Sucrose Self-Administration Procedures
4.7. Open Filed Test
4.8. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Du, R.; Han, R.; Niu, K.; Xu, J.; Zhao, Z.; Lu, G.; Shang, Y. The Multivariate Effect of Ketamine on PTSD: Systematic Review and Meta-Analysis. Front. Psychiatry 2022, 13, 813103. [Google Scholar] [CrossRef] [PubMed]
- Tully, J.L.; Dahlen, A.D.; Haggarty, C.J.; Schioth, H.B.; Brooks, S. Ketamine treatment for refractory anxiety: A systematic review. Br. J. Clin. Pharmacol. 2022, 88, 4412–4426. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, E.; Dadabayev, A.R.; Joshi, S.A.; Glue, P. Systematic review and meta-analysis of randomized controlled trials of ketamine in the treatment of refractory anxiety spectrum disorders. Ther. Adv. Psychopharmacol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.L.; Mateus, C.F.; Malcolm, R.J.; Brady, K.T.; Back, S.E. Efficacy of Ketamine in the Treatment of Substance Use Disorders: A Systematic Review. Front. Psychiatry 2018, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Collins, R.J.; Weeks, J.R.; Cooper, M.M.; Good, P.I.; Russell, R.R. Prediction of abuse liability of drugs using IV self-administration by rats. Psychopharmacology 1984, 82, 6–13. [Google Scholar] [CrossRef] [PubMed]
- De Luca, M.T.; Badiani, A. Ketamine self-administration in the rat: Evidence for a critical role of setting. Psychopharmacology 2011, 214, 549–556. [Google Scholar] [CrossRef]
- De Luca, M.T.; Meringolo, M.; Spagnolo, P.A.; Badiani, A. The role of setting for ketamine abuse: Clinical and preclinical evidence. Rev. Neurosci. 2012, 23, 769–780. [Google Scholar] [CrossRef]
- Suzuki, T.; Kato, H.; Aoki, T.; Tsuda, M.; Narita, M.; Misawa, M. Effects of the non-competitive NMDA receptor antagonist ketamine on morphine-induced place preference in mice. Life Sci. 2000, 67, 383–389. [Google Scholar] [CrossRef]
- Li, F.; Fang, Q.; Liu, Y.; Zhao, M.; Li, D.; Wang, J.; Lu, L. Cannabinoid CB(1) receptor antagonist rimonabant attenuates reinstatement of ketamine conditioned place preference in rats. Eur. J. Pharmacol. 2008, 589, 122–126. [Google Scholar] [CrossRef]
- Liao, Y.; Tang, Y.L.; Hao, W. Ketamine and international regulations. Am. J. Drug. Alcohol. Abuse 2017, 43, 495–504. [Google Scholar] [CrossRef]
- Carlezon, W.A., Jr.; Wise, R.A. Rewarding actions of phencyclidine and related drugs in nucleus accumbens shell and frontal cortex. J. Neurosci. 1996, 16, 3112–3122. [Google Scholar] [CrossRef] [PubMed]
- Contreras, P.C. D-serine antagonized phencyclidine- and MK-801-induced stereotyped behavior and ataxia. Neuropharmacology 1990, 29, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Kanahara, N.; Shimizu, E.; Ohgake, S.; Fujita, Y.; Kohno, M.; Hashimoto, T.; Matsuzawa, D.; Shirayama, Y.; Hashimoto, K.; Iyo, M. Glycine and D: -serine, but not D: -cycloserine, attenuate prepulse inhibition deficits induced by NMDA receptor antagonist MK-801. Psychopharmacology 2008, 198, 363–374. [Google Scholar] [CrossRef]
- Karasawa, J.I.; Hashimoto, K.; Chaki, S. D-Serine and a glycine transporter inhibitor improve MK-801-induced cognitive deficits in a novel object recognition test in rats. Behav. Brain Res. 2008, 186, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Hu, J.; Liu, Y. Ketamine inhibits colorectal cancer cells malignant potential via blockage of NMDA receptor. Exp. Mol. Pathol. 2019, 107, 171–178. [Google Scholar] [CrossRef]
- D’Ascenzo, M.; Podda, M.V.; Grassi, C. The role of D-serine as co-agonist of NMDA receptors in the nucleus accumbens: Relevance to cocaine addiction. Front. Synaptic Neurosci. 2014, 6, 16. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, R.; Guo, L.; Zhen, X.C. Morphine-induced inhibition of Ca2+-dependent D-serine release from astrocytes suppresses excitability of GABAergic neurons in the nucleus accumbens. Addict. Biol. 2017, 22, 1289–1303. [Google Scholar] [CrossRef]
- Seif, T.; Simms, J.A.; Lei, K.; Wegner, S.; Bonci, A.; Messing, R.O.; Hopf, F.W. D-Serine and D-Cycloserine Reduce Compulsive Alcohol Intake in Rats. Neuropsychopharmacology 2015, 40, 2357–2367. [Google Scholar] [CrossRef]
- Zhang, H.X.; Hyrc, K.; Thio, L.L. The glycine transport inhibitor sarcosine is an NMDA receptor co-agonist that differs from glycine. J. Physiol 2009, 587, 3207–3220. [Google Scholar] [CrossRef]
- Yang, S.Y.; Hong, C.J.; Huang, Y.H.; Tsai, S.J. The effects of glycine transporter I inhibitor, N-methylglycine (sarcosine), on ketamine-induced alterations in sensorimotor gating and regional brain c-Fos expression in rats. Neurosci. Lett. 2010, 469, 127–130. [Google Scholar] [CrossRef]
- Pei, J.C.; Hung, W.L.; Lin, B.X.; Shih, M.H.; Lu, L.Y.; Luo, D.Z.; Tai, H.C.; Studer, V.; Min, M.Y.; Lai, W.S. Therapeutic potential and underlying mechanism of sarcosine (N-methylglycine) in N-methyl-D-aspartate (NMDA) receptor hypofunction models of schizophrenia. J. Psychopharmacol. 2019, 33, 1288–1302. [Google Scholar] [CrossRef]
- Ferreira, J.S.; Papouin, T.; Ladepeche, L.; Yao, A.; Langlais, V.C.; Bouchet, D.; Dulong, J.; Mothet, J.P.; Sacchi, S.; Pollegioni, L.; et al. Co-agonists differentially tune GluN2B-NMDA receptor trafficking at hippocampal synapses. Elife 2017, 6, e25492. [Google Scholar] [CrossRef]
- Nong, Y.; Huang, Y.Q.; Ju, W.; Kalia, L.V.; Ahmadian, G.; Wang, Y.T.; Salter, M.W. Glycine binding primes NMDA receptor internalization. Nature 2003, 422, 302–307. [Google Scholar] [CrossRef]
- Witkin, J.M.; Monn, J.A.; Schoepp, D.D.; Li, X.; Overshiner, C.; Mitchell, S.N.; Carter, G.; Johnson, B.; Rasmussen, K.; Rorick-Kehn, L.M. The Rapidly Acting Antidepressant Ketamine and the mGlu2/3 Receptor Antagonist LY341495 Rapidly Engage Dopaminergic Mood Circuits. J. Pharmacol. Exp. Ther. 2016, 358, 71–82. [Google Scholar] [CrossRef]
- Simmler, L.D.; Li, Y.; Hadjas, L.C.; Hiver, A.; van Zessen, R.; Luscher, C. Dual action of ketamine confines addiction liability. Nature 2022, 608, 368–373. [Google Scholar] [CrossRef]
- Caffino, L.; Piva, A.; Mottarlini, F.; Di Chio, M.; Giannotti, G.; Chiamulera, C.; Fumagalli, F. Ketamine Self-Administration Elevates alpha CaMKII Autophosphorylation in Mood and Reward-Related Brain Regions in Rats. Mol. Neurobiol. 2018, 55, 5453–5461. [Google Scholar] [CrossRef]
- Li, H.B.; Rajani, V.; Han, L.; Chung, D.; Cooke, J.E.; Sengar, A.S.; Salter, M.W. Alternative splicing of GluN1 gates glycine site-dependent nonionotropic signaling by NMDAR receptors. Proc. Natl. Acad. Sci. USA 2021, 118, e2026411118. [Google Scholar] [CrossRef]
- Chen, K.T.; Wu, C.H.; Tsai, M.H.; Wu, Y.C.; Jou, M.J.; Huang, C.C.; Wei, I.H. Antidepressant-like effects of long-term sarcosine treatment in rats with or without chronic unpredictable stress. Behav. Brain Res. 2017, 316, 1–10. [Google Scholar] [CrossRef]
- Chen, Z.; Tang, Z.; Zou, K.; Huang, Z.; Liu, L.; Yang, Y.; Wang, W. D-Serine produces antidepressant-like effects in mice through suppression of BDNF signaling pathway and regulation of synaptic adaptations in the nucleus accumbens. Mol. Med. 2021, 27, 127. [Google Scholar] [CrossRef]
- Huang, C.C.; Wei, I.H.; Huang, C.L.; Chen, K.T.; Tsai, M.H.; Tsai, P.; Tun, R.; Huang, K.H.; Chang, Y.C.; Lane, H.Y.; et al. Inhibition of glycine transporter-I as a novel mechanism for the treatment of depression. Biol. Psychiatry 2013, 74, 734–741. [Google Scholar] [CrossRef]
- Yang, F.Y.; Lee, Y.S.; Cherng, C.G.; Cheng, L.Y.; Chang, W.T.; Chuang, J.Y.; Kao, G.S.; Yu, L. D-cycloserine, sarcosine and D-serine diminish the expression of cocaine-induced conditioned place preference. J. Psychopharmacol. 2013, 27, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Radke, A.K.; Sneddon, E.A.; Monroe, S.C. Studying Sex Differences in Rodent Models of Addictive Behavior. Curr. Protoc. 2021, 1, e119. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.N.; Strong, C.E.; Addonizio, M.N.; Brownstein, N.C.; Kabbaj, M. Reinforcing properties of an intermittent, low dose of ketamine in rats: Effects of sex and cycle. Psychopharmacology 2017, 234, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Takarada, T.; Takahata, Y.; Iemata, M.; Hinoi, E.; Uno, K.; Hirai, T.; Yamamoto, T.; Yoneda, Y. Interference With Cellular Differentiation by D-Serine Through Antagonism at N-Methyl-D-Aspartate Receptors Composed of NR1 and NR3A Subunits in Chondrocytes. J. Cell. Physiol. 2009, 220, 756–764. [Google Scholar] [CrossRef]
- Nasyrova, R.F.; Khasanova, A.K.; Altynbekov, K.S.; Asadullin, A.R.; Markina, E.A.; Gayduk, A.J.; Shipulin, G.A.; Petrova, M.M.; Shnayder, N.A. The Role of D-Serine and D-Aspartate in the Pathogenesis and Therapy of Treatment-Resistant Schizophrenia. Nutrients 2022, 14, 5142. [Google Scholar] [CrossRef]
- Avellar, M.; Scoriels, L.; Madeira, C.; Vargas-Lopes, C.; Marques, P.; Dantas, C.; Manhaes, A.C.; Leite, H.; Panizzutti, R. The effect of D-serine administration on cognition and mood in older adults. Oncotarget 2016, 7, 11881–11888. [Google Scholar] [CrossRef]
- Nava-Gomez, L.; Calero-Vargas, I.; Higinio-Rodriguez, F.; Vazquez-Prieto, B.; Olivares-Moreno, R.; Ortiz-Retana, J.; Aranda, P.; Hernandez-Chan, N.; Rojas-Piloni, G.; Alcauter, S.; et al. Aging-Associated Cognitive Decline is Reversed by D-Serine Supplementation. Eneuro 2022, 9, ENEURO.0176-22.2022. [Google Scholar] [CrossRef]
- Wegner, S.A.; Hu, B.; De Oliveira Sergio, T.; Darevsky, D.; Kwok, C.C.; Lei, K.; Hopf, F.W. A novel NMDA receptor-based intervention to suppress compulsion-like alcohol drinking. Neuropharmacology 2019, 157, 107681. [Google Scholar] [CrossRef]
- Chang, C.H.; Lin, C.H.; Liu, C.Y.; Chen, S.J.; Lane, H.Y. Efficacy and cognitive effect of sarcosine (N-methylglycine) in patients with schizophrenia: A systematic review and meta-analysis of double-blind randomised controlled trials. J. Psychopharmacol. 2020, 34, 495–505. [Google Scholar] [CrossRef]
- Kumar, A.; Akhtar, A.; Kuhad, A.; Sah, S.P. Sarcosine (glycine transporter inhibitor) attenuates behavioural and biochemical changes induced by ketamine, in the rat model of schizophrenia. Exp. Brain Res. 2023, 241, 451–467. [Google Scholar] [CrossRef]
- Rajagopal, L.; Burgdorf, J.S.; Moskal, J.R.; Meltzer, H.Y. GLYX-13 (rapastinel) ameliorates subchronic phencyclidine- and ketamine-induced declarative memory deficits in mice. Behav. Brain Res. 2016, 299, 105–110. [Google Scholar] [CrossRef]
- Lee, M.Y.; Hsiao, Y.C.; Chan, M.H.; Chen, H.H. Lamotrigine attenuates the motivation to self-administer ketamine and prevents cue- and prime-induced reinstatement of ketamine-seeking behavior in rats. Drug. Alcohol. Depend. 2019, 194, 257–263. [Google Scholar] [CrossRef]
- Richardson, N.R.; Roberts, D.C. Progressive ratio schedules in drug self-administration studies in rats: A method to evaluate reinforcing efficacy. J. Neurosci. Methods 1996, 66, 1–11. [Google Scholar] [CrossRef]
- Pernot, P.; Maucler, C.; Tholance, Y.; Vasylieva, N.; Debilly, G.; Pollegioni, L.; Cespuglio, R.; Marinesco, S. D-Serine diffusion through the blood-brain barrier: Effect on D-serine compartmentalization and storage. Neurochem. Int. 2012, 60, 837–845. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Self-Administration | Locomotor Activity | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ketamine | Food | |||||||||
| Procedure | PR | Reinstatement | PR | Reinstatement | ||||||
| D-serine | Sarcosine | D-serine | Sarcosine | D-serine | Sarcosine | D-serine | Sarcosine | D-serine | Sarcosine | |
| Numbers | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 12 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsiao, Y.-C.; Lee, M.-Y.; Chan, M.-H.; Chen, H.-H. NMDA Receptor Glycine Binding Site Modulators for Prevention and Treatment of Ketamine Use Disorder. Pharmaceuticals 2023, 16, 812. https://doi.org/10.3390/ph16060812
Hsiao Y-C, Lee M-Y, Chan M-H, Chen H-H. NMDA Receptor Glycine Binding Site Modulators for Prevention and Treatment of Ketamine Use Disorder. Pharmaceuticals. 2023; 16(6):812. https://doi.org/10.3390/ph16060812
Chicago/Turabian StyleHsiao, Yu-Chin, Mei-Yi Lee, Ming-Huan Chan, and Hwei-Hsien Chen. 2023. "NMDA Receptor Glycine Binding Site Modulators for Prevention and Treatment of Ketamine Use Disorder" Pharmaceuticals 16, no. 6: 812. https://doi.org/10.3390/ph16060812
APA StyleHsiao, Y.-C., Lee, M.-Y., Chan, M.-H., & Chen, H.-H. (2023). NMDA Receptor Glycine Binding Site Modulators for Prevention and Treatment of Ketamine Use Disorder. Pharmaceuticals, 16(6), 812. https://doi.org/10.3390/ph16060812