

Exploring the Mechanism of Sempervirine Inhibiting Glioblastoma Invasion Based on Network Pharmacology and Bioinformatics

Abstract
:1. Introduction
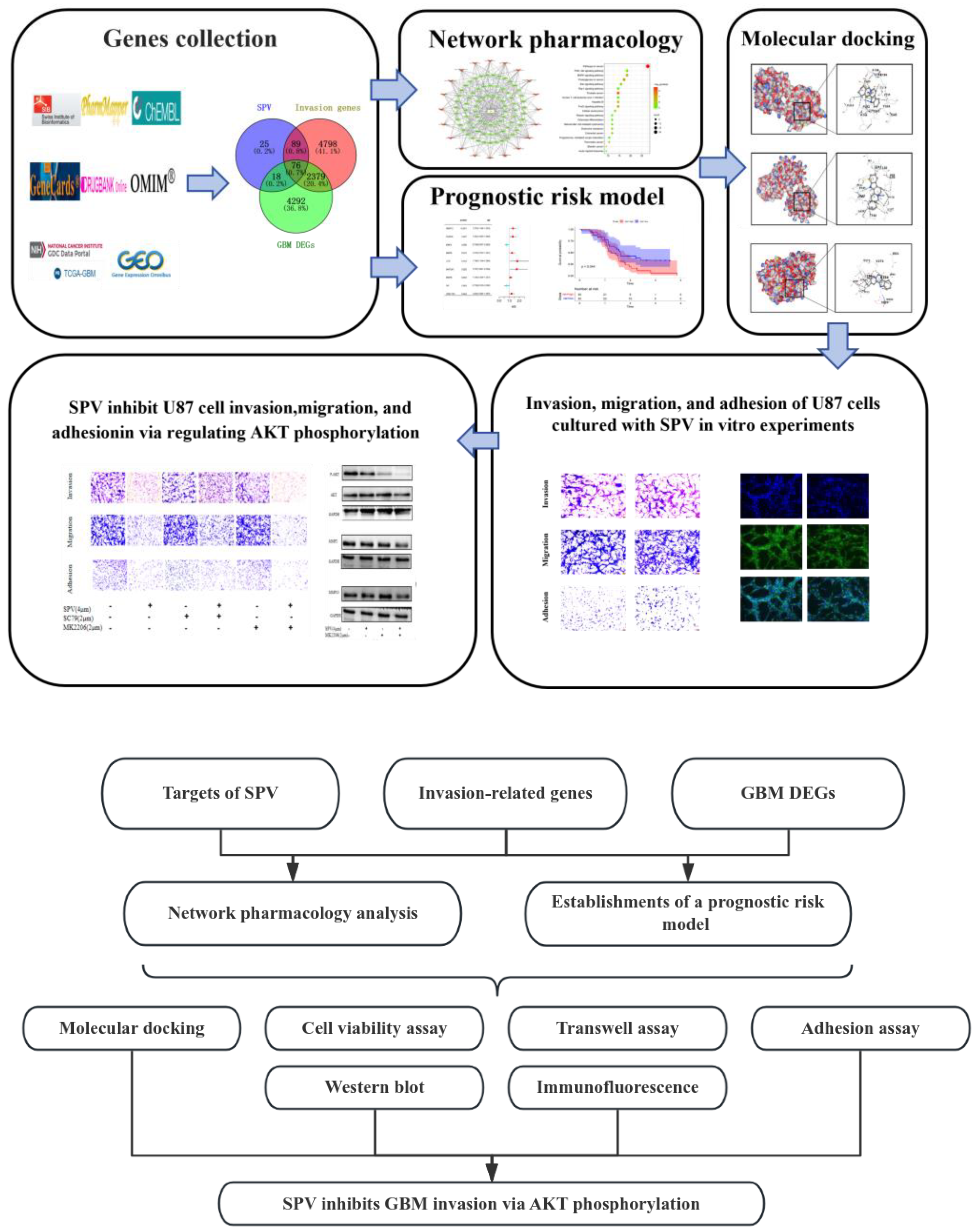
2. Results
2.1. Identifying the Targets and Pathways of SIGI
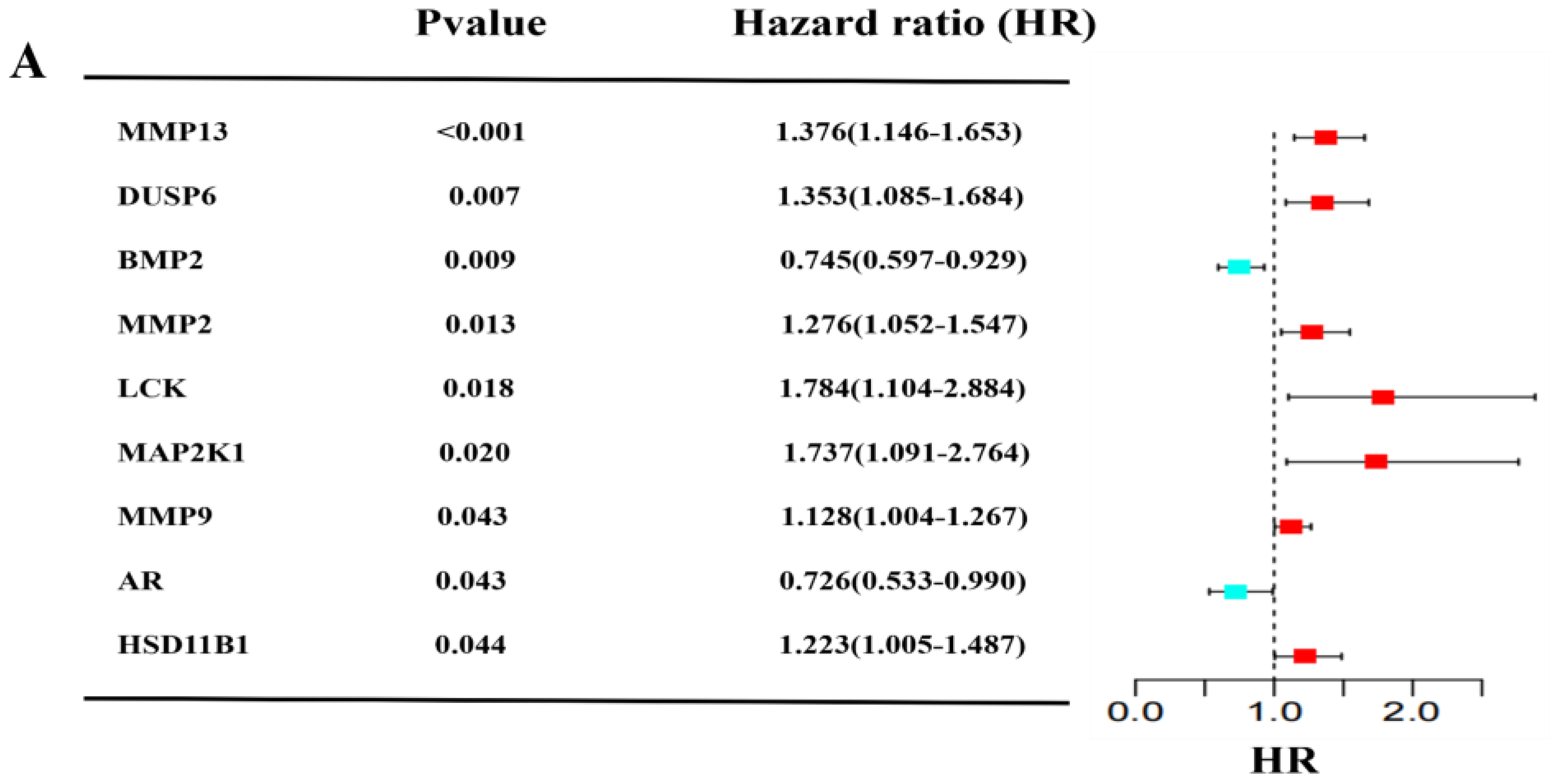
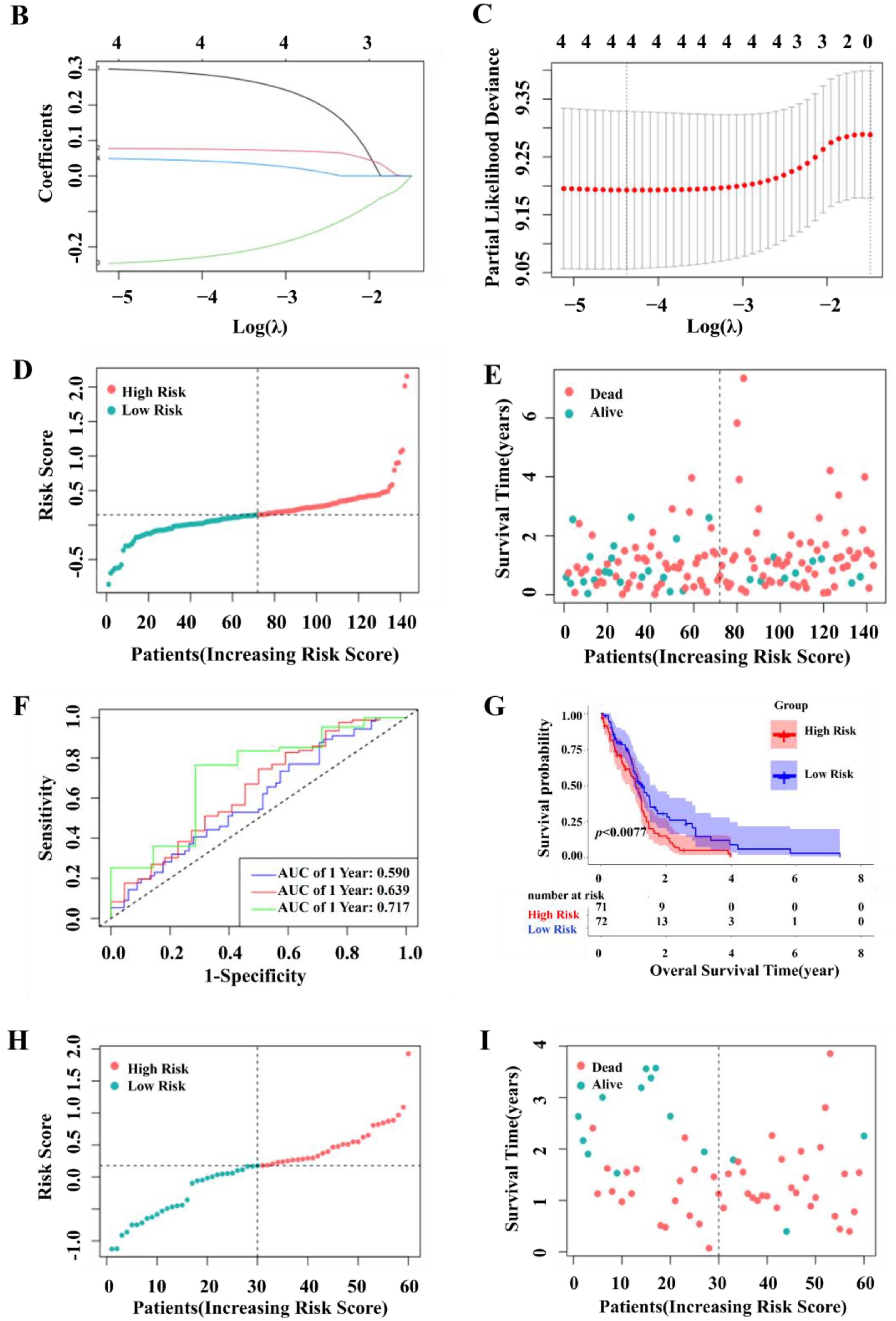
2.2. Construction and Validation of a Prognostic Model for Patients with GBM in the TCGA and GEO Cohort
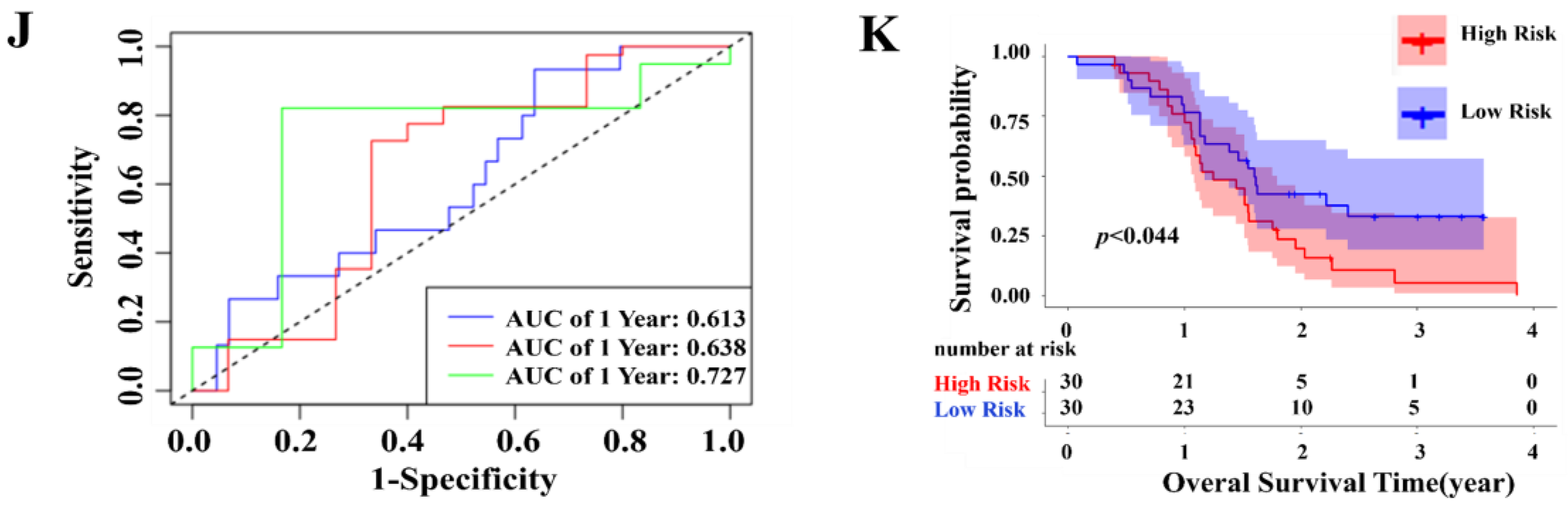
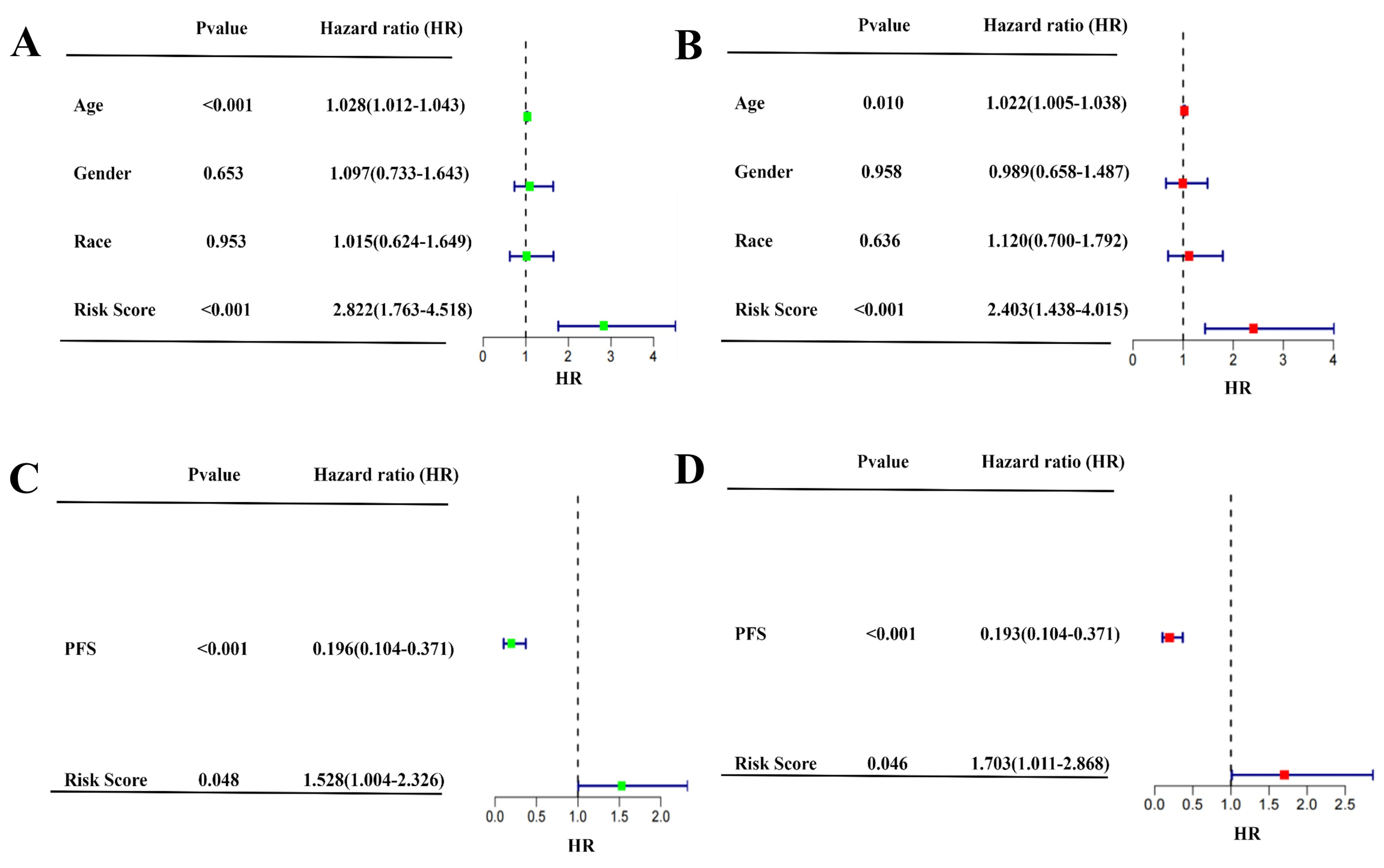
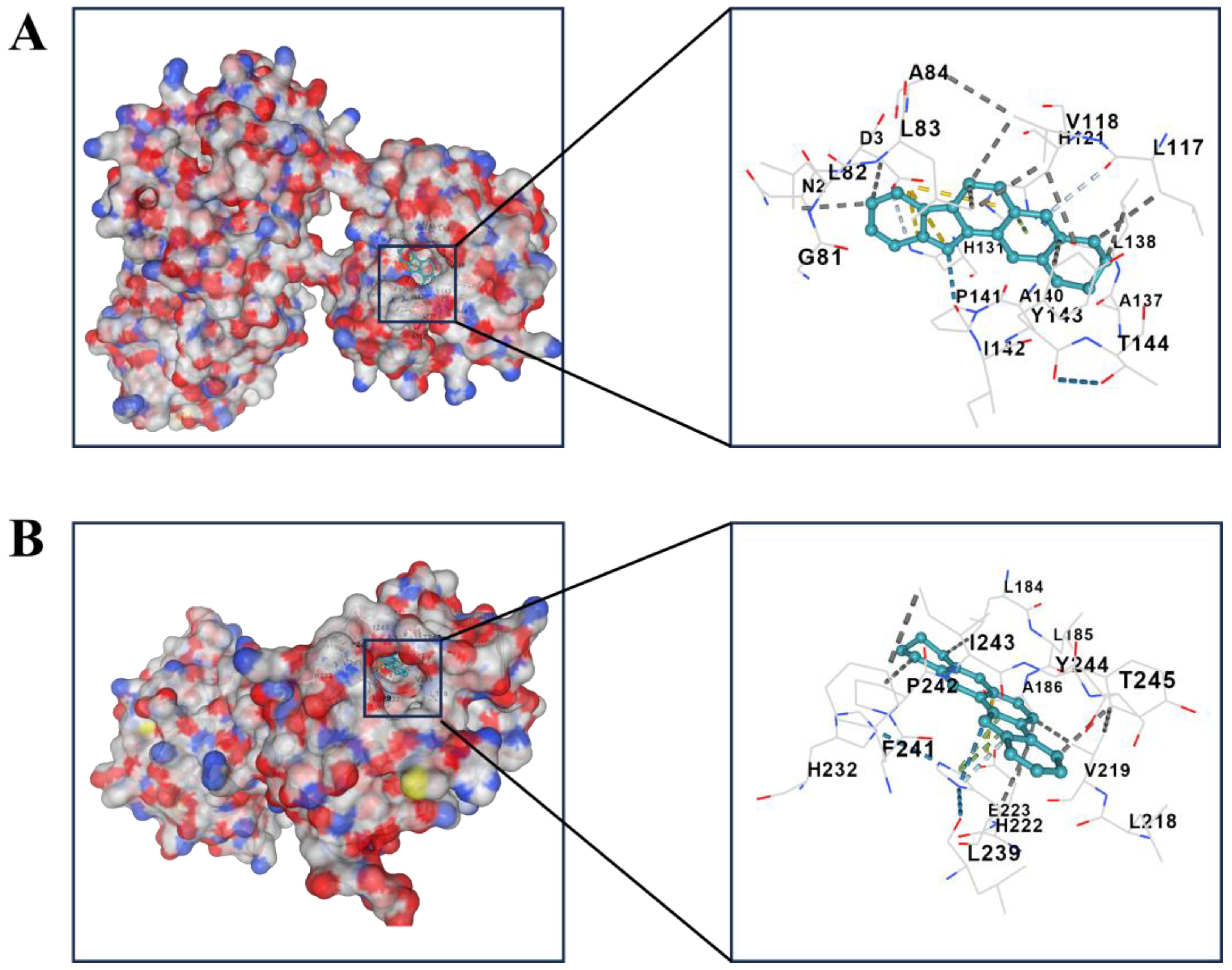
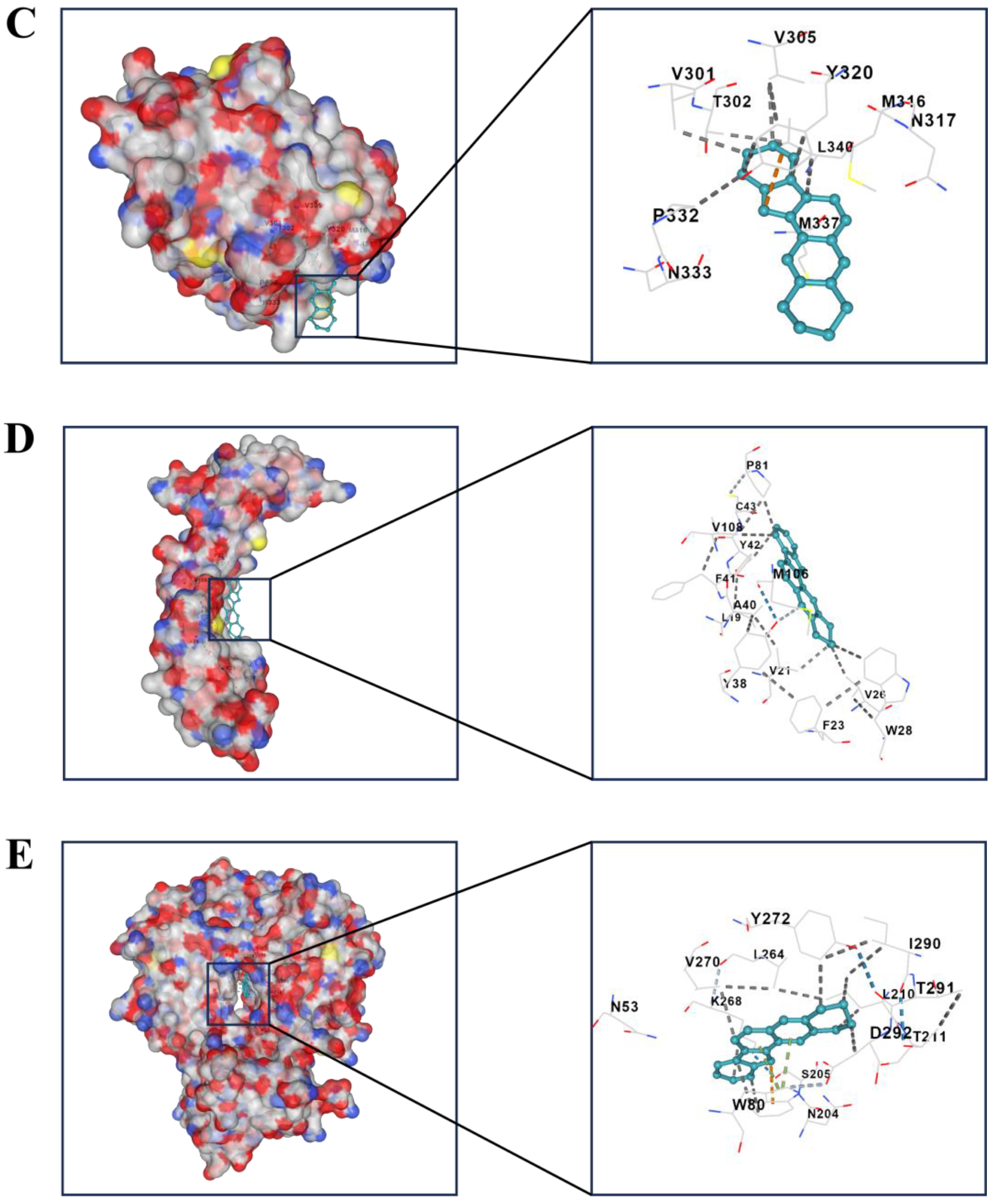
2.3. Independent Prognostic Value of the Four-Gene Prognostic Model and Molecular Docking
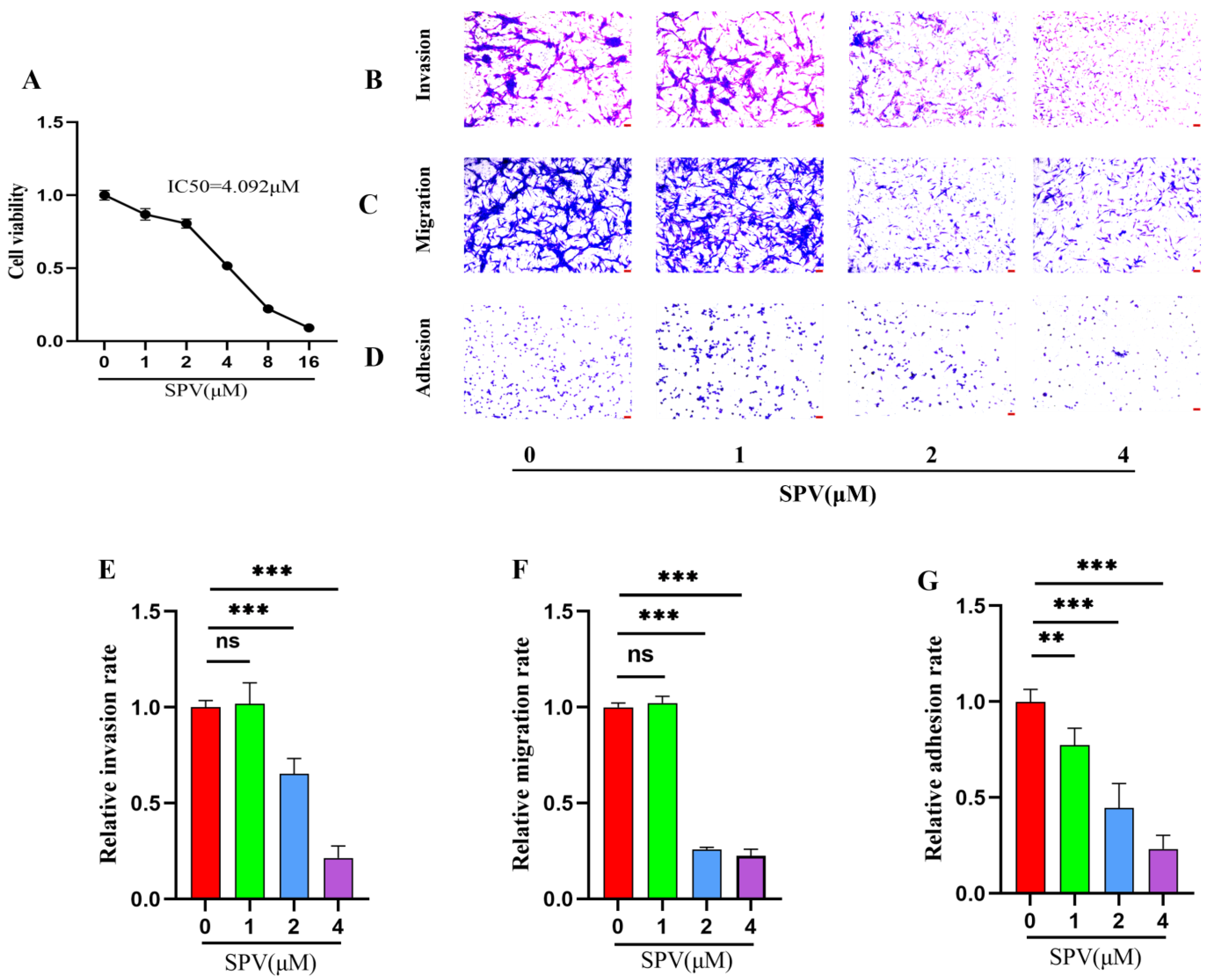
2.4. SPV Efficiently Inhibits Invasion, Migration, and Adhesion on GBM Cells
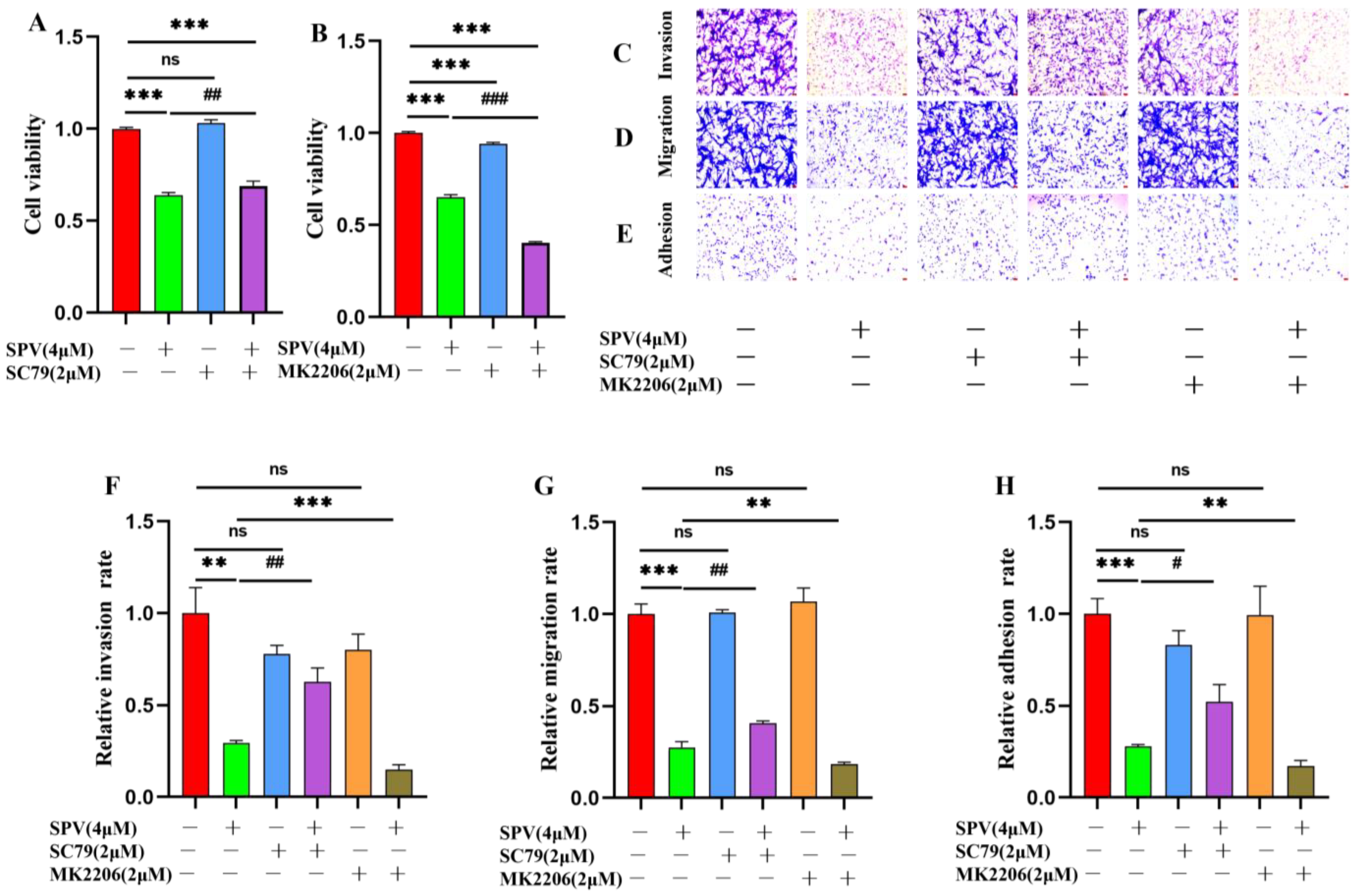
2.5. Regulation of AKT Signaling Affects Invasion, Migration and Adhesion in GBM Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Datasets and Genes Participating in GBM
4.4. Collection of Invasion and SPV-Related Target Genes
4.5. GO, KEGG Pathway Enrichment Analysis, and Compound-Targets-Pathway Network Construction
4.6. Constructing and Validating a Prognostic Gene Signature for the Inhibition of SPV on GBM Invasion
4.7. Evaluation of the Gene Signature Accuracy
4.8. Molecular Docking
4.9. Cell Viability Assay
4.10. Transwell Assay
4.11. Adhesion Assay
4.12. Immunofluorescence Staining
4.13. Western Blot Assay
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shi, P.; Xu, J.; Cui, H. The Recent Research Progress of NF-κB Signaling on the Proliferation, Migration, Invasion, Immune Escape and Drug Resistance of Glioblastoma. Int. J. Mol. Sci. 2023, 24, 10337. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.L.; Comba, A.; Faisal, S.M.; Argento, A.; Franson, A.; Barissi, M.N.; Sachdev, S.; Castro, M.G.; Lowenstein, P.R. Gene Therapy for High Grade Glioma: The Clinical Experience. Expert Opin. Biol. Ther. 2023, 23, 145–161. [Google Scholar] [CrossRef]
- De Fazio, E.; Pittarello, M.; Gans, A.; Ghosh, B.; Slika, H.; Alimonti, P.; Tyler, B. Intrinsic and Microenvironmental Drivers of Glioblastoma Invasion. Int. J. Mol. Sci. 2024, 25, 2563. [Google Scholar] [CrossRef]
- Zou, Y.; Xu, L.; Wang, W.; Zhu, X.; Lin, J.; Li, H.; Chen, J.; Xu, W.; Gao, H.; Wu, X.; et al. Muscone restores anoikis sensitivity in TMZ-resistant glioblastoma cells by suppressing TOP2A via the EGFR/Integrin β1/FAK signaling pathway. Phytomedicine 2024, 129, 155714. [Google Scholar] [CrossRef]
- Mukherjee, A.; Ha, P.; Wai, K.C.; Naara, S. The Role of ECM Remodeling, EMT, and Adhesion Molecules in Cancerous Neural Invasion: Changing Perspectives. Adv. Biol. 2022, 6, e2200039. [Google Scholar] [CrossRef] [PubMed]
- Oishi, T.; Koizumi, S.; Kurozumi, K. Molecular Mechanisms and Clinical Challenges of Glioma Invasion. Brain Sci. 2022, 12, 291. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Weiswald, L.-B.; Poulain, L.; Denoyelle, C.; Meryet-Figuiere, M. Involvement of lncRNAs in cancer cells migration, invasion and metastasis: Cytoskeleton and ECM crosstalk. J. Exp. Clin. Cancer Res. 2023, 42, 173. [Google Scholar] [CrossRef]
- Lee, H.; Tsygankov, A.Y. c-Cbl Regulates Glioma Invasion Through Matrix Metalloproteinase 2. J. Cell. Biochem. 2010, 111, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhu, M.; Yu, S.; Hai, L.; Zhang, L.; Zhang, C.; Zhao, P.; Zhou, H.; Wang, S.; Yang, X. Arg kinase mediates CXCL12/CXCR4-induced invadopodia formation and invasion of glioma cells. Exp. Cell Res. 2020, 389, 111893. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Q.; Chen, Z.; Tian, D.; Xu, H.; Cai, Q.; Liu, B.; Deng, G. EFEMP2 is upregulated in gliomas and promotes glioma cell proliferation and invasion. Int. J. Clin. Exp. Pathol. 2015, 8, 10385–10393. [Google Scholar]
- Zhu, Y.; Yan, L.; Zhu, W.; Song, X.; Yang, G.; Wang, S. MMP2/3 promote the growth and migration of laryngeal squamous cell carcinoma via PI3K/Akt-NF-B-mediated epithelial-mesenchymal transformation. J. Cell. Physiol. 2019, 234, 15847–15855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-Y.; Lu, M.; Liu, Z.-K.; Li, H.; Yong, Y.-L.; Zhang, R.-Y.; Chen, Z.-N.; Bian, H. Rab11a regulates MMP2 expression by activating the PI3K/AKT pathway in human hepatocellular carcinoma cells. Pathol. Res. Pract. 2020, 216, 153046. [Google Scholar] [CrossRef] [PubMed]
- Lipinska, T. General route to the total synthesis of sempervirine analogues containing modified E rings, potential cytostatics. Tetrahedron Lett. 2002, 43, 9565–9567. [Google Scholar] [CrossRef]
- Yue, R.; Liu, H.; Huang, Y.; Wang, J.; Shi, D.; Su, Y.; Luo, Y.; Cai, P.; Jin, G.; Yu, C. Sempervirine Inhibits Proliferation and Promotes Apoptosis by Regulating Wnt/β-Catenin Pathway in Human Hepatocellular Carcinoma. Front. Pharmacol. 2021, 12, 806091. [Google Scholar] [CrossRef]
- Caggiano, C.; Guida, E.; Todaro, F.; Bielli, P.; Mori, M.; Ghirga, F.; Quaglio, D.; Botta, B.; Moretti, F.; Grimaldi, P.; et al. Sempervirine inhibits RNA polymerase I transcription independently from p53 in tumor cells. Cell Death Discov. 2020, 6, 111. [Google Scholar] [CrossRef]
- Li, G.; Zhong, Y.; Wang, W.; Jia, X.; Zhu, H.; Jiang, W.; Song, Y.; Xu, W.; Wu, S. Sempervirine Mediates Autophagy and Apoptosis via the Akt/mTOR Signaling Pathways in Glioma Cells. Front. Pharmacol. 2021, 12, 770667. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Walker, E.; Davis, F. Malignant Primary Brain and Other Central Nervous System Tumours Diagnosed among the Canadian Population from 2009 to 2013. Neuro Oncol. 2018, 20, 83–84. [Google Scholar] [CrossRef]
- Tan, A.C.; Ashley, D.M.; Lopez, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of glioblastoma: State of the art and future directions. CA-A Cancer J. Clin. 2020, 70, 299–312. [Google Scholar] [CrossRef]
- Claes, A.; Idema, A.J.; Wesseling, P. Diffuse glioma growth: A guerilla war. Acta Neuropathol. 2007, 114, 443–458. [Google Scholar] [CrossRef]
- Vollmann-Zwerenz, A.; Leidgens, V.; Feliciello, G.; Klein, C.A.; Hau, P. Tumor Cell Invasion in Glioblastoma. Int. J. Mol. Sci. 2020, 21, 1932. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Transport of molecules, particles, and cells in solid tumors. Annu. Rev. Biomed. Eng. 1999, 1, 241–263. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Delivery of molecular and cellular medicine to solid tumors. Adv. Drug Deliv. Rev. 2012, 64, 353–365. [Google Scholar] [CrossRef]
- Munson, J.; Bonner, M.; Fried, L.; Hofmekler, J.; Arbiser, J.; Bellamkonda, R. Identifying new small molecule anti-invasive compounds for glioma treatment. Cell Cycle 2013, 12, 2200–2209. [Google Scholar] [CrossRef] [PubMed]
- Barrette, A.M.; Ronk, H.; Joshi, T.; Mussa, Z.; Mehrotra, M.; Bouras, A.; Nudelman, G.; Raj, J.G.J.; Bozec, D.; Lam, W.; et al. Anti-invasive efficacy and survival benefit of the YAP-TEAD inhibitor Verteporfin in preclinical glioblastoma models. Neuro Oncol. 2022, 24, 694–707. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, J.; Li, X.; Fu, L.; Gao, D.; Bai, H.; Liu, X. Effects of recombinant human bone morphogenic protein-2 and hyaluronic acid on invasion of brain glioma in vivo. Zhonghua Yi Xue Za Zhi 2002, 82, 90–93. [Google Scholar]
- Qu, Y.; Qi, L.; Hao, L.; Zhu, J. Upregulation of circ-ASPH contributes to glioma cell proliferation and aggressiveness by targeting the miR-599/AR/SOCS2-AS1 signaling pathway. Oncol. Lett. 2021, 21, 388. [Google Scholar] [CrossRef]
- Li, X.; Wu, J.-F. Recent Developments in Patent Anti-Cancer Agents Targeting the Matrix Metalloproteinases (MMPs). Recent Pat. Anti-Cancer Drug Discov. 2010, 5, 109–141. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Li, X.-H.; Tian, G.-W.; Zhang, D.-Y.; Gao, H.; Wang, Z.-Y. MAGI1 Inhibits the Proliferation, Migration and Invasion of Glioma Cells. Oncotargets Ther. 2019, 12, 11281–11289. [Google Scholar] [CrossRef]
- Louca, M.; Stylianou, A.; Minia, A.; Pliaka, V.; Alexopoulos, L.G.; Gkretsi, V.; Stylianopoulos, T. Ras suppressor-1 (RSU-1) promotes cell invasion in aggressive glioma cells and inhibits it in non-aggressive cells through STAT6 phospho-regulation. Sci. Rep. 2019, 9, 7782. [Google Scholar] [CrossRef]
- Zepecki, J.P.; Snyder, K.M.; Moreno, M.M.; Fajardo, E.; Fiser, A.; Ness, J.; Sarkar, A.; Toms, S.A.; Tapinos, N. Regulation of human glioma cell migration, tumor growth, and stemness gene expression using a Lck targeted inhibitor. Oncogene 2019, 38, 1734–1750. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.-J.; Lai, Y.-H.; Manne, R.K.; Tsai, Y.-S.; Sarbassov, D.; Lin, H.-K. Akt: A key transducer in cancer. J. Biomed. Sci. 2022, 29, 76. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yang, P.; Tu, K.; Zhang, H.; Zheng, X.; Yao, Y.; Liu, Q. TPX2 knockdown suppressed hepatocellular carcinoma cell invasion via inactivating AKT signaling and inhibiting MMP2 and MMP9 expression. Chin. J. Cancer Res. 2014, 26, 410–417. [Google Scholar]
- Guan, N.; Huo, X.; Zhang, Z.; Zhang, S.; Luo, J.; Guo, W. Ginsenoside Rh2 inhibits metastasis of glioblastoma multiforme through Akt-regulated MMP13. Tumor Biol. 2015, 36, 6789–6795. [Google Scholar] [CrossRef]
- Jiang, Y.-W.; Cheng, H.-Y.; Kuo, C.-L.; Way, T.-D.; Lien, J.-C.; Chueh, F.-S.; Lin, Y.-L.; Chung, J.-G. Tetrandrine inhibits human brain glioblastoma multiforme GBM 8401 cancer cell migration and invasion in vitro. Environ. Toxicol. 2019, 34, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.-S.; Lin, S.-Y.; Han, X.; Xu, H.-Z.; Gao, Y.-L.; Qin, Z.-Y. Casein kinase 1 (CK1) promotes the proliferation and metastasis of glioma cells via the phosphatidylinositol 3 kinase-matrix metalloproteinase 2 (AKT-MMP2) pathway. Ann. Transl. Med. 2021, 9, 659. [Google Scholar] [CrossRef]
- Justus, C.R.; Marie, M.A.; Sanderlin, E.J.; Yang, L.V. Transwell In Vitro Cell Migration and Invasion Assays. Methods Mol. Biol. 2023, 2644, 349–359. [Google Scholar]
- Rodriguez, L.G.; Wu, X.; Guan, J.-L. Wound-healing assay. Methods Mol. Biol. 2005, 294, 23–29. [Google Scholar] [PubMed]
- Shao, Y.-Y.; Zhang, T.-L.; Wu, L.-X.; Zou, H.-C.; Li, S.; Huang, J.; Zhou, H.-H. AKT Axis, miR-21, and RECK Play Pivotal Roles in Dihydroartemisinin Killing Malignant Glioma Cells. Int. J. Mol. Sci. 2017, 18, 350. [Google Scholar] [CrossRef]
- Qu, L.; Lin, P.; Lin, M.; Ye, S.; Papa Akuetteh, P.D.; Zhu, Y. Fraxetin Inhibits the Proliferation and Metastasis of Glioma Cells by Inactivating JAK2/STAT3 Signaling. Evid.-Based Complement. Altern. Med. 2021, 2021, 5540139. [Google Scholar] [CrossRef]
- Guo, G.; Yao, W.; Zhang, Q.; Bo, Y. Oleanolic Acid Suppresses Migration and Invasion of Malignant Glioma Cells by Inactivating MAPK/ERK Signaling Pathway. PLoS ONE 2013, 8, e72079. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.J.W.; Lin, X.; Chen, H.; Xu, W.; Wu, S. Preparation and quality evaluation of sempervir in echitosan thermosensitive microemulsion-based hydrogel. Chin. J. Hosp. Pharm. 2024, 44, 2015–2020. [Google Scholar]
- Jiang, W.Z.Y.; Li, G.; Song, Y.; Wu, S. Determination of equilibrium solubility, oil-water partition coefficient and dissociation constant of sempervirine. J. Pharm. Res. 2021, 40, 656–659. [Google Scholar]
- Jiang, W.S.Y.; Xu, W.; Guo, Z.; Wu, S. Formulation optimization and safety evaluation of sempervirine microemulsion. Chin. Tradit. Pat. Med. 2023, 45, 3187–3193. [Google Scholar]
- Wang, Z.; Jensen, M.A.; Zenklusen, J.C. A Practical Guide to The Cancer Genome Atlas (TCGA). Methods Mol. Biol. 2016, 1418, 111–141. [Google Scholar]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Farrell, C.M.; Feldgarden, M.; Fine, A.M.; Funk, K.; et al. Database resources of the National Center for Biotechnology Information in 2023. Nucleic Acids Res. 2023, 51, D29–D38. [Google Scholar] [CrossRef] [PubMed]
- Gaulton, A.; Hersey, A.; Nowotka, M.; Bento, A.P.; Chambers, J.; Mendez, D.; Mutowo, P.; Atkinson, F.; Bellis, L.J.; Cibrian-Uhalte, E.; et al. The ChEMBL database in 2017. Nucleic Acids Res. 2017, 45, D945–D954. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. Swiss Target Prediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef]
- Amberger, J.S.; Bocchini, C.A.; Schiettecatte, F.; Scott, A.F.; Hamosh, A. OMIM.org: Online Mendelian Inheritance in Man (OMIM®), an online catalog of human genes and genetic disorders. Nucleic Acids Res. 2015, 43, D789–D798. [Google Scholar] [CrossRef]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Alpi, E.; Bely, B.; Bingley, M.; Britto, R.; Bursteinas, B.; Busiello, G.; et al. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Otasek, D.; Morris, J.H.; Boucas, J.; Pico, A.R.; Demchak, B. Cytoscape Automation: Empowering workflow-based network analysis. Genome Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef]
- Touw, W.G.; Baakman, C.; Black, J.; te Beek, T.A.H.; Krieger, E.; Joosten, R.P.; Vriend, G. A series of PDB-related databanks for everyday needs. Nucleic Acids Res. 2015, 43, D364–D368. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.-X.; Cao, Y. CB-Dock2: Improved protein ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022, 50, W159–W164. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Targets | PDB-ID | Vina Score |
|---|---|---|---|
| Sempervirine | BMP2 | 1REU | −6.5 |
| DUSP6 | 1MKP | −7.8 | |
| MMP2 | 7XGJ | −10.4 | |
| MMP13 | 4JPA | −10.4 | |
| AKT1 | 7NH5 | −10.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Wang, W.; Song, Y.; Chen, H.; Lin, X.; Chen, J.; Chen, Y.; Huang, J.; Li, D.; Wu, S. Exploring the Mechanism of Sempervirine Inhibiting Glioblastoma Invasion Based on Network Pharmacology and Bioinformatics. Pharmaceuticals 2024, 17, 1318. https://doi.org/10.3390/ph17101318
Zhang B, Wang W, Song Y, Chen H, Lin X, Chen J, Chen Y, Huang J, Li D, Wu S. Exploring the Mechanism of Sempervirine Inhibiting Glioblastoma Invasion Based on Network Pharmacology and Bioinformatics. Pharmaceuticals. 2024; 17(10):1318. https://doi.org/10.3390/ph17101318
Chicago/Turabian StyleZhang, Bingqiang, Wenyi Wang, Yu Song, Huixian Chen, Xinxin Lin, Jingjing Chen, Ying Chen, Jinfang Huang, Desen Li, and Shuisheng Wu. 2024. "Exploring the Mechanism of Sempervirine Inhibiting Glioblastoma Invasion Based on Network Pharmacology and Bioinformatics" Pharmaceuticals 17, no. 10: 1318. https://doi.org/10.3390/ph17101318