

Unique Effects of (R)-Ketamine Compared to (S)-Ketamine on EEG Theta Power in Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
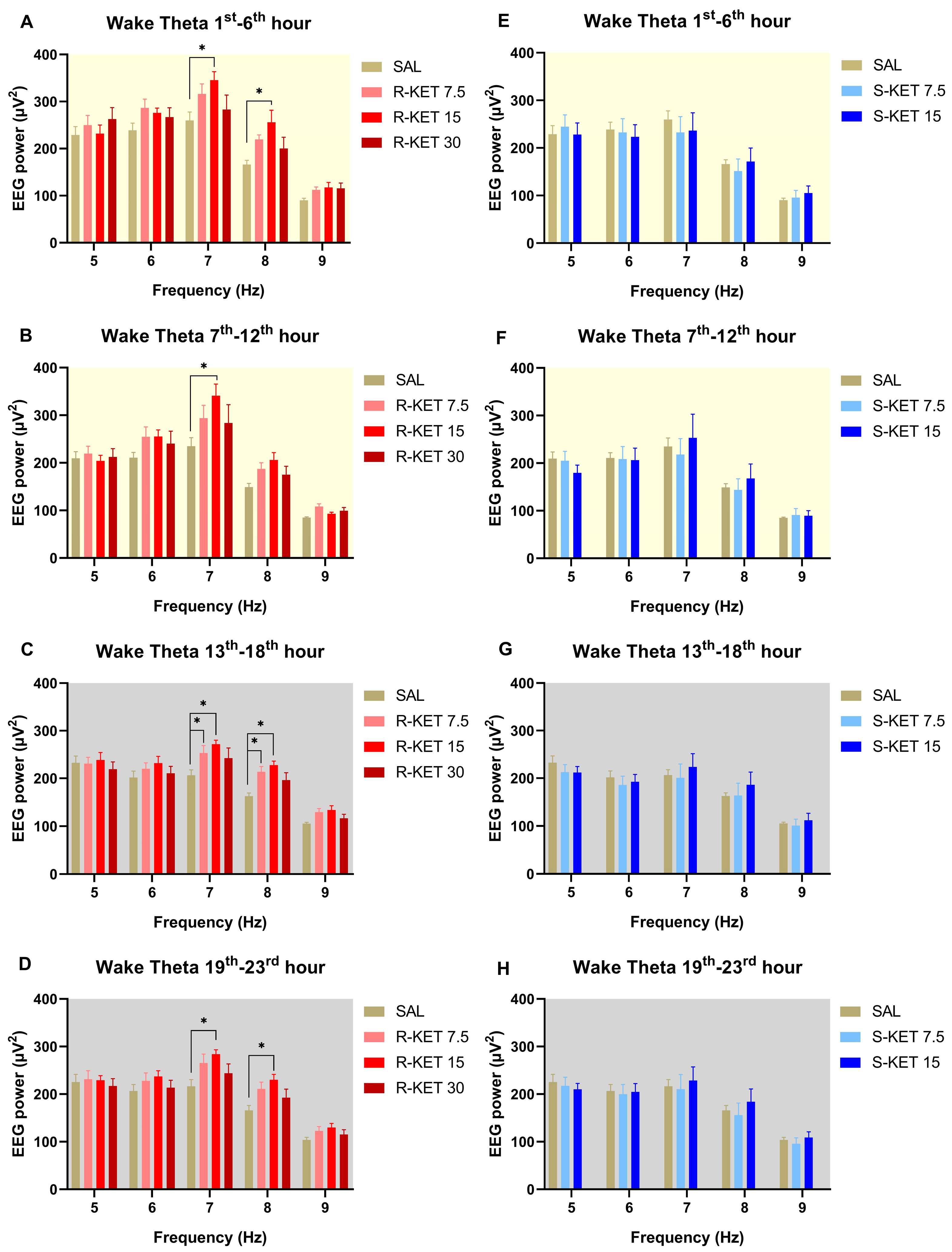
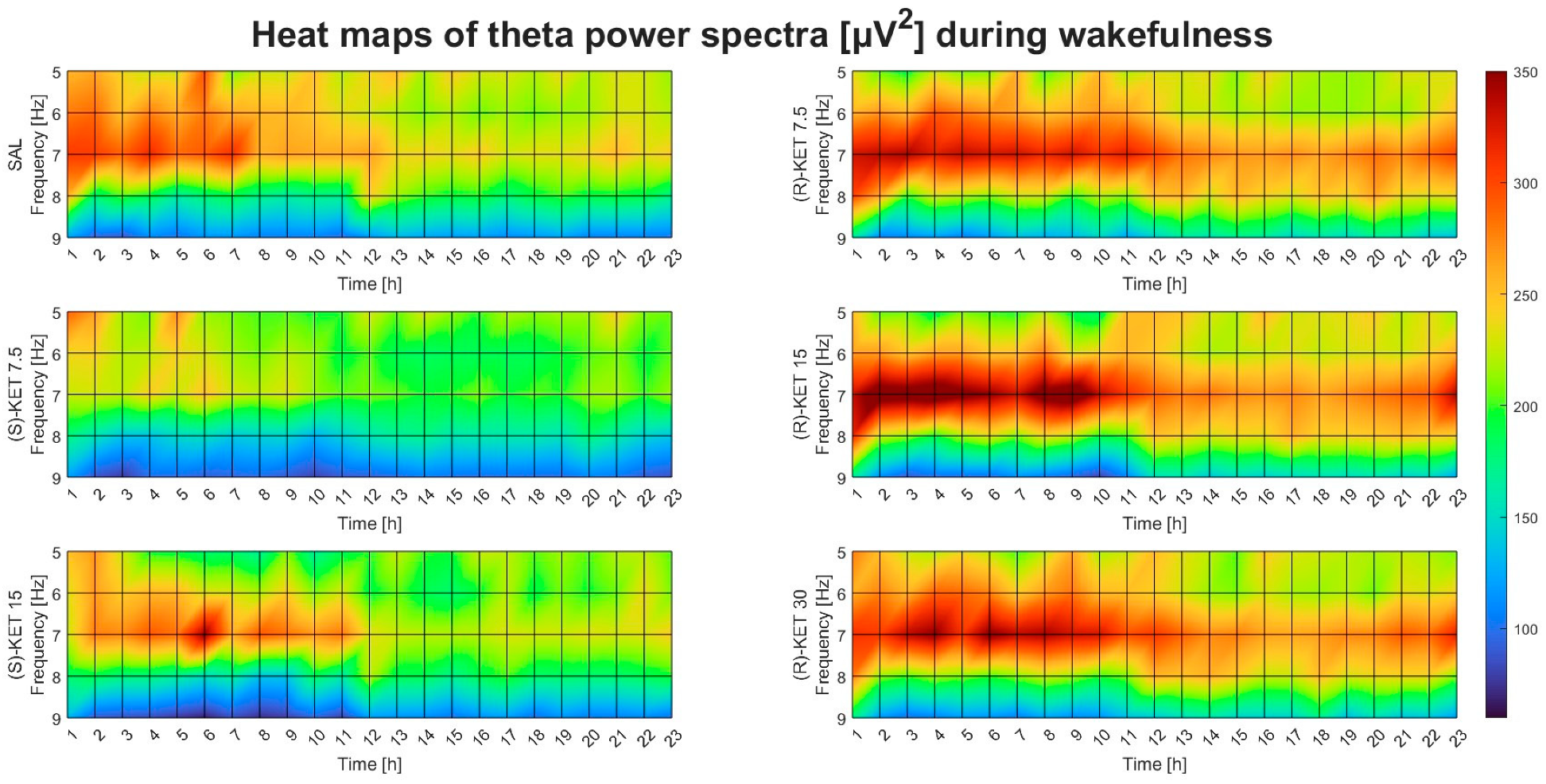
2.1. Effects of (R)-ketamine and (S)-ketamine on qEEG during Wakefulness
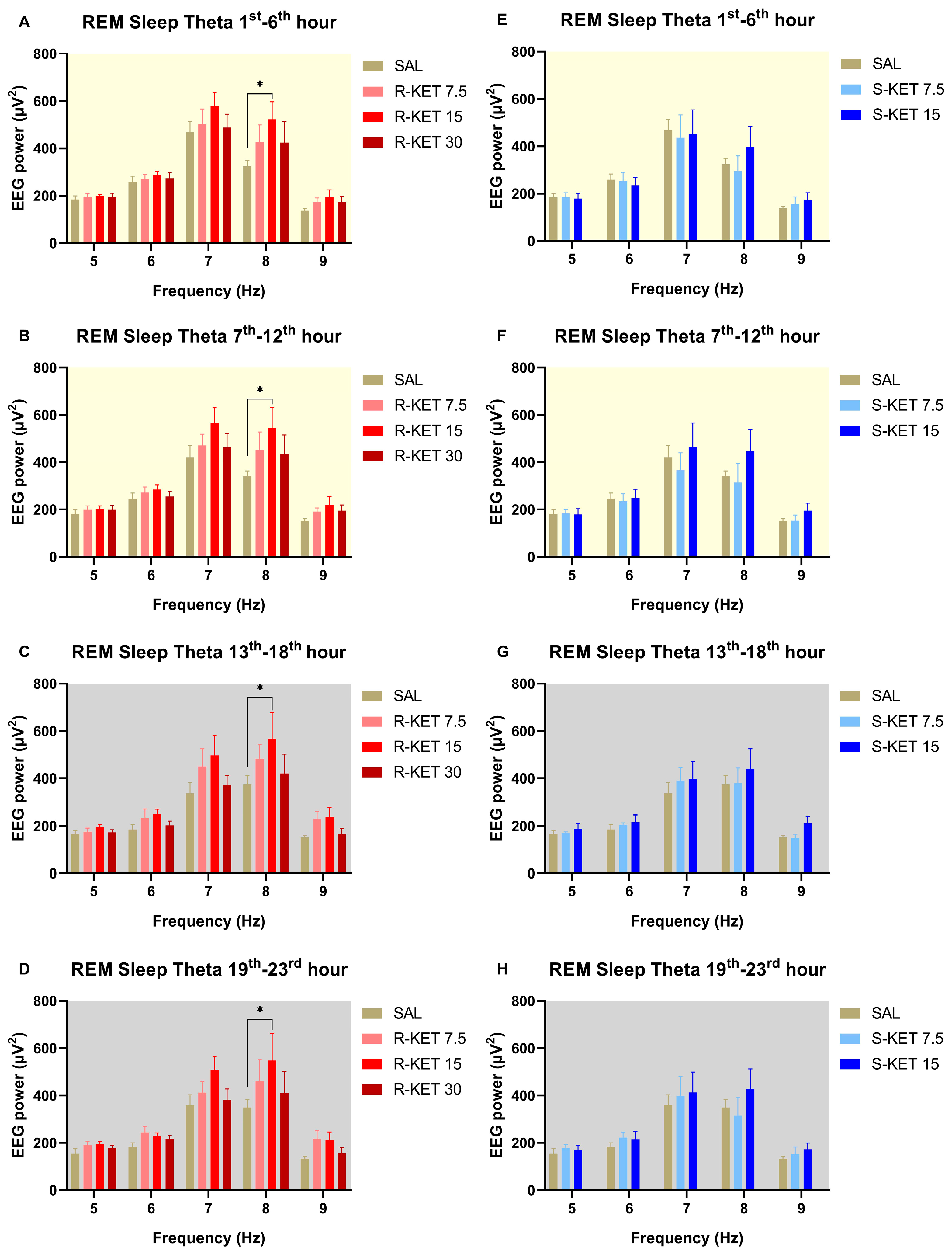
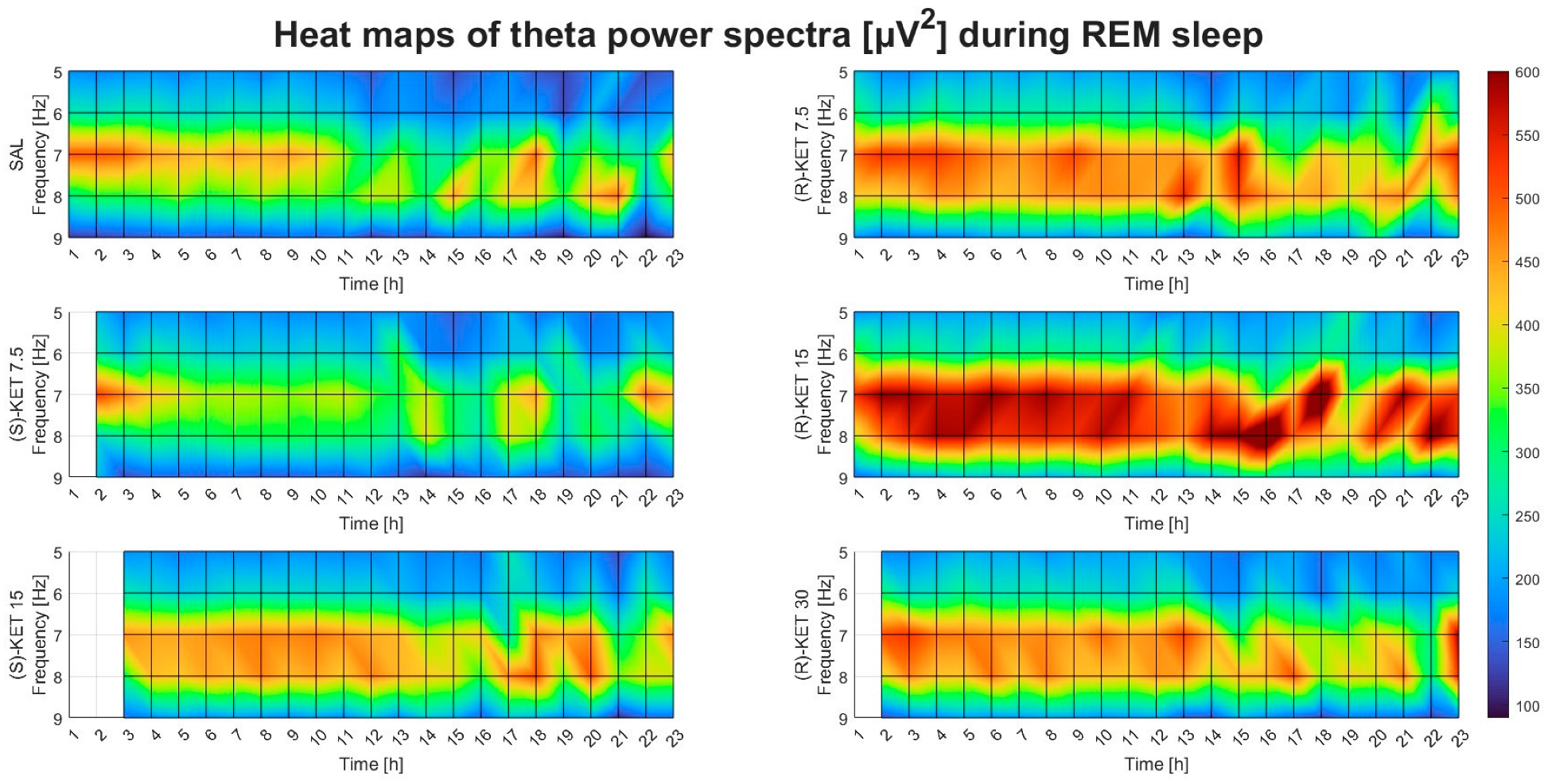
2.2. Effects of (R)-ketamine and (S)-ketamine on qEEG during REM Sleep
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Surgery
4.3. Drugs and Treatments
4.4. EEG Recording and Analysis
4.5. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zanos, P.; Gould, T. Mechanisms of ketamine action as an antidepressant. Mol. Psychiatry 2018, 23, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Ago, Y.; Yokoyama, R.; Asano, S.; Hashimoto, H. Roles of the monoaminergic system in the antidepressant effects of ketamine and its metabolites. Neuropharmacology 2023, 223, 109313. [Google Scholar] [CrossRef]
- Bonaventura, J.; Lam, S.; Carlton, M.; Boehm, M.A.; Gomez, J.L.; Solís, O.; Sánchez-Soto, M.; Morris, P.J.; Fredriksson, I.; Thomas, C.J. Pharmacological and behavioral divergence of ketamine enantiomers: Implications for abuse liability. Mol. Psychiatry 2021, 26, 6704–6722. [Google Scholar] [CrossRef]
- Johnston, J.N.; Henter, I.D.; Zarate, C.A., Jr. The antidepressant actions of ketamine and its enantiomers. Pharmacol. Ther. 2023, 246, 108431. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Shirayama, Y.; Zhang, J.C.; Ren, Q.; Yao, W.; Ma, M.; Dong, C.; Hashimoto, K. R-ketamine: A rapid-onset and sustained antidepressant without psychotomimetic side effects. Transl. Psychiatry 2015, 5, e632. [Google Scholar] [CrossRef]
- Koncz, S.; Papp, N.; Pothorszki, D.; Bagdy, G. (S)-Ketamine but Not (R)-Ketamine Shows Acute Effects on Depression-like Behavior and Sleep-Wake Architecture in Rats. Int. J. Neuropsychopharmacol. 2023, 26, 618–626. [Google Scholar] [CrossRef]
- Tan, Y.; Fujita, Y.; Qu, Y.; Chang, L.; Pu, Y.; Wang, S.; Wang, X.; Hashimoto, K. Phencyclidine-induced cognitive deficits in mice are ameliorated by subsequent repeated intermittent administration of (R)-ketamine, but not (S)-ketamine: Role of BDNF-TrkB signaling. Pharmacol. Biochem. Behav. 2020, 188, 172839. [Google Scholar] [CrossRef] [PubMed]
- Fujita, A.; Fujita, Y.; Pu, Y.; Chang, L.; Hashimoto, K. MPTP-induced dopaminergic neurotoxicity in mouse brain is attenuated after subsequent intranasal administration of (R)-ketamine: A role of TrkB signaling. Psychopharmacology 2020, 237, 83–92. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Tan, Y.; Qu, Y.; Shan, J.; Hashimoto, K. (R)-ketamine ameliorates the progression of experimental autoimmune encephalomyelitis in mice. Brain Res. Bull. 2021, 177, 316–323. [Google Scholar] [CrossRef]
- Wang, X.; Chang, L.; Wan, X.; Tan, Y.; Qu, Y.; Shan, J.; Yang, Y.; Ma, L.; Hashimoto, K. (R)-ketamine ameliorates demyelination and facilitates remyelination in cuprizone-treated mice: A role of gut–microbiota–brain axis. Neurobiol. Dis. 2022, 165, 105635. [Google Scholar] [CrossRef]
- Nuñez, A.; Buño, W. The theta rhythm of the hippocampus: From neuronal and circuit mechanisms to behavior. Front. Cell. Neurosci. 2021, 15, 649262. [Google Scholar] [CrossRef] [PubMed]
- Karakaş, S. A review of theta oscillation and its functional correlates. Int. J. Psychophysiol. 2020, 157, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, I.C.; Rathore, S. The role of REM sleep theta activity in emotional memory. Front. Psychol. 2015, 6, 1439. [Google Scholar] [CrossRef] [PubMed]
- Vas, S.; Kátai, Z.; Kostyalik, D.; Pap, D.; Molnár, E.; Petschner, P.; Kalmár, L.; Bagdy, G. Differential adaptation of REM sleep latency, intermediate stage and theta power effects of escitalopram after chronic treatment. J. Neural Transm. 2013, 120, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Kostyalik, D.; Kátai, Z.; Vas, S.; Pap, D.; Petschner, P.; Molnár, E.; Gyertyán, I.; Kalmár, L.; Tóthfalusi, L.; Bagdy, G. Chronic escitalopram treatment caused dissociative adaptation in serotonin (5-HT) 2C receptor antagonist-induced effects in REM sleep, wake and theta wave activity. Exp. Brain Res. 2014, 232, 935–946. [Google Scholar] [CrossRef]
- Kang, M.J.; Hawken, E.; Vazquez, G.H. The mechanisms behind rapid antidepressant effects of ketamine: A systematic review with a focus on molecular neuroplasticity. Front. Psychiatry 2022, 13, 860882. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K. Arketamine for cognitive impairment in psychiatric disorders. Eur. Arch. Psychiatry Clin. Neurosci. 2023, 273, 1513–1525. [Google Scholar] [CrossRef]
- Costers, L.; Van Schependom, J.; Laton, J.; Baijot, J.; Sjøgård, M.; Wens, V.; De Tiège, X.; Goldman, S.; D’Haeseleer, M.; D’hooghe, M.B. The role of hippocampal theta oscillations in working memory impairment in multiple sclerosis. Hum. Brain Mapp. 2021, 42, 1376–1390. [Google Scholar] [CrossRef]
- Fukumoto, K.; Toki, H.; Iijima, M.; Hashihayata, T.; Yamaguchi, J.-I.; Hashimoto, K.; Chaki, S. Antidepressant potential of (R)-ketamine in rodent models: Comparison with (S)-ketamine. J. Pharmacol. Exp. Ther. 2017, 361, 9–16. [Google Scholar] [CrossRef]
- Hunt, M.J.; Kasicki, S. A systematic review of the effects of NMDA receptor antagonists on oscillatory activity recorded in vivo. J. Psychopharmacol. 2013, 27, 972–986. [Google Scholar] [CrossRef]
- Ishima, T.; Nishimura, T.; Iyo, M.; Hashimoto, K. Potentiation of nerve growth factor-induced neurite outgrowth in PC12 cells by donepezil: Role of sigma-1 receptors and IP3 receptors. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1656–1659. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Su, T.-P. σ-1 receptor ligands: Potential in the treatment of neuropsychiatric disorders. CNS Drugs 2004, 18, 269–284. [Google Scholar] [CrossRef]
- Toyohara, J.; Sakata, M.; Ishiwata, K. Roles of σ 1 receptors in the mechanisms of action of CNS drugs. Transl. Neurosci. 2012, 3, 294–299. [Google Scholar] [CrossRef]
- Schmidt, H.R.; Kruse, A.C. The molecular function of σ receptors: Past, present, and future. Trends Pharmacol. Sci. 2019, 40, 636–654. [Google Scholar] [CrossRef]
- Ago, Y.; Tanabe, W.; Higuchi, M.; Tsukada, S.; Tanaka, T.; Yamaguchi, T.; Igarashi, H.; Yokoyama, R.; Seiriki, K.; Kasai, A. (R)-ketamine induces a greater increase in prefrontal 5-HT release than (S)-ketamine and ketamine metabolites via an AMPA receptor-independent mechanism. Int. J. Neuropsychopharmacol. 2019, 22, 665–674. [Google Scholar] [CrossRef]
- Skosnik, P.D.; Sloshower, J.; Safi-Aghdam, H.; Pathania, S.; Syed, S.; Pittman, B.; D’Souza, D.C. Sub-acute effects of psilocybin on EEG correlates of neural plasticity in major depression: Relationship to symptoms. J. Psychopharmacol. 2023, 37, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Kantor, S.; Jakus, R.; Balogh, B.; Benko, A.; Bagdy, G. Increased wakefulness, motor activity and decreased theta activity after blockade of the 5-HT 2B receptor by the subtype-selective antagonist SB-215505. Br. J. Pharmacol. 2004, 142, 1332–1342. [Google Scholar] [CrossRef] [PubMed]
- Papp, N.; Koncz, S.; Kostyalik, D.; Kitka, T.; Petschner, P.; Vas, S.; Bagdy, G. Acute 5-HT2C Receptor Antagonist SB-242084 Treatment Affects EEG Gamma Band Activity Similarly to Chronic Escitalopram. Front. Pharmacol. 2020, 10, 1636. [Google Scholar] [CrossRef]
- Koncz, S.; Papp, N.; Menczelesz, N.; Pothorszki, D.; Bagdy, G. EEG and Sleep Effects of Tramadol Suggest Potential Antidepressant Effects with Different Mechanisms of Action. Pharmaceuticals 2021, 14, 431. [Google Scholar] [CrossRef]
- Papp, N.; Vas, S.; Bogáthy, E.; Kátai, Z.; Kostyalik, D.; Bagdy, G. Acute and chronic escitalopram alter EEG gamma oscillations differently: Relevance to therapeutic effects. Eur. J. Pharm. Sci. 2018, 121, 347–355. [Google Scholar] [CrossRef]
- Banerjee, P.; Donello, J.E.; Hare, B.; Duman, R.S. Rapastinel, an NMDAR positive modulator, produces distinct behavioral, sleep, and EEG profiles compared with ketamine. Behav. Brain Res. 2020, 391, 112706. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pothorszki, D.; Koncz, S.; Török, D.; Papp, N.; Bagdy, G. Unique Effects of (R)-Ketamine Compared to (S)-Ketamine on EEG Theta Power in Rats. Pharmaceuticals 2024, 17, 194. https://doi.org/10.3390/ph17020194
Pothorszki D, Koncz S, Török D, Papp N, Bagdy G. Unique Effects of (R)-Ketamine Compared to (S)-Ketamine on EEG Theta Power in Rats. Pharmaceuticals. 2024; 17(2):194. https://doi.org/10.3390/ph17020194
Chicago/Turabian StylePothorszki, Dóra, Szabolcs Koncz, Dóra Török, Noémi Papp, and György Bagdy. 2024. "Unique Effects of (R)-Ketamine Compared to (S)-Ketamine on EEG Theta Power in Rats" Pharmaceuticals 17, no. 2: 194. https://doi.org/10.3390/ph17020194