

Effect of Matricaria aurea Essential Oils on Biofilm Development, Virulence Factors and Quorum Sensing-Dependent Genes of Pseudomonas aeruginosa
 , , , ,
, , , ,  , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Chemical Composition of M. aurea EO
2.2. Antibacterial Activity
2.3. Cell Viability
2.4. Antibiofilm
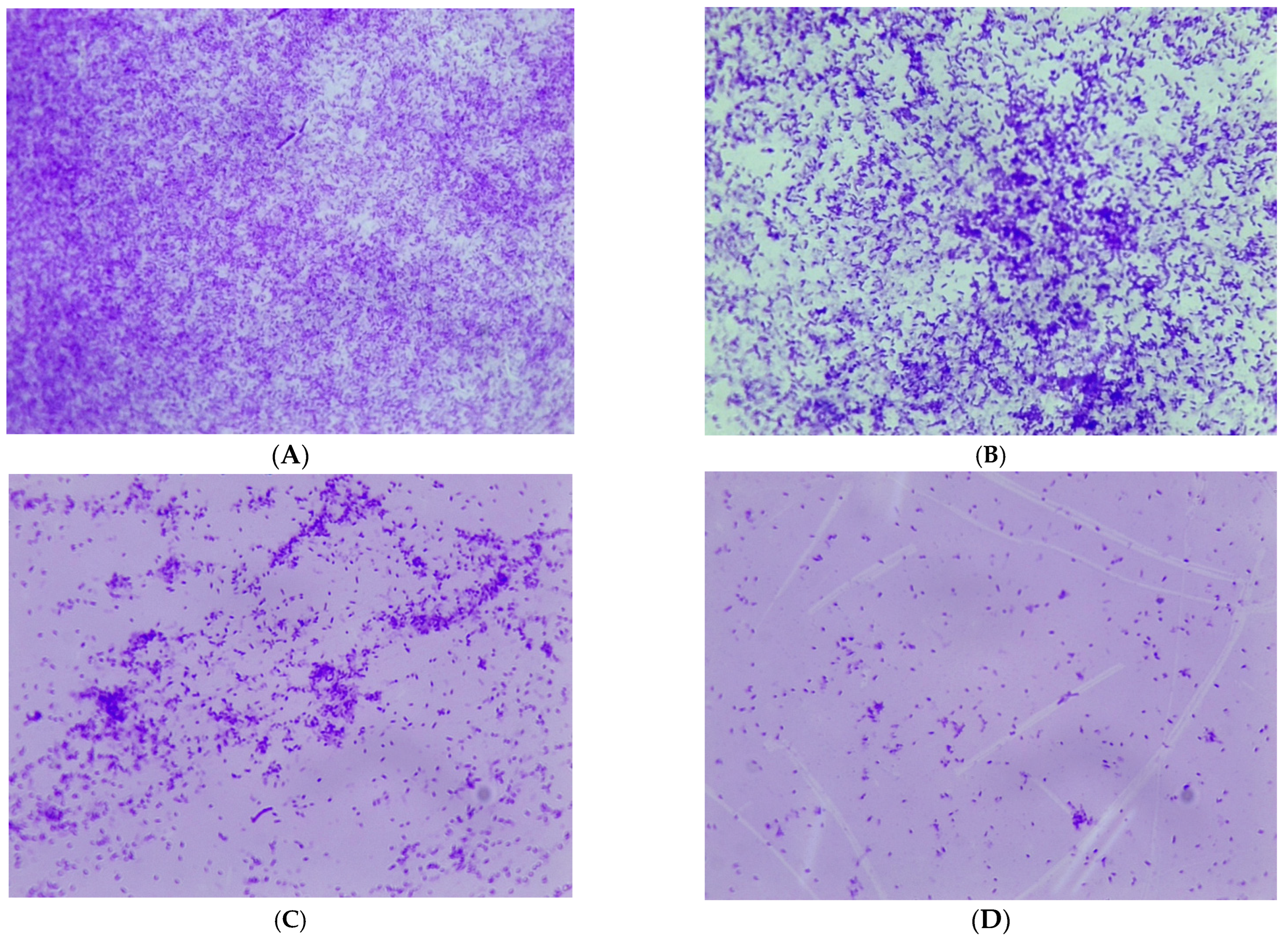
2.5. Biofilm Visualization
2.6. Violacein Inhibition of C. violaceum
2.7. Effect on Biofilm Development
2.7.1. Swarming Motility
2.7.2. Aggregation
2.7.3. Hydrophobicity
2.7.4. EPS Production
2.8. Virulence Factors
2.8.1. Pyocyanin
2.8.2. Rhamnolipids
2.8.3. LasA Protease
2.8.4. Chitinase Activity
2.9. Mechanism of QS
2.9.1. N-Acyl Homoserine Lactones (AHL)
2.9.2. qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Materials and EO Extraction
4.3. Gas Chromatography Mass Spectrometry
4.4. Bacterial Strains
4.5. Antibacterial Activity
4.5.1. Disc Diffusion Assay
4.5.2. MIC
4.6. Cell Viability
4.7. Antibiofilm
4.8. Biofilm Visualization
4.9. Violacein Assay
4.10. Effect on Biofilm Development
4.10.1. Swarming Motility
4.10.2. Aggregation
4.10.3. Hydrophobicity
4.10.4. EPS Production
4.11. Effect on Virulence Factors
4.11.1. Pyocyanin
4.11.2. Rhamnolipids
4.11.3. LasA Protease
4.11.4. Chitinase
4.12. Mechanism of QS
4.12.1. N-Acyl Homoserine Lactones (AHL)
4.12.2. qRT-PCR
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Qin, S.; Xiao, W.; Zhou, C.; Pu, Q.; Deng, X.; Lan, L.; Liang, H.; Song, X.; Wu, M. Pseudomonas aeruginosa: Pathogenesis, Virulence Factors, Antibiotic Resistance, Interaction with Host, Technology Advances and Emerging Therapeutics. Signal Transduct. Target. Ther. 2022, 7, 199. [Google Scholar] [CrossRef]
- Paprocka, P.; Durnaś, B.; Mańkowska, A.; Król, G.; Wollny, T.; Bucki, R. Pseudomonas aeruginosa Infections in Cancer Patients. Pathogens 2022, 11, 679. [Google Scholar] [CrossRef]
- Wood, S.J.; Kuzel, T.M.; Shafikhani, S.H. Pseudomonas aeruginosa: Infections, Animal Modeling, and Therapeutics. Cells 2023, 12, 199. [Google Scholar] [CrossRef] [PubMed]
- Karruli, A.; Catalini, C.; D’Amore, C.; Foglia, F.; Mari, F.; Harxhi, A.; Galdiero, M.; Durante-Mangoni, E. Evidence-Based Treatment of Pseudomonas aeruginosa Infections: A Critical Reappraisal. Antibiotics 2023, 12, 399. [Google Scholar] [CrossRef] [PubMed]
- Tapper, M.L.; Armstrong, D. Bacteremia Due to Pseudomonas aeruginosa Complicating Neoplastic Disease: A Progress Report. J. Infect. Dis. 1974, 130, S14–S23. [Google Scholar] [CrossRef] [PubMed]
- Chatzinikolaou, I.; Abi-Said, D.; Bodey, G.P.; Rolston, K.V.I.; Tarrand, J.J.; Samonis, G. Recent Experience with Pseudomonas aeruginosa Bacteremia in Patients with Cancer: Retrospective Analysis of 245 Episodes. Arch. Intern. Med. 2000, 160, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.-I.; Kim, S.-H.; Kim, H.-B.; Park, S.-W.; Choe, Y.-J.; Oh, M.; Kim, E.-C.; Choe, K.-W. Pseudomonas aeruginosa Bacteremia: Risk Factors for Mortality and Influence of Delayed Receipt of Effective Antimicrobial Therapy on Clinical Outcome. Clin. Infect. Dis. 2003, 37, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Elmassry, M.M.; Colmer-Hamood, J.A.; Kopel, J.; San Francisco, M.J.; Hamood, A.N. Anti-Pseudomonas aeruginosa Vaccines and Therapies: An Assessment of Clinical Trials. Microorganisms 2023, 11, 916. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic Resistance in Pseudomonas aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States, 2019; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019. [Google Scholar]
- Shrivastava, S.R.; Shrivastava, P.S.; Ramasamy, J. World Health Organization Releases Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. J. Med. Soc. 2018, 32, 76–77. [Google Scholar] [CrossRef]
- Zloh, M. AR Mechanism-Based Drug Design. In Antibiotic Drug Resistance; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 275–309. [Google Scholar]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and Resistance of Pseudomonas aeruginosa Biofilms to Antimicrobial Agents—How P. aeruginosa Can Escape Antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef]
- Li, Z.; Nair, S.K. Quorum Sensing: How Bacteria Can Coordinate Activity and Synchronize Their Response to External Signals? Protein Sci. 2012, 21, 1403–1417. [Google Scholar] [CrossRef]
- Naoun, A.A.; Raphael, I.; Forsthuber, T.G. Immunoregulation via Cell Density and Quorum Sensing-like Mechanisms: An Underexplored Emerging Field with Potential Translational Implications. Cells 2022, 11, 2442. [Google Scholar] [CrossRef] [PubMed]
- Duplantier, M.; Lohou, E.; Sonnet, P. Quorum Sensing Inhibitors to Quench P. aeruginosa Pathogenicity. Pharmaceuticals 2021, 14, 1262. [Google Scholar] [CrossRef]
- Bouyahya, A.; Chamkhi, I.; Balahbib, A.; Rebezov, M.; Shariati, M.A.; Wilairatana, P.; Mubarak, M.S.; Benali, T.; El Omari, N. Mechanisms, Anti-Quorum-Sensing Actions, and Clinical Trials of Medicinal Plant Bioactive Compounds against Bacteria: A Comprehensive Review. Molecules 2022, 27, 1484. [Google Scholar] [CrossRef]
- Qaralleh, H. Chemical Composition and Quorum Sensing Inhibitory Effect of Nepeta curviflora Methanolic Extract against ESBL Pseudomonas aeruginosa. J. Pharmacopunct. 2023, 26, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A.; Dakka, N.; Et-Touys, A.; Abrini, J.; Bakri, Y. Medicinal Plant Products Targeting Quorum Sensing for Combating Bacterial Infections. Asian Pac. J. Trop. Med. 2017, 10, 729–743. [Google Scholar] [CrossRef]
- Qaralleh, H.; Khleifat, K.M.; Khlaifat, A.M.; Al-limoun, M.; Al-Tawarah, N.M.; Alhroob, A.M.; Alsaudi, A.B. Chemical Composition, Antioxidant and Inhibitory Effect of Cupressus sempervirens Essential Oils and Methanolic Extract on Beta-Lactamase Producing Isolates. Res. J. Pharm. Technol. 2021, 14, 4673–4679. [Google Scholar] [CrossRef]
- Camele, I.; Elshafie, H.S.; Caputo, L.; De Feo, V. Anti-Quorum Sensing and Antimicrobial Effect of Mediterranean Plant Essential Oils against Phytopathogenic Bacteria. Front. Microbiol. 2019, 10, 2619. [Google Scholar] [CrossRef] [PubMed]
- Oran, S.A.; Al-Eisawi, D.M. Check-List of Medicinal Plants in Jordan. Dirasat 1998, 25, 84–112. [Google Scholar]
- Qnais, E. The Analgesic Effect of the Ethanolic Extract of Matricaria aurea. Turk. J. Biol. 2011, 35, 347–352. [Google Scholar] [CrossRef]
- Kheder, F.B.H.; Mahjoub, M.A.; Zaghrouni, F.; Kwaja, S.; Helal, A.N.; Mighri, Z. Chemical Composition Antioxidant and Antimicrobial Activities of the Essential Oils of Matricaria aurea Loefl. Growing in Tunisia. J. Essent. Oil Bear. Plants 2014, 17, 493–505. [Google Scholar] [CrossRef]
- Høiby, N.; Ciofu, O.; Johansen, H.K.; Song, Z.; Moser, C.; Jensen, P.Ø.; Molin, S.; Givskov, M.; Tolker-Nielsen, T.; Bjarnsholt, T. The Clinical Impact of Bacterial Biofilms. Int. J. Oral Sci. 2011, 3, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Qaralleh, H.; Al-Limoun, M.; Khlaifat, A.; Khleifat, K.; Al-Tawarah, N.; Hashem, A.-H.; Alsharafa, K. Antibacterial and Antibiofilm Activities of a Traditional Herbal Formula against Respiratory Infection Causing Bacteria. Trop. J. Nat. Prod. Res. 2020, 4, 527–534. [Google Scholar]
- Gordya, N.; Yakovlev, A.; Kruglikova, A.; Tulin, D.; Potolitsina, E.; Suborova, T.; Bordo, D.; Rosano, C.; Chernysh, S. Natural Antimicrobial Peptide Complexes in the Fighting of Antibiotic Resistant Biofilms: Calliphora vicina Medicinal Maggots. PLoS ONE 2017, 12, e0173559. [Google Scholar] [CrossRef]
- Qaralleh, H. Thymol Rich Thymbra capitata Essential Oil Inhibits Quorum Sensing, Virulence and Biofilm Formation of Beta Lactamase Producing Pseudomonas aeruginosa. Nat. Prod. Sci. 2019, 25, 172–180. [Google Scholar] [CrossRef]
- Zhao, X.; Yu, Z.; Ding, T. Quorum-Sensing Regulation of Antimicrobial Resistance in Bacteria. Microorganisms 2020, 8, 425. [Google Scholar] [CrossRef] [PubMed]
- Tonkin, M.; Khan, S.; Wani, M.Y.; Ahmad, A. Quorum Sensing-a Stratagem for Conquering Multi-Drug Resistant Pathogens. Curr. Pharm. Des. 2021, 27, 2835–2847. [Google Scholar] [CrossRef]
- Khan, M.; Khan, M.; Alshareef, E.; Alaqeel, S.I.; Alkhathlan, H.Z. Chemical Characterization and Chemotaxonomic Significance of Essential Oil Constituents of Matricaria aurea Grown in Two Different Agro-Climatic Conditions. Plants 2023, 12, 3553. [Google Scholar] [CrossRef]
- Siddiqui, N.A. Chemical Constituents of Essential Oil from Flowers of Matricaria aurea Grown in Saudi Arabia. Indian J. Drugs 2014, 2, 164–168. [Google Scholar]
- Jalali, Z.; Sefidkon, F.; Assareh, M.H.; Attar, F. Comparison of Sesquiterpens in the Essential Oils of Anthemis Hyalina DC., Matricaria recutita L. and Matricaria aurea (Loefl.) Schultz-Bip. Iran. J. Med. Aromat. Plants Res. 2008, 24, 31–37. [Google Scholar]
- Yaz, H.H.; Uysal, I.; Polat, A.O.; Mohammed, F.S.; Sevindik, M. Usage Areas, Biological Activities and Volatile Oil Compounds of Matricaria aurea and Matricaria chamomilla. Lek. Sirovine 2023, 43, 1. [Google Scholar] [CrossRef]
- Rizwana, H.; Alwhibi, M.S.; Soliman, D.A. Research Article Antimicrobial Activity and Chemical Composition of Flowers of Matricaria aurea a Native Herb of Saudi Arabia. Int. J. Pharmacol. 2016, 12, 576–586. [Google Scholar] [CrossRef]
- Khammassi, M.; Polito, F.; Amri, I.; Khedhri, S.; Hamrouni, L.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii. Molecules 2022, 27, 5820. [Google Scholar] [CrossRef]
- Trinh, P.-C.; Thao, L.-T.-T.; Ha, H.-T.-V.; Nguyen, T. DPPH-Scavenging and Antimicrobial Activities of Asteraceae Medicinal Plants on Uropathogenic Bacteria. Evid.-Based Complement. Altern. Med. 2020, 2020, 7807026. [Google Scholar] [CrossRef] [PubMed]
- Perović, J.; Šaponjac, V.T.; Kojić, J.; Krulj, J.; Moreno, D.A.; García-Viguera, C.; Bodroža-Solarov, M.; Ilić, N. Chicory (Cichorium Intybus L.) as a Food Ingredient–Nutritional Composition, Bioactivity, Safety, and Health Claims: A Review. Food Chem. 2021, 336, 127676. [Google Scholar] [CrossRef]
- Sharifi, A.; Nayeri Fasaei, B. Selected Plant Essential Oils Inhibit Biofilm Formation and LuxS-and Pfs-mediated Quorum Sensing by Escherichia coli O157:H7. Lett. Appl. Microbiol. 2022, 74, 916–923. [Google Scholar] [CrossRef]
- Kalia, M.; Yadav, V.K.; Singh, P.K.; Sharma, D.; Pandey, H.; Narvi, S.S.; Agarwal, V. Effect of Cinnamon Oil on Quorum Sensing-Controlled Virulence Factors and Biofilm Formation in Pseudomonas aeruginosa. PLoS ONE 2015, 10, e0135495. [Google Scholar] [CrossRef]
- Emad, F.; Khalafalah, A.K.; El Sayed, M.A.; Mohamed, A.-E.H.; Stadler, M.; Helaly, S.E. Three New Polyacetylene Glycosides (PAGs) from the Aerial Part of Launaea capitata (Asteraceae) with Anti-Biofilm Activity against Staphylococcus aureus. Fitoterapia 2020, 143, 104548. [Google Scholar] [CrossRef]
- Bassey, K.; Mamabolo, P.; Cosa, S. An Andrographolide from Helichrysum caespitium (Dc.) Sond. Ex Harv., (Asteraceae) and Its Antimicrobial, Antiquorum Sensing, and Antibiofilm Potentials. Biology 2021, 10, 1224. [Google Scholar] [CrossRef]
- Masuku, M.; Mozirandi, W.; Mukanganyama, S. Evaluation of the Antibacterial and Antibiofilm Effects of Ethyl Acetate Root Extracts from Vernonia adoensis (Asteraceae) against Pseudomonas aeruginosa. Sci. World J. 2023, 2023, 5782656. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Stoodley, P.; Goeres, D.M.; Hall-Stoodley, L.; Burmølle, M.; Stewart, P.S.; Bjarnsholt, T. The Biofilm Life Cycle: Expanding the Conceptual Model of Biofilm Formation. Nat. Rev. Microbiol. 2022, 20, 608–620. [Google Scholar] [CrossRef]
- Brindhadevi, K.; LewisOscar, F.; Mylonakis, E.; Shanmugam, S.; Verma, T.N.; Pugazhendhi, A. Biofilm and Quorum Sensing Mediated Pathogenicity in Pseudomonas aeruginosa. Process Biochem. 2020, 96, 49–57. [Google Scholar] [CrossRef]
- Shrout, J.D.; Chopp, D.L.; Just, C.L.; Hentzer, M.; Givskov, M.; Parsek, M.R. The Impact of Quorum Sensing and Swarming Motility on Pseudomonas aeruginosa Biofilm Formation Is Nutritionally Conditional. Mol. Microbiol. 2006, 62, 1264–1277. [Google Scholar] [CrossRef]
- Copeland, M.F.; Weibel, D.B. Bacterial Swarming: A Model System for Studying Dynamic Self-Assembly. Soft Matter 2009, 5, 1174–1187. [Google Scholar] [CrossRef]
- O’May, C.; Tufenkji, N. The Swarming Motility of Pseudomonas aeruginosa Is Blocked by Cranberry Proanthocyanidins and Other Tannin-Containing Materials. Appl. Environ. Microbiol. 2011, 77, 3061–3067. [Google Scholar] [CrossRef]
- Kassinger, S.J.; van Hoek, M.L. Biofilm Architecture: An Emerging Synthetic Biology Target. Synth. Syst. Biotechnol. 2020, 5, 1–10. [Google Scholar] [CrossRef]
- McLean, R.J.C.; Pierson, L.S., III; Fuqua, C. A Simple Screening Protocol for the Identification of Quorum Signal Antagonists. J. Microbiol. Methods 2004, 58, 351–360. [Google Scholar] [CrossRef]
- López-Jácome, L.E.; Garza-Ramos, G.; Hernández-Durán, M.; Franco-Cendejas, R.; Loarca, D.; Romero-Martínez, D.; Nguyen, P.T.D.; Maeda, T.; González-Pedrajo, B.; Díaz-Guerrero, M. AiiM Lactonase Strongly Reduces Quorum Sensing Controlled Virulence Factors in Clinical Strains of Pseudomonas aeruginosa Isolated from Burned Patients. Front. Microbiol. 2019, 10, 2657. [Google Scholar] [CrossRef] [PubMed]
- Shravani, V.; Selvi, G.A.S.; Mantravadi, H. Detection of Quorum Sensing Virulence Factor Genes and Its Consanguinity to Antibiotic Sensitivity Profile in the Clinical Isolates of Pseudomonas aeruginosa. Iran. J. Basic Med. Sci. 2023, 26, 899. [Google Scholar] [PubMed]
- Danaraj, J.; Mariasingarayan, Y.; Ayyappan, S.; Karuppiah, V. Seagrass Halodule Pinifolia Active Constituent 4-Methoxybenzioic Acid (4-MBA) Inhibits Quorum Sensing Mediated Virulence Production of Pseudomonas aeruginosa. Microb. Pathog. 2020, 147, 104392. [Google Scholar] [CrossRef]
- Karatuna, O.; Yagci, A. Analysis of Quorum Sensing-Dependent Virulence Factor Production and Its Relationship with Antimicrobial Susceptibility in Pseudomonas aeruginosa Respiratory Isolates. Clin. Microbiol. Infect. 2010, 16, 1770–1775. [Google Scholar] [CrossRef]
- O’Malley, Y.Q.; Reszka, K.J.; Spitz, D.R.; Denning, G.M.; Britigan, B.E. Pseudomonas aeruginosa Pyocyanin Directly Oxidizes Glutathione and Decreases Its Levels in Airway Epithelial Cells. Am. J. Physiol. Cell. Mol. Physiol. 2004, 287, L94–L103. [Google Scholar] [CrossRef]
- Rather, M.A.; Hassan, T.; Dar, B.A.; Shawl, A.S.; Qurishi, M.A.; Ganai, B.A. Essential Oil Composition of Nepeta Raphanorhiza Benth Growing in Kashmir Valley. Rec. Nat. Prod. 2012, 6, 67. [Google Scholar]
- Ashraf, M.I.; Ong, S.-K.; Mujawar, S.; Pawar, S.; More, P.; Paul, S.; Lahiri, C. A Side-Effect Free Method for Identifying Cancer Drug Targets. Sci. Rep. 2018, 8, 6669. [Google Scholar] [CrossRef]
- Pesci, E.C.; Pearson, J.P.; Seed, P.C.; Iglewski, B.H. Regulation of Las and Rhl Quorum Sensing in Pseudomonas aeruginosa. J. Bacteriol. 1997, 179, 3127–3132. [Google Scholar] [CrossRef]
- Soukarieh, F.; Williams, P.; Stocks, M.J.; Camara, M. Pseudomonas aeruginosa Quorum Sensing Systems as Drug Discovery Targets: Current Position and Future Perspectives. J. Med. Chem. 2018, 61, 10385–10402. [Google Scholar] [CrossRef]
- Ueda, A.; Wood, T.K. Connecting Quorum Sensing, c-Di-GMP, Pel Polysaccharide, and Biofilm Formation in Pseudomonas aeruginosa through Tyrosine Phosphatase TpbA (PA3885). PLoS Pathog. 2009, 5, e1000483. [Google Scholar] [CrossRef]
- Muimhneacháin, E.Ó.; Reen, F.J.; O’Gara, F.; McGlacken, G.P. Analogues of Pseudomonas aeruginosa Signalling Molecules to Tackle Infections. Org. Biomol. Chem. 2018, 16, 169–179. [Google Scholar] [CrossRef]
- Yan, S.; Wu, G. Can Biofilm Be Reversed through Quorum Sensing in Pseudomonas aeruginosa? Front. Microbiol. 2019, 10, 1582. [Google Scholar] [CrossRef]
- Turkina, M.V.; Vikström, E. Bacteria-Host Crosstalk: Sensing of the Quorum in the Context of Pseudomonas aeruginosa Infections. J. Innate Immun. 2019, 11, 263–279. [Google Scholar] [CrossRef]
- Lin, J.; Cheng, J.; Wang, Y.; Shen, X. The Pseudomonas quinolone Signal (PQS): Not Just for Quorum Sensing Anymore. Front. Cell. Infect. Microbiol. 2018, 8, 230. [Google Scholar] [CrossRef]
- Mellbye, B.; Schuster, M. Physiological Framework for the Regulation of Quorum Sensing-Dependent Public Goods in Pseudomonas aeruginosa. J. Bacteriol. 2014, 196, 1155–1164. [Google Scholar] [CrossRef]
- Pezzoni, M.; Pizarro, R.A.; Costa, C.S. Role of Quorum Sensing in UVA-Induced Biofilm Formation in Pseudomonas aeruginosa. Microbiology 2020, 166, 735–750. [Google Scholar] [CrossRef]
- Ryder, C.; Byrd, M.; Wozniak, D.J. Role of Polysaccharides in Pseudomonas aeruginosa Biofilm Development. Curr. Opin. Microbiol. 2007, 10, 644–648. [Google Scholar] [CrossRef]
- Ma, L.; Wang, J.; Wang, S.; Anderson, E.M.; Lam, J.S.; Parsek, M.R.; Wozniak, D.J. Synthesis of Multiple Pseudomonas aeruginosa Biofilm Matrix Exopolysaccharides Is Post-transcriptionally Regulated. Environ. Microbiol. 2012, 14, 1995–2005. [Google Scholar] [CrossRef]
- Kreuger, M.R.O.; Grootjans, S.; Biavatti, M.W.; Vandenabeele, P.; D’Herde, K. Sesquiterpene Lactones as Drugs with Multiple Targets in Cancer Treatment: Focus on Parthenolide. Anticancer Drugs 2012, 23, 883–896. [Google Scholar] [CrossRef]
- Owlia, P.; Rasooli, I.; Saderi, H.; Aliahmadi, M. Retardation of Biofilm Formation with Reduced Productivity of Alginate as a Result of Pseudomonas aeruginosa Exposure to Matricaria chamomilla Essential Oil. Pharmacogn. Mag. 2007, 3, 83–89. [Google Scholar]
- Sethupathy, S.; Shanmuganathan, B.; Kasi, P.D.; Karutha Pandian, S. Alpha-Bisabolol from Brown Macroalga padina Gymnospora Mitigates Biofilm Formation and Quorum Sensing Controlled Virulence Factor Production in Serratia marcescens. J. Appl. Phycol. 2016, 28, 1987–1996. [Google Scholar] [CrossRef]
- Wallace, W.E. NIST Chemistry WebBook, NIST Standard Reference Database, 69th ed.; Linstrom, P.J., Mallard, W.G., Eds.; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2014. [Google Scholar]
- Dmour, S.M.; Qaralleh, H.; Al-Limoun, M.; Khleifat, K.M.; Alqaraleh, M.; Alqudah, A.A.; Altarawneh, R.M. Combined Antibacterial Activity of Eucalyptol, γ-Terpinene, p-Cymol and Punicalagin with Cefotaxime against Methicillin (Oxacillin) Resistant Staphylococcus aureus Isolate. Res. J. Pharm. Technol. 2022, 15, 3905–3911. [Google Scholar] [CrossRef]
- Al Assi, G.; Al-Bashaereh, A.; Alsarayreh, A.; Al Qaisi, Y.; Al-Majali, I.; Khleifat, K.; Alqaraleh, M.; Qaralleh, H.; Al-Farrayeh, I. Evaluation of Antibacterial, Antioxidant and Anti-Inflammatory Properties of Methanol Extract of Varthemia iphionoides. Trop. J. Nat. Prod. Res. 2023, 7, 2107–2114. [Google Scholar]
- Gómez-Gómez, B.; Arregui, L.; Serrano, S.; Santos, A.; Pérez-Corona, T.; Madrid, Y. Selenium and Tellurium-Based Nanoparticles as Interfering Factors in Quorum Sensing-Regulated Processes: Violacein Production and Bacterial Biofilm Formation. Metallomics 2019, 11, 1104–1114. [Google Scholar] [CrossRef]
- Sagar, P.K.; Sharma, P.; Singh, R. Inhibition of Quorum Sensing Regulated Virulence Factors and Biofilm Formation by Eucalyptus Globulus against Multidrug-Resistant Pseudomonas aeruginosa. J. Pharmacopunct. 2022, 25, 37. [Google Scholar] [CrossRef]
- Shanks, R.M.Q.; Meehl, M.A.; Brothers, K.M.; Martinez, R.M.; Donegan, N.P.; Graber, M.L.; Cheung, A.L.; O’Toole, G.A. Genetic Evidence for an Alternative Citrate-Dependent Biofilm Formation Pathway in Staphylococcus aureus That Is Dependent on Fibronectin Binding Proteins and the GraRS Two-Component Regulatory System. Infect. Immun. 2008, 76, 2469–2477. [Google Scholar] [CrossRef]
- Krishnan, T.; Yin, W.-F.; Chan, K.-G. Inhibition of Quorum Sensing-Controlled Virulence Factor Production in Pseudomonas aeruginosa PAO1 by Ayurveda Spice Clove (Syzygium aromaticum) Bud Extract. Sensors 2012, 12, 4016–4030. [Google Scholar] [CrossRef]
- Razack, S.A.; Vijayagopal, V.; Viruthagiri, T. Production of Exopolysaccharides by P. aeruginosa, P. fluorescens and Bacillus sp. IUP J. Biotechnol. 2011, 5, 14. [Google Scholar]
- Hossain, M.A.; Lee, S.-J.; Park, N.-H.; Mechesso, A.F.; Birhanu, B.T.; Kang, J.; Reza, M.A.; Suh, J.-W.; Park, S.-C. Impact of Phenolic Compounds in the Acyl Homoserine Lactone-Mediated Quorum Sensing Regulatory Pathways. Sci. Rep. 2017, 7, 10618. [Google Scholar] [CrossRef]
- Luo, J.; Dong, B.; Wang, K.; Cai, S.; Liu, T.; Cheng, X.; Lei, D.; Chen, Y.; Li, Y.; Kong, J. Baicalin Inhibits Biofilm Formation, Attenuates the Quorum Sensing-Controlled Virulence and Enhances Pseudomonas aeruginosa Clearance in a Mouse Peritoneal Implant Infection Model. PLoS ONE 2017, 12, e0176883. [Google Scholar] [CrossRef]
- Andrejko, M.; Zdybicka-Barabas, A.; Janczarek, M.; Cytryńska, M. Three Pseudomonas aeruginosa Strains with Different Protease Profiles. Acta Biochim. Pol. 2013, 60, 83–90. [Google Scholar] [CrossRef]
- Lee, J.; Lee, I.; Nam, J.; Hwang, D.S.; Yeon, K.-M.; Kim, J. Immobilization and Stabilization of Acylase on Carboxylated Polyaniline Nanofibers for Highly Effective Antifouling Application via Quorum Quenching. ACS Appl. Mater. Interfaces 2017, 9, 15424–15432. [Google Scholar] [CrossRef]
- Lenz, A.P.; Williamson, K.S.; Pitts, B.; Stewart, P.S.; Franklin, M.J. Localized Gene Expression in Pseudomonas aeruginosa Biofilms. Appl. Environ. Microbiol. 2008, 74, 4463–4471. [Google Scholar] [CrossRef]
- Li, R.; Yuan, X.; Wei, J.; Zhang, X.; Cheng, G.; Wang, Z.A.; Du, Y. Synthesis and Evaluation of a Chitosan Oligosaccharide-Streptomycin Conjugate against Pseudomonas aeruginosa Biofilms. Mar. Drugs 2019, 17, 43. [Google Scholar] [CrossRef]
- Irie, Y.; Borlee, B.R.; O’Connor, J.R.; Hill, P.J.; Harwood, C.S.; Wozniak, D.J.; Parsek, M.R. Self-Produced Exopolysaccharide Is a Signal That Stimulates Biofilm Formation in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2012, 109, 20632–20636. [Google Scholar] [CrossRef]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rasamiravaka, T.; Stevigny, C.; Duez, P.; Rajaonson, S.; Diallo, B.; Mol, A.; Baucher, M.; El Jaziri, M. The Flavanone Naringenin Reduces the Production of Quorum Sensing-Controlled Virulence Factors in Pseudomonas aeruginosa PAO1. Microbiology 2011, 157, 2120–2132. [Google Scholar] [CrossRef]
- Cabrol, S.; Olliver, A.; Pier, G.B.; Andremont, A.; Ruimy, R. Transcription of Quorum-Sensing System Genes InClinical and Environmental Isolates of Pseudomonas aeruginosa. J. Bacteriol. 2003, 185, 7222–7230. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
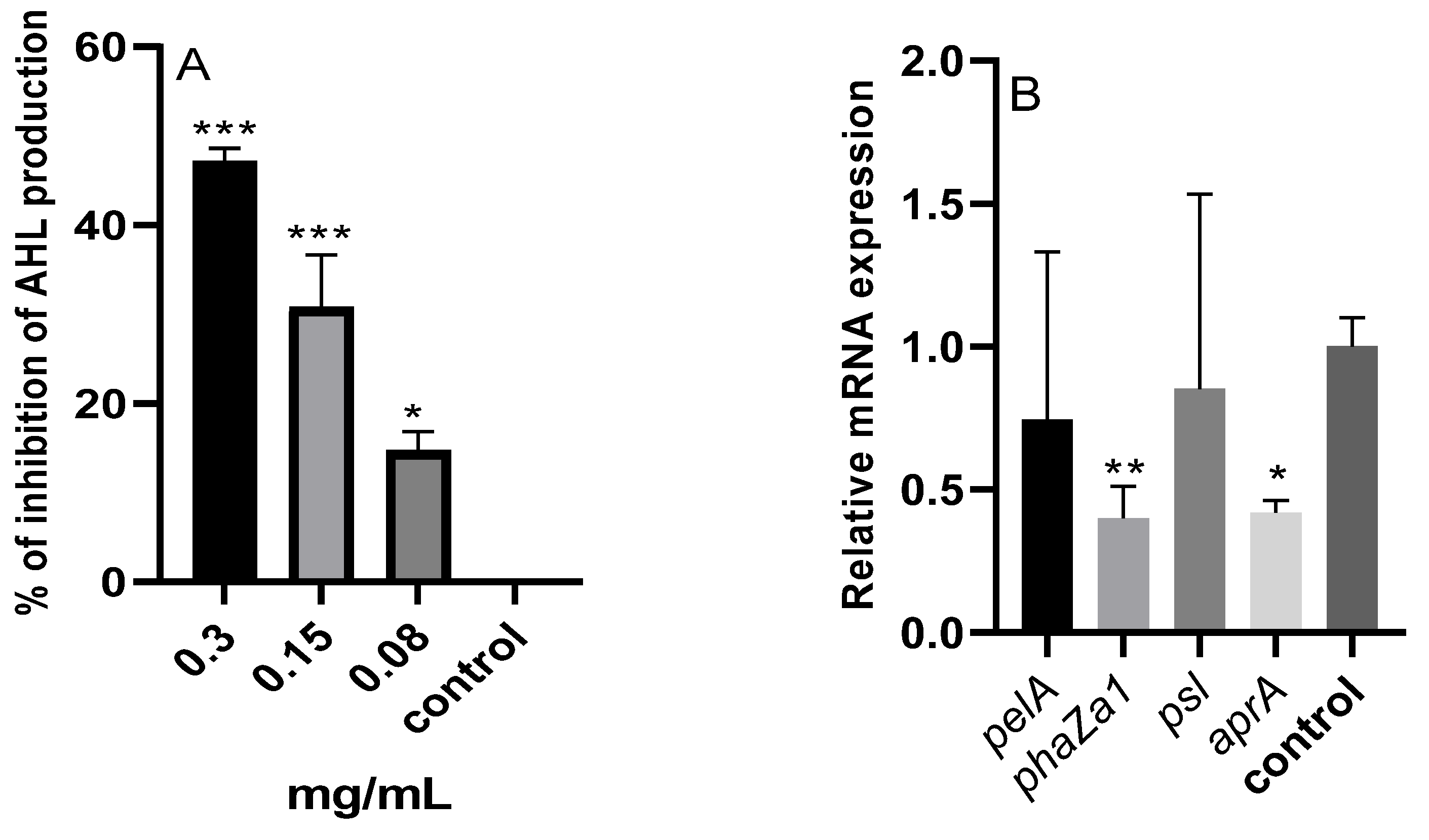{kind=link}
| Peak | Compound | Kical | Kilet | Conc (%) | Method of Identification |
|---|---|---|---|---|---|
| 1 | heptanol | 970 | 935–977 | 0.6 | RI, MS |
| 2 | β-pinene | 978 | 959–982 | 0.3 | RI, MS |
| 3 | 1-octen-3-ol | 980 | 964–986 | 0.3 | RI, MS |
| 4 | α-terpinene | 1010 | 1012–1026 | 0.4 | RI, MS, Co-injection |
| 5 | limonene | 1029 | 995–1039 | 0.2 | RI, MS, Co-injection |
| 6 | 3-octen-2-one | 1037 | 1037–1048 | 0.5 | RI, MS |
| 7 | phenylacetaldehyde | 1043 | 1039–1049 | 0.5 | RI, MS |
| 8 | (E)-2-octenal | 1052 | 1049–1064 | 0.5 | RI, MS |
| 9 | octanol | 1074 | 1068–1078 | 0.3 | RI, MS |
| 10 | terpinolene | 1089 | 1081–1097 | 0.2 | RI, MS |
| 11 | 2-nonanone | 1092 | 1090–1096 | 0.5 | RI, MS |
| 12 | nonanal | 1102 | 1087–1104 | 0.5 | RI, MS |
| 13 | menthone | 1148 | 1124–1155 | 0.3 | RI, MS |
| 14 | hexyl butyrate | 1190 | 1178–1194 | 0.5 | RI, MS |
| 15 | caprylic acid | 1191 | 1170–1192 | 0.2 | RI, MS |
| 16 | octyl acetate | 1200 | 1195–1215 | 1.0 | RI, MS |
| 17 | (E)-2-decenal | 1260 | 1234–1267 | 1.7 | RI, MS |
| 18 | (E, E)-2,4-decadienal | 1318 | 1312–1345 | 0.5 | RI, MS |
| 19 | octyl isobutyrate | 1345 | 1336–1348 | 0.3 | RI, MS |
| 20 | α-copaene | 1372 | 1364–1380 | 0.5 | RI, MS |
| 21 | β-caryophyllene | 1415 | 1394–1451 | 0.4 | RI, MS, Co-injection |
| 22 | α-humulene | 1455 | 1442–1488 | 0.7 | RI, MS, Co-injection |
| 23 | trans-β-farnesene | 1457 | 1456–1461 | 0.5 | RI, MS |
| 24 | Germacrene D | 1474 | 1468–1519 | 1.9 | RI, MS |
| 25 | cuparene | 1504 | 1498–1516 | 0.9 | RI, MS |
| 26 | α-farnesene | 1507 | 1505–1524 | 4.8 | RI, MS |
| 27 | (E, E)-α-Farnesene | 1507 | 1499–1511 | 1.5 | RI, MS |
| 28 | β-bisabolene | 1510 | 1489–1512 | 6.3 | RI, MS |
| 29 | myristicin | 1520 | 1509–1526 | 1.2 | RI, MS, Co-injection |
| 30 | spathulenol | 1576 | 1557–1580 | 0.5 | RI, MS |
| 31 | humulene epoxide II | 1606 | 1601–1620 | 0.7 | RI, MS |
| 32 | isopropyl dodecanoate | 1626 | 1614–1629 | 1.0 | RI, MS |
| 33 | α-Bisabolol | 1682 | 1680–1704 | 0.4 | RI, MS |
| 34 | α-Bisabolol oxide A | 1745 | 1672–1759 | 64.8 | RI, MS |
| 35 | hexahydrofarnesyl acetone | 1830 | 1827–1856 | 0.4 | RI, MS |
| Monoterpenes hydrocarbon | 1.1 | ||||
| Oxygenated monoterpenes | 0.3 | ||||
| Sesquiterpene hydrocarbons | 17.5 | ||||
| Oxygenated sesquiterpenes | 66.8 | ||||
| Others | 10.1 | ||||
| Total | 95.8 |
| Tested material | Inhibition zone (MIC) |
| M. aurea EO (1 mg) | 12.8 ± 0.3 mm (1.25 mg/mL) |
| Erythromycin (10 µg) | 10.33 ± 0.58 |
| DMSO | 0.0 ± 0.0 |
| Gene | Primer Sequence 5′-3′ | References | |
|---|---|---|---|
| 16S rRNA | Forward | CAAAACTACTGAGCTAGAGTACG | [84] |
| Reverse | TAAGATCTCAAGGATCCCAACGGCT | ||
| PelA | Forward | CCTTCAGCCATCCGTTCTTCT | [85] |
| Reverse | TCGCGTACGAAGTCGACCTT | ||
| PslA | Forward | AAGATCAAGAAACGCGTGGAAT | [86] |
| Reverse | TGTAGAGGTCGAACCACACCG | ||
| PhazA | Forward | CGAACCACTTCTGGGTCGAGTGC | [87] |
| Reverse | GGGAATACCGTCACGTTTTATTTGC | ||
| AprA | Forward | GGCAATCCTGGTACCTGATCAA | [88] |
| Reverse | AGCGTCTGGCGCCCGTAGTT | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qaralleh, H.; Saghir, S.A.M.; Al-limoun, M.O.; Dmor, S.M.; Khleifat, K.; Al-Ahmad, B.E.M.; Al-Omari, L.; Tabana, Y.; Mothana, R.A.; Al-Yousef, H.M.; et al. Effect of Matricaria aurea Essential Oils on Biofilm Development, Virulence Factors and Quorum Sensing-Dependent Genes of Pseudomonas aeruginosa. Pharmaceuticals 2024, 17, 386. https://doi.org/10.3390/ph17030386
Qaralleh H, Saghir SAM, Al-limoun MO, Dmor SM, Khleifat K, Al-Ahmad BEM, Al-Omari L, Tabana Y, Mothana RA, Al-Yousef HM, et al. Effect of Matricaria aurea Essential Oils on Biofilm Development, Virulence Factors and Quorum Sensing-Dependent Genes of Pseudomonas aeruginosa. Pharmaceuticals. 2024; 17(3):386. https://doi.org/10.3390/ph17030386
Chicago/Turabian StyleQaralleh, Haitham, Sultan Ayesh Mohammed Saghir, Muhamad O. Al-limoun, Saif M. Dmor, Khaled Khleifat, Basma Ezzat Mustafa Al-Ahmad, Laila Al-Omari, Yasser Tabana, Ramzi A. Mothana, Hanan M. Al-Yousef, and et al. 2024. "Effect of Matricaria aurea Essential Oils on Biofilm Development, Virulence Factors and Quorum Sensing-Dependent Genes of Pseudomonas aeruginosa" Pharmaceuticals 17, no. 3: 386. https://doi.org/10.3390/ph17030386
APA StyleQaralleh, H., Saghir, S. A. M., Al-limoun, M. O., Dmor, S. M., Khleifat, K., Al-Ahmad, B. E. M., Al-Omari, L., Tabana, Y., Mothana, R. A., Al-Yousef, H. M., & Alqahtani, A. M. (2024). Effect of Matricaria aurea Essential Oils on Biofilm Development, Virulence Factors and Quorum Sensing-Dependent Genes of Pseudomonas aeruginosa. Pharmaceuticals, 17(3), 386. https://doi.org/10.3390/ph17030386