
Potential of Lycii Radicis Cortex as an Ameliorative Agent for Skeletal Muscle Atrophy

 ,
,
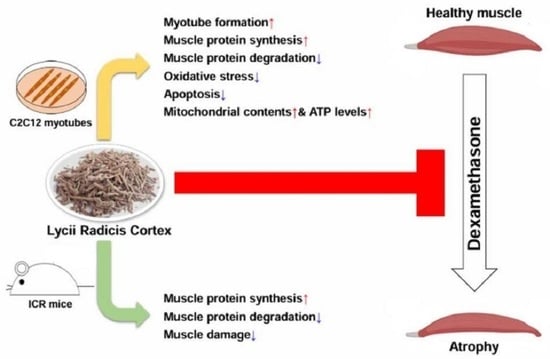
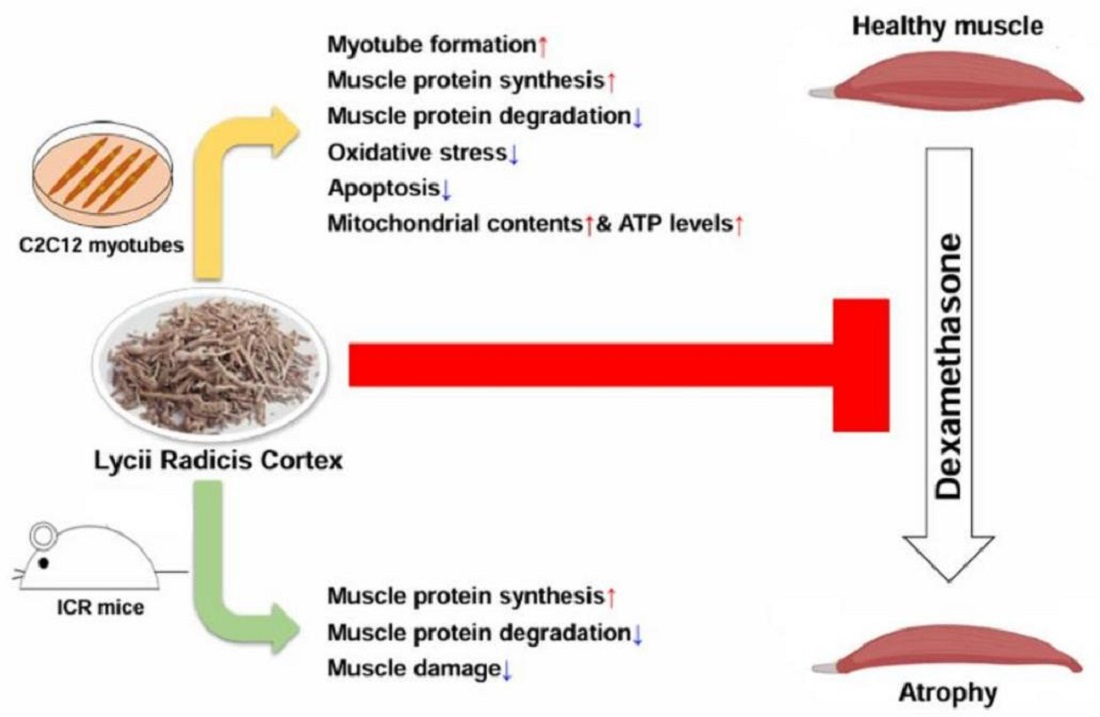
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
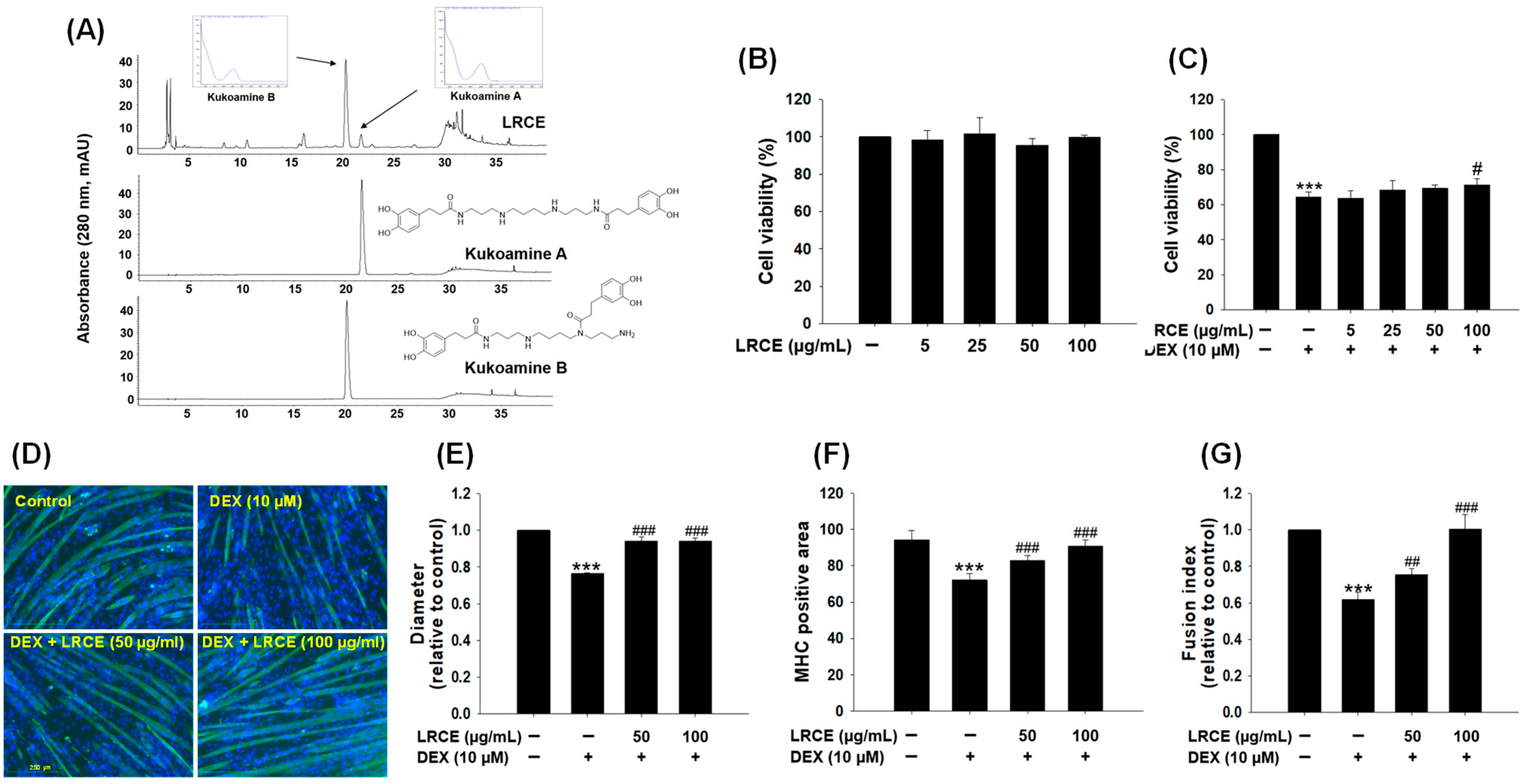
2.1. Effects of LRCE on Viability and Atrophy in DEX-Treated C2C12 Myotubes
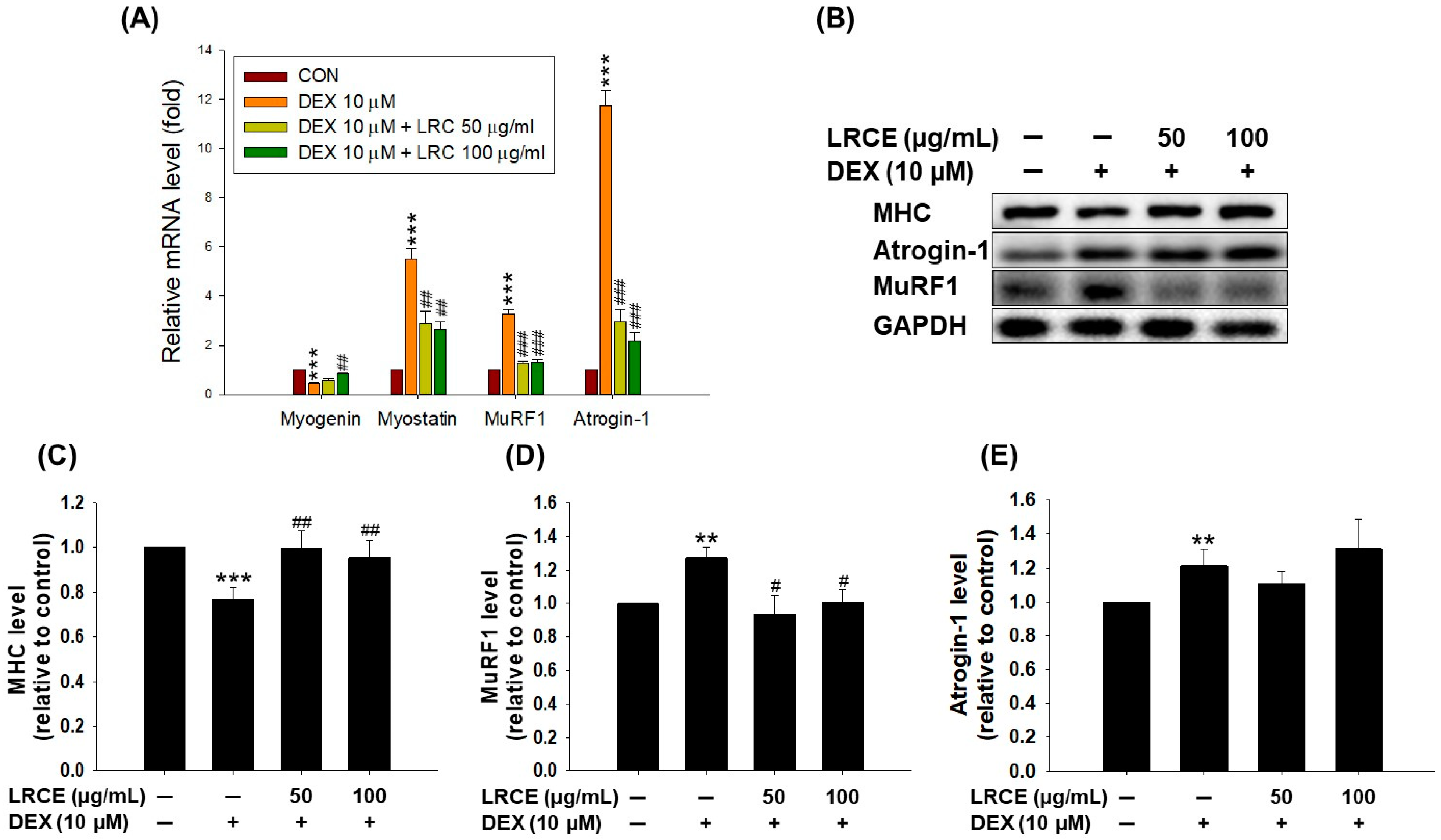
2.2. Effects of LRCE on the Expression of Muscle Atrophy-Related Factors in DEX-Treated C2C12 Myotubes
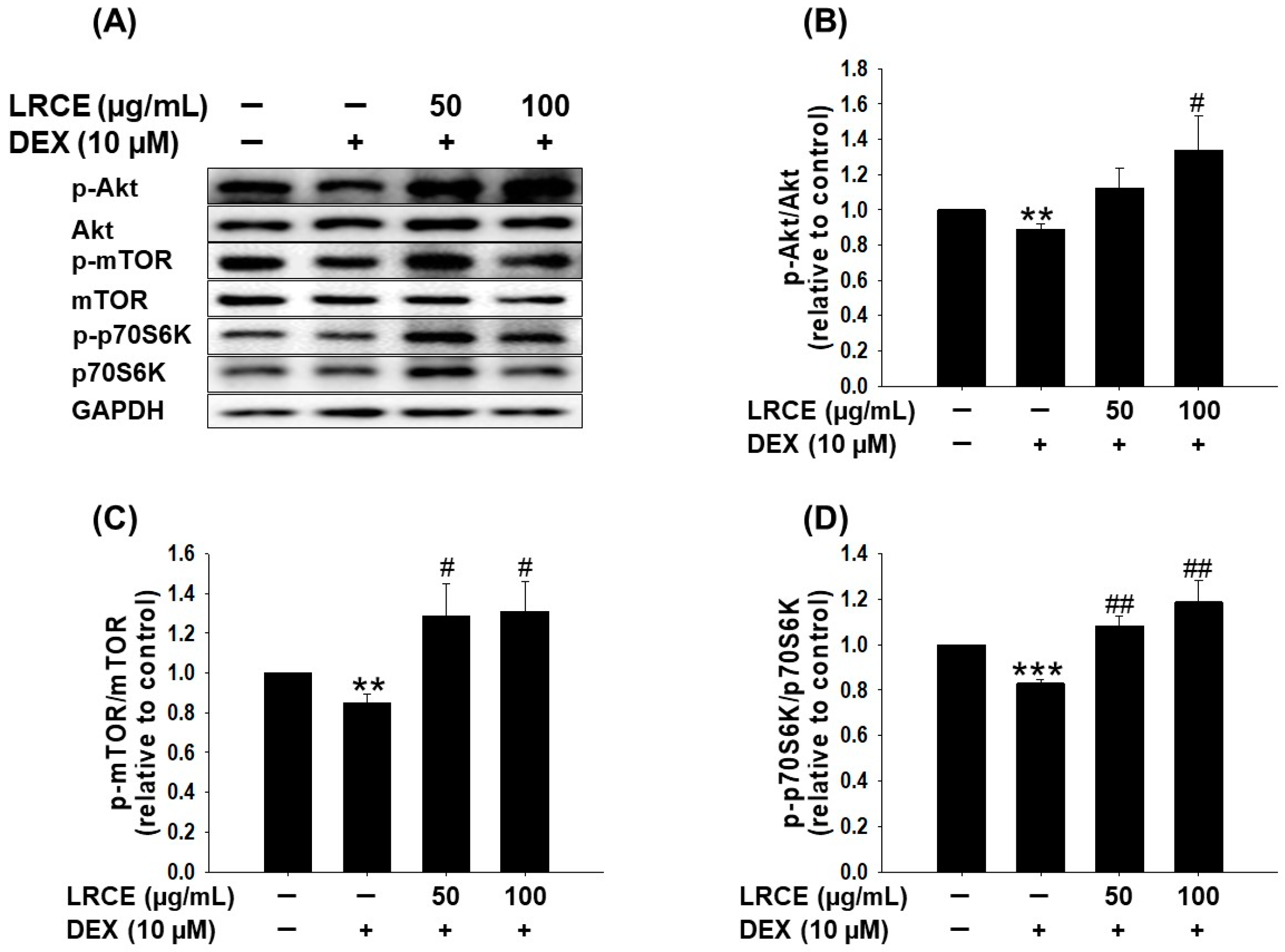
2.3. Effects of LRCE on the Akt/mTOR/p70S6K Pathway in DEX-Treated C2C12 Myotubes
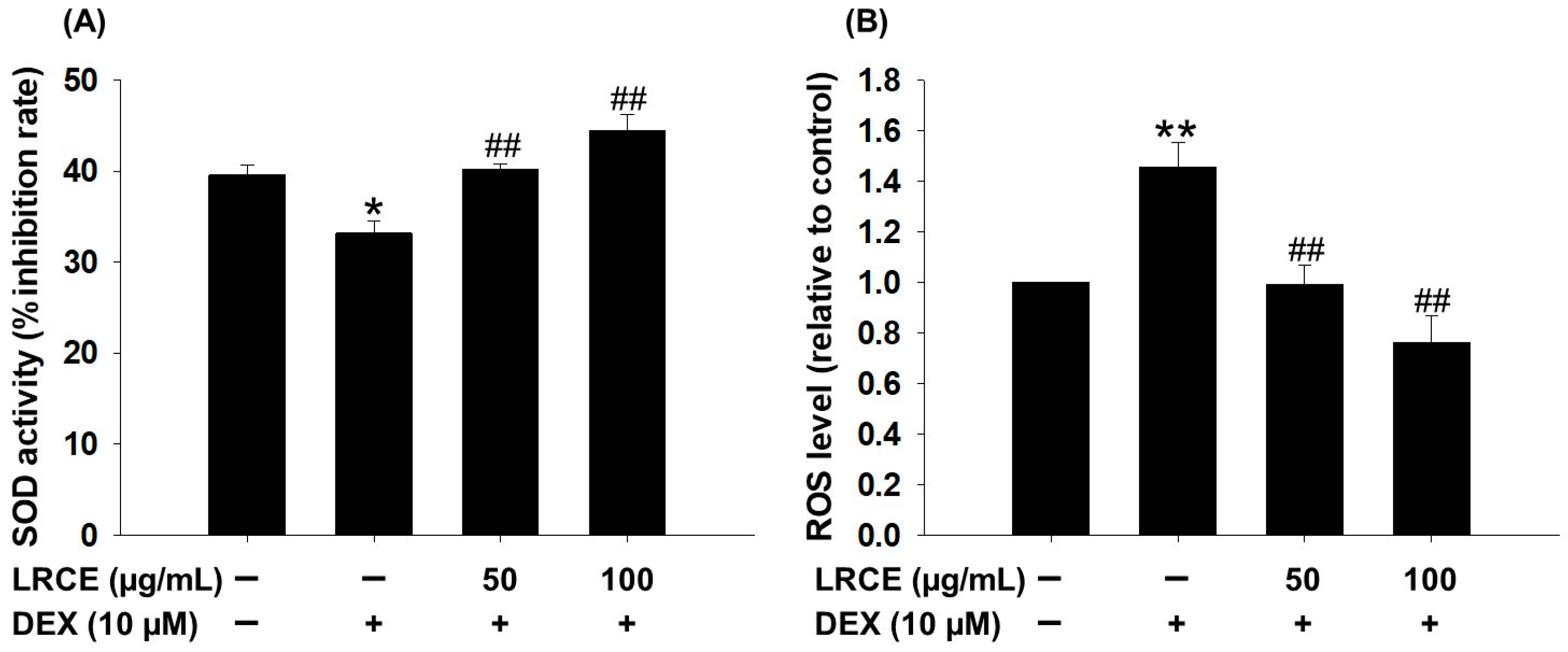
2.4. Effects of LRCE on the SOD Activity and ROS Production in DEX-Treated C2C12 Myotubes
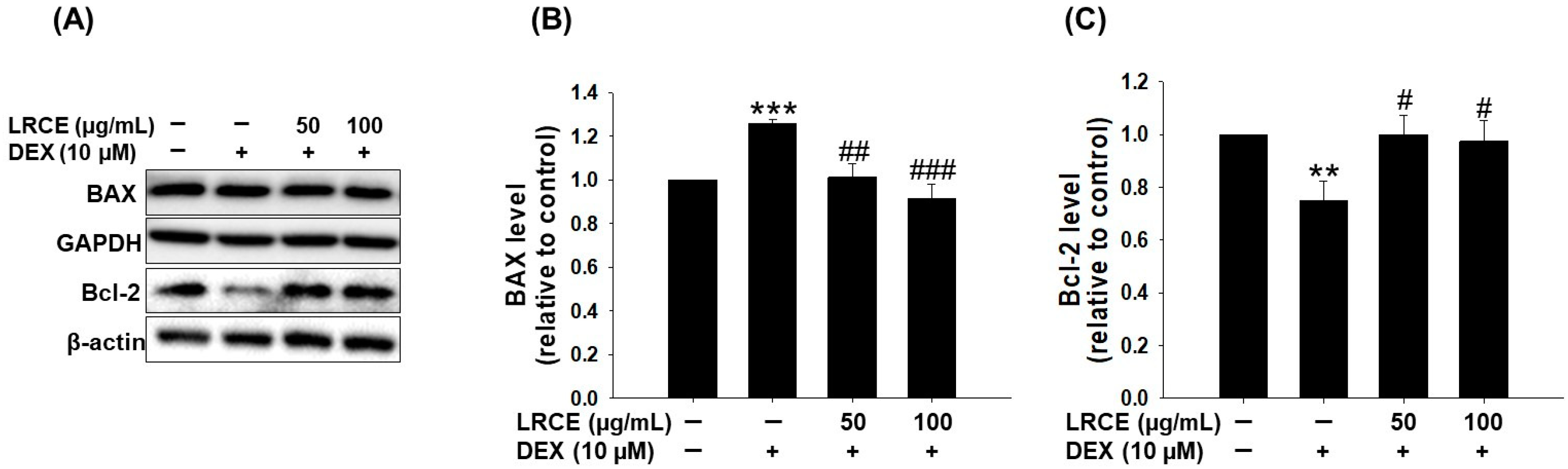
2.5. Effects of LRCE on the Expression of BAX and Bcl-2 in DEX-Treated C2C12 Myotubes
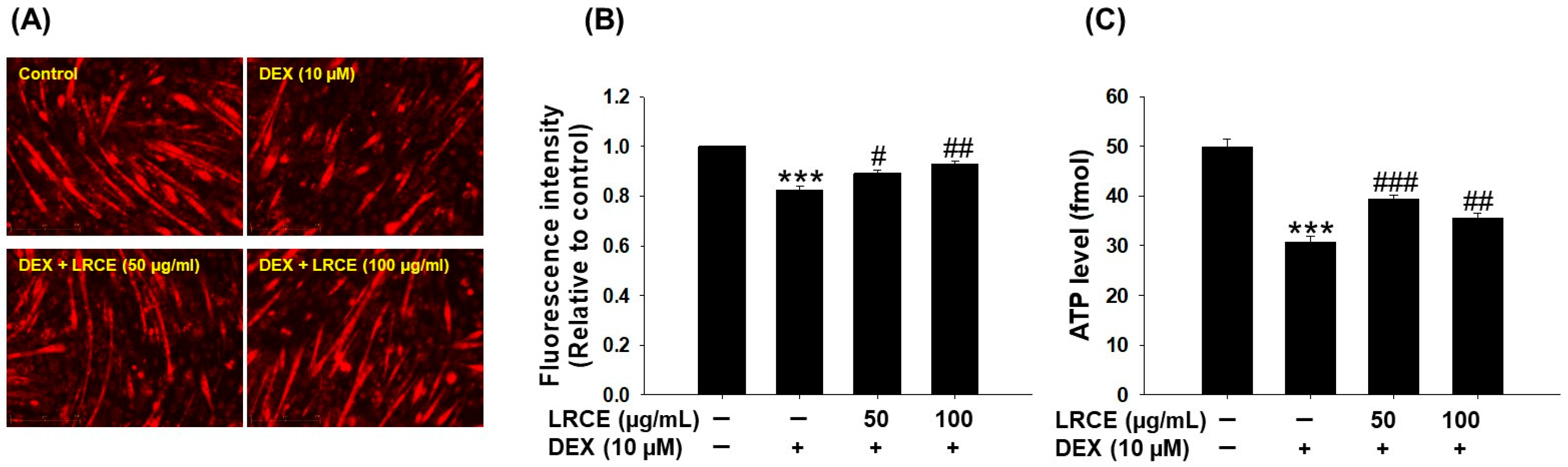
2.6. Effects of LRCE on Mitochondrial Content and ATP Production in DEX-Treated C2C12 Myotubes
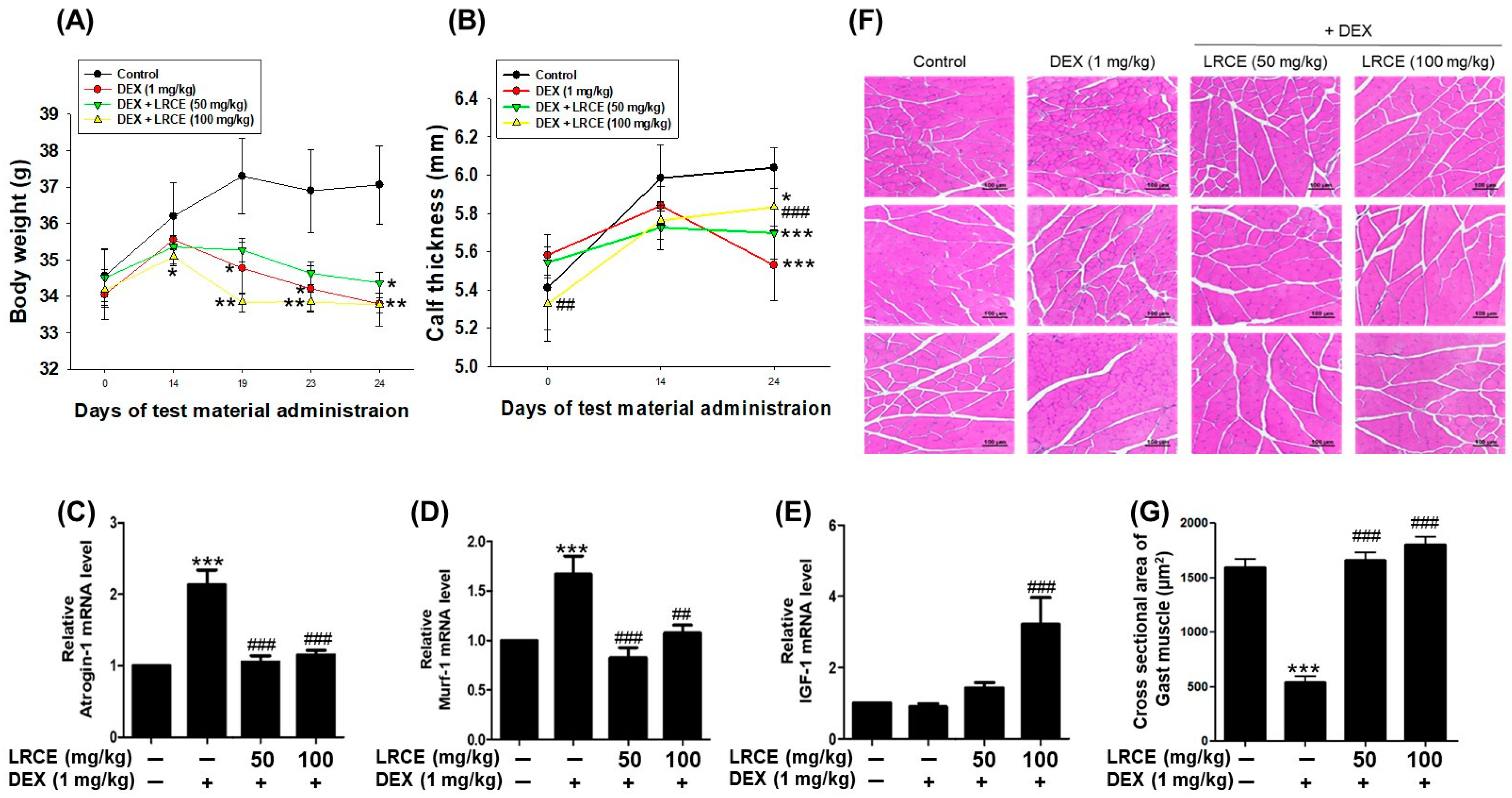
2.7. Effects of LRCE on Body Weight, Calf Thickness, and Gastrocnemius Muscle in DEX-Treated Mice
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Sample Preparation and Optimization of HPLC Analysis
4.3. C2C12 Cell Culture and Differentiation
4.4. Treatment of LRCE and DEX
4.5. Cell Viability Assay
4.6. Myosin Heavy Chain (MHC) Immunofluorescence Staining
4.7. Quantitative Real-Time PCR
4.8. Westerm Blot Analysis
4.9. Measurement of SOD Activity
4.10. Measurement of Intracellular ROS
4.11. Mitochondria Staining
4.12. Determination of ATP Level
4.13. Mouse Preparation and Treatment
4.14. Histological Analysis of Gastrocnemius Muscle
4.15. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Iizuka, K.; Machida, T.; Hirafuji, M. Skeletal muscle is an endocrine organ. J. Pharmacol. Sci. 2014, 125, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Feraco, A.; Gorini, S.; Armani, A.; Camajani, E.; Rizzo, M.; Caprio, M. Exploring the Role of skeletal Muscle in Insulin Resistance: Lessons from Cultured Cells to Animal Models. Int. J. Mol. Sci. 2021, 22, 9327. [Google Scholar] [CrossRef]
- Takada, S.; Sabe, H.; Kinugawa, S. Abnormalities of Skeletal Muscle, Adipocyte Tissue, and Lipid Metabolism in Heart Failure: Practical Therapeutic Targets. Front. Cardiovasc. Med. 2020, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Pauk, M.; Saito, H.; Hesse, E.; Taipaleenmäki, H. Muscle and Bone Defects in Metastatic Disease. Curr. Osteoporos. Rep. 2022, 20, 273–289. [Google Scholar] [CrossRef]
- Mirzoev, T.M. Skeletal Muscle Recovery from Disuse Atrophy: Protein Turnover Signaling Andstrategies for Accelerating Muscle Regrowth. Int. J. Mol. Sci. 2020, 21, 7940. [Google Scholar] [CrossRef]
- Jackman, R.W.; Kandarian, S.C. The Molecular Basis of Skeletal Muscle Atrophy. Am. J. Physiol. Cell Physiol. 2004, 287, C834–C843. [Google Scholar] [CrossRef] [PubMed]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of Muscle Atrophy and Hypertrophy: Implications in Health and Disease. Nat. Commun. 2021, 12, 330. [Google Scholar] [CrossRef] [PubMed]
- Santilli, V.; Bernetti, A.; Mangone, M.; Paoloni, M. Clinical definition of sarcopenia. Clin. Cases Miner. Bone Metab. 2014, 11, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Muscaritoli, M.; Lucia, S.; Molfino, A.; Cederholm, T.; Rossi Fanelli, F. Muscle atrophy in aging and chronic diseases: Is it sarcopenia or cachexia? Intern. Emerg. Med. 2013, 8, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Tey, S.L.; Huynh, D.T.T.; Berde, Y.; Baggs, G.; How, C.H.; Low, Y.L.; Cheong, M.; Chow, W.L.; Tan, N.C.; Chew, S.T.H. Prevalence of low muscle mass and associated factors in community-dwelling older adults in Singapore. Sci. Rep. 2021, 11, 23071. [Google Scholar] [CrossRef] [PubMed]
- Schakman, O.; Kalista, S.; Barbé, C.; Loumaye, A.; Thissen, J.P. Glucocorticoid-induced skeletal muscle atrophy. Int. J. Biochem. Cell Biol. 2013, 45, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Dubashynskaya, N.V.; Bokatyi, A.N.; Skorik, Y.A. Dexamethasone Conjugates: Synthetic Approaches and Medical Prospects. Biomedicines 2021, 9, 341. [Google Scholar] [CrossRef] [PubMed]
- Cea, L.A.; Balboa, E.; Puebla, C.; Vargas, A.A.; Cisterna, B.A.; Escamilla, R.; Regueira, T.; Sáez, J.C. Dexamethasone-induced muscular atrophy is mediated by functional expression of connexin-based hemichannels. Biochim. Biophys. Acta 2016, 1862, 1891–1899. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Baehr, L.M. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef] [PubMed]
- Schakman, O.; Gilson, H.; Thissen, J.P. Mechanisms of glucocorticoid-induced myopathy. J. Endocrinol. 2008, 197, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.S. mTOR as a Key Regulator in Maintaining Skeletal Muscle Mass. Front. Physiol. 2017, 8, 788. [Google Scholar] [CrossRef] [PubMed]
- Go, G.Y.; Lee, S.J.; Jo, A.; Lee, J.; Seo, D.W.; Kang, J.S.; Kim, S.K.; Kim, S.N.; Kim, Y.K.; Bae, G.U. Ginsenoside Rg1 from Panax ginseng enhances myoblast differentiation and myotube growth. J. Ginseng Res. 2017, 41, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Ku, S.K.; Han, M.H.; Kim, K.Y.; Kim, S.G.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Kim, C.M.; Choi, Y.H. The administration of Fructus Schisandrae attenuates dexamethasone-induced muscle atrophy in mice. Int. J. Mol. Med. 2015, 36, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Q.; Quan, H.; Kang, S.G.; Huang, K.; Tong, T. Nutraceuticals in the Prevention and Treatment of the Muscle Atrophy. Nutrients 2021, 3, 1914. [Google Scholar] [CrossRef]
- Xie, L.W.; Atanasov, A.G.; Guo, D.A.; Malainer, C.; Zhang, J.X.; Zehl, M.; Guan, S.H.; Heiss, E.H.; Urban, E.; Dirsch, V.M.; et al. Activity-guided isolation of NF-κB inhibitors and PPARγ agonists from the root bark of Lycium chinense Miller. J. Ethnopharmacol. 2014, 152, 470–477. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, E.Y.; Lee, B.; Min, J.H.; Song, D.U.; Lim, J.M.; Eom, J.W.; Yeom, M.; Jung, H.S.; Sohn, Y. The effects of Lycii Radicis Cortex on RANKL-induced osteoclast differentiation and activation in RAW 264.7 cells. Int. J. Mol. Med. 2016, 37, 649–658. [Google Scholar] [CrossRef]
- Gao, D.; Li, Q.; Liu, Z.; Li, Y.; Liu, Z.; Fan, Y.; Li, K.; Han, Z.; Li, J. Hypoglycemic effects and mechanisms of action of Cortex Lycii Radicis on alloxan-induced diabetic mice. Yakugaku Zasshi 2007, 127, 1715–1721. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Lee, L.; Kim, J.H.; Lee, T.H.; Shim, I. Antidepressant-like effects of lycii radicis cortex and betaine in the forced swimming test in rats. Biomol. Ther. 2013, 21, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Kim, J.; Yeo, S.; Lim, E.; Choi, C.W.; Choi, S.; Li, W.Y.; Lee, J.W.; Park, J.H.; Huh, D.; et al. Anti-Osteoporotic Effects of Combined Extract of Lycii Radicis Cortex and Achyranthes japonica in Osteoblast and Osteoclast Cells and Ovariecto-mized Mice. Nutrients 2019, 11, 2716. [Google Scholar] [CrossRef]
- Morrris, C.A.; Mehdi, S.; Rahman, J.; Rosales, P.; Lee, S.O.; Huh, D.; Yoon, D. Herbal Extracts from Lycii Radicis Corex and Achyranthes japonica Prevent Multiple Myeloma. Blood 2021, 138 (Suppl. S1), 4918. [Google Scholar] [CrossRef]
- Lee, B.; Hong, S.; Kim, M.; Kim, E.Y.; Park, H.J.; Jung, H.S.; Kim, J.H.; Sohn, Y. Lycii radicis cortex inhibits glucocorti-coid-induced bone loss by downregulating Runx2 and BMP-2 expression. Int. J. Mol. Med. 2021, 48, 155. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Kim, J.Y.; Kim, J.H.; Kim, C.Y. Kukoamine B from Lycii Radicis Cortex protects human keratinocyte HaCaT cells through covalent modification by trans-2-nonenal. Plants 2022, 12, 163. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, H.M.; Kim, J.H.; Guo, S.; Lee, D.H.; Lim, G.M.; Kim, W.; Kim, C.Y. Salvia plebeia R.Br. and Rosmarinic Acid Attenuate Dexamethasone-Induced Muscle Atrophy in C2C12 Myotubes. Int. J. Mol. Sci. 2023, 24, 1876. [Google Scholar] [CrossRef]
- Yadav, A.; Yadav, S.S.; Singh, S.; Dabur, R. Natural products: Potential therapeutic agents to prevent skeletal muscle atrophy. Eur. J. Pharmacol. 2022, 925, 174995. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Hoskins, C. Drug development: Lessons from nature. Biomed. Rep. 2017, 6, 612–614. [Google Scholar] [CrossRef]
- Kim, C.; Hwang, J.K. Flavonoids: Nutraceutical potential for counteracting muscle atrophy. Food Sci. Biotechnol. 2020, 29, 1619–1640. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, H.M.; Kim, J.H.; Lee, J.H.; Zhang, K.; Guo, S.; Lee, D.H.; Gao, E.M.; Son, R.H.; Kim, S.M.; et al. Preventive effects of the butanol fraction of Justicia procumbens L. against dexamethasone-induced muscle atrophy in C2C12 myotubes. Heliyon 2022, 8, e11597. [Google Scholar] [CrossRef] [PubMed]
- Funayama, S.; Zhang, G.R.; Nozoe, S.; Kukoamine, B. a spermine alkaloid from Lycium chinense. Phytochemistry 1995, 38, 1529–1531. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, L.; Zhu, Y.; Hou, D.; Li, Y.; Guo, X.; Wang, Y.; Olatunji, O.J.; Wan, P.; Gong, K. Kukoamine B Ameliorate Insulin Resistance, Oxidative Stress, Inflammation and Other Metabolic Abnormalities in High-Fat/High-Fructose-Fed Rats. Diabetes Metab. Syndr. Obes. 2020, 13, 1843–1853. [Google Scholar] [CrossRef]
- Li, X.; Lin, J.; Chen, B.; Xie, H.; Chen, D. Antioxidant and Cytoprotective Effects of Kukoamines A and B: Comparison and Positional Isomeric Effect. Molecules 2018, 23, 973. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Lee, H.; Nirmala, F.S.; Seo, H.D.; Ha, T.Y.; Jung, C.H.; Ahn, J. Antioxidant Activity of Valeriana fauriei Protects against Dexamethasone-Induced Muscle Atrophy. Oxid. Med. Cell. Longev. 2022, 2022, 3645431. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Yang, J.S.; Lu, C.C.; Chiu, Y.J.; Chen, H.C.; Chung, M.I.; Wu, Y.T.; Chen, F.A. Effect of Quercetin on Dexamethasone-Induced C2C12 Skeletal Muscle Cell Injury. Molecules 2020, 25, 3267. [Google Scholar] [CrossRef]
- Shen, S.; Liao, Q.; Liu, J.; Pan, R.; Lee, S.M.; Lin, L. Myricanol rescues dexamethasone-induced muscle dysfunction via a sirtuin1-dependent mechanism. J. Cachexia Sarcopenia Muscle 2019, 10, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Zhiyin, L.; Jinliang, C.; Qiunan, C.; Yunfei, Y.; Qian, X. Fucoxanthin rescues dexamethasone induced C2C12 myotubes atrophy. Biomed. Pharmacother. 2021, 139, 111590. [Google Scholar] [CrossRef] [PubMed]
- Yuenyongchaiwat, K.; Kulchanarat, C.; Satdhabudha, O. Sarcopenia in open heart surgery patients: A cohort study. Heliyon 2020, 6, e05759. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.; Kim, W.; Davie, J. Myogenin is required for assembly of the transcription machinery on muscle genes during skeletal muscle differentiation. PLoS ONE 2021, 16, e0245618. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.; Leinwand, L.A.; Reggiani, C. Developmental myosins: Expression patterns and functional significance. Skelet. Muscle 2015, 5, 22. [Google Scholar] [CrossRef]
- Rodriguez, J.; Vernus, B.; Chelh, I.; Cassar-Malek, I.; Gabillard, J.C.; Hadj Sassi, A.; Seiliez, I.; Picard, B.; Bonnieu, A. Myostatin and the skeletal muscle atrophy and hypertrophy signaling pathways. Cell Mol. Life Sci. 2014, 71, 4361–4371. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Chunthorng-Orn, J.; Lord, S.; Musa, I.; Dawson, P.; Holm, L.; Lai, Y.C. Ubiquitin E3 ligase Atrogin-1 protein is regulated via the rapamycin-sensitive mTOR-S6K1 signaling pathway in C2C12 muscle cells. Am. J. Physiol. Cell Physiol. 2022, 323, C215–C225. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Vuong, T.A.; Go, G.Y.; Song, Y.J.; Lee, S.; Lee, S.Y.; Kim, S.W.; Lee, J.; Kim, Y.K.; Seo, D.W.; et al. An isoflavone compound daidzein elicits myoblast differentiation and myotube growth. J. Funct. Foods 2017, 38, 438–446. [Google Scholar] [CrossRef]
- Kim, R.; Kim, H.; Im, M.; Park, S.K.; Han, H.J.; An, S.; Kang, J.S.; Lee, S.J.; Bae, G.U. BST204 Protects Dexamethasone-Induced Myotube Atrophy through the Upregulation of Myotube Formation and Mitochondrial Function. Int. J. Environ. Res. Public Health 2021, 18, 2367. [Google Scholar] [CrossRef]
- Yuan, Y.; Li, D.; Yu, F.; Kang, X.; Xu, H.; Zhang, P. Effects of Akt/mTOR/p70S6K Signaling Pathway Regulation on Neuron Remodeling Caused by Translocation Repair. Front. Neurosci. 2020, 14, 565870. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Choi, J.W.; Choi, Y.H.; Nam, T.J. Protective Effect of Pyropia yezoensis Peptide on Dexamethasone-Induced Myotube Atrophy in C2C12 Myotubes. Mar. Drugs 2019, 17, 284. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Kang, S.Y.; Kim, S.J.; Park, Y.K.; Jung, H.W. Monotropein Improves Dexamethasone-Induced Muscle Atrophy via the Akt/mTOR/FOXO3a Signaling Pathways. Nutrients 2022, 14, 1859. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, G.; Lin, H.; Lin, M.; Wang, H.; Lin, Y. Chitosan postharvest treatment suppresses the pulp breakdown devel-opment of longan fruit through regulating ROS metabolism. Int. J. Biol. Macromol. 2020, 165 Pt A, 601–608. [Google Scholar] [CrossRef]
- Lian, D.; Chen, M.M.; Wu, H.; Deng, S.; Hu, X. The Role of Oxidative Stress in Skeletal Muscle Myogenesis and Muscle Disease. Antioxidants 2022, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Fang, Q.; Xu, T.; Wu, C.; Xu, L.; Wang, L.; Yang, X.; Yu, S.; Zhang, Q.; Ding, F.; et al. Mechanistic Role of Reactive Oxy-gen Species and Therapeutic Potential of Antioxidants in Denervation- or Fasting-Induced Skeletal Muscle Atrophy. Front. Physiol. 2018, 9, 215. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Smuder, A.J.; Judge, A.R. Oxidative stress and disuse muscle atrophy: Cause or consequence? Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Leeuwenburgh, C.; Gurley, C.M.; Strotman, B.A.; Dupont-Versteegden, E.E. Age-related differences in apoptosis with disuse atrophy in soleus muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1288–R1296. [Google Scholar] [CrossRef]
- Morris, J.L.; Gillet, G.; Prudent, J.; Popgeorgiev, N. Bcl-2 Family of Proteins in the Control of Mitochondrial Calcium Signalling: An Old Chap with New Roles. Int. J. Mol. Sci. 2021, 22, 3730. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, J.G.; Chen, S.T.; Tafani, M.; Snyder, J.W.; Farber, J.L. The overexpression of Bax produces cell death upon induction of the mitochondrial permeability transition. J. Biol. Chem. 1998, 273, 7770–7775. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, R.; Liu, J.; Xu, X.; Sun, G.; Zhao, D.; Sun, L. 20(s) ginseonside Rg3 modulation of AMPK/FoxO3 signaling to attenuate mitochondrial dysfunction in a dexame-thasone injured C2C12 myotube based model of skeletal atrophy in vitro. Mol. Med. Rep. 2021, 23, 306. [Google Scholar] [CrossRef]
- Golpich, M.; Amini, E.; Mohamed, Z.; Azman Ali, R.; Mohamed Ibrahim, N.; Ahmadiani, A. Mitochondrial dysfunction and biogenesis in neurodegenerative diseases: Pathogenesis and treatment. CNS Neurosci. Ther. 2017, 23, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Carafoli, E.; Margreth, A.; Buffa, P. Early biochemical changes in mitochondria from denervated muscle and their relation to the onset of atrophy. Exp. Mol. Pathol. 1964, 3, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Romanello, V.; Sandri, M. The connection between the dynamic remodeling of the mitochondrial network and the regulation of muscle mass. Cell. Mol. Life Sci. 2021, 78, 1305–1328. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Lopez, M.E.; Weindruch, R.; Aiken, J.M. Association of age-related mitochondrial abnormalities with skeletal muscle fiber atrophy. Free Radic. Biol. Med. 1998, 25, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Tryon, L.D.; Vainshtein, A.; Memme, J.M.; Crilly, M.J.; Hood, D.A. Recent advances in mitochondrial turnover during chronic muscle disuse. Integr. Med. Res. 2014, 3, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, K.; Ren, Q.; Yi, L.; Zhu, J.; Zhang, Q.; Mi, M. Dihydromyricetin Attenuates Dexamethasone-Induced Muscle Atrophy by Improving Mitochondrial Function via the PGC-1 Pathway. Cell. Physiol. Biochem. 2018, 49, 758–779. [Google Scholar] [CrossRef]
- Menshikova, E.V.; Ritov, V.B.; Fairfull, L.; Ferrell, R.E.; Kelley, D.E.; Goodpaster, B.H. Effects of exercise on mitochondrial content and function in aging human skeletal muscle. J. Gerontol. A Biol. Sci. Med. Sci. 2006, 61, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Roshanravan, B.; Liu, S.Z.; Ali, A.S.; Shankland, E.G.; Goss, C.; Amory, J.K.; Robertson, H.T.; Marcinek, D.J.; Conley, K.E. In vivo mitochondrial ATP production is improved in older adult skeletal muscle after a single dose of elamipretide in a randomized trial. PLoS ONE 2021, 16, e0253849. [Google Scholar] [CrossRef] [PubMed]
- Seene, T.; Kaasik, P.; Pehme, A.; Alev, K.; Riso, E.M. The effect of glucocorticoids on the myosin heavy chain isoforms’ turn-over in skeletal muscle. J. Steroid Biochem. Mol. Biol. 2003, 86, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Hata, J.; Nakashima, D.; Tsuji, O.; Fujiyoshi, K.; Yasutake, K.; Sera, Y.; Komaki, Y.; Hikishima, K.; Nagura, T.; Matsumoto, M.; et al. Noninvasive technique to evaluate the muscle fiber characteristics using q-space imaging. PLoS ONE 2019, 14, e0214805. [Google Scholar] [CrossRef] [PubMed]
- McCurdy, R.D.; McGrath, J.J.; Mackay-Sim, A. Validation of the comparative quantification method of real-time PCR analysis and a cautionary tale of housekeeping gene selection. Gene Ther. Mol. Biol. 2008, 12, 15–24. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, R.H.; Kim, M.I.; Kim, H.M.; Guo, S.; Lee, D.H.; Lim, G.M.; Kim, S.-M.; Kim, J.-Y.; Kim, C.Y. Potential of Lycii Radicis Cortex as an Ameliorative Agent for Skeletal Muscle Atrophy. Pharmaceuticals 2024, 17, 462. https://doi.org/10.3390/ph17040462
Son RH, Kim MI, Kim HM, Guo S, Lee DH, Lim GM, Kim S-M, Kim J-Y, Kim CY. Potential of Lycii Radicis Cortex as an Ameliorative Agent for Skeletal Muscle Atrophy. Pharmaceuticals. 2024; 17(4):462. https://doi.org/10.3390/ph17040462
Chicago/Turabian StyleSon, Rak Ho, Myeong Il Kim, Hye Mi Kim, Shuo Guo, Do Hyun Lee, Gyu Min Lim, Seong-Min Kim, Jae-Yong Kim, and Chul Young Kim. 2024. "Potential of Lycii Radicis Cortex as an Ameliorative Agent for Skeletal Muscle Atrophy" Pharmaceuticals 17, no. 4: 462. https://doi.org/10.3390/ph17040462