Abstract
The emergence and spread of drug-resistant pathogens, resulting in antimicrobial resistance, continue to compromise our capability to handle commonly occurring infectious diseases. The rapid global spread of multi-drug-resistant pathogens, particularly systemic fungal infections, presents a significant concern, as existing antimicrobial drugs are becoming ineffective against them. In recent decades, there has been a notable increase in systemic fungal infections, primarily caused by Candida species, which are progressively developing resistance to azoles. Moreover, Candida species biofilms are among the most common in clinical settings. In particular, they adhere to biomedical devices, growing as a resilient biofilm capable of withstanding extraordinarily high antifungal concentrations. In recent years, many research programs have concentrated on the development of novel compounds with possible antimicrobial effects to address this issue, and new sources, such as plant-derived antimicrobial compounds, have been thoroughly investigated. Essential oils (EOs), among their numerous pharmacological properties, exhibit antifungal, antibacterial, and antiviral activities and have been examined at a global scale as the possible origin of novel antimicrobial compounds. A recent work carried out by our research group concerned the synergistic antibacterial activities of commercially available and chemically characterized Cinnamomum verum L. essential oil (C. verum EO) in association with sertraline, a selective serotonin reuptake inhibitor whose repositioning as a non-antibiotic drug has been explored over the years with encouraging results. The aim of this work was to explore the synergistic effects of C. verum EO with sertraline on both planktonic and sessile Candida species cells. Susceptibility testing and testing of the synergism of sertraline and C. verum EO against planktonic and sessile cells were performed using a broth microdilution assay and checkerboard methods. A synergistic effect was evident in both the planktonic cells and mature biofilms, with significant reductions in fungal viability. Indeed, the fractional inhibitory concentration index (FICI) was lower than 0.5 for all the associations, thus indicating significant synergism of the associations with the Candida strains examined. Moreover, the concentrations of sertraline able to inhibit Candida spp. strain growth and biofilm formation significantly decreased when it was used in combination with C. verum EO for all the strains considered, with a reduction percentage in the amount of each associated component ranging from 87.5% to 97%.
1. Introduction
Candida species are opportunistic pathogenic microorganisms that can cause systemic infections with a high mortality rate in immunocompromised and at-risk patients, thus representing a serious threat to public health [1]. Indeed, Candida spp. are the primary cause of 50–70% of systemic fungal infections, and candidemia is the most common hospital infection, representing approximately 15% of bloodstream infections [2]. Candida albicans is the most commonly isolated pathogenic species. However, other species, such as Candida glabrata, Candida tropicalis, Candida parapsilosis, Candida krusei, Candida famata, Candida guilliermondii, and Candida lusitaniae, have increasingly been isolated, principally in human immunodeficiency virus (HIV)-infected individuals [3,4]. According to some studies, Candida strains are able to adhere to artificial and mucosal surfaces to produce biofilms, incredibly complex structures [5]. Biofilm formation is a significant virulence factor that lengthens hospital stays, raises death rates, and increases the cost of antifungal therapy in patients affected by this condition [6]. In addition, the growing resistance of yeast to antifungal drugs is one of the main issues among researchers and clinicians. The biological mechanisms that decrease the sensitivity of yeast biofilms to antifungal compounds, including the most commonly used azoles, involve active drug efflux, the limited penetration of molecules through the extracellular matrix, the low metabolic activity of cells in mature biofilms, and the higher expression of various genes in biofilms than in planktonic forms [7,8]. In spite of the harmful effects fungi pose to human health, there are only a few classes of antifungal drugs currently accessible for addressing these potentially life-threatening infections; furthermore, those currently available do not completely provide safe and secure protection [9]. To overcome this issue, numerous pharmaceutical compounds from different therapeutic classes are being evaluated as antimicrobials in the drug repositioning approach to finding new, cost-effective solutions [10]. Non-antifungal medications exhibiting antifungal properties are proven to be effective in combating fungal infections. Certain antidepressant medications have demonstrated antimicrobial effects, with the most potent activity observed in the third-generation antidepressants referred to as selective serotonin reuptake inhibitors (SSRIs), such as fluoxetine, paroxetine, and sertraline [11]. Notably, sertraline, owing to its antimicrobial potential, has been shown to augment the efficacy of various antibiotics, reverse pathogen multidrug resistance, and render them susceptible to previously ineffective drugs [12,13]. Antifungal activity of sertraline has been recently demonstrated against planktonic forms and biofilms of some fungal strains [14,15]. In addition, a recent study demonstrated the synergistic activity of SSRIs, including sertraline, in combination with azoles against Cryptococcus spp. [16]. Another strategy for addressing therapeutic failures in the treatment of fungal infections is the use of plant-derived compounds [17,18,19]. Amongst phytochemicals, essential oils (EOs), yielded by aromatic plants as secondary metabolites, and their bioactive pure compounds have demonstrated an extensive array of noteworthy biological activities, including antibacterials and antifungals [20,21]. EOs can mitigate microbial growth and inhibit biofilm formation through distinct mechanisms associated with the breakdown of the bacterial cell wall, their influence on the breakdown of enzymes or membrane proteins, or the release of cell contents following cytoplasmic membrane failure [22,23]. The intricate composition of EOs can play a fundamental role in counteracting microbial resistance since it can be challenging for pathogens to develop resistance to several substances [24]. This suggests that EOs and antibiotics may work synergistically to combat microbial infections. This aspect matters since it may contribute to lowering the dosage and usage of antibiotics in treatments, thereby decreasing their adverse effects [25]. Among the essential oils with pronounced antimicrobial activities emerge those derived from species within the genus Cinnamomum (Lauraceae), including Cinnamomum verum, commonly referred to as Ceylon cinnamon, or the cinnamon tree [26]. Despite significant variations in the chemical composition observed across oils from different regions worldwide [27,28], trans-cinnamaldehyde predominates as the major constituent, constituting 47–71% of the composition, while their other constituents include eugenol and linalool. The essential oil of C. verum has been used for centuries in various cultural practices due to its medicinal properties. These include its use as an antiseptic, an anti-inflammatory [29], and a preservative agent [30]. Diverse studies have reported the potent antibacterial and antifungal properties of C. verum EO, both independently and in conjunction with antibiotics [31,32]. Indeed, a recent study reported the efficacy of low concentrations of C. verum EO in the inactivation of E. faecalis and S. aureus in carrot juice and white mulberry juice [33]. Furthermore, the efficacy of C. verum leaf essential oil has been demonstrated as a therapeutic alternative for Candida biofilm infections, highlighting its potential in addressing fungal resistance issues [28]. This work is part of our ongoing research program directed toward the development of new strategies to overcome antimicrobial resistance. Indeed, in recent years, we have concentrated our efforts on examining the possible synergies between EOs and antibiotics (gentamicin, oxacillin, and norfloxacin) or repurposed drugs (diclofenac, sertraline, etc.), proving the effectiveness of these combinations and raising the possibility of novel therapeutic applications [34,35,36]. In our previous work, we demonstrated the synergistic antibacterial effect on planktonic cells of sertraline in combination with C. verum EO [37]. Considering the compelling findings acquired and acknowledging the documented capability of sertraline to combat fungal infections and hamper biofilm growth [38,39], the goal of this work was to evaluate the synergic action of the combination of sertraline and C. verum EO on planktonic cells of different Candida strains and their ability to inhibit the growth of biofilms produced by fungal strains.
2. Results
2.1. Cinnamomum verum EO’s Chemical Composition
Gas chromatography–mass spectrometry analyses of the EO were performed following the procedure described in our previous papers [35,37]. The analysis resulted in the identification of 99% of the whole mixture. The principal constituents of C. verum EO are (E)-cinnamaldehyde (72%), linalool (6.78%), eugenol (6%), cinnamyl acetate (5%), and eucalyptol (1.47%). The other constituents are present at quantities of less than 1%, i.e., p-cymene, humulene, a-pinene, etc. Our results, shown in Table 1, are basically in agreement with what has been previously reported [37].

Table 1.
Chemical composition of Cinnamomum verum EO.
2.2. Antifungal Activity of Sertraline in Association with Cinnamomum verum EO against Planktonic Cells of Candida spp.
The synergistic antifungal effect of C. verum EO in combination with sertraline against planktonic cells of several strains of Candida spp. has been explored by means of the microdilution checkerboard technique. Table 2 displays the minimal inhibitory concentration (MIC) values for sertraline and C. verum EO. MICo represents the MIC value of a single component tested alone, while MICc is the MIC value of each component in the association at the most effective inhibition growth. The fractional inhibitory concentration (FIC) is calculated by the MICc/MICo ratio, and the fractional inhibitory concentration index (FICI), a parameter that expresses the synergism of the two compounds, is determined by adding the FIC of sertraline to the FIC of C. verum EO. In the final column of Table 1, R% describes the percentage decrease in the quantity of each combined component compared to each single component. All the FICI values were significantly lower than 0.5, in the range of 0.08–0.16 and thus denoting a marked synergism between C. verum EO and sertraline against all the fungal strains tested. It is worth noting that the MICc value for sertraline is markedly reduced when combined with the EO since it was found to be up to 40-fold lower than the MICo for most of the yeast species under study. Interestingly, the best results were achieved against C. albicans ATCC 90028 and C. kefyr ATCC 204093, for which the MIC values of sertraline decreased from 32.0 (MICo) to 0.80 (MICc) µg/mL and from 64.0 (MICo) to 3.20 (MICc) µg/mL, respectively, by showing a noteworthy reduction percentage of 97% in the first case and 95% in the second, as proven by the FICI values of 0.08 for both species. Concerning the American Type Culture Collection (ATCC) strains, the synergistic activity of the combination against C. albicans ATCC 10231 and C. glabrata ATCC 15126 should be noted. Indeed, a substantial reduction in the sertraline concentration is observed since it switches from 64.0 (MICo) to 3.20 (MICc) µg/mL for both strains, exhibiting a 20-fold reduction. An impressive synergistic effect was also observed on C. albicans A18 and C. krusei 31A29, both derived from clinical isolation, for which the sertraline concentrations decreased from 128.00 to 6.40 and from 64.00 to 3.20 µg/mL, respectively. Interestingly, the combination under study was also shown to be effective against C. albicans 10 A12 (FICI = 0.17), C. krusei ATCC 6258 (FICI = 0.16), and C. tropicalis ATCC 750 (FICI = 0.16). Overall, the clear-cut synergic activity of sertraline and C. verum EO determined a significant reduction percentage for the amount of each component in association with respect to the single component that ranged between 87.5% and 97%.
Table 2.
Antifungal effect of sertraline (µg/mL), Cinnamomum verum EO (mg/mL), and their association on different Candida spp. strains.
2.3. Inhibitory Effects on Candida spp. Biofilm Growth of Sertraline in Association with Cinnamomum verum EO
The encouraging results obtained on the planktonic cells prompted us to investigate the potential inhibitory activity of the association between sertraline and C. verum EO against the growth of biofilms produced by the yeast strains considered in this research. The effects of this association, obtained by means of an XTT assay, expressed as percentage values of the reduction in biofilm growth, along with the amounts of each component in association at the most effective growth inhibition, are reported in Table 3. In addition, all of the sessile minimal inhibitory concentration values (sMIC50) for sertraline and C. verum EO were reported for comparison. The combination of sertraline with C. verum EO was effective in hampering biofilm growth of all the Candida spp. tested, with the inhibition percentages ranging from 59.90 to 86.44%, thus confirming the trend of the results obtained on the planktonic cells. Indeed, the FICI values for this association were between 0.16 and 0.35, demonstrating an evident synergistic effect even on the sessile cells. Particularly interesting are the data obtained against C. albicans ATCC 90028 since the association of sertraline with C. verum EO strongly succeeded in inhibiting the biofilm growth produced by this strain, with a percentage value of 86.44% (FICI = 0.23). The combination also proved to be quite promising in preventing the growth of the biofilms formed by the strains from clinical isolation of C. albicans A 18 (FICI = 0.35) and C. albicans 10 A12 (FICI = 0.23), with percentages of 77.02 and 75.29%, respectively, and those formed by C. albicans ATCC 10231 (FICI = 0.16), with a percentage of 72.8%. As already observed for the planktonic cells, an interesting result was also achieved with sessile cells of C. kefyr ATCC 204093. Indeed, the biofilm formed by this yeast decreased with a percentage amounting to 79.80% (FICI = 0.20). The C. glabrata ATCC 15126, C. krusei ATCC 6558, C. krusei 31 A29, and C. tropicalis ATCC 750 biofilms treated with the sertraline–C. verum EO association also appreciably decreased, with percentages amounting to 59.90, 64.90, 69.44, and 68.25%, respectively. Another parameter that allows us to confirm the effectiveness of the association between these two substances is the significant reduction in the amounts of sertraline able to inhibit biofilm growth when combined with C. verum EO with respect to the amounts of sertraline tested alone. Indeed, the sMIC50 of sertraline ranged between 640.00 and 40.00 µg/mL, whereas the concentration of sertraline in association, for the most effective combination, ranged between 32.00 and 4.00 µg/mL, thus decreasing 10 to 20 times. This decrement was particularly pronounced for C. glabrata ATCC 15126 and C. kefyr ATCC 204093, whereby the sMIC50 of sertraline dropped from 320.00 to 16.00 µg/mL for both, and for C. tropicalis ATCC 750, for which the concentration required to hamper biofilm growth declined from 640.00 to 32.00 µg/mL.
Table 3.
Inhibitory effects (% Red) of Cinnamomum verum EO, sertraline, and their combinations on fungal biofilms.
2.4. Molecular Modeling Studies
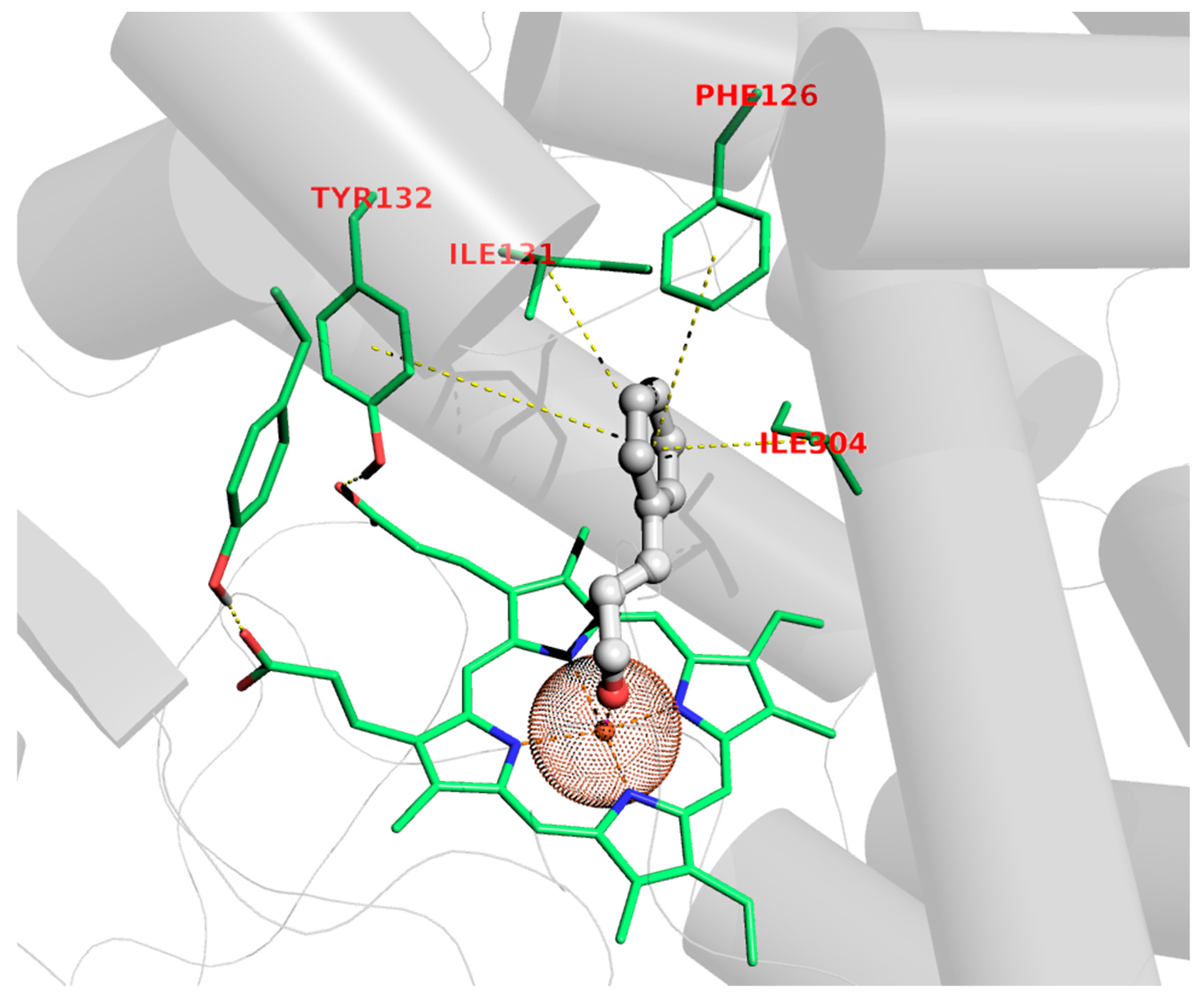
Although the antifungal properties of C. verum EO have not yet been related to a specific mechanism of action or related to exact targets, the C. albicans cytochrome P450-dependent lanosterol 14α-demethylase (CYP51) was enrolled to study the potential mechanisms of C. verum’s main component through molecular modeling experiments. Lanosterol 14α-demethylase (CYP51) is a member of the cytochrome P450 superfamily which catalyzes the oxidative removal of the 14α-methyl group of lanosterol to give ∆14,15-desaturated intermediates in ergosterol biosynthesis [44]. The selection of CYP51 as a plausible site of action is also supported by the evidence that azoles (i.e., fluconazole and miconazole) and other drugs (i.e., benzothiazines and benzoxazine) [45,46,47,48,49,50,51,52,53,54,55,56,57] inhibit cell growth upon ergosterol depletion as determined by CYP51 blockade. On the basis of this evidence, within the EO constituents, the principal chemical entity (E)-cinnamaldehyde was selected as chaperon for this action; hence, binding to CYP51 was assessed through dockings. The pose acquired highlighted some interesting features that might be in charge of the antifungal activity, mainly referring to the possibility of accommodating the whole (E)-cinnamaldehyde molecular scaffold in the middle of the lodge located over the heme group of the target, anchoring nearly perpendicularly by means of a coordination bond involving iron and the aldehydic carbonyl oxygen atom, as depicted in Figure 1.
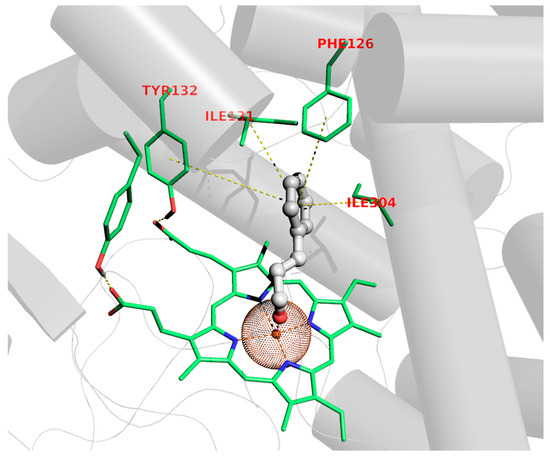
Figure 1.
Binding pose of (E)-cinnamaldehyde to CYP51.
This feature has been experimentally observed in the X-ray structure of other thiazoles in complex with different but functionally related cytochromes [49], as has the chelating ability of cinnamaldehyde also been proven by X-ray data diffraction [50,51]. In addition to this, π-π stackings of the Phe126 and Tyr118 aromatic rings and hydrophobic contact with the Ile121 and Ile304 sidechains handle the binding to the cavity dome of CYP51, as corroborated by favorable docking scores (see Section 4.2).
3. Discussion
The antifungal resistance of Candida spp. to azoles and echinocandins, the main therapeutic options for the treatment of candidiasis, continues to be an issue of global concern [58], along with biofilm formation [59]. Microbial biofilms are intricate, spatially organized communities of microorganisms ensconced within an extracellular matrix and adhered to a surface. Candida species are known to form intricate and dynamic biofilms consisting of diverse morphological manifestations of the fungus, including yeasts, pseudohyphae, and hyphae. According to the National Institutes of Health (NIH), more than 80% of human microbial infections are linked to biofilms [60]. Candida spp. biofilms play a significant role in recurrent and chronic infections, demonstrating resilience and/or resistance to diverse antifungal compounds and the innate immune system [61]. Due to the limitations in treating Candida spp. biofilms with conventional drugs, alternative options are urgently required. In this scenario, the drug repositioning approach, which aims to identify new therapeutic potential among already approved drugs, is part of an encouraging development in the field of antimicrobial drug discovery. The literature has reported that selective serotonin reuptake inhibitors (SSRIs) impact antimicrobial activity [11,38,62,63]. Sertraline (an SSRI), in addition to its primary purpose, exhibits antimicrobial activity against various yeast strains, either independently or when combined with antifungal agents [64,65]. Furthermore, effects against Candida spp. biofilms have been reported [39,66]. The precise mechanism by which sertraline acts is still not uniquely understood; however, evidence supports the hypothesis that sertraline, a serotonin reuptake pump inhibitor in humans, can also behave as an efflux pump inhibitor in bacteria [63]. In addition, some research corroborates the ability of sertraline to cause cell wall and membrane damage in Candida spp., also affecting ergosterol synthesis [14,67]. Another promising strategy for finding new treatments for acute fungal infections is the use of EOs in combination with traditional antifungal agents or repositioned drugs, known for their antifungal potential. This strategy aims to improve the treatment efficacy of conventional drugs and decrease toxicity, side effects, and resistance problems. Indeed, several EOs have been recognized as a significant reservoir of compounds suitable for application in fungal infections, especially in the treatment of biofilms [68,69,70]. The efficacy of these associations was substantiated by our previous studies related to the synergy of EOs with certain commercially available antimicrobials and diclofenac, a widely recognized non-steroidal anti-inflammatory drug, which displayed notable antimicrobial and antifungal efficacy, thereby supporting the potential therapeutic application of these combinations [35,36,71,72,73]. In addition, scientific studies reported synergistic activity of C. verum EO when used in combination with various antibiotics. Indeed, it was demonstrated that the synergistic combination of C. verum EO with piperacillin effectively tackled a beta-lactamase-resistant strain of Escherichia coli. One noteworthy finding revealed a remarkable two-fold reduction in the MIC of piperacillin, decreasing from 1024 µg/mL to 256 µg/mL, when used in association with the EO [74]. The capability of C. verum EO to cause irreversible membrane damage and to reduce the bacterial surface charge has been hypothesized as a plausible mechanism of action. One study also suggested the capability of C. verum EO to function as a quorum-sensing inhibitor, thus reversing E. coli’s resistance to piperacillin [75]. C. verum EO has been also demonstrated to enhance the antimicrobial activity of ampicillin or chloramphenicol against multi-resistant strains such as Bacillus subtilis and Escherichia coli [32]. More recently, we demonstrated the synergistic antibacterial effect of the non-antibiotic drug sertraline in combination with C. verum EO on several bacterial strains [36]. Besides its antibacterial potential, some studies have highlighted the synergistic activity of C. verum EO when combined with antifungal agents in addressing Candida infections. Indeed, it has been demonstrated that combined treatment of C. verum EO and fluconazole (a FIC value of 0.37) led to 69.51% killing of C. albicans cells after 3 h of incubation and allowed us to decrease the effective concentrations four- to eight-fold compared to the MIC values. Furthermore, this combination was found to inhibit ergosterol synthesis at sub-MIC values [76]. The same potent synergistic effect was demonstrated for the C. verum EO/amphotericin B association (FIC value of 0.37) against Candida spp. biofilm. Notably, this combination inhibited C. albicans biofilm formation with a percentage of 94%. This association has also been reported to affect the production of secreted aspartic proteases, a virulent factor for C. albicans ATCC 10231, while transcriptomic analyses demonstrated the combined effect of C. verum EO and amphotericin B in down-regulating Ras-cAMP-Efg and MAPK signaling genes and inhibiting the expression of adhesion and secreted aspartyl proteinase virulence factor genes [77]. Building upon these encouraging findings and what was achieved in our previous work, we aimed to extend our research to the evaluation of the antifungal effects of the association between C. verum EO and sertraline on different Candida spp. and their biofilms. As underlined in our in vitro assays, the fungal strains under study displayed sensitivity to the compounds tested, both individually and in combination, as shown in Table 1, thus confirming a clear-cut synergism between sertraline and C. verum EO. Particularly interesting was the significant reduction in the sertraline concentration needed for planktonic cells when it was used in combination with C. verum EO for all the yeast strains under study. A noteworthy decrease in sertraline’s active concentration was observed for C. albicans ATCC 90028, for which the MIC value of sertraline was found to decrease from 32.0 to 0.80 µg/mL. Meanwhile, for C. albicans ATCC 10231, C. albicans 10 A12, C. glabrata ATCC 15126, C. kefyr ATCC 204093, and C. krusei 31A29, the active concentration dropped from 64.00 to 3.20 µg/mL. These results are particularly interesting considering that these strains are responsible for most nosocomial infections of the bloodstream. The FICI values were significantly lower than the limit value of 0.5 for both the planktonic and sessile cells, thus confirming the substantial synergism between sertraline and C. verum EO. Indeed, the sertraline–C. verum EO combination considerably prevented the biofilm growth of all Candida spp. tested, with the inhibition percentages ranging from 59.90 to 86.44%. Also, in this case, it was possible to observe a significant decrease in the sertraline concentration necessary to inhibit biofilm growth when it was used in the combination. This outcome was particularly noteworthy in the case of C. albicans ATCC 90028. Indeed, while the use of sertraline alone necessitated 40.00 µg/mL, when combined with C. verum EO, a mere 4.00 µg/mL was adequate to achieve a substantial reduction in biofilm growth, by 86.4%. This clear synergistic effect was corroborated by a corresponding FICI value of 0.23, markedly lower than the threshold value of 0.5. The data also indicated a considerable synergistic effect on those strains from clinical isolation. Indeed, the concentration of sertraline combined with EO that was able to inhibit 69.44 ± 0.80% of C. krusei 31 A29’s biofilms amounted to 8.00 µg/mL, whereas sertraline alone necessitates 160.00 µg/mL. These data are particularly encouraging since the remarkable antimicrobial efficacy of this combination particularly stands out when compared to the associations between C. verum EO and the conventional antibiotics or antifungals reported in the literature, as documented by their lower FIC values. Moreover, the lower FIC values imply a more effective synergistic effect, whereby the combined action of this association achieves greater inhibition of microbial growth at lower concentrations than either compound alone. This is underscored by the considerable decrease in the concentration of the respective components within the association.
The effects achieved could be attributed to active substances in C. verum EO such as (E)-cinnamaldehyde, which is the major component, and others such as eugenol, caryophyllene, and cinnamyl acetate that are among the minor constituents, as reported in our previous work [37]. Our docking studies revealed that (E)-cinnamaldehyde binds effectively to the active site of CYP51, suggesting a potential mechanism by which this compound exerts its antifungal activity. This interaction is particularly significant as CYP51 is a key enzyme involved in the biosynthesis of ergosterol, an essential component of the fungal cell membrane. The inhibition of CYP51 can disrupt ergosterol production, leading to increased cell membrane permeability and ultimately cell death. This finding supports the hypothesis that (E)-cinnamaldehyde, as the principal constituent of C. verum essential oil, plays a major role in the observed antifungal effects. However, the mixture of these components present in C. verum EO conceivably contribute to the antifungal activity of the EO and work alongside with sertraline for the observed synergistic effect [78,79]. Indeed, it could be hypothesized that synergy arises from concerted action on multiple cellular targets. As previously reported, C. verum EO could act non-specifically against the cell envelope of microorganisms. Indeed, it has been reported that some of the constituents of C. verum EO, such as (E)-cinnamaldehyde and eugenol, could be responsible for damaging cell membrane integrity [80,81]. In addition, (E)-cinnamaldehyde could promote ROS generation in microbial cells, thus triggering cell death [82]. In this context, sertraline could act affecting by a specific target, such as ergosterol synthesis, or by hampering efflux pumps, which may, in turn, enhance the whole inhibitory effect. Otherwise, membrane permeabilization by C. verum EO’s components could enhance sertraline uptake [14,65,83].
4. Materials and Methods
4.1. Materials
4.1.1. Essential Oil
The C. verum EO (Lot 140/0000324, 10.2018, 10 mL) was supplied by Erbe Nobili srl (Corato, Bari, Italy) and was stored in a vial shielded from light and at temperatures ranging from 0 to 4 °C until its use. The EO samples had already undergone gas chromatographic analysis (GC-MS) in our prior study [37].
4.1.2. Chemicals
Sertraline was obtained from Sigma-Aldrich srl (Milan, Italy). The filters used were provided by Agilent Technologies Italia spa (Milan, Italy). The culture media employed were Sabouraud 2% dextrose broth (Oxoid, Rodano, Italy) and Yeast Malt Broth (Oxoid, Rodano, Italy).
4.1.3. Fungal Strains
The antifungal activity was assessed against numerous fungal strains, encompassing various strains sourced from the American Type Culture Collection (ATCC, Rockville, MD, USA), which were C. albicans (ATCC 10231, ATCC 90028), C. glabrata (ATCC 15126), C. tropicalis (ATCC 750), C. kefyr (ATCC 204093), and C. krusei (ATCC 6258), and Candida strains derived from clinical isolation (C. albicans 10 A12, C. albicans A18, and C. krusei 31 A29). All the clinical isolates were obtained from patients admitted to the intensive care unit of the Department of Biomedical Sciences and Human Oncology, the University of Bari “Aldo Moro”, Italy. The isolation procedures were carried out in the hygiene section of the Department utilizing conventional physiological and morphological methods alongside an API system.
4.2. Methods
4.2.1. Antifungal Testing
The strains were preserved at −80 °C in yeast peptone dextrose broth with 10–25% glycerol (Oxoid, Rodano, Italy) solution. Glycerol stocks were prepared for all the strains, stored at −20 °C, and subcultured on antimicrobial agent-free Sabouraud dextrose agar plates (BioMerieux, Marcy L’Etoile, France) to ensure their viability and purity before the start of the study. The antifungal activity of sertraline was assessed by means of the microdilution method, as described by the Clinical and Laboratory Standards Institute (CLSI, M27-A3) [84].
4.2.2. Medium and Culture Conditions
Each frozen stock culture was inoculated in Sabouraud dextrose broth and incubated at 37 °C for 24 h in an orbital shaker at 60 rpm. Following incubation, the cells were transferred into a tube containing RPMI 1640 broth medium with L-glutamine and without bicarbonate buffered to a pH of 7 with MOPS, 3-(N-morpholino) propane sulfonic acid (165 M, Sigma, Milan, Italy). A standardized suspension of 1 × 106 CFU/mL was obtained and immediately used.
4.2.3. Preparation of the Test Solution
C. verum EO was dissolved in ethanol at a ratio of 1:5 and then further diluted in RPMI 1640 broth medium supplemented with Tween 80. Sertraline was appropriately dissolved in DMSO and subsequently in the culture medium (DMSO < 5%).
4.2.4. The Checkerboard Test
The checkerboard method was employed to assess the synergistic, additive, or antagonistic effects of the combination of sertraline and C. verum EO. The tested dilutions were based on the MIC of the two substances. The combination of the two compounds was synergistic when the FICI was ≤0.5, additive when the FICI was >0.5 and <1, and antagonistic when the FICI was >1. The test was conducted using sterile 96-well microtiter plates containing sertraline and EO at two-fold serial concentrations. The MICs were obtained following incubation at 37 °C for 48 h. Each test was carried out in triplicate.
4.2.5. Biofilm Biomass Measurement and Reduction
To assess the synergistic antibiofilm action of C. verum EO in association with sertraline, we conducted, in vitro, a colorimetric XTT (2,3-bis(2-methoxy-4-nitro-5-sulphophenyl)-5-[(phenylamino)carbonyl]-2H-tetrazolium hydroxide) assay. Briefly, 200 µL of yeast culture (106 CFU/mL) was added to each well of a 96-well flat microtiter plate and incubated for 24 h at 37 °C by shaking it on a rocker table to facilitate cell attachment and biofilm formation. Subsequently, 200 µL of antibiotic was added as a positive control, while the negative control contained only RPMI 1640 without the sertraline–C. verum EO association. After incubation, the contents of each well were aspirated, and the wells were washed with 100 µL of sterile PBS to remove loosely adherent and nonviable cells. Following incubation, 200 µL of each sertraline–C. verum EO combination was added to the wells. Four double serial dilutions of the 40% ethanol C. verum EO with Tween 80 0.1% were prepared using the same method as described in our previous works for MIC evaluation. Dilutions of the C. verum EO were prepared alongside a series of double dilutions of the drug and the EO. This method ensured that all the sertraline dilutions were combined with appropriate concentrations of EO, resulting in a range of sertraline–EO concentration combinations [85]. These concentrations were prepared to represent 40%, 20%, 10%, and 5% of the MIC values for the EO and 25%, 12.5%, 6.25%, and 3.12% of the MIC values for sertraline [86,87]. Following incubation with the drug, the media was aspirated from each well, and the supports were washed with 1 mL of PBS. The biofilms were then plunged in 2 mL of PBS with the addition of 180 μL of XTT solution (1 mg/mL) and Menadione (0.4 mM) at a ratio of 6:1. This solution was prepared by dissolving XTT in sterile water (obtained by filtration) and Menadione in dimethyl sulfoxide. The plate with the supports was incubated for 2 h at 37 °C. After 2 h, the plate was read with a microplate spectrophotometer Tecan Infinite M1000 Pro multiplate reader (Tecan, Cernusco S.N., Italy) at a 490 nm wavelength, where there was a maximum Formazan absorbance peak. The sessile minimal inhibitory concentration (sMIC50) was determined as the lowest antibiofilm concentration for both compounds causing a visible reduction in the growth of the biofilm compared with the growth in the control well without the presence of sertraline or the EO. The percentage of the reduction in the biofilm obtained with the combinations studied was calculated using the following formula: percentage of biofilm reduction = (OD control well−OD experimental)/(OD control well) × 100 [31]. Antimicrobial susceptibility tests of the biofilms were performed in triplicate on different days. In our experimental protocols, the combinations of the substances were analyzed by calculating the FIC index (FICI). Generally, the FICI value was interpreted as (i) a synergistic effect when it was ≤0.5; (ii) an additive or indifferent effect when it was >0.5 or <1; and (iii) an antagonistic effect when it was >1 [88]. The results are expressed as means ± SDs of at least three distinct measurements performed in triplicate.
4.2.6. Molecular Modeling Studies
The (E)-cinnamaldehyde SMILES string was converted into a three-dimensional structure and its energy relaxed with Open Babel [51] after 10,000 steepest descent iterations. The target protein (pdb code 5V5Z [52]) was prepared for docking with the Maestro Protein Preparation Wizard [53]. The binding poses were sampled throughout 1000 runs of the Lamarckian Genetic Algorithm (LGA) implemented in AutoDock 4.2.6 [54] using the GPU-OpenCL algorithm version [55], exploring a 85 × 85 × 85 Å3 box with its center on the iron atom and a spacing of 0.375 Å. The Gasteiger charges were calculated for the ligand and the heme group with QUACPAC [56] and the AMBER UNITED force field [57] for the peptide chain of CYP51. The best free energy of binding (−5.30 kcal/mol) and the most populated (1000/1000) pose were selected.
5. Conclusions
Recent research has shown promising outcomes in antimicrobial therapies that combine repositioned drugs with essential oils. Our previous study demonstrated the synergistic efficacy of C. verum EO and sertraline against various bacterial strains. Given the public health threat posed by Candida infections and the role of microbial biofilms in fostering resistance, our study aimed to assess the synergistic effects of sertraline and C. verum EO on Candida spp. in both planktonic and sessile states. Our results highlight the potent effect of this combination, significantly reducing the quantity of sertraline required to suppress fungal strains and decrease biofilm production. These results hold paramount importance since demonstrating the enhanced efficacy of the association at lower concentrations compared to the components taken individually, this novel combination offers a path towards more sustainable and effective treatment options. While our study was preliminary and conducted in vitro, further investigations are needed to clarify the mechanism underlying this synergism and to assess its clinical efficacy, particularly against biofilms on medical devices. These findings offer a promising foundation for exploring alternative antifungal therapies targeting biofilm infections and potentially overcoming the challenges associated with conventional antibiotics.
Author Contributions
Conceptualization, A.C. (Alessia Carocci), A.R. and G.F.; methodology, A.B., A.C. (Alessia Carocci) and A.R.; investigation, A.B., R.T. and G.F.; data curation, A.R., G.F. and A.C. (Alessia Carocci); formal analysis, L.F.; software, A.C. (Antonio Carrieri); writing—original draft, A.B.; writing—review and editing, A.C. (Alessia Carocci); validation, supervision, L.F.; project administration, A.C. (Alessia Carocci); funding acquisition, A.C. (Alessia Carocci) and F.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained within the manuscript.
Conflicts of Interest
The authors reported no potential conflicts of interest.
References
- Guinea, J. Global Trends in the Distribution of Candida Species Causing Candidemia. Clin. Microbiol. Infect. 2014, 20, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.C.D.O.; Vasconcelos, C.C.; Lopes, A.J.O.; Cartágenes, M.D.S.D.S.; Filho, A.K.D.B.; Nascimento, F.R.F.D.; Ramos, R.; Pires, E.R.R.B.; Andrade, M.; Rocha, F.M.G.; et al. Candida Infections and Therapeutic Strategies: Mechanisms of Action for Traditional and Alternative Agents. Front. Microbiol. 2018, 9, 1351. [Google Scholar]
- Barchiesi, F.; Orsetti, E.; Osimani, P.; Catassi, C.; Santelli, F.; Manso, E. Factors related to outcome of bloodstream infections due to Candida parapsilosis complex. BMC Infect. Dis. 2016, 16, 387. [Google Scholar] [CrossRef]
- Pathakumari, B.; Liang, G.; Liu, W. Immune defence to invasive fungal infections: A comprehensive review. Biomed. Pharmacother. 2020, 130, 110550. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, E.G.; Andes, D.R. Candida Biofilm Tolerance: Comparison of Planktonic and Biofilm Resistance Mechanisms. In Candida albicans: Cellular and Molecular Biology; Springer International Publishing: Cham, Switzerland, 2017; pp. 77–92. [Google Scholar]
- De Barros, P.P.; Rossoni, R.D.; De Souza, C.M.; Scorzoni, L.; Fenley, J.C.; Junqueira, J.C. Candida Biofilms: An Update on Developmental Mechanisms and Therapeutic Challenges. Mycopathologia, 2020; published online ahead of print. [Google Scholar]
- Roemer, T.; Krysan, D.J. Antifungal drug development: Challenges, unmet clinical needs, and new approaches. Cold Spring Harb. Perspect. 2014, 4, a019703. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, C.H.; Morelli, K.A.; Schultz, D.; Nadell, C.D.; Cramer, R.A. Fungal biofilm architecture produces hypoxic microenvironments that drive antifungal resistance. Proc. Natl. Acad. Sci. USA 2020, 117, 22473–22483. [Google Scholar] [CrossRef] [PubMed]
- Trubenová, B.; Roizman, D.; Moter, A.; Rolff, J.; Regoes, R.R. Population genetics, biofilm recalcitrance, and antibiotic resistance evolution. Trends Microbiol. 2022, 30, 841–852. [Google Scholar] [CrossRef]
- Moraes, D.C.; Ferreira-Pereira, A. Insights on the anti Candidal activity of non-antifungal drugs. J. Mycol. Med. 2019, 29, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Caldara, M.; Marmiroli, N. Antimicrobial Properties of Antidepressants and Antipsychotics—Possibilities and Implications. Pharmaceuticals 2021, 14, 915. [Google Scholar] [CrossRef]
- Bottega, A.; Serafin, M.B.; Da Rosa, T.F.; Foletto, V.S.; Machado, C.D.S.; Coelho, S.S.; Mainardi, A.; Hörner, R. Antimicrobial and Antineoplastic Properties of Sertraline. Am. J. Ther. 2020, 27, e632–e635. [Google Scholar] [CrossRef]
- Muthu, D.; Gowri, M.; Kumar, G.S.; Kattimani, V.S.; Girija, E.K. Repurposing of antidepression drug sertraline for antimicrobial activity against Staphylococcus aureus: A potential approach for the treatment of osteomyelitis. New J. Chem. 2019, 43, 5315–5324. [Google Scholar] [CrossRef]
- Gowri, M.; Jayashree, B.; Jeyakanthan, J.; Girija, E. Sertraline as a promising antifungal agent: Inhibition of growth and biofilm of Candida auris with special focus on the mechanism of action in vitro. J. Appl. Microbiol. 2019, 128, 426–437. [Google Scholar] [CrossRef]
- Treviño-Rangel, R.D.J.; Villanueva-Lozano, H.; Méndez-Galomo, K.S.; Solís-Villegas, E.M.; Becerril-García, M.A.; Montoya, A.M.; Robledo-Leal, E.R.; González, G.M. In vivo evaluation of the antifungal activity of sertraline against Aspergillus fumigatus. J. Antimicrob. Chemother. 2019, 74, 663–666. [Google Scholar] [CrossRef] [PubMed]
- da Silva, C.R.; Sá, L.G.D.A.V.; Ferreira, T.L.; Leitão, A.C.; de Farias Cabral, V.P.; Rodrigues, D.S.; Barbosa, A.D.; Moreira, L.E.A.; Filho, H.L.P.; de Andrade Neto, J.B.; et al. Antifungal activity of selective serotonin reuptake inhibitors against Cryptococcus spp. and their possible mechanism of action. J. Mycol. Med. 2023, 33, 101431. [Google Scholar] [CrossRef]
- Lagrouh, F.; Dakka, N.; Bakri, Y. The antifungal activity of Moroccan plants and the mechanism of action of secondary metabolites from plants. J. Med. Mycol. 2017, 27, 303–311. [Google Scholar] [CrossRef]
- Arif, T.; Bhosale, J.D.; Kumar, N.; Mandal, T.K.; Bendre, R.S.; Lavekar, G.S.; Dabur, R. Natural products—Antifungal agents derived from plants. J. Asian Nat. Prod. Res. 2009, 11, 621–638. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V. Essential oils and antifungal activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef]
- Angane, M.; Swift, S.; Huang, K.; Butts, C.A.; Quek, S.Y. Essential oils and their major components: An updated review on antimicrobial activities, mechanism of action and their potential application in the food industry. Foods 2022, 11, 464. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Rikke, L.M. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial Activity of Some Essential Oils—Present Status and Future Perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; El-Saadony, M.T.; Alagawany, M.; Arif, M.; Batiha, G.E.; Khafaga, A.F.; Elwan, H.A.M.; Elnesr, S.S.; El-Hack, M.E.A. Using essential oils to overcome bacterial biofilm formation and their antimicrobial resistance. Saudi J. Biol. Sci. 2021, 28, 5145–5156. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Ullah, F.; Ovais, M.; Ahmed, J.; Devkota, H.P. Synergistic interactions of phytochemicals with antimicrobial agents: Potential strategy to counteract drug resistance. Chem. Biol. Interact. 2019, 308, 294–303. [Google Scholar] [CrossRef]
- Singh, N.; Rao, A.S.; Nandal, A.; Kumar, S.; Yadav, S.S.; Ganaie, S.A.; Narasimhan, B. Phytochemical and pharmacological review of Cinnamomum verum J. Presl-a versatile spice used in food and nutrition. Food Chem. 2020, 338, 127773. [Google Scholar] [CrossRef] [PubMed]
- Al-Mijalli, S.H.; Mrabti, H.N.; El Hachlafi, N.; El Kamili, T.; Elbouzidi, A.; Abdallah, E.M.; Flouchi, R.; Assaggaf, H.; Qasem, A.; Zengin, G.; et al. Integrated analysis of antimicrobial, antioxidant, and phytochemical properties of Cinnamomum verum: A comprehensive In vitro and In silico study. Biochem. Syst. Ecol. 2023, 110, 104700. [Google Scholar] [CrossRef]
- Wijesinghe, G.K.; de Oliveira, T.R.; Maia, F.C.; de Feiria, S.B.; Barbosa, J.P.; Joia, F.; Boni, G.C.; Höfling, J.F. Efficacy of true cinnamon (Cinnamomum verum) leaf essential oil as a therapeutic alternative for Candida biofilm infections. Iran J. Basic Med. Sci. 2021, 24, 787–795. [Google Scholar] [PubMed]
- Pagliari, S.; Forcella, M.; Lonati, E.; Sacco, G.; Romaniello, F.; Rovellini, P.; Fusi, P.; Palestini, P.; Campone, L.; Labra, M.; et al. Antioxidant and Anti-Inflammatory Effect of Cinnamon (Cinnamomum verum J. Presl) Bark Extract after In Vitro Digestion Simulation. Foods 2023, 12, 452. [Google Scholar] [CrossRef]
- Valková, V.; Ďúranová, H.; Galovičová, L.; Vukovic, N.L.; Vukic, M.; Kowalczewski, P.Ł.; Kačániová, M. Application of Three Types of Cinnamon Essential Oils as Natural Antifungal Preservatives in Wheat Bread. Appl. Sci. 2022, 12, 10888. [Google Scholar] [CrossRef]
- Narayanankutty, A.; Kunnath, K.; Alfarhan, A.; Rajagopal, R.; Ramesh, V. Chemical composition of Cinnamomum verum leaf and flower essential oils and analysis of their antibacterial, insecticidal, and larvicidal properties. Molecules 2021, 26, 6303. [Google Scholar] [CrossRef]
- Padalia, H.; Rathod, T.; Moteriya, P.; Chanda, S. Antimicrobial efficacy of Cinnamonum verum essential oil alone and in combination with antibiotics and other essential oils. Int. J. Curr. Microbiol. Appl. 2017, 6, 3377–3395. [Google Scholar] [CrossRef]
- Erkmen, O. Antimicrobial effects of Cinnamomum verum essential oil against Enterococcus faecalis and Staphylococcus aureus in mulberry and carrot juices. Int. J. Food Sci. Technol. 2024, 59, 2297–2304. [Google Scholar] [CrossRef]
- Rosato, A.; Sblano, S.; Salvagno, L.; Carocci, A.; Clodoveo, M.L.; Corbo, F.; Fracchiolla, G. Anti-biofilm inhibitory synergistic effects of combinations of essential oils and antibiotics. Antibiotics 2020, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Altini, E.; Sblano, S.; Salvagno, L.; Maggi, F.; de Michele, G.; Carocci, A.; Clodoveo, M.L.; Corbo, F.; Fracchiolla, G. Synergistic Activity of New Diclofenac and Essential Oils Combinations against Different Candida spp. Antibiotics 2021, 10, 688. [Google Scholar] [CrossRef]
- Barbarossa, A.; Rosato, A.; Carrieri, A.; Tardugno, R.; Corbo, F.; Clodoveo, M.L.; Fracchiolla, G.; Carocci, A. Antifungal Biofilm Inhibitory Effects of Combinations of Diclofenac and Essential Oils. Antibiotics 2023, 12, 1673. [Google Scholar] [CrossRef] [PubMed]
- Barbarossa, A.; Sblano, S.; Rosato, A.; Carrieri, A.; Corbo, F.; Clodoveo, M.L.; Fracchiolla, G.; Carocci, A. Synergistic Action of Cinnamomum verum Essential Oil with Sertraline. Antibiotics 2022, 11, 1617. [Google Scholar] [CrossRef]
- Oliveira, A.S.; Martinez-de-Oliveira, J.; Donders, G.G.G.; Palmeira-de-Oliveira, R.; Palmeira-de-Oliveira, A. Anti-Candida Activity of Antidepressants Sertraline and Fluoxetine: Effect upon Pre-Formed Biofilms. Med. Microbiol. Immunol. 2018, 207, 195–200. [Google Scholar] [CrossRef]
- Rodrigues, D.S.; Cabral, V.P.D.F.; Barbosa, A.D.; Sá, L.G.D.A.V.; Moreira, L.E.A.; de Andrade Neto, J.B.; da Silva, C.R.; de Moraes, M.O.; Silva, J.; Marinho, E.S.; et al. Sertraline has in vitro activity against both mature and forming biofilms of different Candida species. J. Med. Microbiol. 2023, 72, 001664. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Steam, IL, USA, 1995; ISBN 978-1-932633-21-4. [Google Scholar]
- NIST Chemistry WebBook 2022. Available online: https://webbook.nist.gov/chemistry/ (accessed on 20 April 2022).
- Koo, I.; Kim, S.; Zhang, X. Comparative analysis of mass spectral matching-based compound identification in gas chromatographymass spectrometry. J. Chromatogr. A 2013, 1298, 132–138. [Google Scholar] [CrossRef]
- Wan, K.X.; Vidavsky, I.; Gross, M.L. From similarity index to spectral contrast angle. J. Am. Soc. Mass Spectrom. 2002, 13, 85–88. [Google Scholar] [CrossRef]
- Vanden Bossche, H.; Koymans, L. Cytochromes P450 in fungi. Mycoses 1998, 41 (Suppl. S1), 32–38. [Google Scholar]
- Sheehan, D.J.; Hitchcock, C.A.; Sibley, C.M. Current and emerging azole antifungal agents. Clin. Microbiol. Rev. 1999, 12, 40–79. [Google Scholar] [CrossRef] [PubMed]
- Lamb, D.C.; Kelly, D.E.; Kelly, S.L. Molecular diversity of sterol 14α-demethylase substrates in plants, fungi and humans. FEBS Lett. 1998, 425, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Lepesheva, G.I.; Waterman, M.R. Sterol 14alpha-demethylase cytochrome P450 (CYP51), a P450 in all biological kingdoms. Biochim. Biophy. Acta 2007, 1770, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Sevrioukova, I.F.; Poulos, T.L. Structure and mechanism of the complex between cytochrome P4503A4 and ritonavir. Proc. Nat. Acad. Sci. USA 2010, 107, 18422–18427. [Google Scholar] [CrossRef]
- De Cian, A.; Weiss, R. Structures de complexes π carbonyés du fer. II. Structure cristalline et moléculaire de l’aldéhyde cinnamique fertricarbonyle: C6H5CH=CH–CHO.Fe(CO)3. Acta Cryst. 1972, B28, 3273–3280. [Google Scholar] [CrossRef]
- Sacerdoti, M.; Bertolasi, V.; Gilli, G. The structure of dicarbonyl(η4-cinnamaldehyde)(triphenylphosphine)iron(0). Acta Cryst. 1980, B36, 1061–1065. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Keniya, M.V.; Sabherwal, M.; Wilson, R.K.; Woods, M.A.; Sagatova, A.A.; Tyndall, J.D.A.; Monk, B.C. Crystal Structures of Full-Length Lanosterol 14α-Demethylases of Prominent Fungal Pathogens Candida albicans and Candida glabrata Provide Tools for Antifungal Discovery. Antimicrob. Agents Chemother. 2018, 62, 1134. [Google Scholar] [CrossRef]
- Schrödinger Release 2023-4: Maestro; Schrödinger, LLC.: New York, NY, USA, 2023.
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Khoury, L.E.; Santos-Martins, D.; Sasmal, S.; Eberhardt, J.; Bianco, G.; Ambrosio, F.A.; Solis-Vasquez, L.; Koch, A.; Forli, S.; Mobley, D.L. Comparison of affinity ranking using AutoDock-GPU and MM-GBSA scores for BACE-1 inhibitors in the D3R Grand Challenge 4. J. Comput. Aided Mol. Des. 2019, 33, 1011–1020. [Google Scholar] [CrossRef]
- QUACPAC 2.1.0.4, OpenEye Scientific Software, Santa Fe, NM. Available online: http://www.eyesopen.com (accessed on 5 August 2024).
- Cornell, W.D.; Cieplak, P.; Bayly, C.I.; Gould, I.R.; Merz, K.M.; Ferguson, D.M.; Spellmeyer, D.C.; Fox, T.; Caldwell, J.W.; Kollman, P.A. A second generation force field for the simulation of proteins, nucleic acids, and organic molecules. J. Am. Chem. Soc. 1995, 117, 5179–5197. [Google Scholar] [CrossRef]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Mathe, L.; Van Dijck, P. Recent insights into Candida albicans biofilm resistance. Curr. Genet. 2013, 59, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Foletto, V.S.; da Rosa, T.F.; Serafin, M.B.; Bottega, A.; Hörner, R. Repositioning of non-antibiotic drugs as an alternative to microbial resistance: A systematic review. Int. J. Antimicrob. Agents 2021, 58, 106380. [Google Scholar] [CrossRef] [PubMed]
- Caldara, M.; Marmiroli, N. Tricyclic antidepressants inhibit Candida albicans growth and biofilm formation. Int. J. Antimicrob. Agents 2018, 52, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Alanís-Ríos, S.A.; González, G.M.; Andrade, A.; Becerril-García, M.A.; Bonifaz, A.; Robledo-Leal, E.R.; Montoya, A.M.; Treviño-Rangel, R.D.J. Evaluation of the synergistic antifungal activity of micafungin and voriconazole plus sertraline against Candida auris. Braz. J. Microbiol. 2022, 53, 2003–2008. [Google Scholar] [CrossRef]
- Alkhalifa, B.A.; Bulatova, N.R.; abuRokba, W.; Darwish, R.M. Serotonin reuptake inhibitors effect on fluconazole activity against resistant Candida glabrata strains. J. Glob. Antimicrob. Resist. 2022, 29, 49–54. [Google Scholar] [CrossRef]
- Ahmed, E.I.; Alhuwaydi, A.M.; Taha, A.E.; Abouelkheir, M. Anti-Candidal Activity of Reboxetine and Sertraline Antidepressants: Effects on Pre-Formed Biofilms. Antibiotics 2023, 12, 881. [Google Scholar] [CrossRef]
- Rodrigues, D.S.; Cabral, V.P.; Barbosa, A.D.; Valente Sá, L.G.; Silva, C.R.; Moreira, L.E.; Neto, J.B.A.; Silva, J.; Santos, H.S.; Marinho, E.S.; et al. Sertraline has fungicidal activity against Candida spp. and acts by inhibiting membrane and cell wall biosynthesis. Future Microbiol. 2023, 18, 1025–1039. [Google Scholar] [CrossRef] [PubMed]
- Sahal, G.; Woerdenbag, H.J.; Hinrichs, W.L.; Visser, A.; Tepper, P.G.; Quax, W.J.; van der Mei, H.C.; Bilkay, I.S. Antifungal and biofilm inhibitory effect of Cymbopogon citratus (lemongrass) essential oil on biofilm forming by Candida tropicalis isolates; an in vitro study. J. Ethnopharmacol. 2020, 246, 112188. [Google Scholar] [CrossRef]
- Garcia, L.G.S.; da Rocha, M.G.; Lima, L.R.; Cunha, A.P.; de Oliveira, J.S.; de Andrade, A.R.C.; Ricardo, N.M.P.S.; Pereira-Neto, W.A.; Sidrim, J.J.C.; Rocha, M.F.G.; et al. Essential oils encapsulated in chitosan microparticles against Candida albicans biofilms. Int. J. Biol. Macromol. 2021, 166, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Barbarossa, A.; Mustafa, A.M.; Bonacucina, G.; Perinelli, D.R.; Petrelli, R.; Maggi, F.; Spinozzi, E. Comprehensive Evaluation of the Antibacterial and Antifungal Activities of Carlina acaulis L. Essential Oil and Its Nanoemulsion. Antibiotics 2021, 10, 1451. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Carocci, A.; Catalano, A.; Clodoveo, M.L.; Franchini, C.; Corbo, F.; Carbonara, G.G.; Carrieri, A.; Fracchiolla, G. Elucidation of the synergistic action of Mentha Piperita essential oil with common antimicrobials. PLoS ONE 2018, 13, e0200902. [Google Scholar] [CrossRef] [PubMed]
- Salvagno, L.; Sblano, S.; Fracchiolla, G.; Corbo, F.; Clodoveo, M.L.; Rosato, A. Antibiotics—Mentha piperita essential oil synergism inhibits mature bacterial biofilm. Chem. Today 2020, 38, 49–52. [Google Scholar]
- Rosato, A.; Vitali, C.; De Laurentis, N.; Armenise, D.; Milillo, M.A. Antibacterial effect of some essential oils administered alone or in combination with Norfloxacin. Phytomedicine 2007, 14, 727–732. [Google Scholar] [CrossRef]
- Yap, P.S.; Lim, S.H.; Hu, C.P.; Yiap, B.C. Combination of essential oils and antibiotics reduce antibiotic resistance in plasmid-conferred multidrug resistant bacteria. Phytomedicine 2013, 20, 710–713. [Google Scholar] [CrossRef]
- Yap, P.S.; Krishnan, T.; Chan, K.G.; Lim, S.H. Antibacterial Mode of Action of Cinnamomum verum Bark Essential Oil, Alone and in Combination with Piperacillin, Against a Multi-Drug-Resistant Escherichia coli Strain. J. Microbiol. Biotechnol. 2015, 25, 1299–1306. [Google Scholar] [CrossRef]
- Essid, R.; Hammami, M.; Gharbi, D.; Karkouch, I.; Hamouda, T.B.; Elkahoui, S.; Limam, F.; Tabbene, O. Antifungal Mechanism of the Combination of Cinnamomum Verum and Pelargonium Graveolens Essential Oils with Fluconazole against Pathogenic Candida Strains. Appl. Microbiol. Biotechnol. 2017, 101, 6993–7006. [Google Scholar] [CrossRef]
- Essid, R.; Gharbi, D.; Abid, G.; Karkouch, I.; Ben Hamouda, T.; Fares, N.; Trabelsi, D.; Mhadhbi, H.; Elkahoui, S.; Limam, F.; et al. Combined effect of Thymus capitatus and Cinnamomum verum essential oils with conventional drugs against Candida albicans biofilm formation and elucidation of the molecular mechanism of action. Ind. Crop. Prod. 2019, 140, 111720. [Google Scholar] [CrossRef]
- Shariati, A.; Didehdar, M.; Razavi, S.; Heidary, M.; Soroush, F.; Chegini, Z. Natural Compounds: A Hopeful Promise as an Antibiofilm Agent against Candida Species. Front. Pharmacol. 2022, 13, 917787. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing. Available online: https://clsi.org/media/3481/m100ed30_sample.pdf (accessed on 15 May 2023).
- OuYang, Q.; Duan, X.; Li, L.; Tao, N. Cinnamaldehyde exerts its antifungal activity by disrupting the cell wall integrity of Geotrichum citri-aurantii. Front. Microbiol. 2019, 10, 55. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Pereira, F.; Mendes, J.M.; de Oliveira Lima, E. Investigation on mechanism of antifungal activity of eugenol against Trichophyton rubrum. Med. Mycol. 2013, 51, 507–513. [Google Scholar] [CrossRef]
- Lee, J.E.; Jung, M.; Lee, S.C.; Huh, M.J.; Seo, S.M.; Park, I.K. Antibacterial mode of action of trans-cinnamaldehyde derived from cinnamon bark (Cinnamomum verum) essential oil against Agrobacterium tumefaciens. Pestic. Biochem. Phys. 2020, 165, 104546. [Google Scholar] [CrossRef]
- Giske, C.G.; Turnidge, J.; Cantón, R.; Kahlmeter, G. Update from the European committee on antimicrobial susceptibility testing (EUCAST). J. Clin. Microbiol. 2022, 60, e00276-21. [Google Scholar] [CrossRef]
- Eloff, J.N. Quantifying the bioactivity of plant extracts during screening and bioassay-guided fractionation. Phytomedicine 2004, 11, 370–371. [Google Scholar] [CrossRef] [PubMed]
- Williamson, E.M. Synergy and other interactions in phytomedicines. Phytomedicine 2001, 8, 401–409. [Google Scholar] [CrossRef]
- White, D.C.; Arrage, A.A.; Nivens, D.E.; Palmer, R.J.; Rice, J.F.; Sayler, G.S. Biofilm ecology: On-line methods bring new insights into mic and microbial biofouling. Biofouling 1996, 10, 3–16. [Google Scholar] [CrossRef]
- Stepanovíc, S.; Vukovíc, D.; Dakíc, I.; Savíc, B.; Švabíc-Vlahovíc, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Saracino, I.M.; Foschi, C.; Pavoni, M.; Spigarelli, R.; Valerii, M.C.; Spisni, E. Antifungal Activity of Natural Compounds vs. Candida spp.: A Mixture of Cinnamaldehyde and Eugenol Shows Promising In Vitro Results. Antibiotics 2022, 11, 73. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).