
Application of Mammalian Nudix Enzymes to Capped RNA Analysis
Department of Biophysics, Faculty of Physics, University of Warsaw, Pasteura 5, 02-093 Warsaw, Poland
Pharmaceuticals 2024, 17(9), 1195; https://doi.org/10.3390/ph17091195
Submission received: 30 June 2024
/
Revised: 3 September 2024
/
Accepted: 9 September 2024
/
Published: 11 September 2024
(This article belongs to the Special Issue Design, Synthesis, and Evaluation of Nucleobase, Nucleoside, and Nucleotide Analogues)
Abstract
:Following the success of mRNA vaccines against COVID-19, mRNA-based therapeutics have now become a great interest and potential. The development of this approach has been preceded by studies of modifications found on mRNA ribonucleotides that influence the stability, translation and immunogenicity of this molecule. The 5′ cap of eukaryotic mRNA plays a critical role in these cellular functions and is thus the focus of intensive chemical modifications to affect the biological properties of in vitro-prepared mRNA. Enzymatic removal of the 5′ cap affects the stability of mRNA in vivo. The NUDIX hydrolase Dcp2 was identified as the first eukaryotic decapping enzyme and is routinely used to analyse the synthetic cap at the 5′ end of RNA. Here we highlight three additional NUDIX enzymes with known decapping activity, namely Nudt2, Nudt12 and Nudt16. These enzymes possess a different and some overlapping activity towards numerous 5′ RNA cap structures, including non-canonical and chemically modified ones. Therefore, they appear as potent tools for comprehensive in vitro characterisation of capped RNA transcripts, with special focus on synthetic RNAs with therapeutic activity.
Keywords:
mRNA cap; synthetic cap analogues; NUDIX family; Nudt16; Nudt12; Nudt5; Nudt2; mRNA therapeutics1. Introduction
mRNA-based technology has recently come into focus due to the development and worldwide use of new-generation vaccines against SARS-CoV-2 [1,2]. In the pioneering work, it was shown that in-vitro-synthesized mRNA supports protein synthesis after direct injection into skeletal muscle [3]. Thus, in addition to mRNA-based vaccines against infectious diseases, mRNA appears as a promising platform for other medically relevant applications, such as cancer immunotherapies or protein-replacement therapies [4]. This is highlighted by the 2023 Nobel Prize awarded to Katalin Kariko and Drew Weissman for their research on nucleoside base modifications of mRNA [5] and the long-standing research of other pioneers at the early stages of mRNA characterization [6].
In addition to the diverse modifications present on the eukaryotic mRNA chain [7], the 5′ end of mRNA end bears a specific cap structure, comprising of 7-methylguanosine (m7Guo) connected via 5′-5′ triphosphate bond to the first transcribed nucleotide of mRNA chain (m7GpppN, or Cap 0) [8] (Figure 1). Due to the biological roles of the 5′ cap in mRNA cellular functions (translation, stability, immunogenicity), it is a “hot-spot” for designing and synthesising chemically modified cap-structure analogues (e.g., [9,10]). For instance, the modified cap m27,3′OGpppAmpG is already part of modified mRNAs in designed and distributed vaccine [1].
These chemically modified and synthesized cap analogues, once incorporated into mRNA, require characterisation in terms of capping efficiency, proper cap orientation, support of efficient cap-dependent translation, induction of cellular immunogenic response, and stability to enzymatic decapping. In this review, we focus on selected mammalian decapping enzymes of the NUDIX protein family, namely Dcp2, Nudt16, Nudt12, Nudt2 and Nudt5. These enzymes have been shown to act on distinct RNA cap structures in vivo and/or in vitro. Dcp2 and Nudt16, due to their initial discovery in the 5′ cap processing of mRNA and snRNA/snoRNA [11,12,13,14,15], have mostly been used to date for the detailed characterization and validation of mRNA transcripts with chemically modified 5′ caps. Including a wider set of Nudt decapping proteins with different or overlapping specificities towards chemically modified cap structures could be beneficial for a comprehensive understanding of the susceptibility to enzymatic degradation of in vitro designed RNAs. Furthermore, the use of human enzymes provides a more physiologically relevant environment for the analysis of modified caps, which is pivotal for the design, optimisation, and translation of therapeutic (m)RNAs into the clinic.
2. Diversity of 5′ RNA Cap Structures—Natural Caps
Beyond the aforementioned Cap 0 structure, the 5′ mRNA cap can be considered an extended structure where the first transcribed RNA nucleotides (namely N1 and N2 in the m7GpppN1pN2 structure) are 2′O-methylated, resulting in the m7Gpppm2′ON1 (Cap 1) or m7Gpppm2′ON12′ON2 (Cap 2) structures [16]. Cap 0 is generally found in lower eukaryotes, whereas Cap 1 and Cap 2 are hallmarks of higher eukaryotes, including humans [8]. The most extensively methylated cap, Cap 4, has been identified in Kinetoplastida (e.g., Typanosoma brucei, Trypanosoma cruzi and Leishmania major), where the first four transcribed nucleotides adjacent to m7Guo (m7GpppN1pN2N3pN4) are heavily methylated (m7Gpppm36,6,2′OApm2′OApm2′OC pm23,2′OU) [17]. Additionally, adenosine as the first transcribed nucleotide of Cap 1 can be also methylated at the N6 position of the purine ring (m6A) both in the absence or presence of the 2′O-methylation. This modification has been shown to have a positive effect on translation efficiency in vitro [18] and on mRNA stability due to increased resistance to Dcp2 [19].
Eukaryotic RNA (e.g., snRNA, snoRNA, primary miRNA, pre-tRNA) and specific mRNAs (selenoprotein encoding transcripts, or junD, dhx9 and tgs1 mRNA) are modified at their 5′ ends with other cap structures, such as: trimethylguanosine cap (m32,2,7GpppN), 5′-gamma-methyl (tri)phosphate cap (CH3pppN), and alfa-dimethyl monophosphate cap [20,21,22,23,24]. Additional 5′ cap structures have been detected on short RNAs (~20–200 nt), including non-methylated caps (GpppG and GpppA/ApppG), and next to the m7Gp or m32,2,7Gp moiety, the dimethylguanosine (m22,7Gp and m22,2Gp), monomethylguanosine (m1Gp, m2Gp), and monomethylcitidine (mCp) structures. Moreover, the 2′O-methylation of the first transcribed nucleotide (N1) has been detected together with the aforementioned structures and trimethylated m32,2,7Gp also with dimethyladenosine m6Am [25,26], Table 1.
Although initially identified on bacterial RNAs [27], the diversity of naturally occurring 5′ caps present on mRNA and other types of eukaryotic cellular RNAs is further expanded by the detection of structures where the cap is composed of a non-canonical initiating nucleotide (NCIN): nicotinamide adenine dinucleotide NAD+ (NAD-cap) [28,29], flavin adenine dinucleotide cap (FAD-cap), dephospho-CoA (dpCoA-cap) or UDP-glucose derived caps (uridine diphosphate–glucose UDP-Glc cap and UDP-N-acetyloglucosoamie UDP-GlcNAc), so called “metabolic” cap structures (Figure 2) [30,31,32]. A very recent report showed that adenosine dinucleotide tetraphosphate (AppppA) can also serve as yet another type of non-canonical RNA cap (Ap4A-RNA) in eukaryotic cells (HEK293T, RBL-2H3 cell lines) [33].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Diversity of naturally occurring cap structures on Eukaryotic cellular RNAs.
| Cap Structure | RNA Type | Cell Type | Ref. |
|---|---|---|---|
| m7GpppN1 m7Gpppm2′ON1 m7Gpppm6,2′OA m7Gpppm2′ON12′ON2 | mRNA | Eukaryotic | [8,19,20] |
| m32,2,7GpppN | snRNA (U1, U2, U4, U5), snoRNA primary miRNA (pri-miRNA) selenoprotein encoded transcripts, junD, dhx9 and tgs1 mRNA | Eukaryotic HPFs (human foreskin fibroblasts) HEK293, OSCA-40 canine osteosarcoma, HEK293FT | [20] [34] [22,23,24] |
| CH3pppN 5′-gamma-methyl (tri)phosphate cap | U6 and 7SK snRNAs | Eukaryotic | [35,36] |
| (CH3)2pN Alfa-dimethyl-phosphate cap | Precursor RNA of miR-145 (pre-miR-145) | MCF-7 and MDA-MB-231 breast cancer cells | [21] |
| m7GpppA(G) m22,7GpppA(G) m32,2,7GpppA(G) | pre-tRNA | S. cerevisiae HeLa | [25] |
| m7GpppA(G) mGpppC GpppG, GpppA m32,2,7Gp, m22,7Gp, m22,2Gp m1Gp, m2Gp, mCp | Fraction of short RNAs (~20–200 nt), obtained from total RNA preparation | THP-1 (acute monocyte leukemia cell line) | [26] |
| NAD+ (5′ nicotinamide-adenine dinucleotide cap) | mRNA Intronic snoRNA mRNA | S. cerevisiae HEK293T Human (CCRF-SB cell line), mouse (C57BL/6 tissues), S.cervisiae | [28] [29] [37] |
| FAD | mRNA | Human (CCRF-SB cell line), mouse (C57BL/6 tissues), S.cervisiae | [37] |
| dpCoA | Mouse liver RNA | Mouse liver RNA | [32] |
| AppppA | RNA (>200 nt) | HEK293T RBL-2H3 (rat basophilic leukemia cells) | [33] |
| UDP-GlcNAc UDP-Glc | mRNA | Human (CCRF-SB cell line), mouse (C57BL/6 tissues), S.cervisiae | [37] |
3. Diversity of Synthetic Cap Structures
Synthetic cap analogues, and synthetic capped (m)RNAs have proven to be invaluable tools for studying various cellular processes where the cap plays a pivotal role, including mRNA translation, splicing, intracellular transport, and RNA decay [38]. They are also important in studying cap-binding proteins and enzymes, including potential inhibitors of translation initiation factor eIF4E overexpressed in certain human malignancies and cancers (e.g., breast, colon, prostate, neck cancers) [39,40,41]. Now, following pioneering work demonstrating the first successful example of protein expression in muscle tissue after injection of in-vitro-transcribed capped mRNA [3] and the recent launch of anti-COVID-19 vaccines [1,2], capped RNAs have emerged as potent therapeutic agents with almost unlimited potential.
In vitro transcription (IVT) methodology is a convenient approach for producing mRNA transcripts from a provided DNA template, allowing for co-transcriptional capping with synthetic dinucleotide cap analogues, resulting in capped-mRNA within a single reaction [42,43,44,45]. One of the problems with the direct using IVT-capped transcripts in therapeutic applications is the heterogeneity of the 5′ ends of the transcription reaction products, resulting in both m7GpppG-capped and reversely oriented Gppp(m7G)-capped transcripts due to priming of the polymerisation reaction by bacteriophage RNA polymerases (T7, SP6) from both ends of the m7GpppG cap dinucleotide [46]. The breakthrough in this issue was the development of anti-reverse cap analogues (ARCAs) by the Darzykiewicz lab, where the 3′-OH group of 7-methyl-guanosine (m7G) was either replaced by methyl group (m27,3′OGpppG) (Figure 3) or removed (in 7-methyl-(3′deoxy)GpppG analogue) [47]. Such modifications prevent RNA polymerase from initiating from m7G moiety, resulting in the synthesis of transcripts capped exclusively in correct orientation, and thus in biologically active form. Similar ARCA-type properties have also been shown for dinucleotide cap analogues with 2′-OH methylated 7-methyl-guanosine [48].
A natural choice for further chemical modifications was the triphosphate bridge of the dinucleotide cap structure (Figure 4), as it may protect the resulting analogues against decapping enzyme (e.g., Dcp2, [11,12,13,14]). Additionally, elongation of the triphosphate bridge to tetra- or pentaphosphate increases binding affinity towards eIF4E and has a positive effect on cap-dependent translation in vitro [48,49,50].
Numerous different substituents have been used in synthetic cap analogs where at least one oxygen atom in the triphosphate bride was replaced, either the non-bridging α, β or γ oxygen (e.g., O-to-S, O-to-Se, O-to-BH3) [51,52,53,54,55,56], or bridging α-β and β-γ oxygen (e.g., O to NH, O to CH2, and O to CCl2, to CH2 or to CF2 in tetraphosphate analogs) [56,57,58,59]. In addition to cap analogues with a single replacement within the tri- or tetra-phosphate bridge, a series of multiple-modified compounds were synthesized, e.g., with double thiophosphate or double boranophosphate moieties, with or without O to CH2 exchange [60,61]. Another site of O-to-S substitution was the 5′-phosphoester bond in the triphosphate chain, resulting in a cap dinucleotide analogues with a phosphorothiolate moiety [62]. By applying a “click-chemistry” approach, a series of cap analogues containing the triazole moiety within the oligophosphate chain (with accompanying oxygen substitutions mentioned above in the text) were obtained, expanding the library of diverse substituents for the triphosphate bride modification [63,64,65] (Figure 5).
Further modifications that utilize chemical modifications of either the 2′-OH or/and 3′-OH on m7G, similar to ARCA cap analogues, have also been reported. These include substitution of 2′-OH with a fluorine or allyl moiety [66,67], 2′-propagyl substituted di- and tri-nucleotide cap analogues [68,69], isopropylidene analogues [70,71], anthraniolyl and methylanthraniolyl fluorescent cap analogues [72], and locked nucleic acid (LNA) di- and tri-nucleotide cap analogues [73,74]. Analogues with 2′-amino groups instead of 2′-OH of m7G were next subject for biotin attachment, facilitating the capture of mRNA–protein complexes [75,76]. Further development of this series of cap analogues resulted in compounds carrying amino-functionalized linkers at 2′-OH or 3′-OH of m7G, allowing subsequent attachment of fluorescent moieties (e.g., fluorescein), biotin [77], or photoreactive tags such as trifluoromethyl diazirine, and 6-nitroveratryl alcohol, as very recently reported by Warminski et al. [78]. Similarly, azido-functionalized analogues with Cy3 or Cy5 florescent dyes, were reported [79]. The 2′-propagyl substituted cap analogue was also the subject of fluorescent labelling (with Alexa Fluor) [69] (Figure 6).
Modifications of the cap structure at the N7, N2 or C8-position of the nucleobase of m7G have also been widely described. Initial focus was on the substitution of the N7 methyl moiety, as it is one of the structural features of the cap required for specific recognition by the eIF4E translation initiation factor, as revealed by crystallographic and biophysical data [80,81,82]. In turn, the N7-substituted compounds with diverse alkyl and aryl groups were synthesized [49,83,84,85]. Among those, N7-benzyl substitution increased inhibitory properties of mononucleotide analogues, and its pronucleotide counterpart appeared promising for targeting lung and breast cancer [39,49,86]. Furthermore, mRNA transcripts bearing N7-benzyl (or its derivatives) dinucleotide cap analogues showed higher translational activity [83,85,87]. Another favourable modification site was the N2 position of m7G, where the presence of a single methyl group has a positive effect on inhibition and translation [49,83]. Several other N2-alkyl- and -aryl-substituted analogues, including N2-benzyl and N2-(4-metoxybenzyl) ARCA, were next reported [88,89]. Further N2-modified analogues included triazole-, isoxazole-, and thiazole-ring-based substituents [10,90,91,92]. Combined modifications at N2 position of m7G and triphosphate bridge of dinucleotide and trinucleotide cap analogues (Cap 1–type) have also been described [93]. Recently, the synergistic effect that increase the inhibitory potency of synthesized compounds was shown for simultaneous modification at both N7 and N2 positions of m7G of mono- and dinucleotide analogs [41]. Photolabile groups put at the N2 (such as ortho-nitrobenzyl [94,95,96]) or at the N7 position (e.g., bezophenone or coumarin [97,98], represent yet another series of 7-methyl-guanosine modified cap analogues that can be incorporated into mRNA transcripts. Upon irradiation and photodeprotection of the cap structure, these analogues trigger the cap-dependent translation. Inagaki et al. [96] also synthesized cap analogues where the nitrobenzyl photocleavable tag was introduced at 2′-OH or 3′-OH position of m7G. Additionally, the m7G nucleobase of the cap structure can be modified at the C8-position, as demonstrated by Wojtczak et al. [99] (Figure 7).
Further development of synthetic cap analogues has led to the creation of trinucleotide and tetranucleotide compounds. Those, from one point of view, facilitated in vitro capping of mRNA transcripts in a correct orientation (such as ARCA analogues [9]; from another perspective they allowed the acquisition of analogues that reproduce the Cap-1-type and the Cap-2-type structures, that are present on mRNA transcripts in higher eukaryotes, including mammals [8,37], that are involved in suppression of the innate response activation trough IFIT1 and RIG-I [100,101,102] and confer resistance towards DXO activity [103]. Following the report of a series of m7GpppApG trinucleotide cap analogues (with differentially methylated adenosine: A, Am, m6A, m6Am) [18] and commercially available CleanCap trinucleotides [104], the trinucleotide analogues with the remaining nucleobases (in relation adenosine) were synthesized (m7GpppNpG, where N = C,Cm, U, Um, G, Gm) [105], as were also the tetranucleotide cap analogues (with Cap 2 signature) [103]. Additionally, other tri- and tetranucleotide cap analogues with modifications within the triphosphate bridge and/or m7G have been described [65,69,74,93,96,106,107].
Following the discovery of non-canonical cap structures, the synthesis of their analogs (both di- and trinucleotide series, with additional modification within the phosphate linkage) represents yet another series of compounds that facilitate the in vitro synthesis of NAD-, FAD-, or UDP-glucose-capped RNA transcripts [108,109,110].
4. NUDIX Protein Family
NUDIX enzymes are evolutionary conserved family of pyrophosphohydrolases found across all domains of life, including viruses. NUDIX (namely NUcleoside DIphosphates linked to a moiety X) possess a broad range of substrate specificity, including canonical (d)NTPs, oxidized nucleotides, dinucleotide polyphosphates, nucleotide sugars, capped RNAs, cap structures, and dinucleotide coenzymes (NAD, FAD, dpCoA) and their derivatives [111,112]. The E. coli MutT protein is referred to as a founding member of this superfamily, that is characterized by a highly conserved NUDIX motif (GX5EX5[UA]XREX2EEXGU, where U is hydrophobic and X any amino acid) [111,113]. The glutamic acid residues within the NUDIX motif (REUXEE) play role in the binding of divalent metal ions required for the catalytic activity of NUDIX enzymes [111]. The mechanism of substrate hydrolysis by NUDIX enzymes proceeds, in most cases, through nucleophilic substitution by water molecule at specific phosphorus of a di-(or polyphosphate) chain, but hydrolysis can occur also by nucleophilic substitution at carbon as it is in the case of the GDP-mannose hydrolase NUDIX subfamily [114]. Available data (based on crystal structures, mechanistic and kinetic studies) have indicated that NUDIX hydrolases usually require two or three metal ions for catalysis [114,115]. A recent study that used time-resolved X-ray crystalography of MutT, with its substrate (8-oxo-dGTP) and Mn2+, allowed for detailed visualization of the reaction process that proceed through nucleophilic substitution by water molecule with three Mn2+ ions with captured structures of intermediate reaction states [115].
Among eukaryotic NUDIX hydrolases, Dcp2 (Nudt20) was the first identified enzyme involved in removing the cap structure present at the 5′ end of mRNA [11,12,13,14]. An additional set of seven NUDIX mammalian enzymes possess mRNA cap hydrolysis activity in vitro (Nudt2, Nudt3, Nudt12, Nudt15, Nudt16, Nudt17 and Nudt19 [116,117]) and in vivo (Nudt3 and Nudt16 [15,118,119]). Aforementioned NUDIX proteins cleave within the triphosphate bridge of a cap structure, between α-β, β-γ or both α-β/β-γ phosphates (Figure 1) (e.g., [117]). Enzymatic activity of these NUDIX-decapping enzymes is influenced by the presence of specific modifications in the cap structure (e.g., by the presence of 7-methylganosie or methylation of adenosine adjacent to m7G for hDcp2 activity [14,19]; by the nucleobase type, as caps with adenosine (or 2′-O adenosine) are not subject to effective hydrolysis with hNudt16 [120,121]; or by the preference of hNutd16 for unmethylated cap [120]. As all these sites of the mRNA cap structure are subject of intensive/plethora chemical modifications in order to optimize its effect on mRNA stability, translation efficiency, and immunogenicity (Figure 4), the NUDIX proteins with different specificities towards cap structure(s) emerge as a powerful tool in their analysis.
5. RNA Decapping with Mammalian Nudt2, Nudt5, Nudt12, Nudt16, and Dcp2
In the case of the in-vitro-transcribed mRNA (IVT-mRNA), the presence of the cap structure at the 5′ end is one of the key features that enhances its translational activity and stability, thus underpinning the development of mRNA-based therapeutics. As the Dcp2 (Nudt20) is the major decapping enzyme in the 5′- to -3′ mRNA degradation pathway [122], recombinant hDcp2 is used in the in vitro assays to test stability of RNA transcripts capped with modified cap analogues or with analogues of the Cap 1 and Cap 2 structures (e.g., [55,56,83,103,105]). Dcp2 interacts with several regulatory proteins that act as decapping activators, including Dcp1 (S. pombe Dcp1 enhanced the Dcp2 catalytic activity 10-fold in vitro [123] or PNRC2 that, together with Dcp1, synergistically stimulates decapping activity of hDcp2 [124]. Both mentioned Dcp2 protein complexes were used in decapping susceptibility analysis of in vitro capped RNA: with N2-modified caps (here SpDcp1/Dcp2complex [89] or with N6-benzylated cap analogues (here: PNRC2-hDcp1/Dcp2 complex [107]. With regards to in-vitro-designed therapeutic (m)RNAs, next to the principal confirmation of enzymatic resistance of modified-cap –RNAs, a few additional aspects regarding Dcp2-mediated decapping should be taken into account. First, Dcp2 is not ubiquitously present throughout different human tissues [119]. Second, it is not involved in bulk mRNA decay in cells but rather is important for specific degradation pathways (e.g., nonsense-mediated decay, microRNA-mediated mRNA decay, or degradation in response to interferon [125,126]. Third, methylation status of adenosine adjacent to 7-methylguanosine in the cap structure (m7Gpppm6Am) was shown to be involved in decreased susceptibility to Dcp2 decapping [19] and that this phenomenon is related to selected transcripts rather than to general mRNA stabilization [105,127] (Table 2).
hNudt16 is another NUDIX enzyme used in analysis of modified-cap –RNA transcripts. It was initially identified as U8 snoRNA binding protein with decapping activity, which was shown for in vitro m7G- and m32,2,7G- capped RNA [15]. Further studies revealed that it is ubiquitously detected in all analysed human tissues, is responsible for decapping at least of a subset of mRNA in cells [119] and is able to hydrolyse m7G-capped RNA transcripts in vitro [119,128]. Recombinant hNudt16 appeared as a potent tool to test triphosphate bridge modification of trimethylated m7G cap analogues and capped RNAs in a study on m32,2,7G cap as a nuclear import signal and m32,2,7G-mediated transport of therapeutics [121,129]. A direct comparative analysis pointed out to non-methylated cap structure at the 5′ RNA transcript end (GpppG-, GpppA-) as preferred substrate for hNudt16 decapping [120] and that decapping of RNAs with mono- and trimethylated cap structures is influenced by the nucleotide type adjacent to m7G (or m32,2,7G)—those containing adenosine (or 2′-O adenosine) are very poor substrates to hNudt16 [120,121] (Table 2). This is in line with hNudt16 substrate specificity towards m7G dinucleotide cap analogues, where m7GpppA, m7GpppAm, and m7Gpppm6Am appeared resistant to hNudt16 hydrolysis [130]. Additionally, modification of guanosine moiety (but not m7G) at the C8-position in the dinucleotide compounds resulted in their resistance to hNudt16 (except for the pyrene substituent) [99]. hNudt16 preference to the exposed unmethylated guanosine in those C8-modified cap analogues was shown also for dinucleotide diphosphate compounds, where the methylated m7GppG was hydrolysed around eight-fold slower in comparison to GppG [131]. This specific hNudt16 feature was used in developing of the “cap-orientation” test in analysis of RNA transcripts with N2-susbstituted caps [92]. Here, the GpppG-capped transcript underwent complete decapping with hNudt16, whereas m7GpppG-RNA SP6 polymerase transcript (that is a mixture of correctly and in-reverse-orientation capped products, m7GpppG-RNA and GpppGm7-RNA) was decapped at around 45%. In the correct orientation, ARCA-capped transcript was decapped at around 8% in the same experimental conditions, which is attributed to the lower activity of hNudt16 towards m7G-capped RNA [92,120]. Functionality of that test was shown for mRNA transcripts capped in vitro with a series of N2-modified dinucleotide cap analogues (triazole, isoxazole or thiazole substituted) [92,93]. RNA transcripts bearing cap structure with the attached FAM moitety at 2′-OH or 3′-OH of m7G were also subject to decapping with hNudt16 [77]. Next to the aforementioned analogues of the canonical cap structure, hNudt16 showed enzymatic activity towards RNA capped with non-canonical metabolite 5′ caps (NAD, dpCoA and FAD) [132]. Such wide activity of hNudt16 towards different 5′ cap structures on in vitro prepared RNA transcripts points out to it as a convenient, single enzyme that can be widely used in testing functionality of introduced chemical modifications within a polyphosphate linkage or within a nucleotide part of cap structure, and in test of cap-orientation and capping efficiency of non-ARCA cap analogues. It can be also considered in removing of the incorrectly (reversely) capped RNA transcripts after transcription in vitro with RNA polymerase. In such a case, hNudt16 treatment would expose monophosphate at the 5′ end of RNA (e.g., pGm7-RNA, after GpppGm7-RNA decapping), which is then degraded by Xrn1 processesive 5′ to 3′ exoribonuclease (similarly to removing of 5′ triphosphorylated RNA from the primary transcripts after 5′RNA polyphosphatase treatment, e.g., [107]).
Mammalian Nudt12 NUDIX enzyme was also shown to possess decapping activity of m7GpppG- and GpppG-capped transcripts in vitro [116]. Subsequent reports identified human and murine Nudt12 as a cytosolic RNA deNADing enzyme [133,134] and its decapping activity towards dpCoA-capped RNA [132]. Trimethylated cap structure at the 5′ end of RNA has been also effectively hydrolyzed with Nudt12, similarly to unmethylated an monomethylated caps [130]. Studies on differentially methylated dinucleotide cap analogs pointed out towards compounds containing adenosine (with different methylation status, e.g., A, Am or m6Am) as preferred substrates for Nudt12, which is different in comparison to Nudt16 activity. For example, in the case of trimethylated analogues (m32,2,7GpppG and m32,2,7GpppA), hNudt12 efficiently hydrolysed dinucleotides with adenine but not guanine, whereas hNudt16 showed the opposite effect [130]. Thus, it seems reasonable to replace Nudt16 with Nudt12 in functional decapping studies of RNA transcripts modified cap analogues with adenosine as a nucleoside adjacent to initial m7G, as deNADing enzyme Nudt12 was already used in decapping test of RNAs with synthetic NAD analogues modified within diphosphate bridge [108].
Nudt2 was among other mammalian NUDIX enzymes with decapping activity towards canonical cap and metabolite FAD-cap and dpCoA cap [116,132]. In a recent report, following the discovery of diadenosine tetraphosphate capped RNAs, Nudt2 was shown to be responsible for the hydrolysis of AppppA-RNA [33]. Extended data for in vitro Nudt2-mediated decapping of ApnA-RNA, GpnG-RNA, ApnG-RNA, and GpnA-RNA showed its primary preference towards ApnA-capped substrates (Table 2). Among other dinucleotide structures those with tetraphosphate bridge were also effectively hydrolyzed. Interestingly, Ap3G-RNA appeared resistant to Nudt2 decapping, opposite to Gp3A-RNA [33]. Interestingly, Nudt2 hydrolase triphosphorylated pppN-RNA into monophosporylated pN-RNA, which serves as a substrate for Xrn1 exonuclease [135] and as such could be considered as another enzymatic activity that can be used in removing uncapped pppN-RNA transcripts, similarly to the earlier-mentioned approach that utilizes commercially available 5′ polyphosphatase.
Table 2.
Comparison of decapping activity of Dcp2, Nud16, Nudt12, Nudt2 towards RNA transcripts with indicated chemically modified and natural 5′ cap structures.
Table 2.
Comparison of decapping activity of Dcp2, Nud16, Nudt12, Nudt2 towards RNA transcripts with indicated chemically modified and natural 5′ cap structures.
| Dcp2 | Nudt16 | Nudt12 | Nudt2 | |
|---|---|---|---|---|
| m7GpppN | + [105] | |||
| m7GpppNm | + [105] | |||
| m7GpppG | + [92,105,120,136] | +/− [92,93,120] | + [116,130] | + [116] |
| m7GpppA | + [105] | − [120] | ||
| m7GpppGm | + [105] | +/− [120] | ||
| m7GpppAm | + [105] | − [120] | ||
| m7Gpppm6Am | + [105,107] − [19] (transcript dependent) | |||
| m7GpppAmGm | + [103] | |||
| m7Gpppbn6AmG | + [107] | |||
| GpppG | − [14] | + [92,93,120] | + [116,130] | + [33,116] |
| GpppA | + [120] | + [33] | ||
| ApppA | + [33] | |||
| ApppG | − [33] | |||
| GppppG | + [33] | |||
| GppppA | + [33] | |||
| AppppA | + [33] | |||
| AppppG | + [33] | |||
| m27,3′OGpppG | + [58,89,136] | − [92,93,120] | ||
| m27,2′OGpppG | + [38] | |||
| m27,2′OGppppG | + [136] | |||
| m27,2′-OGppCH2ppG | − [136] | |||
| m27,2′-OGppCCl2ppG | − [136] | |||
| m27,2′-OGppCF2ppG | − [136] | |||
| m27,2′-OGpp-tz-C2H4OppG | − [64] | |||
| m27,2′-OGppp-tz-C2H4OpG | + [64] | |||
| m27,2′OGppNHpN | − [56] | |||
| m27,3′OGppCH2pN | − [56,58] | |||
| m27,2′OGppSpG (D1) | −/+ [38] | |||
| m27,2′OGppSpG (D2) | −/+ [38] | |||
| m27,2′OGppBH3pG (D2) | − [38] | |||
| bn2m7GpppG | + [92] | −/+ [92] | ||
| bn2m7GppSpG | − [93] | |||
| bn2m27,3′OGpppG | + [89] | |||
| bn2m27,2′OGpppG | + [89] | |||
| bn2m7GppppG | + [93] | |||
| bn2m7GpppAmG | − [93] | |||
| (4-Cl-bn)2m7GpppG | + [92] | −/+ [92] | ||
| (4-Cl-bn)2m7GppppG | + [93] | |||
| (4-Cl-bn)2m7GpppAmG | − [93] | |||
| (4-OCH3-bn)2m7GpppG | + [92] | −/+ [92] | ||
| (4-OCH3-bn)2m27,3′OGpppN | + [89] | |||
| (4-di(OCH3-bn)-tz)2m7GpppG | + [92] | − [92] | ||
| (4-di(OCH3-bn)-tz-CH2)2m7GpppG | − [93] | |||
| (4-di(OCH3-bn)-tz-(CH2)2)2m7GpppG | +/− [93] | |||
| (4-di(OCH3-bn)-tz-(CH2)4)2m7GpppG | − [93] | |||
| (4-bn-isx)2m7GpppG | + [92] | −/+ [92] | ||
| (4-CH3-th)2m7GpppG | + [92] | −/+ [92] | ||
| 2′-O/3′-O-(FAM-L6N)-m7GpppG | + [77] | + [77] | ||
| m32,2,7GpppG | + [120,129] | + [130] | ||
| m32,2,7GpppA | −/+ [121] |
Human Nudt5 is the NUDIX hydrolase that has been linked to nucleotide metabolism and cancer [137]. Nudt5 did not show decapping activity towards m7GpppG- and GpppG-capped RNAs [116]. Recently, for the recombinant hNudt5, it showed decapping activity towards non-canonical UDP-GlcNAc cap on RNA transcripts in vitro [110] (Table 3). Thus, it could be a potential tool to analysis of UDP-glucose-derived cap synthetic analogues for future investigation of this 5′ end RNA modification in mammalian cells [37].
6. Other RNA Decapping Enzymes
Decapping of (m)RNA cap structures was shown in vitro for NUDIX enzymes in other domains of life—in plants [138], bacteria, or viruses [139]. Among four tested enzymes of the model plant Arabidopsis thaliana, the AtNUDT27 appeared as effective enzymes with a broad activity towards various cap structures: unmethylated, monomethylated m7G-caps, and non-canonical caps [138]. RppH bacterial NUDIX hydrolase was initially identified as activity that hydrolyzes 5′-triphoshate from RNA leaving 5′-monophosphate RNA [140], and subsequently it was demonstrated to cleave m7GpppG- and GpppG-capped RNA [116] and NAD-capped transcripts [133]. deNADing activity was also attributed to bacterial NudC enzyme [141].
Apart from the NUDIX-type decapping enzymes, at least three other classes of enzymes are involved in processing cap structures at the 5′–RNA end: DXO family proteins, HIT (histidine triad) proteins, and ApaH-like phosphatase found in trypanosomes [139]. In the case of DXO, it removes cap structure through cleavage of the entire dinucleotide from the RNA body (e.g., GpppG, m7GpppG, m32,2,7GpppG), with a preference towards unmethylated structure, which was connected to functions in the mechanism of 5′cap quality control [142]. 2′O-methylation of the first and the second transcribed nucleotide found in the Cap 1 and Cap 2 structures of mRNA, respectively, caused resistance to DXO decapping [101,143]. Apart from decapping activity, DXO can also remove non-canonical caps (NAD, FAD, and dpCoA) and has activity toward uncapped triphosphorylated RNA [144].
7. Discussion
Among mammalian NUDIX hydrolases with (m)RNA decapping in vitro, Dcp2 and Nudt16 have been used in the analysis of diverse analogues of canonical cap structure. However, mRNA decapping by Dcp2 is a complex process involving multiple regulatory proteins, such as Dcp1, Edc3, and Edc4. The formation of various Dcp2-containing multiprotein complexes ensures precise control over mRNA stability and degradation in vivo, which is crucial for cellular homeostasis and response to environmental changes [145,146]. Moreover, as mentioned earlier, Dcp2 controls stability only of a subset of mRNA in cells [115], exemplified by the recent global profiling study that revealed a number of Dcp2 mRNA substrates, including innate immunity-related transcripts [147,148]. Thus, for the more physiologically relevant results, in vitro decapping with Dcp2 should also be reconsidered in terms of the decapping complex with regulatory proteins and designed gene-specific mRNA (e.g., involved in innate immunity).
Available evidence points out to Nudt16 as a decapping enzyme with activity towards the widest range of different cap structures at 5′ end of RNA transcripts: umethylated (GpppN-), mono-methylated (m7GpppN-), trimethylated (m32,2,7GpppN-), NAD-, FAD-, and dpCoA-cap. Bearing in mind the broad substrate specificity, Nudt16 can be considered as the novel major standard in analysis of modified cap structures at (m)RNA transcripts produced in vitro, next to Dcp2. It already proved to be a potent tool in studies of caps modified in a triphosphate bridge and within 7-methylguanosine (N2 and 2′-OH or 3′-OH of m7G). Additionally, it could be used in cap-orientation assay and removing of the reversely capped (with non-ARCA cap analogues) RNA transcripts from IVT reactions (when combined with Xrn1 exoribonuclease). Important advantage of Nudt16 is that it acts effectively as a single subunit, without additional regulatory proteins, what can facilitate its use and optimization. One of the possible limitations is its use in regards to transcripts bearing at their 5′ end dinucleotide cap analogues with adenosine adjacent to the m7G (or m32,2,7G), which are poor substrates for Nudt16 decapping. However, the Nudt12 hydrolase could be used in such case instead. Finally, Nudt2 enzyme is active in vitro towards unmethylated NpnN-RNAs (where N = G or A), with lower activity towards those with triphosphate bridge (Np3N-RNAs) in comparison to di- or tetraphosphate-containing dinucleotide structures. As such, Nudt2 can be an attractive tool in testing modifications introduced into the polyphosphate chain of synthetic non-methylated dinucleotide analogues (e.g., GppXpG). Those in turn, after incorporation into (m)RNA during IVT reaction, could be subject to methylation by guanine-7-methyltranferase specific activity (e.g., Vaccina Capping Enzyme, VCE) or introduction, e.g., benzylic moieties with in-vitro-engineered MAT/MTase (methionine adenosylotrasferase/methyltransferse) activities [87]. Nudt2 also showed in vitro activity to m7GpppN-capped transcripts. Thus, it cannot be excluded from analysis of methylated cap structures. However, further characterization with a wider set of such analogues is needed in that context. Another Nudt2 activity of potentially great interest is its ability to convert triphosphorylated pppN-RNA into monophosporylated pN-RNA, the latter being a substrate for Xrn1-riboexonuclease-mediated removal from IVT reaction. Among identified mammalian NUDIX proteins with in vitro decapping activity towards canonical methylated and unmethylated cap structures are four additional enzymes: Nudt3, Nudt15, Nud17, and Nudt19 [116,117]. These represent an as-yet-unexplored field in the study of decapping of RNAs with modified cap analogues. As Nudt3 was shown also to be active in vivo toward a subset of mRNAs involved in cell migration regulation [118], it can be considered, next to the Nudt proteins described above, in future assay(s) aimed at the stability of RNA transcripts.
8. Conclusions
Next to the Dcp2, at least three other NUDIX enzymes were reviewed here—Nudt16, Nudt12, and Nudt2 comprise a set of activities that can be used in the 5′ capped RNA transcripts analysis that bears a diverse chemical modifications introduced into 5′ cap structure. They can be also considered in preparation of homogenous RNAs as a result of removal of incorrectly capped or 5′ triphosphorylated transcripts, or preparation of 5′ RNA end for downstream methodologies (e.g., CapZyme-Seq [149]). Given the known differences in the specificity of Nudt16, Nudt12, Nudt2, and Dcp2 towards natural and modified cap structures, the combinatorial use of these enzymes of human origin as a “NUDIX decapping toolbox” seems more relevant in the context of further development and use of capped mRNAs as a therapeutic molecule and their translation into the clinic. Finally, as they act effectively without additional activatory proteins, this facilitates their use and optimization.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T-cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Corbett, K.S.; Edwards, D.K.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schäfer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature 2020, 586, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef]
- Sahin, U.; Karikó, K.; Türeci, Ö. mRNA-based therapeutics—Developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef]
- Aygün, I.; Barciszewski, J. The forerunners and successful partnerships behind the BioNTech mRNA vaccine. J. Appl. Genet. 2023, 65, 47–55. [Google Scholar] [CrossRef]
- Shi, H.; Chai, P.; Jia, R.; Fan, X. Novel insight into the regulatory roles of diverse RNA modifications: Re-defining the bridge between transcription and translation. Mol. Cancer 2020, 19, 1–17. [Google Scholar] [CrossRef]
- Furuichi, Y. Discovery of m7G-cap in eukaryotic mRNAs. Proc. Jpn. Acad. Ser. B 2015, 91, 394–409. [Google Scholar] [CrossRef]
- Warminski, M.; Mamot, A.; Depaix, A.; Kowalska, J.; Jemielity, J. Chemical Modifications of mRNA Ends for Therapeutic Applications. Accounts Chem. Res. 2023, 56, 2814–2826. [Google Scholar] [CrossRef]
- Kurpiejewski, K.; Jankowska-Anyszka, M.; Grzela, R. N2 modified cap analogues as translation inhibitors and substrates for preparation of therapeutic mRNA. Eur. Biophys. J. 2023, 52, 511–519. [Google Scholar] [CrossRef]
- Dunckley, T.; Parker, R. The DCP2 protein is required for mRNA decapping in Saccharomyces cerevisiae and contains a functional MutT motif. EMBO J. 1999, 18, 5411–5422. [Google Scholar] [CrossRef]
- Lykke-Andersen, J. Identification of a Human Decapping Complex Associated with hUpf Proteins in Nonsense-Mediated Decay. Mol. Cell. Biol. 2002, 22, 8114–8121. [Google Scholar] [CrossRef] [PubMed]
- van Dijk, E.; Cougot, N.; Meyer, S.; Babajko, S.; Wahle, E.; Séraphin, B. Human Dcp2: A catalytically active mRNA decapping enzyme located in specific cytoplasmic structures. EMBO J. 2002, 21, 6915–6924. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jiao, X.; Carr-Schmid, A.; Kiledjian, M. The hDcp2 protein is a mammalian mRNA decapping enzyme. Proc. Natl. Acad. Sci. USA 2002, 99, 12663–12668. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.; Peterson, B.; Tomasevic, N.; Peculis, B.A. Xenopus U8 snoRNA Binding Protein Is a Conserved Nuclear Decapping Enzyme. Mol. Cell 2004, 13, 817–828. [Google Scholar] [CrossRef]
- Meyer, K.D.; Jaffrey, S.R. The dynamic epitranscriptome: N6-methyladenosine and gene expression control. Nat. Rev. Mol. Cell Biol. 2014, 15, 313–326. [Google Scholar] [CrossRef]
- Zamudio, J.R.; Mittra, B.; Zeiner, G.M.; Feder, M.; Bujnicki, J.M.; Sturm, N.R.; Campbell, D.A. Complete Cap 4 Formation Is Not Required for Viability in Trypanosoma brucei. Eukaryot. Cell 2006, 5, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Murai, R.; Hagiwara, H.; Hoshino, T.; Suyama, K. Preparation of eukaryotic mRNA having differently methylated adenosine at the 5′-terminus and the effect of the methyl group in translation. Nucleic Acids Symp. Ser. 2009, 53, 129–130. [Google Scholar] [CrossRef]
- Mauer, J.; Luo, X.; Blanjoie, A.; Jiao, X.; Grozhik, A.V.; Patil, D.P.; Linder, B.; Pickering, B.F.; Vasseur, J.-J.; Chen, Q.; et al. Reversible methylation of m6Am in the 5′ cap controls mRNA stability. Nature 2017, 541, 371–375. [Google Scholar] [CrossRef]
- Warminski, M.; Sikorski, P.J.; Kowalska, J.; Jemielity, J. Applications of Phosphate Modification and Labeling to Study (m)RNA Caps. Top. Curr. Chem. 2017, 375, 1–29. [Google Scholar] [CrossRef]
- Xhemalce, B.; Robson, S.C.; Kouzarides, T. Human RNA Methyltransferase BCDIN3D Regulates MicroRNA Processing. Cell 2012, 151, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Wurth, L.; Gribling-Burrer, A.-S.; Verheggen, C.; Leichter, M.; Takeuchi, A.; Baudrey, S.; Martin, F.; Krol, A.; Bertrand, E.; Allmang, C. Hypermethylated-capped selenoprotein mRNAs in mammals. Nucleic Acids Res. 2014, 42, 8663–8677. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Fritz, S.E.; Seufzer, B.; Boris-Lawrie, K. The mRNA encoding the JUND tumor suppressor detains nuclear RNA-binding proteins to assemble polysomes that are unaffected by mTOR. J. Biol. Chem. 2020, 295, 7763–7773. [Google Scholar] [CrossRef] [PubMed]
- Zucko, D.; Boris-Lawrie, K. Blocking tri-methylguanosine synthase 1 (TGS1) stops anchorage-independent growth of canine sarcomas. Cancer Gene Ther. 2023, 30, 1274–1284. [Google Scholar] [CrossRef]
- Ohira, T.; Suzuki, T. Precursors of tRNAs are stabilized by methylguanosine cap structures. Nat. Chem. Biol. 2016, 12, 648–655. [Google Scholar] [CrossRef]
- Abdelhamid, R.F.; Plessy, C.; Yamauchi, Y.; Taoka, M.; de Hoon, M.; Gingeras, T.R.; Isobe, T.; Carninci, P. Multiplicity of 5′ Cap Structures Present on Short RNAs. PLoS ONE 2014, 9, e102895. [Google Scholar] [CrossRef]
- Mattay, J. Noncanonical metabolite RNA caps: Classification, quantification, (de)capping, and function. Wiley Interdiscip. Rev. RNA 2022, 13, e1730. [Google Scholar] [CrossRef]
- Walters, R.W.; Matheny, T.; Mizoue, L.S.; Rao, B.S.; Muhlrad, D.; Parker, R. Identification of NAD + capped mRNAs in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2016, 114, 480–485. [Google Scholar] [CrossRef]
- Jiao, X.; Doamekpor, S.K.; Bird, J.G.; Nickels, B.E.; Tong, L.; Hart, R.P.; Kiledjian, M. 5′ End Nicotinamide Adenine Dinucleotide Cap in Human Cells Promotes RNA Decay through DXO-Mediated deNADding. Cell 2017, 168, 1015–1027.e10. [Google Scholar] [CrossRef]
- Kiledjian, M. Eukaryotic RNA 5′-End NAD + Capping and DeNADding. Trends Cell Biol. 2018, 28, 454–464. [Google Scholar] [CrossRef]
- Wiedermannová, J.; Julius, C.; Yuzenkova, Y. The expanding field of non-canonical RNA capping: New enzymes and mechanisms. R. Soc. Open Sci. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Zhang, H.; Zhu, Z.; Ji, F.; He, Z.; Yang, Z.; Xia, Y.; Cai, Z. DpCoA tagSeq: Barcoding dpCoA-Capped RNA for Direct Nanopore Sequencing via Maleimide-Thiol Reaction. Anal. Chem. 2023, 95, 11124–11131. [Google Scholar] [CrossRef] [PubMed]
- Potužník, J.F.; Nešuta, O.; Škríba, A.; Voleníková, B.; Mititelu, M.; Mancini, F.; Serianni, V.; Fernandez, H.; Spustová, K.; Trylčová, J.; et al. Diadenosine Tetraphosphate (Ap4A) Serves as a 5′ RNA Cap in Mammalian Cells. Angew. Chem. Int. Ed. 2023, 63, e202314951. [Google Scholar] [CrossRef]
- Martinez, I.; Hayes, K.E.; Barr, J.A.; Harold, A.D.; Xie, M.; Bukhari, S.I.A.; Vasudevan, S.; Steitz, J.A.; DiMaio, D. An Ex-portin-1-dependent microRNA biogenesis pathway during human cell quiescence. Proc. Natl. Acad. Sci. USA 2017, 114, E4961–E4970. [Google Scholar] [CrossRef]
- Singh, R.; Reddy, R. Gamma-monomethyl phosphate: A cap structure in spliceosomal U6 small nuclear RNA. Proc. Natl. Acad. Sci. USA 1989, 86, 8280–8283. [Google Scholar] [CrossRef]
- Gupta, S.; Busch, R.K.; Singh, R.; Reddy, R. Characterization of U6 small nuclear RNA cap-specific antibodies. Identification of gamma-monomethyl-GTP cap structure in 7SK and several other human small RNAs. J. Biol. Chem. 1990, 265, 19137–19142. [Google Scholar] [CrossRef]
- Wang, J.; Chew, B.L.A.; Lai, Y.; Dong, H.; Xu, L.; Balamkundu, S.; Cai, W.M.; Cui, L.; Liu, C.F.; Fu, X.-Y.; et al. Quantifying the RNA cap epitranscriptome reveals novel caps in cellular and viral RNA. Nucleic Acids Res. 2019, 47, e130. [Google Scholar] [CrossRef] [PubMed]
- Grudzien-Nogalska, E.; Stepinski, J.; Jemielity, J.; Zuberek, J.; Stolarski, R.; Rhoads, R.E.; Darzynkiewicz, E. Synthesis of anti-reverse cap analogs (ARCAs) and their applications in mRNA translation and stability. Methods Enzymol. 2007, 431, 203–227. [Google Scholar] [CrossRef]
- Li, S.; Jia, Y.; Jacobson, B.; McCauley, J.; Kratzke, R.; Bitterman, P.B.; Wagner, C.R. Treatment of Breast and Lung Cancer Cells with a N-7 Benzyl Guanosine Monophosphate Tryptamine Phosphoramidate Pronucleotide (4Ei-1) Results in Chemosensitization to Gemcitabine and Induced eIF4E Proteasomal Degradation. Mol. Pharm. 2013, 10, 523–531. [Google Scholar] [CrossRef]
- Golojuch, S.; Kopcial, M.; Strzelecka, D.; Kasprzyk, R.; Baran, N.; Sikorski, P.J.; Kowalska, J.; Jemielity, J. Exploring tryptamine conjugates as pronucleotides of phosphate-modified 7-methylguanine nucleotides targeting cap-dependent transla-tion. Bioorganic Med. Chem. 2020, 28, 115523. [Google Scholar] [CrossRef]
- Kurpiejewski, K.; Piecyk, K.; Lukaszewicz, M.; Kamel, K.; Chmurski, K.; Kmiecik, S.; Jankowska-Anyszka, M. The Synergistic Effect of N2 and N7 Modifications on the Inhibitory Efficacy of mRNA Cap Analogues. Pharmaceuticals 2024, 17, 632. [Google Scholar] [CrossRef] [PubMed]
- Konarska, M.M.; Padgett, R.A.; Sharp, P.A. Recognition of cap structure in splicing in vitro of mRNA precursors. Cell 1984, 38, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Darzynkiewicz, E.; Stepinski, J.; Ekiel, I.; Jin, Y.; Haber, D.; Sijuwade, T.; Tahara, S. Beta-globin mRNAs capped with m7G, m22.7G or m32.2.7G differ in intrinsic translation efficiency. Nucleic Acids Res. 1988, 16, 8953–8962. [Google Scholar] [CrossRef]
- Yisraeli, J.K.; Melton, D.A. Synthesis of long, capped transcripts in vitro by SP6 and T7 RNA polymerases. Methods Enzymol. 1989, 180, 42–50. [Google Scholar] [CrossRef]
- Coleman, T.M. Superior 5′ homogeneity of RNA from ATP-initiated transcription under the T7 2.5 promoter. Nucleic Acids Res. 2004, 32, 14e. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Dahlberg, J.E.; Lund, E. Reverse 5′ caps in RNAs made in vitro by phage RNA polymerases. RNA 1995, 1, 957–967. [Google Scholar]
- Stepinski, J.; Waddell, C.; Stolarski, R.; Darzynkiewicz, E.; Rhoads, R.E. Synthesis and properties of mRNAs containing the novel “anti-reverse” cap analogs 7-methyl(3′-O-methyl)GpppG and 7-methyl (3′-deoxy)GpppG. RNA 2001, 7, 1486–1495. [Google Scholar]
- Jemielity, J.; Fowler, T.; Zuberek, J.; Stepinski, J.; Lewdorowicz, M.; Niedzwiecka, A.; Stolarski, R.; Darzynkiewicz, E.; Rhoads, R.E. Novel “anti-reverse” cap analogs with superior translational properties. RNA 2003, 9, 1108–1122. [Google Scholar] [CrossRef] [PubMed]
- Cai, A.; Jankowska-Anyszka, M.; Centers, A.; Chlebicka, L.; Stepinski, J.; Stolarski, R.; Darzynkiewicz, E.; Rhoads, R.E. Quantitative Assessment of mRNA Cap Analogues as Inhibitors of in Vitro Translation. Biochemistry 1999, 38, 8538–8547. [Google Scholar] [CrossRef]
- Zuberek, J.; Wyslouch-Cieszynska, A.; Niedzwiecka, A.; Dadlez, M.; Stepinski, J.; Augustyniak, W.; Gingras, A.-C.; Zhang, Z.; Burley, S.K.; Sonenberg, N.; et al. Phosphorylation of eIF4E attenuates its interaction with mRNA cap analogs by electrostatic repulsion: Intein-mediated protein ligation strategy to obtain phosphorylated protein. RNA 2003, 9, 52–61. [Google Scholar] [CrossRef]
- Kowalska, J.; Lewdorowicz, M.; Zuberek, J.; Bojarska, E.; Wojcik, J.; Cohen, L.S.; Davis, R.E.; Stepinski, J.; Stolarski, R.; Darzynkiewicz, E.; et al. Synthesis and Properties of Mrna cap Analogs Containing Phosphorothioate Moiety In 5′,5′-Triphosphate Chain. Nucleosides Nucleotides Nucleic Acids 2005, 24, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, J.; Lewdorowicz, M.; Zuberek, J.; Grudzien-Nogalska, E.; Bojarska, E.; Stepinski, J.; Rhoads, R.E.; Darzynkiewicz, E.; Davis, R.E.; Jemielity, J. Synthesis and characterization of mRNA cap analogs containing phosphorothioate substitutions that bind tightly to eIF4E and are resistant to the decapping pyrophosphatase DcpS. RNA 2008, 14, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, J.; Lukaszewicz, M.; Zuberek, J.; Darzynkiewicz, E.; Jemielity, J. Phosphoroselenoate Dinucleotides for Modification of mRNA 5′ End. ChemBioChem 2009, 10, 2469–2473. [Google Scholar] [CrossRef]
- Kowalska, J.; del Nogal, A.W.; Darzynkiewicz, Z.M.; Buck, J.; Nicola, C.; Kuhn, A.N.; Lukaszewicz, M.; Zuberek, J.; Strenkowska, M.; Ziemniak, M.; et al. Synthesis, properties, and biological activity of boranophosphate analogs of the mRNA cap: Versatile tools for manipulation of therapeutically relevant cap-dependent processes. Nucleic Acids Res. 2014, 42, 10245–10264. [Google Scholar] [CrossRef]
- Grudzien-Nogalska, E.; Jemielity, J.; Kowalska, J.; Darzynkiewicz, E.; Rhoads, R.E. Phosphorothioate cap analogs stabilize mRNA and increase translational efficiency in mammalian cells. RNA 2007, 13, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Slepenkov, S.; Grudzien-Nogalska, E.; Kowalska, J.; Kulis, M.; Zuberek, J.; Lukaszewicz, M.; Darzynkiewicz, E.; Jemielity, J.; Rhoads, R.E. Translation, stability, and resistance to decapping of mRNAs containing caps substituted in the triphosphate chain with BH3, Se, and NH. RNA 2011, 17, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Kalek, M.; Jemielity, J.; Darzynkiewicz, Z.M.; Bojarska, E.; Stepinski, J.; Stolarski, R.; Davis, R.E.; Darzynkiewicz, E. Enzymatically stable 5′ mRNA cap analogs: Synthesis and binding studies with human DcpS decapping enzyme. Bioorganic Med. Chem. 2006, 14, 3223–3230. [Google Scholar] [CrossRef]
- Grudzien, E.; Kalek, M.; Jemielity, J.; Darzynkiewicz, E.; Rhoads, R.E. Differential Inhibition of mRNA Degradation Pathways by Novel Cap Analogs. J. Biol. Cgem. 2006, 281, 1857–1867. [Google Scholar] [CrossRef]
- Rydzik, A.M.; Lukaszewicz, M.; Zuberek, J.; Kowalska, J.; Darzynkiewicz, Z.M.; Darzynkiewicz, E.; Jemielity, J. Synthetic dinucleotide mRNA cap analogs with tetraphosphate 5′,5′ bridge containing methylenebis(phosphonate) modification. Org. Biomol. Chem. 2009, 7, 4763–4776. [Google Scholar] [CrossRef]
- Ziemniak, M.; Kowalska, J.; Lukaszewicz, M.; Zuberek, J.; Wnek, K.; Darzynkiewicz, E.; Jemielity, J. Phosphate-modified analogues of m 7 GTP and m 7 Gppppm 7 G—Synthesis and biochemical properties. Bioorganic Med. Chem. 2015, 23, 5369–5381. [Google Scholar] [CrossRef]
- Strenkowska, M.; Grzela, R.; Majewski, M.; Wnek, K.; Kowalska, J.; Lukaszewicz, M.; Zuberek, J.; Darzynkiewicz, E.; Kuhn, A.N.; Sahin, U.; et al. Cap analogs modified with 1,2-dithiodiphosphate moiety protect mRNA from decapping and enhance its translational potential. Nucleic Acids Res. 2016, 44, 9578–9590. [Google Scholar] [CrossRef] [PubMed]
- Wojtczak, B.A.; Sikorski, P.J.; Fac-Dabrowska, K.; Nowicka, A.; Warminski, M.; Kubacka, D.; Nowak, E.; Nowotny, M.; Kowalska, J.; Jemielity, J. 5′-Phosphorothiolate Dinucleotide Cap Analogues: Reagents for Messenger RNA Modification and Potent Small-Molecular Inhibitors of Decapping Enzymes. J. Am. Chem. Soc. 2018, 140, 5987–5999. [Google Scholar] [CrossRef] [PubMed]
- Walczak, S.; Nowicka, A.; Kubacka, D.; Fac, K.; Wanat, P.; Mroczek, S.; Kowalska, J.; Jemielity, J. A novel route for preparing 5′ cap mimics and capped RNAs: Phosphate-modified cap analogues obtained via click chemistry. Chem. Sci. 2017, 8, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Walczak, S.; Sikorski, P.J.; Kasprzyk, R.; Kowalska, J.; Jemielity, J. Exploring the potential of phosphotriazole 5′ mRNA cap analogues as efficient translation initiators. Org. Biomol. Chem. 2018, 16, 6741–6748. [Google Scholar] [CrossRef]
- Kozarski, M.; Drazkowska, K.; Bednarczyk, M.; Warminski, M.; Jemielity, J.; Kowalska, J. Towards superior mRNA caps accessible by click chemistry: Synthesis and translational properties of triazole-bearing oligonucleotide cap analogs. RSC Adv. 2023, 13, 12809–12824. [Google Scholar] [CrossRef]
- Kore, A.R.; Shanmugasundaram, M.; Charles, I.; Cheng, A.M.; Barta, T.J. Synthesis and application of 2′-fluoro-substituted cap analogs. Bioorganic Med. Chem. Lett. 2007, 17, 5295–5299. [Google Scholar] [CrossRef]
- Kore, A.R.; Charles, I. Synthesis and evaluation of 2′-O-allyl substituted dinucleotide cap analog for mRNA translation. Bioorganic Med. Chem. 2010, 18, 8061–8065. [Google Scholar] [CrossRef]
- Shanmugasundaram, M.; Charles, I.; Kore, A.R. Design, synthesis and biological evaluation of dinucleotide mRNA cap analog containing propargyl moiety. Bioorganic Med. Chem. 2016, 24, 1204–1208. [Google Scholar] [CrossRef]
- Senthilvelan, A.; Vonderfecht, T.; Shanmugasundaram, M.; Potter, J.; Kore, A.R. Click-iT trinucleotide cap analog: Synthesis, mRNA translation, and detection. Bioorganic Med. Chem. 2023, 77, 117128. [Google Scholar] [CrossRef]
- Kore, A.R.; Shanmugasundaram, M.; Vlassov, A.V. Synthesis and application of a new 2′,3′-isopropylidene guanosine substituted cap analog. Bioorganic Med. Chem. Lett. 2008, 18, 4828–4832. [Google Scholar] [CrossRef]
- Warminski, M.; Kowalska, J.; Buck, J.; Zuberek, J.; Lukaszewicz, M.; Nicola, C.; Kuhn, A.N.; Sahin, U.; Darzynkiewicz, E.; Jemielity, J. The synthesis of isopropylidene mRNA cap analogs modified with phosphorothioate moiety and their evaluation as promoters of mRNA translation. Bioorganic Med. Chem. Lett. 2013, 23, 3753–3758. [Google Scholar] [CrossRef] [PubMed]
- Ziemniak, M.; Szabelski, M.; Lukaszewicz, M.; Nowicka, A.; Darzynkiewicz, E.; Rhoads, R.E.; Wieczorek, Z.; Jemielity, J. Synthesis and evaluation of fluorescent cap analogues for mRNA labelling. RSC Adv. 2013, 3, 20943–20958. [Google Scholar] [CrossRef] [PubMed]
- Kore, A.R.; Shanmugasundaram, M.; Charles, I.; Vlassov, A.V.; Barta, T.J. Locked Nucleic Acid (LNA)-Modified Dinucleotide mRNA Cap Analogue: Synthesis, Enzymatic Incorporation, and Utilization. J. Am. Chem. Soc. 2009, 131, 6364–6365. [Google Scholar] [CrossRef] [PubMed]
- Senthilvelan, A.; Vonderfecht, T.; Shanmugasundaram, M.; Pal, I.; Potter, J.; Kore, A.R. Trinucleotide Cap Analogue Bearing a Locked Nucleic Acid Moiety: Synthesis, mRNA Modification, and Translation for Therapeutic Applications. Org. Lett. 2021, 23, 4133–4136. [Google Scholar] [CrossRef]
- Jemielity, J.; Lukaszewicz, M.; Kowalska, J.; Czarnecki, J.; Zuberek, J.; Darzynkiewicz, E. Synthesis of biotin labelled cap analogue–incorporable into mRNA transcripts and promoting cap-dependent translation. Org. Biomol. Chem. 2012, 10, 8570–8574. [Google Scholar] [CrossRef]
- Bednarek, S.; Madan, V.; Sikorski, P.J.; Bartenschlager, R.; Kowalska, J.; Jemielity, J. mRNAs biotinylated within the 5′ cap and protected against decapping: New tools to capture RNA–protein complexes. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20180167. [Google Scholar] [CrossRef]
- Warminski, M.; Sikorski, P.J.; Warminska, Z.; Lukaszewicz, M.; Kropiwnicka, A.; Zuberek, J.; Darzynkiewicz, E.; Kowalska, J.; Jemielity, J. Amino-Functionalized 5′ Cap Analogs as Tools for Site-Specific Sequence-Independent Labeling of mRNA. Bioconjugate Chem. 2017, 28, 1978–1992. [Google Scholar] [CrossRef]
- Warminski, M.; Grab, K.; Szczepanski, K.; Spiewla, T.; Zuberek, J.; Kowalska, J.; Jemielity, J. Photoactivatable mRNA 5′ Cap Analogs for RNA-Protein Crosslinking. Adv. Sci. 2024, e2400994. [Google Scholar] [CrossRef]
- Mamot, A.; Sikorski, P.J.; Warminski, M.; Kowalska, J.; Jemielity, J. Azido-Functionalized 5′ Cap Analogues for the Preparation of Translationally Active mRNAs Suitable for Fluorescent Labeling in Living Cells. Angew. Chem. Int. Ed. 2017, 56, 15628–15632. [Google Scholar] [CrossRef]
- Matsuo, H.; Li, H.; McGuire, A.M.; Fletcher, C.M.; Gingras, A.-C.; Sonenberg, N.; Wagner, G. Structure of translation factor elF4E bound to m7GDP and interaction with 4E-binding protein. Nat. Struct. Mol. Biol. 1997, 4, 717–724. [Google Scholar] [CrossRef]
- Marcotrigiano, J.; Gingras, A.-C.; Sonenberg, N.; Burley, S.K. Cocrystal Structure of the Messenger RNA 5′ Cap-Binding Protein (eIF4E) Bound to 7-methyl-GDP. Cell 1997, 89, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Niedzwiecka, A.; Marcotrigiano, J.; Stepinski, J.; Jankowska-Anyszka, M.; Wyslouch-Cieszynska, A.; Dadlez, M.; Gingras, A.-C.; Mak, P.; Darzynkiewicz, E.; Sonenberg, N.; et al. Biophysical Studies of eIF4E Cap-binding Protein: Recognition of mRNA 5′ Cap Structure and Synthetic Fragments of eIF4G and 4E-BP1 Proteins. J. Mol. Biol. 2002, 319, 615–635. [Google Scholar] [CrossRef] [PubMed]
- Grudzien, E.; Stepinski, J.; Jankowska-Anyszka, M.; Stolarski, R.; Darzynkiewicz, E.; Rhoads, R.E. Novel cap analogs for in vitro synthesis of mRNAs with high translational efficiency. RNA 2004, 10, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Kore, A.R.; Xiao, Z.; Li, M. Synthesis and biological validation of N7-(4-chlorophenoxyethyl) substituted dinucleotide cap analogs for mRNA translation. Bioorganic Med. Chem. 2013, 21, 4570–4574. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, R.; Baranowski, M.R.; Markiewicz, L.; Kubacka, D.; Bednarczyk, M.; Baran, N.; Wojtczak, A.; Sikorski, P.J.; Zuberek, J.; Kowalska, J.; et al. Novel N7-Arylmethyl Substituted Dinucleotide mRNA 5′ cap Analogs: Synthesis and Evaluation as Modulators of Translation. Pharmaceutics 2021, 13, 1941. [Google Scholar] [CrossRef]
- Jia, Y.; Chiu, T.-L.; Amin, E.A.; Polunovsky, V.; Bitterman, P.B.; Wagner, C.R. Design, synthesis and evaluation of analogs of initiation factor 4E (eIF4E) cap-binding antagonist Bn7-GMP. Eur. J. Med. Chem. 2010, 45, 1304–1313. [Google Scholar] [CrossRef]
- Cornelissen, N.V.; Mineikaitė, R.; Erguven, M.; Muthmann, N.; Peters, A.; Bartels, A.; Rentmeister, A. Post-synthetic benzylation of the mRNA 5′ cap via enzymatic cascade reactions. Chem. Sci. 2023, 14, 10962–10970. [Google Scholar] [CrossRef]
- Piecyk, K.; Davis, R.E.; Jankowska-Anyszka, M. Synthesis of N2-modified 7-methylguanosine 5′-monophosphates as nematode translation inhibitors. Bioorganic Med. Chem. 2012, 20, 4781–4789. [Google Scholar] [CrossRef]
- Kocmik, I.; Piecyk, K.; Rudzinska, M.; Niedzwiecka, A.; Darzynkiewicz, E.; Grzela, R.; Jankowska-Anyszka, M. Modified ARCA analogs providing enhanced translational properties of capped mRNAs. Cell Cycle 2018, 17, 1624–1636. [Google Scholar] [CrossRef]
- Piecyk, K.; Lukaszewicz, M.; Darzynkiewicz, E.; Jankowska-Anyszka, M. Triazole-containing monophosphate mRNA cap analogs as effective translation inhibitors. RNA 2014, 20, 1539–1547. [Google Scholar] [CrossRef]
- Piecyk, K.; Lukaszewicz, M.; Kamel, K.; Janowska, M.; Pietrow, P.; Kmiecik, S.; Jankowska-Anyszka, M. Isoxazole-containing 5′ mRNA cap analogues as inhibitors of the translation initiation process. Bioorganic Chem. 2020, 96, 103583. [Google Scholar] [CrossRef] [PubMed]
- Grzela, R.; Piecyk, K.; Stankiewicz-Drogon, A.; Pietrow, P.; Lukaszewicz, M.; Kurpiejewski, K.; Darzynkiewicz, E.; Jankowska-Anyszka, M. N2 modified dinucleotide cap analogs as a potent tool for mRNA engineering. RNA 2023, 29, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Kurpiejewski, K.; Stankiewicz-Drogon, A.; Piecyk, K.; Rajkowska, E.; Skrzypczyk, P.; Geng, J.; Darzynkiewicz, E.; Grzela, R.; Jankowska-Anyszka, M. The potential of N2-modified cap analogues for precise genetic manipulation through mRNA engineering. Front. Mol. Biosci. 2024, 10, 1269028. [Google Scholar] [CrossRef]
- Klöcker, N.; Weissenboeck, F.P.; van Dülmen, M.; Špaček, P.; Hüwel, S.; Rentmeister, A. Photocaged 5′ cap analogues for optical control of mRNA translation in cells. Nat. Chem. 2022, 14, 905–913. [Google Scholar] [CrossRef]
- Weissenboeck, F.P.; Klöcker, N.; Špaček, P.; Hüwel, S.; Rentmeister, A. Stabilized 5′ Cap Analogue for Optochemical Activation of mRNA Translation. ACS Omega 2024, 9, 12810–12816. [Google Scholar] [CrossRef]
- Inagaki, M.; Abe, N.; Li, Z.; Nakashima, Y.; Acharyya, S.; Ogawa, K.; Kawaguchi, D.; Hiraoka, H.; Banno, A.; Meng, Z.; et al. Cap analogs with a hydrophobic photocleavable tag enable facile purification of fully capped mRNA with various cap structures. Nat. Commun. 2023, 14, 1–17. [Google Scholar] [CrossRef]
- Anhäuser, L.; Klöcker, N.; Muttach, F.; Mäsing, F.; Špaček, P.; Studer, A.; Rentmeister, A. A Benzophenone-Based Photocaging Strategy for the N7 Position of Guanosine. Angew. Chem. Int. Ed. 2019, 59, 3161–3165. [Google Scholar] [CrossRef]
- Bollu, A.; Schepers, H.; Klöcker, N.; Erguven, M.; Lawrence-Dörner, A.; Rentmeister, A. Visible Light Activates Coumarin-Caged mRNA for Cytoplasmic Cap Methylation in Cells. Chem.–A Eur. J. 2023, 30, e202303174. [Google Scholar] [CrossRef] [PubMed]
- Wojtczak, B.A.; Bednarczyk, M.; Sikorski, P.J.; Wojtczak, A.; Surynt, P.; Kowalska, J.; Jemielity, J. Synthesis and Evaluation of Diguanosine Cap Analogs Modified at the C8-Position by Suzuki–Miyaura Cross-Coupling: Discovery of 7-Methylguanosine-Based Molecular Rotors. J. Org. Chem. 2023, 88, 6827–6846. [Google Scholar] [CrossRef]
- Hyde, J.L.; Diamond, M.S. Innate immune restriction and antagonism of viral RNA lacking 2′-O methylation. Virology 2015, 479-480, 66–74. [Google Scholar] [CrossRef]
- Schuberth-Wagner, C.; Ludwig, J.; Bruder, A.K.; Herzner, A.-M.; Zillinger, T.; Goldeck, M.; Schmidt, T.; Schmid-Burgk, J.L.; Kerber, R.; Wolter, S.; et al. A Conserved Histidine in the RNA Sensor RIG-I Controls Immune Tolerance to N1-2′O-Methylated Self RNA. Immunity 2015, 43, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Despic, V.; Jaffrey, S.R. mRNA ageing shapes the Cap2 methylome in mammalian mRNA. Nature 2023, 614, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Drazkowska, K.; Tomecki, R.; Warminski, M.; Baran, N.; Cysewski, D.; Depaix, A.; Kasprzyk, R.; Kowalska, J.; Jemielity, J.; Sikorski, P.J. 2′-O-Methylation of the second transcribed nucleotide within the mRNA 5′ cap impacts the protein production level in a cell-specific manner and contributes to RNA immune evasion. Nucleic Acids Res. 2022, 50, 9051–9071. [Google Scholar] [CrossRef]
- Henderson, J.M.; Ujita, A.; Hill, E.; Yousif-Rosales, S.; Smith, C.; Ko, N.; McReynolds, T.; Cabral, C.R.; Escamilla-Powers, J.R.; Houston, M.E. Cap 1 Messenger RNA Synthesis with Co-transcriptional CleanCap® Analog by In Vitro Transcription. Curr. Protoc. 2021, 1, e39. [Google Scholar] [CrossRef] [PubMed]
- Sikorski, P.J.; Warminski, M.; Kubacka, D.; Ratajczak, T.; Nowis, D.; Kowalska, J.; Jemielity, J. The identity and methylation status of the first transcribed nucleotide in eukaryotic mRNA 5′ cap modulates protein expression in living cells. Nucleic Acids Res. 2020, 48, 1607–1626. [Google Scholar] [CrossRef]
- Wojtczak, A.; Kasprzyk, R.; Warmiński, M.; Ubych, K.; Kubacka, D.; Sikorski, P.J.; Jemielity, J.; Kowalska, J. Evaluation of carboxyfluorescein-labeled 7-methylguanine nucleotides as probes for studying cap-binding proteins by fluorescence anisotropy. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Warminski, M.; Trepkowska, E.; Smietanski, M.; Sikorski, P.J.; Baranowski, M.R.; Bednarczyk, M.; Kedzierska, H.; Majewski, B.; Mamot, A.; Papiernik, D.; et al. Trinucleotide mRNA Cap Analogue N6-Benzylated at the Site of Posttranscriptional m6Am Mark Facilitates mRNA Purification and Confers Superior Translational Properties In Vitro and In Vivo. J. Am. Chem. Soc. 2024, 146, 8149–8163. [Google Scholar] [CrossRef]
- Mlynarska-Cieslak, A.; Depaix, A.; Grudzien-Nogalska, E.; Sikorski, P.J.; Warminski, M.; Kiledjian, M.; Jemielity, J.; Kowalska, J. Nicotinamide-Containing Di- and Trinucleotides as Chemical Tools for Studies of NAD-Capped RNAs. Org. Lett. 2018, 20, 7650–7655. [Google Scholar] [CrossRef]
- Depaix, A.; Grudzien-Nogalska, E.; Fedorczyk, B.; Kiledjian, M.; Jemielity, J.; Kowalska, J. Preparation of RNAs with non-canonical 5′ ends using novel di- and trinucleotide reagents for co-transcriptional capping. Front. Mol. Biosci. 2022, 9, 854170. [Google Scholar] [CrossRef]
- Weber, F.; Motzkus, N.A.; Brandl, L.; Möhler, M.; Alempijevic, A.; Jäschke, A. Identification and in vitro characterization of UDP-GlcNAc-RNA cap-modifying and decapping enzymes. Nucleic Acids Res. 2024, 52, 5438–5450. [Google Scholar] [CrossRef]
- McLennan, A.G. The Nudix hydrolase superfamily. Cell. Mol. Life Sci. 2006, 63, 123–143. [Google Scholar] [CrossRef] [PubMed]
- McLennan, A.G. Substrate ambiguity among the nudix hydrolases: Biologically significant, evolutionary remnant, or both? Cell. Mol. Life Sci. 2013, 70, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Bessman, M.J.; Frick, D.N.; O’Handley, S.F. The MutT Proteins or “Nudix” Hydrolases, a Family of Versatile, Widely Distributed, “Housecleaning” Enzymes. J. Biol. Chem. 1996, 271, 25059–25062. [Google Scholar] [CrossRef] [PubMed]
- Mildvan, A.; Xia, Z.; Azurmendi, H.; Saraswat, V.; Legler, P.; Massiah, M.; Gabelli, S.; Bianchet, M.; Kang, L.-W.; Amzel, L. Structures and mechanisms of Nudix hydrolases. Arch. Biochem. Biophys. 2005, 433, 129–143. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamagata, Y. Visualization of mutagenic nucleotide processing by Escherichia coli MutT, a Nudix hydrolase. Proc. Natl. Acad. Sci. USA 2022, 119, e2203118119. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-G.; Bail, S.; Kiledjian, M. Multiple Nudix family proteins possess mRNA decapping activity. RNA 2013, 19, 390–399. [Google Scholar] [CrossRef]
- Grudzien-Nogalska, E.; Kiledjian, M. New insights into decapping enzymes and selective mRNA decay. Wiley Interdiscip. Rev. RNA 2016, 8, e1379. [Google Scholar] [CrossRef]
- Grudzien-Nogalska, E.; Jiao, X.; Song, M.-G.; Hart, R.P.; Kiledjian, M. Nudt3 is an mRNA decapping enzyme that modulates cell migration. RNA 2016, 22, 773–781. [Google Scholar] [CrossRef]
- Song, M.-G.; Li, Y.; Kiledjian, M. Multiple mRNA Decapping Enzymes in Mammalian Cells. Mol. Cell 2010, 40, 423–432. [Google Scholar] [CrossRef]
- Grzela, R.; Nasilowska, K.; Lukaszewicz, M.; Tyras, M.; Stepinski, J.; Jankowska-Anyszka, M.; Bojarska, E.; Darzynkiewicz, E.; Grzela, R.; Nasilowska, K.; et al. Hydrolytic activity of human Nudt16 enzyme on dinucleotide cap analogs and short capped oligonucleotides. RNA 2018, 24, 633–642. [Google Scholar] [CrossRef]
- Wojtczak, B.A.; Warminski, M.; Kowalska, J.; Lukaszewicz, M.; Honcharenko, M.; Smith, C.I.E.; Strömberg, R.; Darzynkiewicz, E.; Jemielity, J. Clickable trimethylguanosine cap analogs modified within the triphosphate bridge: Synthesis, conjugation to RNA and susceptibility to degradation. RSC Adv. 2016, 6, 8317–8328. [Google Scholar] [CrossRef]
- Garneau, N.L.; Wilusz, J.; Wilusz, C.J. The highways and byways of mRNA decay. Nat. Rev. Mol. Cell Biol. 2007, 8, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Floor, S.N.; Jones, B.N.; Hernandez, G.A.; Gross, J.D. A split active site couples cap recognition by Dcp2 to activation. Nat. Struct. Mol. Biol. 2010, 17, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.; Cho, H.; Liu, Z.; Bowler, M.W.; Piao, S.; Parker, R.; Kim, Y.K.; Song, H. Structural Basis of the PNRC2-Mediated Link between mRNA Surveillance and Decapping. Structure 2012, 20, 2025–2037. [Google Scholar] [CrossRef]
- Li, Y.; Song, M.; Kiledjian, M. Differential utilization of decapping enzymes in mammalian mRNA decay pathways. RNA 2011, 17, 419–428. [Google Scholar] [CrossRef]
- Li, Y.; Dai, J.; Song, M.; Fitzgerald-Bocarsly, P.; Kiledjian, M. Dcp2 Decapping Protein Modulates mRNA Stability of the Critical Interferon Regulatory Factor (IRF) IRF-7. Mol. Cell. Biol. 2012, 32, 1164–1172. [Google Scholar] [CrossRef]
- Boulias, K.; Toczydłowska-Socha, D.; Hawley, B.R.; Liberman, N.; Takashima, K.; Zaccara, S.; Guez, T.; Vasseur, J.-J.; Debart, F.; Aravind, L.; et al. Identification of the m6Am Methyltransferase PCIF1 Reveals the Location and Functions of m6Am in the Transcriptome. Mol. Cell 2019, 75, 631–643.e8. [Google Scholar] [CrossRef]
- Lu, G.; Zhang, J.; Li, Y.; Li, Z.; Zhang, N.; Xu, X.; Wang, T.; Guan, Z.; Gao, G.F.; Yan, J. hNUDT16: A universal decapping enzyme for small nucleolar RNA and cytoplasmic mRNA. Protein Cell 2011, 2, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Zytek, M.; Kowalska, J.; Lukaszewicz, M.; Wojtczak, B.A.; Zuberek, J.; Ferenc-Mrozek, A.; Darzynkiewicz, E.; Niedzwiecka, A.; Jemielity, J. Towards novel efficient and stable nuclear import signals: Synthesis and properties of trimethylguanosine cap analogs modified within the 5′,5′-triphosphate bridge. Org. Biomol. Chem. 2014, 12, 9184–9199. [Google Scholar] [CrossRef]
- Lukaszewicz, M.; Ferenc-Mrozek, A.; Bojarska, E.; Stelmach, J.; Stepinski, J.; Darzynkiewicz, E. Contribution of Nudt12 enzyme to differentially methylated dinucleotides of 5′RNA cap structure. Biochim. Biophys. Acta (BBA) Gen. Subj. 2023, 1867, 130400. [Google Scholar] [CrossRef]
- Chrabąszczewska, M.; Winiewska-Szajewska, M.; Ostrowska, N.; Bojarska, E.; Stępiński, J.; Mancewicz, Ł.; Łukaszewicz, M.; Trylska, J.; Taube, M.; Kozak, M.; et al. Insight into the Binding and Hydrolytic Preferences of hNudt16 Based on Nucleotide Diphosphate Substrates. Int. J. Mol. Sci. 2021, 22, 10929. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Grudzien-Nogalska, E.; Hamilton, K.; Jiao, X.; Yang, J.; Tong, L.; Kiledjian, M. Mammalian Nudix proteins cleave nucleotide metabolite caps on RNAs. Nucleic Acids Res. 2020, 48, 6788–6798. [Google Scholar] [CrossRef]
- Grudzien-Nogalska, E.; Wu, Y.; Jiao, X.; Cui, H.; Mateyak, M.K.; Hart, R.P.; Tong, L.; Kiledjian, M. Structural and mechanistic basis of mammalian Nudt12 RNA deNADding. Nat. Chem. Biol. 2019, 15, 575–582. [Google Scholar] [CrossRef]
- Wu, H.; Li, L.; Chen, K.-M.; Homolka, D.; Gos, P.; Fleury-Olela, F.; McCarthy, A.A.; Pillai, R.S. Decapping Enzyme NUDT12 Partners with BLMH for Cytoplasmic Surveillance of NAD-Capped RNAs. Cell Rep. 2019, 29, 4422–4434.e13. [Google Scholar] [CrossRef]
- Laudenbach, B.T.; Krey, K.; Emslander, Q.; Andersen, L.L.; Reim, A.; Scaturro, P.; Mundigl, S.; Dächert, C.; Manske, K.; Moser, M.; et al. NUDT2 initiates viral RNA degradation by removal of 5′-phosphates. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Rydzik, A.M.; Warminski, M.; Sikorski, P.J.; Baranowski, M.R.; Walczak, S.; Kowalska, J.; Zuberek, J.; Lukaszewicz, M.; Nowak, E.; Claridge, T.D.W.; et al. mRNA cap analogues substituted in the tetraphosphate chain with CX2: Identification of O-to-CCl2 as the first bridging modification that confers resistance to decapping without impairing translation. Nucleic Acids Res. 2017, 45, 8661–8675. [Google Scholar] [CrossRef] [PubMed]
- Page, B.D.G.; Valerie, N.C.K.; Wright, R.H.G.; Wallner, O.; Isaksson, R.; Carter, M.; Rudd, S.G.; Loseva, O.; Jemth, A.-S.; Almlöf, I.; et al. Targeted NUDT5 inhibitors block hormone signaling in breast cancer cells. Nat. Commun. 2018, 9, 250. [Google Scholar] [CrossRef] [PubMed]
- Mititelu, M.-B.; Hudeček, O.; Gozdek, A.; Benoni, R.; Nešuta, O.; Krasnodębski, S.; Kufel, J.; Cahová, H. Arabidopsis thaliana NudiXes have RNA-decapping activity. RSC Chem. Biol. 2023, 4, 223–228. [Google Scholar] [CrossRef]
- Kramer, S.; McLennan, A.G. The complex enzymology of mRNA decapping: Enzymes of four classes cleave pyrophosphate bonds. Wiley Interdiscip. Rev. RNA 2018, 10, e1511. [Google Scholar] [CrossRef]
- Deana, A.; Celesnik, H.; Belasco, J.G. The bacterial enzyme RppH triggers messenger RNA degradation by 5′ pyrophosphate removal. Nature 2008, 451, 355–358. [Google Scholar] [CrossRef]
- Cahová, H.; Winz, M.-L.; Höfer, K.; Nübel, G.; Jäschke, A. NAD captureSeq indicates NAD as a bacterial cap for a subset of regulatory RNAs. Nature 2015, 519, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Chang, J.H.; Kilic, T.; Tong, L.; Kiledjian, M. A Mammalian Pre-mRNA 5′ End Capping Quality Control Mechanism and an Unexpected Link of Capping to Pre-mRNA Processing. Mol. Cell 2013, 50, 104–115. [Google Scholar] [CrossRef]
- Picard-Jean, F.; Brand, C.; Tremblay-Létourneau, M.; Allaire, A.; Beaudoin, M.C.; Boudreault, S.; Duval, C.; Rainville-Sirois, J.; Robert, F.; Pelletier, J.; et al. 2′-O-methylation of the mRNA cap protects RNAs from decapping and degradation by DXO. PLoS ONE 2018, 13, e0193804. [Google Scholar] [CrossRef]
- Doamekpor, S.K.; Grudzien-Nogalska, E.; Mlynarska-Cieslak, A.; Kowalska, J.; Kiledjian, M.; Tong, L. DXO/Rai1 enzymes remove 5′-end FAD and dephospho-CoA caps on RNAs. Nucleic Acids Res. 2020, 48, 6136–6148. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-T.; Bercovich, N.; Loh, B.; Jonas, S.; Izaurralde, E. The activation of the decapping enzyme DCP2 by DCP1 occurs on the EDC4 scaffold and involves a conserved loop in DCP1. Nucleic Acids Res. 2014, 42, 5217–5233. [Google Scholar] [CrossRef]
- Borbolis, F.; Syntichaki, P. Biological implications of decapping: Beyond bulk mRNA decay. FEBS J. 2021, 289, 1457–1475. [Google Scholar] [CrossRef]
- Luo, Y.; Schofield, J.A.; Simon, M.D.; Slavoff, S.A. Global Profiling of Cellular Substrates of Human Dcp2. Biochemistry 2020, 59, 4176–4188. [Google Scholar] [CrossRef]
- Luo, Y.; Schofield, J.A.; Na, Z.; Hann, T.; Simon, M.D.; Slavoff, S.A. Discovery of cellular substrates of human RNA-decapping enzyme DCP2 using a stapled bicyclic peptide inhibitor. Cell Chem. Biol. 2020, 28, 463–474.e7. [Google Scholar] [CrossRef]
- Vvedenskaya, I.O.; Nickels, B.E. CapZyme-Seq: A 5′-RNA-Seq Method for Differential Detection and Quantitation of NAD-Capped and Uncapped 5′-Triphosphate RNA. STAR Protoc. 2020, 1, 100002. [Google Scholar] [CrossRef]
Figure 1.
mRNA 5′ end cap structures. Methylations of m7Guo and at 2′O position of riboses of the first two transcribed nucleotides, which result in Cap 1 and Cap 2 structures, are indicated. Inset the unmethylated cap variant, and m6Am adenosine containing cap variant (m7Gppp m6Am) are shown. Structures were drawn with ACD/ChemSketch, version 2022.1.2, Advanced Chemistry Development, Inc. (ACD/Labs).
Figure 1.
mRNA 5′ end cap structures. Methylations of m7Guo and at 2′O position of riboses of the first two transcribed nucleotides, which result in Cap 1 and Cap 2 structures, are indicated. Inset the unmethylated cap variant, and m6Am adenosine containing cap variant (m7Gppp m6Am) are shown. Structures were drawn with ACD/ChemSketch, version 2022.1.2, Advanced Chemistry Development, Inc. (ACD/Labs).
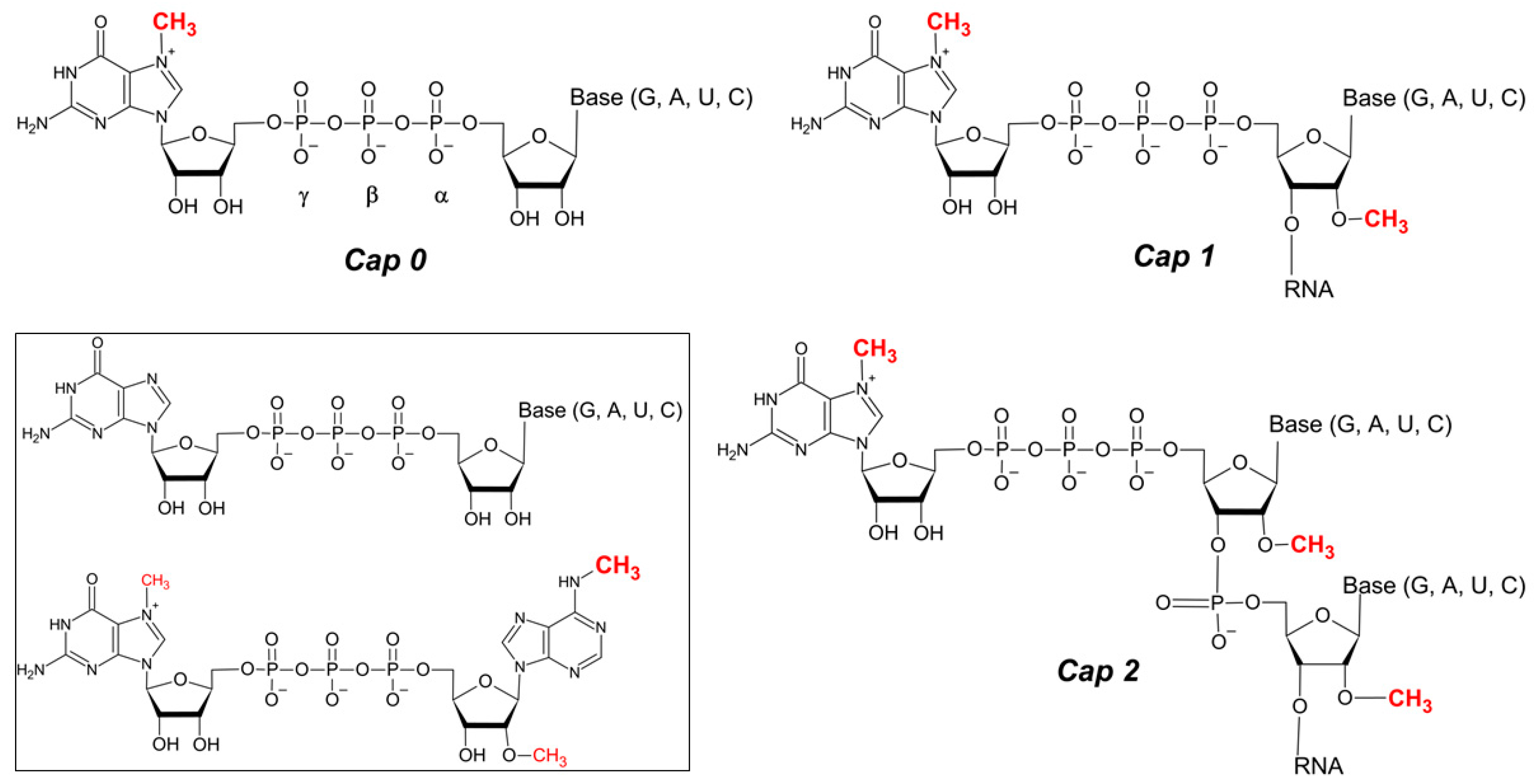
Figure 2.
Structures of non-canonical RNA caps.

Figure 3.
Anti-reverse cap analogue (ARCA).
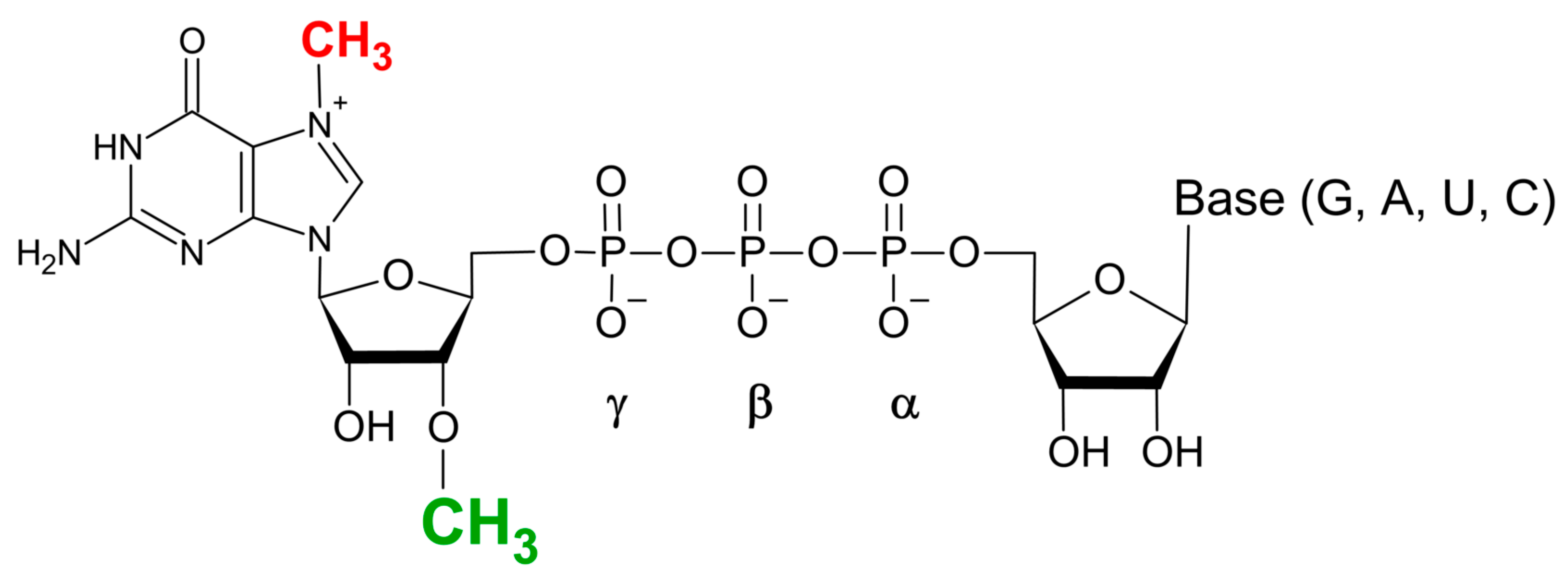
Figure 4.
Sites of chemical modifications of the canonical cap structures. (1) 2′OH and 3′OH of m7G, (2) the triphosphate bridge, (3–5) substitutions at the N7, N2, and C8 position of guanine, respectively, (6) modifications at N6 position of adenine adjacent to the m7G in the cap nucleotide structure.
Figure 4.
Sites of chemical modifications of the canonical cap structures. (1) 2′OH and 3′OH of m7G, (2) the triphosphate bridge, (3–5) substitutions at the N7, N2, and C8 position of guanine, respectively, (6) modifications at N6 position of adenine adjacent to the m7G in the cap nucleotide structure.

Figure 5.
Examples of the modifications introduced into triphosphate bridge of mRNA cap dinucleotide. (a) X1, X2 indicate sites of bridging oxygen substitution with -NH, -CH2, -CCl2, or -CF2, and Y1, Y2, and Y3 indicate sites of non-bridging oxygen substitution with -S, -Se, or-BH3; (b) representative phosphorothiolate modifications after O-to-S substitution within 5′-phosphoester bond in the triphosphate chain; (c) examples of triazole moiety introduced within cap oligophosphate linkage.
Figure 5.
Examples of the modifications introduced into triphosphate bridge of mRNA cap dinucleotide. (a) X1, X2 indicate sites of bridging oxygen substitution with -NH, -CH2, -CCl2, or -CF2, and Y1, Y2, and Y3 indicate sites of non-bridging oxygen substitution with -S, -Se, or-BH3; (b) representative phosphorothiolate modifications after O-to-S substitution within 5′-phosphoester bond in the triphosphate chain; (c) examples of triazole moiety introduced within cap oligophosphate linkage.

Figure 6.
Examples of the modifications introduced at 2′-OH or 3′-OH of m7Guanine. (a) propagyl group; (b) allyl group; (c) 2′-OH to Fluor substitution; (d) isopropylidene group; (e) locked nucleic acid (LNA) modification; (f) anthraniolyl (R: -H) and methylanthraniolyl (R: -CH3) fluorescent moieties; (g) 2′ amino group; (h) example of carbamate modification, possible label moieties include: trifluoromethyl diazirine (i), 6-nitroveratryl alcohol (NVA) (ii), fluorescein or biotin; (i) example of modification with azide group.
Figure 6.
Examples of the modifications introduced at 2′-OH or 3′-OH of m7Guanine. (a) propagyl group; (b) allyl group; (c) 2′-OH to Fluor substitution; (d) isopropylidene group; (e) locked nucleic acid (LNA) modification; (f) anthraniolyl (R: -H) and methylanthraniolyl (R: -CH3) fluorescent moieties; (g) 2′ amino group; (h) example of carbamate modification, possible label moieties include: trifluoromethyl diazirine (i), 6-nitroveratryl alcohol (NVA) (ii), fluorescein or biotin; (i) example of modification with azide group.

Figure 7.
Examples of the modifications of N7, N2, and C8 sites of m7Guanine. (A) N7 substitutions include: (a) benzyl, (b) 4-nitrobenzyl, (c) naphtalene, (d) 2,4-difluorobenzyl, (e) 4-chlorophenoxyethyl, (f) 7-(diethylamino)-4-methyl-coumarin, (g) benzophenone. (B) N2 substitutions include: (a) methyl, (b) benzyl, (c) 4-methoxybenzyl, (d) triazole, (e) 1-phenyl-triazole, (f) 2,6-dimethoxyphenyl triazole, (g) 3-phenyl-isoxazole, (h) 4-methyl-thiazole. (C) C8 substitutions include: (a) methyl, (b) 1-pyrene, (c) benzyl, (d) 4-dimethylaminophenyl, (e) 4-cyanophenyl.
Figure 7.
Examples of the modifications of N7, N2, and C8 sites of m7Guanine. (A) N7 substitutions include: (a) benzyl, (b) 4-nitrobenzyl, (c) naphtalene, (d) 2,4-difluorobenzyl, (e) 4-chlorophenoxyethyl, (f) 7-(diethylamino)-4-methyl-coumarin, (g) benzophenone. (B) N2 substitutions include: (a) methyl, (b) benzyl, (c) 4-methoxybenzyl, (d) triazole, (e) 1-phenyl-triazole, (f) 2,6-dimethoxyphenyl triazole, (g) 3-phenyl-isoxazole, (h) 4-methyl-thiazole. (C) C8 substitutions include: (a) methyl, (b) 1-pyrene, (c) benzyl, (d) 4-dimethylaminophenyl, (e) 4-cyanophenyl.
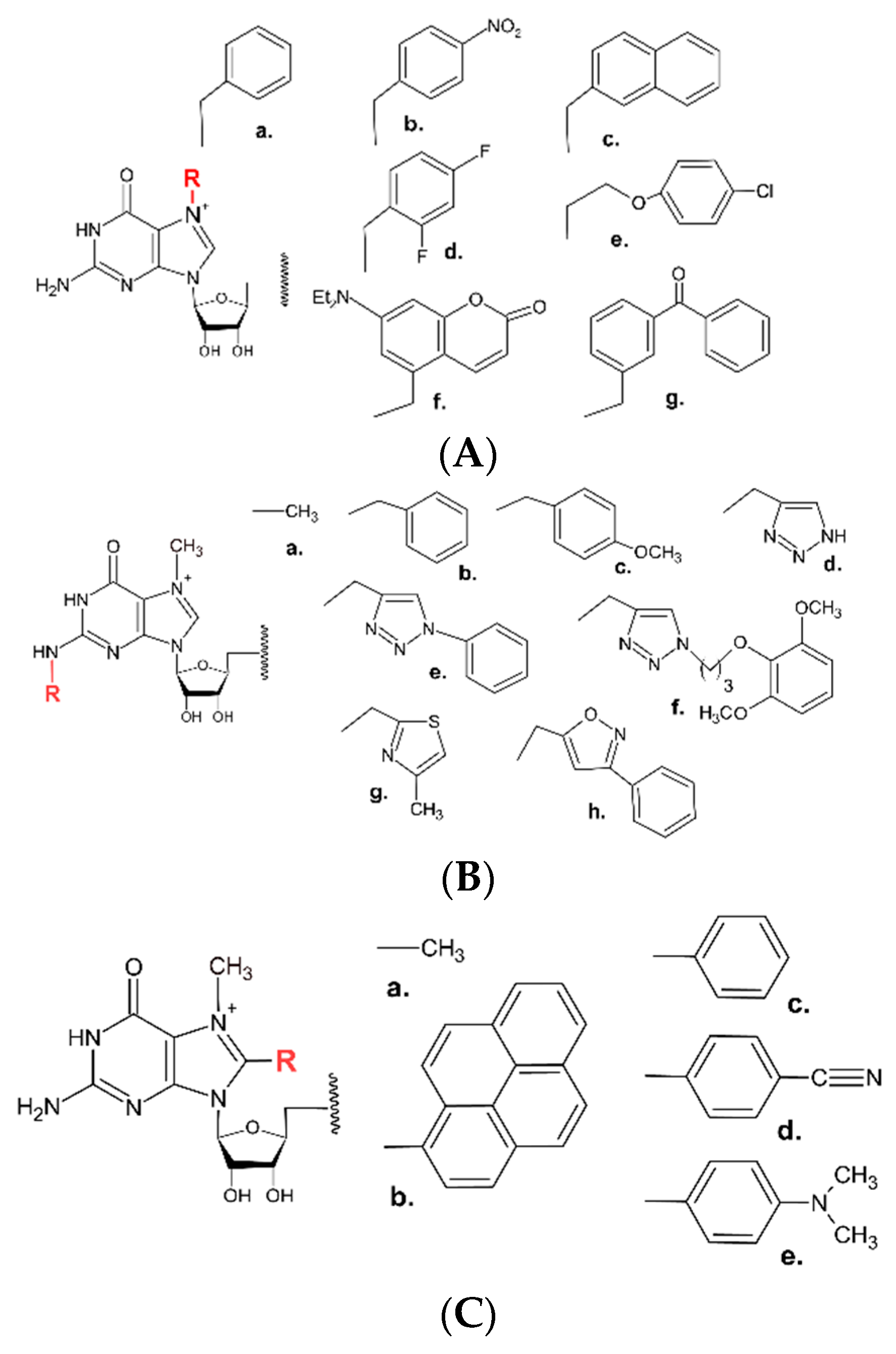
Table 3.
Activity of selected NUDIX proteins towards metabolite non-canonical RNA caps.
| NAD Cap | FAD Cap | dpCoA Cap | UDP-GlcNAc Cap | |
|---|---|---|---|---|
| Nudt16 | + | + | + | |
| Nudt12 | + | + | ||
| Nudt5 | + | |||
| Nudt2 | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lukaszewicz, M. Application of Mammalian Nudix Enzymes to Capped RNA Analysis. Pharmaceuticals 2024, 17, 1195. https://doi.org/10.3390/ph17091195
AMA Style
Lukaszewicz M. Application of Mammalian Nudix Enzymes to Capped RNA Analysis. Pharmaceuticals. 2024; 17(9):1195. https://doi.org/10.3390/ph17091195
Chicago/Turabian StyleLukaszewicz, Maciej. 2024. "Application of Mammalian Nudix Enzymes to Capped RNA Analysis" Pharmaceuticals 17, no. 9: 1195. https://doi.org/10.3390/ph17091195
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.
Article Metrics
Article metric data becomes available approximately 24 hours after publication online.