Cannabinoids and Reproduction: A Lasting and Intriguing History

Abstract
:Contents
- The Cannabinergic System: A Historical Overview
- 1.1.1
- Cannabinoid receptors
- 1.1.2
- Exogenous and endogenous ligands
- 1.1.3
- Endocannabinoids biosynthesis and degradation
- Interactions of the Cannabinergic System with Different Neuronal Networks in Reproductive Perspective
- The Cannabinergic System in Male Reproductive Tracts: From Spermatogenesis to Sperm Physiology
- 3.1.1
- Testis
- 3.1.2
- Excurrent duct system
- Effects of the Cannabinergic Sytem on Female Reproduction: From Ovary to Utero-placental Relationship
- Closing Remarks
1. The Cannabinergic System: A Historical Overview
1.1. Cannabinoid receptors
1.2. Exogenous and endogenous ligands

{kind=link}
| Discovery | Ref. |
|---|---|
| Δ9-THC | [10] |
| CB1 receptor | [15] |
| CB2 receptor | [16] |
| TRPV1 | [61] |
| GPR119 | [52] |
| GPR55 | [51] |
| AEA | [71] |
| 2-AG | [72] |
| SR141716A | [79] |
| SR144528 | [80] |
| AM251 and/or AM281 | [82] |
| AM630 | [83] |
| Classical cannabinoids |
|---|
| Δ-9-tetrahydrocannabinol, Δ9-THC [10, 17] |
| Δ-8-tetrahydrocannabinol, Δ8-THC [12, 17] |
| Cannabinol, CBN [12, 17] |
| Cannabidiol, CBD [12, 17] |
| Cannabicyclol, CBL [12, 17] |
| Cannabichromene, CBC [12, 17] |
| Cannabigerol, CBG [12, 17] |
| HU-210 [13, 17] |
| Desacetyl-l-nantradol [13, 17] |
| Non classical cannabinoids |
| CP55940 [70] |
| R-(+)-WIN55212 [70] |
| Endocannabinoids |
| AEA [71] |
| 2-AG [72] |
| noladin, 2-AGE [73] |
| virodhamine [74] |
| N-arachidonoyldopamine (NADA) [75] |
1.3. Endocannabinoids biosynthesis and degradation
| Member | Description | Function | Ref. |
|---|---|---|---|
| CB1 | Cannabinoid receptor type 1 | Bind CBs and eCBs | [15] |
| CB2 | Cannabinoid receptor type 2 | Bind CBs and eCBs | [16] |
| TRPV1 | Vanilloid receptor | Bind AEA | [61,62] |
| EMT | Endocannabinoids Membrane Transporter | Mediate eCBs diffusion across cellular membrane | [100] |
| NAPE-PLD | N-acylphosphatidylethanolamine phospholipase D | Biosynthesise AEA | [86] |
| FAAH | Fatty Acid Amide Hydrolase | Hydrolyse AEA and to a lesser content 2-AG | [107] |
| DAGL | Dyacilglycerol lipase | Biosynthesise | [93] |
| MAGL | Monoacylglycerol lipase | Hydrolyse 2-AG | [108] |
2. Interactions of the Cannabinergic System with Different Neuronal Networks from a Reproductive Perspective
3. The Cannabinergic System in the Male Reproductive Tract: From Spermatogenesis to Sperm Physiology
3.1. Testis
3.2. Excurrent duct system

4. Effects of the Cannabinergic System on Female Reproduction: From Ovary to Utero-placental Relationship
5. Closing Remarks
Abbreviations
| 2-AG | 2-arachydonoyl glycerol |
| 2-AGE | 2-arachidonoylglyceryl ether |
| AC | adenylyl cyclase |
| ACTH | adenocorticotropin hormone |
| AEA | anandamide |
| AM404 | N-(4-hydroxyphenyl)-arachidonylamide |
| APOA | anterior preoptic area |
| AR | acrosome reaction |
| ARC | arcuate nucleus CB1, cannabinoid receptor type-1 |
| CB2 | cannabinoid receptor type-2 |
| CBC | cannabichromene |
| CBD | cannabidiol |
| CBG | cannabigerol |
| CBL | cannabicyclol |
| CBN | cannabinol |
| CBRs | cannabinoid receptors |
| CBs | cannabinoids |
| cNOS | nitric oxide synthase |
| CNS | central nervous system |
| CORT | corticosterone |
| COXs | cyclooxygenases |
| CPS | capsaicin |
| CPZ | capsazepine |
| CRH | corticotropin-releasing hormone |
| CS | cannabinergic system |
| D | dopamine |
| 1,2-DAG | 1,2-diacylglycerol |
| DAGL | diacylglycerol lipase |
| dpp | days post partum |
| E2 | estradiol |
| eCBs | endocannabinoids |
| EMT | endocannabinoid membrane transporter |
| ERK | extracellular signal-regulated protein kinases |
| eSPT | elongating SPT |
| FAAH | fatty acid amide hydrolase |
| FABP | fatty acid binding proteins |
| FSH | follicle-stimulating hormone |
| GABA | γ-aminobutyric acid |
| GAPDS | glyceraldehyde 3-phosphate dehydrogenase S |
| GCs | glucocorticoids |
| GnIH | gonadotropin-inhibitory hormone |
| GnRH1 | gonadotropin-releasing hormone 1 |
| GPCR | GTP-binding protein-coupled receptors |
| GPR54 | G protein-coupled receptor 54 |
| GPR55 | G protein-coupled receptor 55 |
| GPR119 | G protein-coupled receptor 119 |
| GSK-3 | glycogen synthase kinase 3 |
| HAEAs | hydroanandamides |
| HPA | hypothalamic-pituitary-adrenal |
| HPG | hypothalamic-pituitary-gonad |
| HU-210 | 11- hydroxy-Δ8-THC-dimethylheptyl |
| ICM | inner cell mass |
| IFN-γ | interferon-γ |
| IL | interleukin |
| IVF | in vitro fertilization |
| KO | knock-out |
| LCs | Leydig cells |
| LH | luteinizing-hormone |
| LIF | leukaemia inhibitory factor |
| LOXs | lipoxygenases |
| LPA | lysophosphatidic acid |
| LPI | l-α-lysophosphatidylinositol |
| lyso-NArPE | lysophosphatidylethanolamine |
| MAGL | monoacylglycerol lipase |
| MAP | mitogen-activated protein |
| Met-AEA | methanandamide |
| NADA | N-arachidonoyldopamine |
| NAE | N-acylethanolamine |
| NAPE | N-acylphosphatidylethanolamine |
| NAPE-PLD | NAPE-specific phospholipase D |
| N-ArPE | N-arachidonylphosphatidylethanolamine |
| NAT | N-acyl-transacylase |
| NE | norephinephrine |
| NFAT | nuclear factor of activated T cells |
| NK | natural killer |
| NMDA | N-methyl-d-aspartate |
| NO | nitric oxide |
| OVX | ovariectomized |
| P | progesterone |
| p-AEA | phospho-AEA |
| PC | phosphatidylcholine |
| PE | phosphatidyl-ethanolamine |
| PGE2 | prostaglandin E2 |
| PG-EA | prostaglandin ethanolamine |
| PG-G | prostaglandin glyceryl ester |
| PI | phosphatidylinositol |
| PKA | protein kinase A |
| PKC | protein kinase C |
| PLA1 | phospholipase A1 |
| PLC | Phospholipase C |
| PM | plasma membrane |
| PPARα | peroxisome proliferator activator receptor alpha |
| PRL | prolactin |
| R-123 | rhodamine |
| rSPT | round spermatids |
| SCs | Sertoli cells |
| SPC | spermatocytes |
| SPG | spermatogonia |
| sPLA2 | secretory phospholipase 2 |
| SPT | spermatids |
| SPZ | spermatozoa |
| T | testosterone |
| Δ9-THC | Δ-9-tetrahydrocannabinol |
| Th | T-helper |
| THS | thyroid-stimulating hormone |
| TM | transmembrane |
| TRPV1 | transient potential vanilloid channel type-1 |
| URB597 | cyclohexyl carbamic acid 3’-carbamoyl-byphenyl-3-yl ester |
| ZP | zona pellucida |
Acknowledgements
References
- Read, B.E. Chinese medicinal plants. In Peking Natural History Bulletin; Mit Press: Beijing, China, 1936; p. 152. Columns IV and V. [Google Scholar]
- Johnson, M.R.; Melvin, L.S. Cannabinoids as Therapeutic Agents; Mechoulam, R., Ed.; CRC Press: New York, NY, USA, 1986; pp. 121–145. [Google Scholar]
- Bowker, J. The Oxford Dictionary of World Religions; Oxford University Press: Oxford, UK, 1997; p. 142. [Google Scholar]
- Booth, M. Cannabis: A History. Doubleday 2003, 339. [Google Scholar]
- Baker, D.; Pryce, G.; Giovannoni, G.; Thompson, A.J. The therapeutic potential of cannabis. Lancet Neurol. 2003, 2, 291–298. [Google Scholar]
- Robson, P. Therapeutic aspects of cannabis and cannabinoids. Br. J. Psychiatry. 2001, 178, 107–115. [Google Scholar]
- Solomon, P. Medical management of drug dependence. J. Amer. Medic. Assoc. 1968, 206, 1521–1526. [Google Scholar]
- Carter, G.T.; Weydt, P.; Kyashna-Tocha, M.; Abrams, D.I. Medicinal cannabis: rational guidelines for dosing. IDrugs 2004, 7, 464–470. [Google Scholar]
- Wilkinson, D.J. Myths and mandrakes. J. R. Soc. Med. 2003, 96, 255–256. [Google Scholar]
- Mechoulam, R.; Gaoni, Y.J. A total synthesis of delta 1 tetrahycrocannabinol, the active constituents of hashish. Am. Chem. Soc. 1965, 87, 3273–3275. [Google Scholar]
- Elsohly, M.A.; Slade, D. Chemical constituents of marijuana: the complex mixture of natural cannabinoids. Life Sci. 2005, 78, 539–548. [Google Scholar]
- Smith, F.L.; Fujimori, K.; Lowe, J.; Welch, S.P. Characterization of delta9-tetrahydrocannabinol and anandamide antinociception in nonarthritic and arthritic rats. Pharmacol. Biochem. Behav. 1998, 60, 183–191. [Google Scholar]
- Melvin, L.S.; Johnson, M.R. Structure-activity relationships of tricyclic and nonclassical bicyclic cannabinoids. NIDA Res. Monogr. 1987, 79, 31–47. [Google Scholar]
- Devane, W.A.; Dysarz, F.A., 3rd; Johnson, M.R.; Melvin, L.S.; Howlett, A.C. Determination and characterization of a cannabinoid receptor in rat brain. Mol. Pharmacol. 1988, 34, 605–613. [Google Scholar] [PubMed]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar]
- Howlett, A.C. The cannabinoid receptors. Prost. Other Lipid Mediat. 2002, 68, 619–631. [Google Scholar]
- Childers, S.R. Activation of G-proteins in brain by endogenous and exogenous cannabinoids. AAPS J. 2006, 8, E112–E117. [Google Scholar]
- Howlett, A.C.; Blume, L.C.; Dalton, G.D. CB(1) cannabinoid receptors and their associated proteins. Curr. Med. Chem. 2010, 17, 1382–1393. [Google Scholar]
- Howlett, A.C. The CB1 cannabinoid receptor in the brain. Neurobiol Dis 1998, 5, 405–416. [Google Scholar]
- Glass, M.; Felder, C.C. Concurrent stimulation of cannabinoid CB1 and dopamine D2 receptors augments cAMP accumulation in striatal neurons: evidence for a Gs linkage to the CB1 receptor. J. Neurosci. 1997, 17, 5327–5333. [Google Scholar]
- Twitchell, W.; Brown, S.; Mackie, K. Cannabinoids inhibit N- and P/Q-type calcium channels in cultured rat hippocampal neurons. J. Neuriphysiol. 1997, 78, 43–50. [Google Scholar]
- Caulfield, M.P.; Brown, D.A. Cannabinoid receptor agonists inhibit Ca current in NG108-15 neuroblastoma cells via a pertussis toxin-sensitive mechanism. Br. J. Pharmacol. 1992, 106, 231–232. [Google Scholar]
- Mackie, K.; Hille, B. Cannabinoids inhibit N-type calcium channels in neuroblastoma-glioma cells. Proc. Natl. Acad. Sci. USA 1992, 89, 3825–3829. [Google Scholar]
- Oz, M. Receptor-independent effects of endocannabinoids on ion channels. Curr. Pharm. Des. 2006, 12, 227–239. [Google Scholar]
- Lauckner, J.E.; Hille, B.; Mackie, K. The cannabinoid agonist WIN55,212-2 increases intracellular calcium via CB1 receptor coupling to Gq/11 G proteins. Proc. Natl. Acad. Sci. USA 2005, 102, 19144–19149. [Google Scholar]
- Netzeband, J.G.; Conroy, S.M.; Parson, K.L.; Groul, D.L. Cannabinoids enhance NMDA-elicited calcium signals in cerebellar granule neurons in culture. J. Neurosci. 1999, 19, 8765–8777. [Google Scholar]
- Fimiani, C.; Mattocks, D.; Cavani, F.; Salzet, M.; Deutsch, D.G.; Pryor, S.; Bilfinger, T.V.; Stefano, G.B. Morphine and anandamide stimulate intracellular calcium transients in human arterial endothelial cells: coupling to nitric oxide release. Cell Signal 1999, 3, 189–193. [Google Scholar]
- Bouaboula, M.; Poinot-Chazel, C.; Bourrie, B.; Canat, X.; Calandra, B.; Rinaldi-Carmona, M.; Le Fur, G.; Casellas, P. Activation of mitogen-activated protein kinases by stimulation of the central cannabinoid receptor CB1. Biochem. J. 1995, 312, 637–641. [Google Scholar]
- Derkinderen, P.; Valjent, E.; Toutant, M.; Corvol, J.C.; Enslen, H.; Ledent, C.; Trzaskos, J.; Caboche, J.; Girault, J.A. Regulation of extracellular signal-regulated kinase by cannabinoids in hippocampus. J. Neurosci. 2003, 23, 2371–2382. [Google Scholar]
- Felder, C.C.; Joyce, K.E.; Briley, E.M.; Mansouri, J.; Mackie, K.; Blond, O.; Lay, Y.; Ma, A.L.; Mitchell, R.L. Comparison of the pharmacology and signal transduction of the human cannabinoid CB1 and CB2 receptors. Mol. Pharmacol. 1995, 48, 443–450. [Google Scholar]
- Diaz-Laviada, I.; Ruiz-Llorente, L. Signal transduction activated by cannabinoid receptors. Mini Rev. Med. Chem. 2005, 5, 619–630. [Google Scholar]
- Wenger, T.; Fernández-Ruiz, J.J.; Ramos, J.A. Immunocytochemical demonstration of CB1 cannabinoid receptors in the anterior lobe of the pituitary gland. J. Neuroendocrinol. 1999, 11, 873–878. [Google Scholar]
- Kaminski, N.E.; Abood, M.E.; Kessler, F.K.; Martin, B.R.; Schatz, A.R. Identification of a functionally relevant cannabinoid receptor on mouse spleen cells that is involved in cannabinoid-mediated immune modulation. Mol. Pharmacol. 1992, 42, 736–742. [Google Scholar]
- Bouaboula, M.; Rinaldi, M.; Carayon, P.; Carillon, C.; Delpech, B.; Shire, D.; Le Fur, G.; Casellas, P. Cannabinoid-receptor expression in human leukocytes. Eur. J. Biochem. 1993, 214, 173–180. [Google Scholar]
- Taylor, A.H.; Ang, C.; Bell, S.C.; Konje, J.C. The role of the endocannabinoid system in gametogenesis, implantation and early pregnancy. Hum. Reprod. Update 2007, 13, 501–513. [Google Scholar]
- Kulkarni-Narla, A.; Brown, D.R. Localization of CB1-cannabinoid receptor immunoreactivity in the porcine enteric nervous system. Cell Tissue Res. 2000, 302, 73–80. [Google Scholar]
- Ishac, E.J.; Jiang, L.; Lake, K.D.; Varga, K.; Abood, M.E.; Kunos, G. Inhibition of exocytotic noradrenaline release by presynaptic cannabinoid CB1 receptors on peripheral sympathetic nerves. Br. J. Pharmacol. 1996, 118, 2023–2028. [Google Scholar]
- Wenger, T.; Ledent, C.; Csernus, V.; Gerendai, I. The central cannabinoid receptor inactivation suppresses endocrine reproductive functions. Biochem. Biophys. Res. Commun. 2001, 284, 363–368. [Google Scholar]
- Rice, W.; Shannon, J.M.; Burton, F.; Fiedeldey, D. Expression of a brain-type cannabinoid receptor (CB1) in alveolar Type II cells in the lung: regulation by hydrocortisone. Eur. J. Pharmacol. 1997, 327, 227–232. [Google Scholar]
- Farquhar-Smith, W.P.; Jaggar, S.I.; Rice, A.S. Attenuation of nerve growth factor-induced visceral hyperalgesia via cannabinoid CB(1) and CB(2)-like receptors. Pain 2002, 97, 11–21. [Google Scholar]
- Niederhoffer, N.; Hansen, H.H.; Fernandez-Ruiz, J.J.; Szabo, B. Effects of cannabinoids on adrenaline release from adrenal medullary cells. Br. J. Pharmacol. 2001, 134, 1319–1327. [Google Scholar]
- Cota, D.; Marsicano, G.; Tschöp, M.; Grübler, Y.; Flachskamm, C.; Schubert, M.; Auer, D.; Yassouridis, A.; Thöne-Reineke, C.; Ortmann, S.; Tomassoni, F.; Cervino, C.; Nisoli, E.; Linthorst, A.C.; Pasquali, R.; Lutz, B.; Stalla, G.K.; Pagotto, U. The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J. Clin. Invest. 2003, 112, 423–431. [Google Scholar]
- Osei-Hyiaman, D.; De Petrillo, M.; Pacher, P.; Liu, J.; Radaeva, S.; Bátkai, S.; Harvey-White, J.; Mackie, K.; Offertáler, L.; Wang, L.; Kunos, G. Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity. J. Clin. Invest. 2005, 115, 1298–1305. [Google Scholar] [PubMed]
- Galiegue, S.; Mary, S.; Marchand, J; Dussossoy, D.; Carriere, D.; Carayon, P.; Bouaboula, M.; Shire, D.; Le Fur, G.; Casellas, P. Expression of central and peripheral cannabinoid receptors in human immune tissues and leukocyte subpopulations. Eur. J. Biochem. 1995, 232, 54–61. [Google Scholar]
- Pettit, D.A.; Anders, D.L.; Harrison, M.P.; Cabral, G.A. Cannabinoid receptor expression in immune cells. Adv. Exp. Med. Biol. 1996, 402, 119–129. [Google Scholar]
- Facci, L.; Dal Toso, R.; Romanello, S.; Buriani, A.; Skaper, S.D.; Leon, A. Mast cells express a peripheral cannabinoid receptor with differential sensitivity to anandamide and palmitoylethanolamide. Proc. Natl. Acad. Sci. USA 1995, 92, 3376–3380. [Google Scholar]
- Walter, L.; Franklin, A.; Witting, A.; Wade, C.; Xie, Y.; Kunos, G.; Mackie, K.; Stella, N. Nonpsychotropic cannabinoid receptors regulate microglial cell migration. J. Neurosci. 2003, 23, 1398–1405. [Google Scholar]
- Van Sickle, M.D.; Duncan, M.; Kingsley, P.J.; Mouihate, A.; Urbani, P.; Mackie, K.; Stella, N.; Makriyannis, A.; Piomelli, D.; Davison, J.S.; Marnett, L.J.; Di Marzo, V.; Pittman, Q.J.; Patel, K.D.; Sharkey, K.A. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 2005, 310, 329–332. [Google Scholar] [PubMed]
- Onaivi, E.S. Neuropsychobiological evidence for the functional presence and expression of cannabinoid CB2 receptors in the brain. Neuropsychobiology 2006, 54, 231–246. [Google Scholar]
- Begg, M.; Pacher, P.; Batkai, S.; Osei-Hyiaman, D.; Offertaler, L.; Mo, F.M.; Liu, J.; Kunos, G. Evidence for novel cannabinoid receptors. Pharmacol. Ther. 2005, 106, 133–145. [Google Scholar]
- Fredriksson, R.; Hoglund, P.J.; Gloriam, D.E.; Lagerstrom, M.C.; Schioth, H.B. Seven evolutionarily conserved human rhodopsin G protein-coupled receptors lacking close relatives. FEBS Lett. 2003, 554, 381–388. [Google Scholar]
- Lauckner, J.E.; Bensen, J.B.; Chen, H.Y.; Lu, H.C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar]
- Henstridge, C.M.; Balenga, N.A.B.; Ford, L.A.; Ross, R.A.; Waldhoer, M.; Irving, A.J. The GPR55 ligand L-α-lysophosphatidylinositol promotes RhoA-dependent Ca2+ signaling and NFAT activation. FASEB J. 2009, 23, 183–193. [Google Scholar]
- Mackie, K.; Stella, N. Cannabinoid receptors and endocannabinoids: evidence for new players. Am. Assoc. Pharm. Sci. J. 2006, 8, E298–E306. [Google Scholar]
- Kapur, A.; Zhao, P.; Sharir, H.; Bai, Y.; Caron, M.G.; Barak, L.S.; Abood, M.E. Atypical responsiveness of the orphan receptor GPR55 to cannabinoid ligands. J. Biol. Chem. 2009, 284, 29817–29827. [Google Scholar]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar]
- Daly, C.J.; Ross, R.A.; Whyte, J.; Henstridge, C.M.; Irving, A.J.; McGrath, J.C. Fluorescent ligand binding reveals heterogeneous distribution of adrenoceptors and 'cannabinoid-like' receptors in small arteries. Br. J. Pharmacol. 2010, 159, 787–796. [Google Scholar]
- Sakamoto, Y.; Inoue, H.; Kawakami, S.; Miyawaki, K.; Miyamoto, T.; Mizuta, K.; Itakura, M. Expression and distribution of Gpr119 in the pancreatic islets of mice and rats: predominant localization in pancreatic polypeptide-secreting PP-cells. Biochem. Biophys. Res. Commun. 2006, 351, 474–480. [Google Scholar]
- Overton, H.A.; Babbs, A.J.; Doel, S.M.; Fyfe, M.C.; Gardner, L.S.; Griffin, G.; Jackson, H.C.; Procter, M.J.; Rasamison, C.M.; Tang-Christensen, M.; Widdowson, P.S.; Williams, G.M.; Reynet, C. Deorphanization of a G protein-coupled receptor for oleoylethanolamide and its use in the discovery of small-molecule hypophagic agents. Cell Metab. 2006, 3, 167–175. [Google Scholar]
- Kedei, N.; Szabo, T.; Lile, J.D.; Treanor, J.J.; Olah, Z.; Iadarola, M.J.; Blumberg, P.M. Analysis of the native quaternary structure of vanilloid receptor 1. J. Biol. Chem. 2001, 276, 28613–28619. [Google Scholar]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar]
- Gunthorpe, M.J; Harries, M.H.; Prinjha, R.K.; Davis, J.B.; Randall, A. Voltage- and time-dependent properties of the recombinant rat vanilloid receptor (rVR1). J. Physiol. 2000, 525, 747–759. [Google Scholar]
- Bhave, G.; Hu, H.J.; Glauner, K.S.; Zhu, W.; Wang, H.; Brasier, D.J.; Oxford, G.S.; Gereau, R.W. Protein kinase C phosphorylation sensitizes but does not activate the capsaicin receptor transient receptor potential vanilloid 1 (TRPV1). Proc. Natl. Acad. Sci. USA 2003, 100, 12480–12485. [Google Scholar]
- De Petrocellis, L.; Bisogno, T.; Maccarrone, M.; Davis, J.B.; Finazzi-Agrò, A.; Di Marzo, V. The activity of anandamide at vanilloid VR1 receptors requie facilitated tran sport across the cell membrane and is limited by intracellular metabolism. J. Biol. Chem. 2001, 276, 12856–12863. [Google Scholar]
- Yamamoto, Y.; Taniguchi, K. Immunolocalization of VR1 and VRL1 in rat larynx. Auton. Neurosci. 2005, 117, 62–65. [Google Scholar]
- Liu, L.; Chen, L.; Liedtke, W.; Simon, S.A. Changes in osmolality sensitize the response to capsaicin in trigeminal sensory neurons. J. Neurophysiol. 2007, 97, 2001–2015. [Google Scholar]
- Maccarrone, M.; Barboni, B.; Paradisi, A.; Bernabo, N.; Gasperi, V.; Pistilli, M.G.; Fezza, F.; Lucidi, P.; Mattioli, M. Characterization of the endocannabinoid system in boar spermatozoa and implications for sperm capacitation and acrosome reaction. J. Cell Sci. 2005, 118, 4393–4404. [Google Scholar]
- Grimaldi, P.; Orlando, P.; Di Siena, S.; Lolicato, F.; Petrosino, S.; Bisogno, T.; Geremia, R.; De Petrocellis, L.; Di Marzo, V. The endocannabinoid system and pivotal role of the CB2 receptor in mouse spermatogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 11131–11136. [Google Scholar]
- Pertwee, R.G. Pharmacology of cannabinoid receptor ligands. Curr. Med. Chem. 1999, 6, 635–664. [Google Scholar]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [PubMed]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [PubMed]
- Sugiura, T.; Kodaka, T.; Nakane, S.; Miyashita, T.; Kondo, S.; Suhara, Y.; Takayama, H.; Waku, K.; Seki, C.; Baba, N.; Ishima, Y. Evidence that the cannabinoid CB1 receptor is a 2-arachidonoylglycerol receptor. Structure-activity relationship of 2-arachidonoylglycerol, ether-linked analogues, and related compounds. J. Biol. Chem. 1999, 274, 2794–2801. [Google Scholar] [PubMed]
- Porter, A.C.; Sauer, J.M.; Knierman, M.D.; Becker, G.W.; Berna, M.J.; Bao, J.; Nomikos, G.G.; Carter, P.; Bymaster, F.P.; Leese, A.B.; Felder, C.C. Characterization of a novel endocannabinoid, virodhamine, with antagonist activity at the CB1 receptor. J. Pharmacol. Exp. Ther. 2002, 301, 1020–1024. [Google Scholar]
- Ralevic, V. Cannabinoid modulation of peripheral autonomic and sensory neurotransmission. Eur. J. Pharmacol. 2003, 472, 1–21. [Google Scholar]
- Di Marzo, V.; De Petrocellis, L.; Bisogno, T.; Melck, D. Metabolism of anandamide and 2-arachidonoylglycerol: an historical overview and some recent developments. Lipids 1999, 34 Suppl, S319–S325. [Google Scholar]
- Ross, R.A.; Gibson, T.M.; Brockie, H.C.; Leslie, M.; Pashmi, G.; Craib, S.J.; Di Marzo, V.; Pertwee, R.G. Structure-activity relationship for the endogenous cannabinoid, anandamide, and certain of its analogues at vanilloid receptors in transfected cells and vas deferens. Br. J. Pharmacol. 2001, 132, 631–640. [Google Scholar] [PubMed]
- Lo Verme, J.; Fu, J.; Astarita, G.; La Rana, G.; Russo, R.; Calignano, A.; Piomelli, D. The nuclear receptor peroxisome proliferator-activated receptor-alpha mediates the anti-inflammatory actions of palmitoylethanolamide. Mol. Pharmacol. 2005, 67, 15–19. [Google Scholar] [PubMed]
- Rinaldi-Carmona, M.; Barth, F.; Heaulme, M.; Shire, D.; Calandra, B.; Congy, C.; Martinez, S.; Maruani, J.; Neliat, G.; Caput, D. SR141716A, a potent and selective antagonist of the brain cannabinoid receptor. FEBS Lett. 1994, 350, 240–244. [Google Scholar]
- Rinaldi-Carmona, M.; Barth, F.; Millan, J.; Derocq, J.M.; Casellas, P.; Congy, C.; Oustric, D.; Sarran, M.; Bouaboula, M.; Calandra, B.; Portier, M.; Shire, D.; Breliere, J.C.; Le Fur, G.L. SR 144528, the first potent and selective antagonist of the CB2 cannabinoid receptor. J. Pharmacol. Exp. Ther. 1998, 284, 644–650. [Google Scholar]
- MacLennan, S.J.; Reynen, P.H.; Kwan, J.; Bonhaus, D.W. Evidence for inverse agonism of SR141716A at human recombinant cannabinoid CB1 and CB2 receptors. Br. J. Pharmacol. 1998, 124, 619–622. [Google Scholar]
- Al-Hayani, A.; Davies, S.N. Cannabinoid receptor mediated inhibition of excitatory synaptic transmission in the rat hippocampal slice is developmentally regulated. Br. J. Pharmacol. 2000, 131, 663–665. [Google Scholar]
- White, R.; Ho, W.S.; Bottrill, F.E.; Ford, W.R.; Hiley, C.R. Mechanisms of anandamide-induced vasorelaxation in rat isolated coronary arteries. Br. J. Pharmacol. 2001, 134, 921–929. [Google Scholar]
- Di Marzo, V.; Fontana, A.; Cadas, H.; Schinelli, S.; Cimino, G.; Schwartz, J.C.; Piomelli, D. Formation and inactivation of endogenous cannabinoid anandamide in central neurons. Nature 1994, 372, 686–691. [Google Scholar]
- Cadas, H.; di Tomaso, E.; Piomelli, D. Occurrence and biosynthesis of endogenous cannabinoid precursor, N-arachidonoyl phosphatidylethanolamine, in rat brain. J. Neurosci. 1997, 17, 1226–1242. [Google Scholar] [PubMed]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Tonegawa, T.; Nakane, S.; Yamashita, A.; Waku, K. Enzymatic synthesis of anandamide, an endogenous cannabinoid receptor ligand, through N-acylphosphatidylethanolamine pathway in testis: involvement of Ca(2+)-dependent transacylase and phosphodiesterase activities. Biochem. Biophys. Res. Commun. 1996, 218, 113–117. [Google Scholar] [PubMed]
- Schmid, P.C.; Reddy, P.V.; Natarajan, V.; Schmid, H.H. Metabolism of N-acylethanolamine phospholipids by a mammalian phosphodiesterase of the phospholipase D type. J. Bio.l Chem. 1983, 258, 9302–9306. [Google Scholar]
- Okamoto, Y.; Morishita, J.; Tsuboi, K.; Tonai, T.; Ueda, N. Molecular characterization of a phospholipase D generating anandamide and its congeners. J. Biol. Chem. 2004, 279, 5298–5305. [Google Scholar]
- Leung, D.; Saghatelian, A.; Simon, G.M.; Cravatt, B.F. Inactivation of N-acyl phosphatidylethanolamine phospholipase D reveals multiple mechanisms for the biosynthesis of - endocannabinoids. Biochemisty 2006, 45, 4720–4726. [Google Scholar]
- Sun, Y.X.; Tsuboi, K.; Okamoto, Y.; Tonai, T.; Murakami, M.; Kudo, I.; Ueda, N. Biosynthesis of anandamide and N-palmitoylethanolamine by sequential actions of phospholipase A2 and lysophospholipase D. Biochem. J. 2004, 380, 749–756. [Google Scholar]
- Liu, J.; Wang, L.; Harvey-White, J.; Osei-Hyiaman, D.; Razdan, R.; Gong, Q.; Chan, A.C.; Zhou, Z.; Huang, B.X.; Kim, H.Y.; Kunos, G. A biosynthetic pathway for anandamide. Proc. Natl. Acad. Sci. USA 2006, 103, 13345–13350. [Google Scholar]
- Simon, G.M.; Cravatt, B.F. Endocannabinoid biosynthesis proceeding through glycerophospho-N-acyl ethanolamine and a role for alpha/beta-hydrolase 4 in this pathway. J. Biol. Chem. 2006, 281, 26465–26472. [Google Scholar]
- Bisogno, T.; Melck, D.; De Petrocellis, L.; Di Marzo, V. Phosphatidic acid as the biosynthetic precursor of the endocannabinoid 2-arachidonoylglycerol in intact mouse neuroblastoma cells stimulated with ionomycin. J. Neurochem. 1999, 72, 2113–2119. [Google Scholar]
- Stella, N.; Schweitzer, P.; Piomelli, D. A second endogenous cannabinoid that modulates long-term potentiation. Nature 1997, 388, 773–778. [Google Scholar]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar]
- Bisogno, T.; Howell, F.; Williams, G.; Minassi, A.; Cascio, M.G.; Ligresti, A.; Matias, I.; Schiano-Moriello, A.; Paul, P.; Williams, E.J.; et al. Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain. J. Cell. Biol. 2003, 163, 463–468. [Google Scholar]
- Beltramo, M.; Stella, N.; Calignano, A.; Lin, S.Y.; Makriyannis, A.; Piomelli, D. Functional role of high-affinity anandamide transport, as revealed by selective inhibition. Science 1997, 277, 1094–1097. [Google Scholar]
- Bojesen, I.N.; Hansen, H.S. Membrane transport of anandamide through resealed human red blood cell membranes. Lipid. Res. 2005, 46, 1652–1659. [Google Scholar]
- Glaser, S.T.; Abumrad, N.A.; Fatade, F.; Kaczocha, M.; Studholme, K.M.; Deutsch, D.G. Evidence against the presence of an anandamide transporter. Proc. Natl. Acad. Sci. U S A. 2003, 100, 4269–4274. [Google Scholar]
- Beltramo, M.; Piomelli, D. Carrier-mediated transport and enzymatic hydrolysis of the endogenous cannabinoid 2-arachidonylglycerol. Neuroreport 2000, 27, 1231–1235. [Google Scholar]
- Ligresti, A.; Morera, E.; Van Der Stelt, M.; Monory, K.; Lutz, B.; Ortar, G.; Di Marzo, V. Further evidence for the existence of a specific process for the membrane transport of anandamide. Biochem. J. 2004, 380, 265–272. [Google Scholar]
- Ligresti, A.; De Petrocellis, L.; Hernán Pérez de la Ossa, D.; Aberturas, R.; Cristino, L.; Moriello, A.S.; Finizio, A.; Gil, M.E.; Torres, A.I.; Molpeceres, J.; Di Marzo, V. Exploiting nanotechnologies and TRPV1 channels to investigate the putative anandamide membrane transporter. PLoS One 2010, 5, e10239. [Google Scholar]
- Rodgers, W.; Smith, K. Properties of glycolipid-enriched membrane rafts in antigen presentation. Crit. Rev. Immunol. 2005, 25, 19–30. [Google Scholar]
- Bari, M.; Oddi, S.; De Simone, C.; Spagnolo, P.; Gasperi, V.; Battista, N.; Centonze, D.; Maccarrone, M. Type-1 cannabinoid receptors colocalize with caveolin-1 in neuronal cells. Neuropharmacology 2008, 54, 45–50. [Google Scholar]
- Pike, L.J. Growth factor receptors, lipid rafts and caveolae: an evolving story. Biochem. Biophys. Acta 2005, 1746, 260–273. [Google Scholar]
- Kaczocha, M.; Glaser, S.T.; Deutsch, D.G. Identification of intracellular carriers for the ndocannabinoid anandamide. Proc. Natl. Acad. Sci. USA 2009, 106, 6375–6380. [Google Scholar]
- Schug, T.T.; Berry, D.C.; Shaw, N.S.; Travis, S.N.; Noy, N. Opposing effects of retinoic acid on cell growth result from alternate activation of two different nuclear receptors. Cell 2007, 129, 723–733. [Google Scholar]
- Cravatt, B.F.; Giang, D.K.; Mayfield, S.P.; Boger, D.L.; Lerner, R.A.; Gilula, N.B. Molecular characterization of an enzyme that degrades neuromodulatory fatty-acid amides. Nature 1996, 384, 83–87. [Google Scholar]
- Dinh, T.P.; Freund, T.F.; Piomelli, D. A role for monoglyceride lipase in 2-arachidonoylglycerol inactivation. Chem. Phys. Lipids. 2002, 121, 149–158. [Google Scholar]
- Kozak, K.R; Marnett, L.J. Oxidative metabolism of endocannabinoids. Prost. Leukot. Essent. Fatty Acids 2002, 66, 211–220. [Google Scholar] [CrossRef]
- van der Stelt, M.; van Kuik, J.A.; Bari, M.; van Zadelhoff, G.; Leeflang, B.R.; Veldink, G.A.; Finazzi-Agrò, A.; Vliegenthart, J.F.; Maccarrone, M. Oxygenated metabolites of anandamide and 2-arachidonoylglycerol: conformational analysis and interaction with cannabinoid receptors, membrane transporter, and fatty acid amide hydrolase. J. Med. Chem. 2002, 45, 3709–3720. [Google Scholar]
- Craib, S.J.; Ellington, H.C.; Pertwee, R.G.; Ross, R.A. A possible role of lipoxygenase in the activation of vanilloid receptors by anandamide in the guinea-pig bronchus. Br. J. Pharmacol. 2001, 134, 30–37. [Google Scholar]
- Kozak, K.R.; Gupta, R.A.; Moody, J.S.; Ji, C.; Boeglin, W.E.; DuBois, R.N.; Brash, A.R.; Marnett, L.J. 15-Lipoxygenase metabolism of 2-arachidonylglycerol. Generation of a peroxisome proliferator-activated receptor alpha agonist. J. Biol. Chem. 2002, 277, 23278–23286. [Google Scholar] [PubMed]
- Weber, A.; Ni, J.; Ling, K.H.; Acheampong, A.; Tang-Liu, D.D.; Burk, R.; Cravatt, B.F.; Woodward, D. Formation of prostamides from anandamide in FAAH knockout mice analyzed by HPLC with tandem mass spectrometry. J. Lipid. Res. 2004, 45, 757–763. [Google Scholar]
- Ross, R.A.; Craib, S.J.; Stevenson, L.A.; Pertwee, R.G.; Henderson, A.; Toole, J.; Ellington, H.C. Pharmacological characterization of the anandamide cyclooxygenase metabolite: prostaglandin E2 ethanolamide. J. Pharmacol. Exp. Ther. 2002, 301, 900–907. [Google Scholar]
- Cravatt, B.F.; Lichtman, A.H. The enzymatic inactivation of the fatty acid amide class of signaling lipids. Chem. Phys. Lipids. 2002, 121, 135–148. [Google Scholar]
- McKinney, M.K.; Cravatt, B.F. Structure and function of fatty acid amide hydrolase. Annu. Rev. Biochem. 2005, 74, 411–432. [Google Scholar]
- Wei, B.Q.; Mikkelsen, T.S.; McKinney, M.K.; Lander, E.S.; Cravatt, B.F. A second fatty acid amide hydrolase with variable distribution among placental mammals. J. Biol. Chem. 2006, 281, 36569–36578. [Google Scholar]
- Deutsch, D.G.; Chin, S.A. Enzymatic synthesis and degradation of anandamide, a cannabinoid receptor agonist. Biochem. Pharmacol. 1993, 46, 791–796. [Google Scholar]
- Mor, M.; Rivara, S.; Lodola, A.; Plazzi, P.V.; Tarzia, G.; Duranti, A.; Tontini, A.; Piersanti, G.; Kathuria, S.; Piomelli, D. Cyclohexylcarbamic acid 3'- or 4'-substituted biphenyl-3-yl esters as fatty acid amide hydrolase inhibitors: synthesis, quantitative structure-activity relationships, and molecular modeling studies. J. Med. Chem. 2004, 47, 4998–5008. [Google Scholar] [PubMed]
- Maccarrone, M.; Salvati, S.; Bari, M.; Finazzi-Agró. Anandamide and 2-arachidonoylglycerol inhibit fatty acid amide hydrolase by activating the lipoxygenase pathway of the arachidonate cascade. Biochem. Biophys. Res. Commun. 2000, 278, 576–583. [Google Scholar]
- Kathuria, S.; Gaetani, S.; Fegley, D.; Valiño, F.; Duranti, A.; Tontini, A.; Mor, M.; Tarzia, G.; La Rana, G.; Calignano, A.; Giustino, A.; Tattoli, M.; Palmery, M.; Cuomo, V.; Piomelli, D. Modulation of anxiety through blockade of anandamide hydrolysis. Nat. Med. 2003, 9, 76–81. [Google Scholar]
- Karlsson, M.; Reue, K.; Xia, Y.R.; Lusis, A.J.; Langin, D.; Tornqvist, H.; Holm, C. Exon-intron organization and chromosomal localization of the mouse monoglyceride lipase gene. Gene 2001, 272, 11–18. [Google Scholar]
- Gulyas, A.I.; Cravatt, B.F.; Bracey, M.H.; Dinh, T.P.; Piomelli, D.; Boscia, F.; Freund, T.F. Segregation of two endocannabinoid-hydrolyzing enzymes into pre- and postsynaptic compartments in the rat hippocampus, cerebellum and amygdala. Eur. J. Neurosci. 2004, 20, 441–458. [Google Scholar]
- Lu, D.; Vemuri, V.K.; Duclos, R.I.; Makriyannis, A. The cannabinergic system as a target for anti-inflammatory therapies. Curr. Top. Med. Chem. 2006, 6, 1401–1426. [Google Scholar] [PubMed]
- Walter, L.; Stella, N. Cannabinoids and neuroinflammation. Br. J. Pharmacol. 2004, 141, 775–785. [Google Scholar]
- Guzman, M.; Sanchez, C.; Galve-Roperh, I. Cannabinoids and cell fate. Pharmacol. Ther. 2002, 95, 175–184. [Google Scholar]
- Berdyshev, E.V. Cannabinoid receptors and the regulation of immune response. Chem. Phys. Lipids 2000, 108, 169–190. [Google Scholar]
- Cota, D.; Marsicano, G.; Lutz, B.; Vicennati, V.; Stalla, G.K.; Pasquali, R.; Pagotto, U. Endogenous cannabinoid system as a modulator of food intake. Int. J. Obes. Relat Metab Disord. 2003, 27, 289–301. [Google Scholar]
- Shepherd Gordon, M. The Synaptic Organization of the Brain, Fifth ed; Oxford University Press: Oxford, UK/ New York, NY, USA, 2004. [Google Scholar]
- Finlay, B.L.; Darlington, R.B.; Nicastro, N. Developmental structure in brain evolution. Behav. Brain Sci. 2001, 20, 263–308. [Google Scholar]
- Wilson, R.I.; Nicoll, R.A. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 2001, 410, 588–592. [Google Scholar]
- Adermark, L.; Lovinger, D.M. Retrograde endocannabinoid signalling at striatal synapses requires a regulated postsynaptic release step. Proc. Natl. Acad. Sci. USA 2007, 104, 20564–20569. [Google Scholar]
- Gong, J.P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 receptors: immunohistochemical localization in rat brain. Brain Res. 2006, 1071, 10–23. [Google Scholar]
- Terasawa, E. Gonadotropin-releasing hormone II: is this neuropeptide important for mammalian reproduction? Endocrinology 2003, 144, 3–4. [Google Scholar]
- Pierantoni, R.; Cobellis, G.; Meccariello, R.; Fasano, S. Evolutionary aspects of cellular comunication in the vertebrate hypothalamo-hypophysio-gonadal axis. Int. Rev. Cytol. 2002, 218, 69–141. [Google Scholar]
- Tsou, K.; Brown, S.; Sanudo-pena, M.C.; Mackie, K.; Walker, J.M. Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system. Neuroscience 1998, 83, 393–411. [Google Scholar]
- Cristino, L.; De Petrocellis, L.; Pryce, G.; Baker, D.; Guglielmotti, V.; Di Marzo, V. Immunohistochemical localization of cannabinoid type 1 and vanilloid transient receptor potential vanilloid type 1 receptors in the mouse brain. Neuroscience 2006, 139, 1405–1415. [Google Scholar]
- Yang, H.T.; Karoum, F.; Felder, C.; Badger, H.; Wang, T.L.; Markey, S.P. GC/MS analysis of anandamide and quantification of N-arachidonoylphosphatidylethanolamides in various brain regions, spinal cord, testis, and spleen of the rat. J. Neurochem. 1999, 72, 1959–1968. [Google Scholar] [PubMed]
- Blankman, J.; Simon, G.M.; Cravatt, B. A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem. Biol. 2007, 14, 1347–1356. [Google Scholar]
- Salio, C.; Doly, S.; Fischer, J.; Franzoni, M.F.; Conrath, M. Neuronal and astrocytic localization of the cannabinoid receptor-1 in the dorsal horn of the rat spinal cord. Neurosci. Lett. 2002, 329, 13–16. [Google Scholar]
- Salio, C.; Fischer, J.; Franzoni, M.F.; Conrath, M. Pre- and postsynaptic localizations of the CB1 cannabinoid receptor in the dorsal horn of the rat spinal cord. Neuroscience 2002, 110, 755–764. [Google Scholar]
- Soderstrom, K.; Leid, M.; Moore, F.L.; Murray, T.F. Behavioral, pharmacological and molecular characterization of an amphibian cannabinoid receptor. J. Neurochem. 2000, 75, 413–423. [Google Scholar]
- Meccariello, R.; Chianese, R.; Cacciola, G.; Cobellis, G.; Pierantoni, R.; Fasano, S. Type-1 cannabinoid receptor expression in the frog, Rana esculenta, tissues: a possible involvement in the regulation of testicular activity. Mol. Reprod. Develop. 2006, 73, 551–558. [Google Scholar]
- Cottone, E.; Forno, S.; Campantico, E.; Guastalla, A.; Voltrono, L.; Mackie, K.; Franzoni, M.F. Expression and distribution of CB1 cannabinoid receptors in the central nervous system of the African cichlid fish Pelvicachromis pulcher. J. Comp. Neurol. 2005, 485, 293–303. [Google Scholar]
- Valenti, M.; Cottone, E.; Martinez, R.; De Pedro, N.; Rubio, M.; Viveros, M.P.; Franzoni, M.F.; Delgado, M.J.; Di Marzo, V. The endocannabinoid system in the brain of Carassius auratus and its possible role in the control of food intake. J. Neurochem. 2005, 95, 662–672. [Google Scholar]
- Meccariello, R.; Franzoni, M.F.; Chianese, R.; Cottone, E.; Scarpa, D.; Donna, D.; Cobellis, G.; Guastalla, A.; Pierantoni, R.; Fasano, S. Interplay between the endocannabinoid system and GnRH1 in the forebrain of the anuran amphibian Rana esculenta. Endocrinol. 2008, 149, 2149–2158. [Google Scholar]
- Schuel, H.; Burkman, L.J. A tale of two cells: endocannabinoid-signaling regulates functions of neurons and sperm. Biol. Reprod. 2005, 73, 1078–1086. [Google Scholar]
- Stella, N. How might cannabinoids influence sexual behaviour? Proc. Natl. Acad. Sci. USA 2001, 98, 793–795. [Google Scholar] [CrossRef]
- Murphy, L.L.; Adrian, B.A.; Kohli, M. Inhibition of luteinizing hormone secretion by delta9-tetrahydrocannabinol in the ovariectomized rat: effect of pretreatment with neurotransmitter or neuropeptide receptor antagonists. Steroids 1999, 64, 664–671. [Google Scholar]
- Pagotto, U.; Marsicano, G.; Cota, D.; Lutz, B.; Pasquali, R. The emerging role of the endocannabinoid system in endocrine regulation and energy balance. Endoc. Rev. 2006, 27, 73–100. [Google Scholar]
- Gonzalez, S.; Manzanares, J.; Berrendero, F.; Wenger, T.; Corchero, J.; Bisogno, T.; Romero, J.; Fuentes, J.A.; Di Marzo, V.; Ramos, J.A.; Fernández-Ruiz, J. Identification of endocannabinoids and cannabinoid CB1 receptor mRNA in the pituitary gland. Neuroendocrinology 1999, 70, 137–145. [Google Scholar]
- Gonzalez, S.; Bisogno, T.; Wenger, T.; Manzanares, J.; Milone, A.; Berrendero, F.; Di Marzo, V.; Ramos, J.A.; Fernandez-Ruiz, J.J. Sex steroid influence on cannabinoids CB1 receptor mRNA and endocannabinoid levels in the anterior pituitary gland. Biochem. Biophys. Res. Commun. 2000, 270, 260–266. [Google Scholar]
- Wenger, T.; Toth, B.E.; Juaneda, C.; Leonardelli, J.; Tramu, G. The effects of cannabinoids on the regulation of reproduction. Life Sci. 1999, 65, 695–702. [Google Scholar]
- Pagotto, U.; Marsicano, G.; Fezza, F.; Theodoropoulou, M.; Gruebler, Y.; Stalla, J.; Arzberger, T.; Milone, A.; Losa, M.; Di Marzo, V.; Lutz, B.; Stalla, G.K. Normal human pituitary gland and pituitary adenomas express cannabinoid receptor type 1 and synthesize endogenous cannabinoids: first evidence for a direct role of cannabinoids on hormone modulation at the human pituitary level. J. Clin. Endocrinol. Metab. 2001, 86, 2687–2696. [Google Scholar]
- Cesa, R.; Guastalla, A.; Cottone, E.; Mackie, K.; Beltramo, M.; Franzoni, M.F. Relationships between CB1 cannabinoid receptors and pituitary endocrine cells in Xenopus laevis: an immunohistochemical study. Gen. Comp. Endocrinol. 2002, 125, 17–24. [Google Scholar]
- Fernandez-Ruiz, J.J.; Munoz, R.M.; Romer, J.; Villanua, M.A.; Makryannis, A.; Ramos, J.A. Time course of the effects of differents cannabimimetics on prolactin and gonadotropin secretion: evidence for the presence of CB1 receptors in hypothalamic structures and their involvement in the effects of cannabimimetics. Biochem. Pharmacol. 1997, 53, 1919–1927. [Google Scholar]
- Bonnin, A.; Ramos, J.A.; Rodriguez de Fonseca, F.; Cebeira, M.; Fernandez-Ruiz, J.J. Acute effects of Δ9-tetrahydrocannbinol on tuberoinfundibular dopaminergic activity, anterior pituitary sensitivity to dopamine and prolactin release vary as a function of estrous cycle. Neuroendocrinology 1993, 58, 280–286. [Google Scholar] [PubMed]
- Rodriguez de Fonseca, F.; Cebeira, M.; Ramos, J.A.; Martin, M.; Fernandez-Ruiz, J.J. Cannabinoid receptors in rat brain areas: sexual differences, fluctuations during estrous cycle and changes after gonadectomy and sex steroid replacement. Life Sci. 1994, 54, 159–170. [Google Scholar]
- Hughes, C.L.; Everett, J.W.; Tyrey, L. Δ9-Tetrahydrocannabinol suppression of prolactin secretion is modulated by estrogens. Proc. Natl. Acad. Sci. USA 2003, 100, 2134–2139. [Google Scholar]
- Scorticati, C.; Mohn, C.; De Laurentis, A.; Vissio, P.; Fernandez-Solari, J.; Seilicovich, A.; McCann, S.M.; Rettori, V. The effect of anandamide on prolactin secretion is modulated by estrogen. Proc. Natl. Acad. Sci. USA 2003, 100, 2134–2139. [Google Scholar]
- Ho, B.Y.; Stadnicka, A.; Prather, P.L.; Buckley, A.R.; Current, L.L.; Bosnjak, Z.L.; Kwok, W.M. Cannabinoid CB1 receptor-mediated inhibition of prolactin release and signaling mechanisms in GH4C1 cells. Endocrinology 2000, 141, 1675–1685. [Google Scholar]
- Meldeson, J.H.; Mello, N.K.; Ellingboe, J.; Skupny, A.S.; Lex, B.W.; Griffin, M. Marijuana smoking suppresses luteinizing hormone in women. J. Pharmacol. Exp. Ther. 1986, 237, 862–866. [Google Scholar]
- Symons, A.M.; Teale, J.D.; Marks, V. Proceedings : effect of Δ9-tetrahydrocannabinol on the hypothalamic-pituitary-gonadal system in the maturing male rat. J. Endocrinol. 1976, 68, 43–44. [Google Scholar]
- Dixit, V.P.; Sharma, V.N.; Lohiya, N.K. The effect of chronically administered cannabis extract on the testicular function of mice. Eur. J. Pharmacol. 1974, 26, 111–114. [Google Scholar]
- Fernandez-Solari, J.; Scorticati, C.; Mohn, C.; De Laurentis, A.; Billi, S.; Franchi, A.; McCann, S.M.; Rettori, V. Alcohol inhibits luteinizing hormone-releasing hormone release by activating the endocannabinoid system. Proc. Natl. Acad. Sci. USA 2004, 101, 3264–3268. [Google Scholar]
- Scorticati, C.; Fernandez-Solari, J.; De Laurentis, A.; Mohn, C.; Prestifilippo, J.P.; Lasaga, M.; Seilicovich, A.; Billi, S.; Franchi, A.; McCann, S.M.; et al. The inhibitory effect of anandamide on luteinizing hormone-releasing hormone secretion is reversed by estrogen. Proc. Natl. Acad. Sci. USA 2004, 101, 11891–11896. [Google Scholar]
- de Miguel, R.; Romero, J.; Munoz, R.M.; Garcia-Gil, L.; Gonzalez, S.; Villanua, M.A.; Makriyannis, A.; Ramos, J.A.; Fernandez-Ruiz, J.J. Effects of cannabinoids on prolactin and gonadotropin secretion: involvement of changes in hypothalamic gamma-aminobutyric acid (GABA) inputs. Biochem. Pharmacol. 1998, 56, 1331–1338. [Google Scholar]
- Gammon, C.M.; Freeman, G.M.; Xie, W.; Petersen, S.L.; Wetsel, W. Regulation of gonadotropin-releasing hormone secretion by cannabinoids. Endocrinology 2005, 146, 4491–4499. [Google Scholar]
- Todman, M.G.; Han, S.K.; Herbison, A.E. Profiling neurotransmitter receptor expression in mouse gonadotropin-releasing hormone neurons using green fluorescent protein-promoter transgenics and microarrays. Neuroscience 2005, 132, 705–712. [Google Scholar]
- Herrick, C.J. The brain of the tiger salamander. The University of Chicago Press: Chicago, 1948. [Google Scholar]
- Fasano, S.; Goos, G.J.; Janssen, C.; Pierantoni, R. Two GnRHs fluctuate in correlation with androgen levels in the male frog Rana esculenta. J. Exp. Zool. 1993, 266, 277–283. [Google Scholar]
- Chianese, R.; Cobellis, G.; Pierantoni, R.; Fasano, S.; Meccariello, R. Non-mammalian vertebrate models and the endocannabinoid system: relationship with gonadotropin-releasing hormone. Mol. Cell Endocrinol. 2008, 286, S46–S51. [Google Scholar]
- Fernandez-Fernandez, R.; Martini, A.C.; Navarro, V.M.; Castellano, J.M.; Dieguez, C.; Aguilar, E.; Pinilla, L.; Tena-Sempere, M. Novel signals for the integration of energy bilance and reproduction. Mol. Cell Endocrinol. 2006, 254-255, 127–132. [Google Scholar] [PubMed]
- Wynne, K.; Stanley, S.; McGowan, B.; Bloom, S. Appetite control. J. Endocrinol. 2005, 184, 291–318. [Google Scholar]
- Hatami-Baroogh, L.; Razavi, S.; Zarkesh-Esfahani, H.; Tavalaee, M.; Tanhaei, S.; Ghaedi, K.; Deemeh, M.R.; Rabiee, F.; Nasr-Esfahani, M.H. Evaluation of the leptin receptor in human spermatozoa. Reprod. Biol. Endocrinol. 2010, 23, 8–17. [Google Scholar]
- Arias-Alvarez, M.; Garcia-Garcia, R.M.; Torres-Rovira, L.; Gonzalez-Bulnes, A.; Rebollar, P.G.; Lorenzo, P.L. Influence of leptin on in vitro maturation and steroidogenic secretion of cumulus-oocyte complexes through JAK2/STAT3 and MEK1/2 pathways in the rabbit model. Reproduction 2010, 139, 523–532. [Google Scholar]
- Sarkar, M.; Schilffarth, S.; Scams, D.; Meyer, H.H.; Berisha, B. The expression of leptin and its receptor during different physiological stages in the bovine ovary. Mol. Reprod. Dev. 2010, 77, 174–181. [Google Scholar]
- Mah, P.M.; Wittert, G.A. Obesity and testicular function. Mol. Cell. Endocrinol. 2010, 316, 180–186. [Google Scholar]
- Backholer, K.; Smith, J.T.; Rao, A.; Pereira, A.; Iqbal, J.; Ogawa, S.; Li, Q.; Clarke, I.J. Kisspeptin cells in the ewe brain respond to leptin and communicate with neuropeptides Y and proopiomelanocortin cells. Endocrinology 2010, 151. in press. [Google Scholar]
- Popa, S.M.; Clifton, D.K.; Steiner, R.A. The role of kisspeptins and GPR54 in the neuroendocrine regulation of reproduction. Annu. Rev. Physiol. 2008, 70, 213–238. [Google Scholar]
- Tena-Sempere, M. Kiss-1 and reproduction: focus on its role in the metabolic regulation of fertility. Neuroendocrinology 2006, 83, 275–281. [Google Scholar]
- Di Marzo, V.; Goparaju, S.K.; Wang, L.; Liu, J.; Batkai, S.; Jarai, Z.; Fezza, F.; Miura, G.I.; Palmiter, R.D.; Sugiura, T.; et al. Leptin-regulated endocannabinoids are involved in maintaining food intake. Nature 2001, 410, 822–825. [Google Scholar] [PubMed]
- Hanus, L.; Avraham, Y.; Ben-Shushan, D.; Zolotarev, O.; Berry, E.M.; Mechoulam, R. Short-term fasting and prolonged semistarvation have opposite effects on 2-AG levels in mouse brain. Brain Res. 2003, 983, 144–151. [Google Scholar]
- Mani, S.K.; Mitchell, A.; O’Malley, B.W. Progesterone receptor and dopamine receptors are required in Δ9-tetrahydrocannabinol modulation of sexual receptivity in female rats. Proc. Natl. Acad. Sci. USA 2001, 98, 1249–1254. [Google Scholar]
- Mani, S.K.; Allen, J.M.C.; Lydon, J.P.; Mulac-Jericevic, B.; Blaustein, J.D.; DeMayo, F.J.; Conneely, O.M.; O’Malley, B.W. Dopamine requires unoccupied progesterone receptor to induce sex behavior in mice. Mol. Endocrinol. 1996, 10, 1728–1737. [Google Scholar]
- Turley, W.A.; Floody, O.R. Delta-9-tetrahydrocannabinol stimulates receptive and proceptive sexual behaviors in female hamsters. Pharmacol. Biochem. Behav. 1981, 14, 745–747. [Google Scholar]
- Coddington, E.; Moore, F.L. Neuroendocrinology of context-dependent stress responses: vasocitin alters the effect of corticosterone on amphibian behaviors. Horm. Behav. 2003, 43, 222–228. [Google Scholar]
- Coddington, E.; Lewis, C.; Rose, J.D.; Moore, F.L. Endocannabinoids mediate the effects of acute stress and corticosterone on sex behavior. Endocrinology 2007, 148, 493–500. [Google Scholar]
- Wetzel, D.M.; Kelley, D.B. Androgen and gonadotropin effects on male mate calls in South African clawed frogs, Xenopus laevis. Horm. Behav. 1983, 17, 388–404. [Google Scholar] [PubMed]
- Kelley, D.B.; Denninson, J. The vocal motor neurons of Xenopus laevis: development of sex differences in axon number. J. Neurobiol. 1990, 21, 869–882. [Google Scholar]
- Brahic, C.J.; Kelley, D.B. Vocal circuitry in Xenopus laevis telencephalon to laryngeal motor neurons. J. Comp. Neurol. 2003, 464, 115–130. [Google Scholar]
- Cottone, E.; Guastalla, A.; Mackie, K.; Franzoni, M.F. Endocannabinoids affect the reproductive functions in teleosts and amphibian. Mol. Cell Endocrinol. 2008, 286, S41–S45. [Google Scholar]
- Burmeister, S.; Somes, C.; Wilczynski, W. Behavioral and hormonal effects of exogenous vasotocin and corticosterone in the green treefrog. Gen. Comp. Endocrinol. 2001, 122, 189–197. [Google Scholar]
- Zornik, E.; Kelley, D.B. Breathing and calling: neuronal networks in the Xenopus laevis hindbrain. J. Comp. Neurol. 2007, 501, 303–315. [Google Scholar]
- Pierantoni, R.; Cobellis, G.; Meccariello, R.; Cacciola, G.; Chianese, R.; Chioccarelli, T.; Fasano, S. CB1 activity in male reproduction: mammalian and nonmammalian animal models. Vitam. Horm. 2009, 81, 367–387. [Google Scholar]
- Kalantaridou, S.N.; Makrigiannakis, A.; Zoumakis, E.; Chrousos, G.P. Reproductive functions of corticotropin-releasing hormone. Research and potential clinical utility of antalarmins (CRH receptor type 1 antagonists). Am. J. Reprod. Immunol. 2004, 51, 269–274. [Google Scholar] [PubMed]
- Dubey, A.K.; Plant, T.M. A suppression of gonadotropin secretion by cortisol in castrated male reshus monkeys (Macaca mulatta) mediated by the interruption of hypothalamic gonadotropin-relesasing hormone release. Biol. Reprod. 1985, 33, 423–431. [Google Scholar] [PubMed]
- Lerman, S.A.; Miller, G.K.; Bohlman, K.; Albaladejo, V.; Leonard, J.F.; Devas, V.; Clark, R.L. Effects of corticosterone on reproduction in male Sprague-Dawley rats. Reprod. Toxicol. 1997, 11, 799–805. [Google Scholar]
- Rivier, C.; Rivest, S. Effect of stress on the activity of the hypothalamic-pituitary-gonadal axis: peripheral and central mechanisms. Biol. Reprod. 1991, 45, 523–532. [Google Scholar]
- Tsutsui, K.; Saigoh, E.; Ukana, K.; Teranishi, H.; Fujisawa, Y.; Kikuchi, M.; Ishii, S.; Sharp, P.J. A novel avian hypothalamic peptide inhibiting gonadotropin release. Biochem. Biophys. Res. Commun. 2000, 275, 661–667. [Google Scholar]
- Kirby, E.D.; Geraghty, A.C.; Ubuka, T.; Bently, G.E.; Kaufer, D. Stress increases putative gonadotropin inhibitory hormone and decreases luteinizing hormone in male rats. Proc. Natl. Acad. Sci. USA 2009, 106, 11324–11329. [Google Scholar]
- Patel, S.; Hillard, C.J. Adaptations in endocannabinoid signalling in response to repeated homotypic stress: a novel mechanism for stress habituation. Eur. J. Neurosci. 2008, 27, 2821–2829. [Google Scholar]
- Ledent, C.; Valverde, O.; Cossu, G.; Petitet, F.; Aubert, J.F.; Beslot, F.; Bohme, G.A.; Imperato, A.; Pedrazzini, T.; Roques, B.P.; Vassart, G.; Fratta, W.; Parmentier, M. Unresponsiveness to cannabinoids and reduced addictive effects of opiates in CB1 receptor knockout mice. Science 1999, 283, 401–404. [Google Scholar]
- Wenger, T.; Ledent, C.; Tramu, G. The endogenous cannabinoid, anandamide, activates the hypothalamo-pituitary-adrenal axis in CB1 cannabinoid receptor knockout mice. Neuroendocrinology 2003, 78, 294–300. [Google Scholar]
- Van Pett, K.; Viau, V.; Bittencourt, J.C.; Chan, R.K.; Li, H.Y.; Arias, C.; Prins, G.S.; Perrin, M.; Vale, W.; Sawchenko, P.E. Distribution of mRNAs encoding CRF receptors in brain and pituitary of rat and mouse. J. Comp. Neurol. 2000, 428, 191–212. [Google Scholar]
- Hermann, H.; Lutz, B. Coexpression of the cannabinoid receptor type 1 with the corticotrophin-releasing hormone receptor type 1 in distinct regions of the adult mouse forebrain. Neurosci. Lett. 2005, 375, 13–18. [Google Scholar]
- Cyr, N.E.; Romero, L.M. Identifying hormonal habituation in field studies of stress. Gen. Comp. Endocrinology 2009, 161, 295–303. [Google Scholar]
- Hill, M.N.; McLaughlin, R.J.; Bingham, B.; Shrestha, L.; Lee, T.T.Y.; Megan Gray, J.; Hillard, C.J.; Gorzalka, B.B.; Viau, V. Endogenous cannabinoid signalling is essential for stress adaptation. Proc. Natl. Acad. Sci. USA 2010, 107, 9406–9411. [Google Scholar]
- Patel, S.; Roelke, C.T.; Rademacher, D.J.; Cullinan, W.E.; Hillard, C.J. Endocannabinoid signalling negatively modulates stress-induced activation of the hypothalamic-pituitary-adrenal axis. Endocrinology 2004, 145, 5431–5438. [Google Scholar]
- Bayatti, N.; Hermann, H.; Lutz, B.; Behl, C. Corticotropin-releasing hormone-mediated induction of intracellular signaling pathways and brain-derived neurotrophic factor expression is inhibited by the activation of the endocannabinoid system. Endocrinology 2005, 146, 1205–1213. [Google Scholar]
- Di Marzo, V. The endocannabinoid system: its general strategy of action, tools for its pharmacological manipulation and potential therapeutical exploitation. Pharmacol. Res. 2009, 60, 77–84. [Google Scholar]
- Fawcett, D.W.; Leak, L.V.; Heidger, P.M., Jr. Electron microscopic observations on the structural components of the blood-testis barrier. J. Reprod. Fertil. Suppl 1970, 10, 105–122. [Google Scholar]
- Heindel, J.J.; Keith, W.B. Specific inhibition of FSH-stimulated cAMP accumulation by delta 9-tetrahydrocannabinol in cultures of rat Sertoli cells. Toxicol. Appl. Pharmacol. 1989, 101, 124–134. [Google Scholar]
- Leblond, C.P.; Clermont, Y. Spermiogenesis of rat, mouse, hamster and guinea pig as revealed by the periodic acid-fuchsin sulfurous acid technique. Am. J. Anat. 1952, 90, 167–215. [Google Scholar]
- Ross, M.H. The Sertoli cell junctional specialization during spermiogenesis and at spermiation. Anat. Rec. 1976, 186, 79–87. [Google Scholar]
- Harclerode, J. Endocrine effects of marijuana in the male: preclinical studies. NIDA Res. Monogr. 1984, 44, 46–64. [Google Scholar]
- Dalterio, S.; Badr, F.; Bartke, A.; Mayfield, D. Cannabinoids in male mice: effects on fertility and spermatogenesis. Science 1982, 216, 315–316. [Google Scholar]
- Dalterio, S.; Bartke, A.; Roberson, C.; Watson, D.; Burstein, S. Direct and pituitary-mediated effects of delta9-THC and cannabinol on the testis. Pharmacol. Biochem. Behav. 1978, 8, 673–678. [Google Scholar]
- Fujimoto, G.I.; Morrill, G.A.; O'Connell, M.E.; Kostellow, A.B.; Retura, G. Effects of cannabinoids given orally and reduced appetite on the male rat reproductive system. Pharmacol. 1982, 24, 303–313. [Google Scholar]
- Wang, H.; Dey, S.K.; Maccarrone, M. Jekyll and Hyde: two faces of cannabinoid signaling in male and female fertility. Endocr. Rev. 2006, 27, 427–448. [Google Scholar]
- Pierantoni, R.; Cobellis, G.; Meccariello, R.; Cacciola, G.; Chianese, R.; Chioccarelli, T.; Fasano, S. Testicular gonadotropin-releasing hormone activity, progression of spermatogenesis, and sperm transport in vertebrate. Ann. N. Y. Acad. Sci. 2009, 1163, 279–291. [Google Scholar]
- Sun, X.; Wang, H.; Okabe, M.; Mackie, K.; Kingsley, P.J.; Marnett, L.J.; Cravatt, B.F.; Dey, S.K. Genetic loss of Faah compromises male fertility in mice. Biol. Reprod. 2009, 80, 235–242. [Google Scholar]
- Cobellis, G.; Cacciola, G.; Scarpa, D.; Meccariello, R.; Chianese, R.; Franzoni, M.F.; Mackie, K.; Pierantoni, R.; Fasano, S. Endocannabinoid system in frog and rodent testis: type-1 cannabinoid receptor and fatty acid amide hydrolase activity in male germ cells. Biol. Reprod. 2006, 75, 82–89. [Google Scholar]
- Cacciola, G.; Chioccarelli, T.; Mackie, K.; Meccariello, R.; Ledent, C.; Fasano, S.; Pierantoni, R.; Cobellis, G. Expression of type-1 cannabinoid receptor during rat postnatal testicular development: possible involvement in adult leydig cell differentiation. Biol. Reprod. 2008, 79, 758–765. [Google Scholar]
- Maccarrone, M.; Cecconi, S.; Rossi, G.; Battista, N.; Pauselli, R.; Finazzi-Agro, A. Anandamide activity and degradation are regulated by early postnatal aging and follicle-stimulating hormone in mouse Sertoli cells. Endocrinology 2003, 144, 20–28. [Google Scholar]
- Rossi, G.; Gasperi, V.; Paro, R.; Barsacchi, D.; Cecconi, S.; Maccarrone, M. Follicle-stimulating hormone activates fatty acid amide hydrolase by protein kinase A and aromatase-dependent pathways in mouse primary Sertoli cells. Endocrinology 2007, 148, 1431–1439. [Google Scholar]
- Sharpe, R.M.; McKinnell, C.; Kivlin, C.; Fisher, J.S. Proliferation and functional maturation of Sertoli cells, and their relevance to disorders of testis function in adulthood. Reproduction 2003, 125, 769–784. [Google Scholar]
- Atanassova, N.; McKinnell, C.; Walker, M.; Turner, K.J.; Fisher, J.S.; Morley, M.; Millar, M.R.; Groome, N.P.; Sharpe, R.M. Permanent effects of neonatal estrogen exposure in rats on reproductive hormone levels, Sertoli cell number, and the efficiency of spermatogenesis in adulthood. Endocrinology 1999, 140, 5364–5373. [Google Scholar]
- Rastogi, R.K. The control of spermatogenesis in the green frog, Rana esculenta. J. Exp. Zool. 1976, 169, 151–166. [Google Scholar]
- D'Istria, M.; Delrio, G.; Botte, V.; Chieffi, G. Radioimmunoassay of testosterone, 17beta-oestradiol and oestrone in the male and female plasma of plasma of Rana esculenta during sexual cycle. Steroids Lipids Res. 1974, 5, 42–48. [Google Scholar]
- Pierantoni, R.; Iela, L.; d'Istria, M.; Fasano, S.; Rastogi, R.K.; Delrio, G. Seasonal testosterone profile and testicular responsiveness to pituitary factors and gonadotrophin releasing hormone during two different phases of the sexual cycle of the frog (Rana esculenta). J. Endocrinol. 1984, 102, 387–392. [Google Scholar]
- Meccariello, R.; Chianese, R.; Cobellis, G.; Pierantoni, R.; Fasano, S. Cloning of type 1 cannabinoid receptor in Rana esculenta reveals differences between genomic sequence and cDNA. FEBS J. 2007, 274, 2909–2920. [Google Scholar]
- Tsai, S.J.; Wang, Y.C.; Hong, C.J. Association study of a cannabinoid receptor gene (CNR1) polymorphism and schizophrenia. Psychiatr Genet. 2000, 10, 149–151. [Google Scholar]
- Martínez-Gras, I.; Hoenicka, J.; Ponce, G.; Rodríguez-Jiménez, R.; Jiménez-Arriero, M.A.; Pérez-Hernandez, E.; Ampuero, I.; Ramos-Atance, J.A.; Palomo, T.; Rubio, G. (AAT)n repeat in the cannabinoid receptor gene, CNR1: association with schizophrenia in a Spanish population. Eur Arch Psychiatry Clin Neurosci. 2006, 256, 437–441. [Google Scholar]
- Fasano, S.; Meccariello, R.; Cobellis, G.; Chianese, R.; Cacciola, G.; Chioccarelli, T.; Pierantoni, R. The endocannabinoid system: an ancient signaling involved in the control of male fertility. Ann. N. Y. Acad. Sci. 2009, 1163, 112–124. [Google Scholar]
- Gye, M.C.; Kang, H.H.; Kang, H.J. Expression of cannabinoid receptor 1 in mouse testes. Arch. Androl 2005, 51, 247–255. [Google Scholar]
- Cacciola, G.; Chioccarelli, T.; Ricci, G.; Meccariello, R.; Fasano, S.; Pierantoni, R.; Cobellis, G. The endocannabinoid system in vertebrate male reproduction: a comparative overview. Mol. Cell. Endocrinology 2008, 286, S24–S30. [Google Scholar]
- Habert, R.; Lejeune, H.; Saez, J.M. Origin, differentiation and regulation of fetal and adult Leydig cells. Mol. Cell. Endocrinol. 2001, 179, 47–74. [Google Scholar]
- Liu, Q.R.; Pan, C.H.; Hishimoto, A.; Li, C.Y.; Xi, Z.X.; Llorente-Berzal, A.; Viveros, M.P.; Ishiguro, H.; Arinami, T.; Onaivi, E.S.; Uhl, G.R. Species differences in cannabinoid receptor 2 (CNR2 gene): identification of novel human and rodent CB2 isoforms, differential tissue expression and regulation by cannabinoid receptor ligands. Genes Brain Behav. 2009, 8, 519–530. [Google Scholar]
- Mizrak, S.C.; van Dissel-Emiliani, F.M. Transient receptor potential vanilloid receptor-1 confers heat resistance to male germ cells. Fertil. Steril. 2008, 90, 1290–1293. [Google Scholar]
- Yanagimachi, R. Fertility of mammalian spermatozoa: its development and relativity. Zygote 1994, 2, 371–372. [Google Scholar]
- Orgebin-Crist, M.C. Sperm maturation in rabbit epididymis. Nature 1967, 216, 816–818. [Google Scholar] [PubMed]
- Suarez, S.S.; Pacey, A.A. Sperm transport in the female reproductive tract. Hum. Reprod. Update 2006, 12, 23–37. [Google Scholar]
- Ho, H.C.; Suarez, S.S. Hyperactivation of mammalian spermatozoa: function and regulation. Reproduction 2001, 122, 519–526. [Google Scholar]
- Wassarman, P.; Chen, J.; Cohen, N.; Litscher, E.; Liu, C.; Qi, H.; Williams, Z. Structure and function of the mammalian egg zona pellucida. J. Exp. Zool. 1999, 285, 251–258. [Google Scholar]
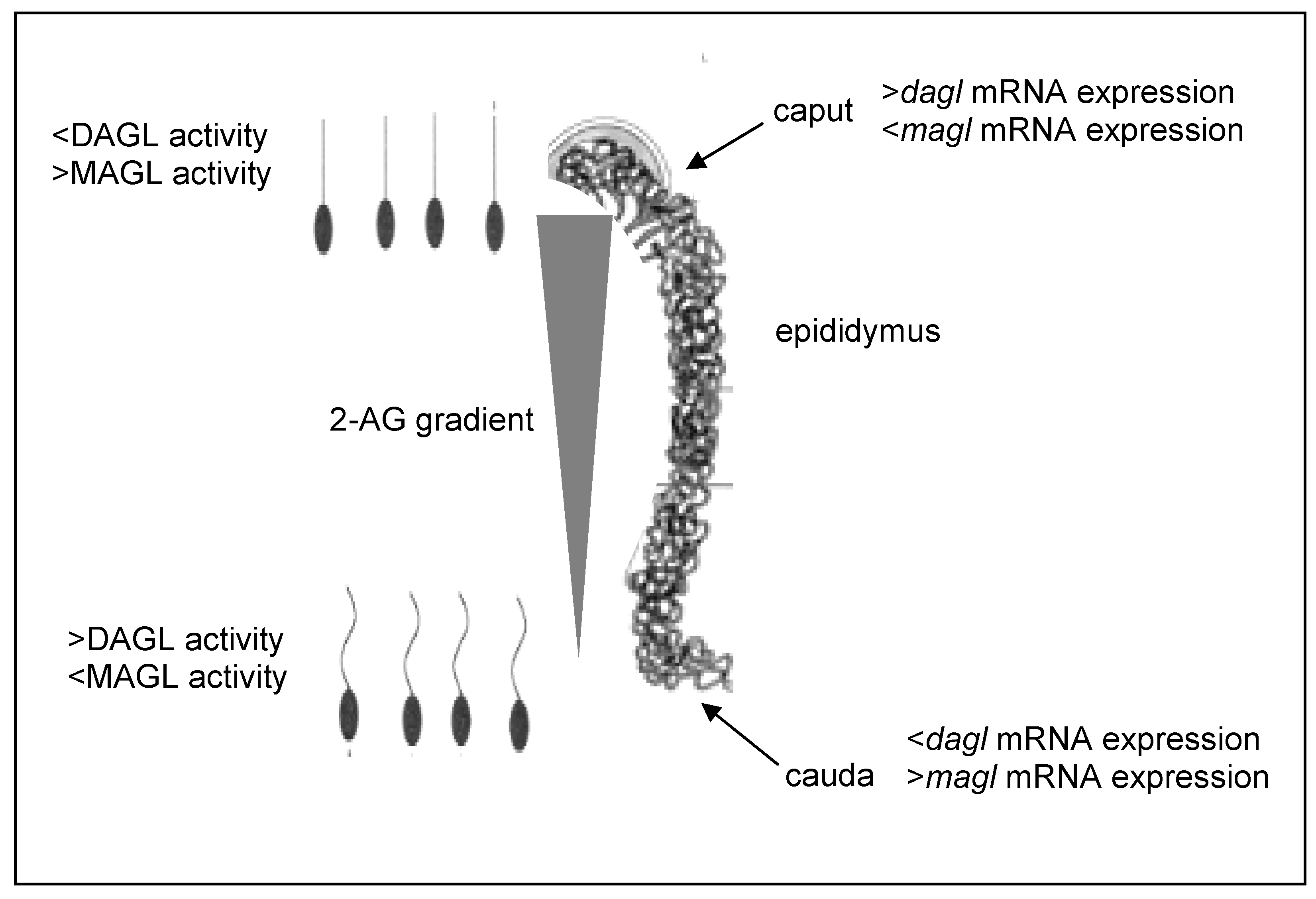
- Cobellis, G.; Ricci, G.; Cacciola, G.; Orlando, P.; Petrosino, S.; Cascio, M.G.; Bisogno, T.; De Petrocellis, L.; Chioccarelli, T.; Altucci, L.; Fasano, S.; Meccariello, R.; Pierantoni, R.; Ledent, C.; Di Marzo, V. A gradient of 2-arachidonoylglycerol regulates mouse epididymal sperm cell start-up. Biol. Reprod. 2010, 82, 451–458. [Google Scholar]
- Schuel, H.; Burkman, L.J.; Lippes, J.; Crickard, K.; Forester, E.; Piomelli, D.; Giuffrida, A. N-Acylethanolamines in human reproductive fluids. Chem. Phys. Lipids 2002, 121, 211–227. [Google Scholar]
- Nahas, G.G.; Frick, H.C.; Lattimer, J.K.; Latour, C.; Harvey, D. Pharmacokinetics of THC in brain and testis, male gametotoxicity and premature apoptosis of spermatozoa. Hum. Psychopharmacol. 2002, 17, 103–113. [Google Scholar]
- Francavilla, F.; Battista, N.; Barbonetti, A.; Vassallo, M.R.; Rapino, C.; Antonangelo, C.; Pasquariello, N.; Catanzaro, G.; Barboni, B.; Maccarrone, M. Characterization of the endocannabinoid system in human spermatozoa and involvement of transient receptor potential vanilloid 1 receptor in their fertilizing ability. Endocrinology 2009, 150, 4692–4700. [Google Scholar]
- Schuel, H.; Burkman, L.J.; Lippes, J.; Crickard, K.; Mahony, M.C.; Giuffrida, A.; Picone, R.P.; Makriyannis, A. Evidence that anandamide-signaling regulates human sperm functions required for fertilization. Mol. Reprod. Dev. 2002, 63, 376–387. [Google Scholar]
- Aquila, S.; Guido, C.; Santoro, A.; Perrotta, I.; Laezza, C.; Bifulco, M.; Sebastiano, A. Human sperm anatomy: ultrastructural localization of the cannabinoid1 receptor and a potential role of anandamide in sperm survival and acrosome reaction. Anat. Rec. (Hoboken) 2010, 293, 298–309. [Google Scholar]
- Rossato, M.; Ion, P.F.; Ferigo, M.; Clari, G.; Foresta, C. Human sperm express cannabinoid receptor Cb1, the activation of which inhibits motility, acrosome reaction, and mitochondrial function. J. Clin. Endocrinol. Metab. 2005, 90, 984–991. [Google Scholar] [PubMed]
- Agirregoitia, E.; Carracedo, A.; Subiran, N.; Valdivia, A.; Agirregoitia, N.; Peralta, L.; Velasco, G.; Irazusta, J. The CB(2) cannabinoid receptor regulates human sperm cell motility. Fertil. Steril. 2010, 93, 1378–1387. [Google Scholar]
- Chang, M.C.; Berkery, D.; Schuel, R.; Laychock, S.G.; Zimmerman, A.M.; Zimmerman, S.; Schuel, H. Evidence for a cannabinoid receptor in sea urchin sperm and its role in blockade of the acrosome reaction. Mol. Reprod. Dev. 1993, 36, 507–516. [Google Scholar]
- Schuel, H.; Burkman, L.J. A tale of two cells: endocannabinoid-signaling regulates functions of neurons and sperm. Biol. Reprod. 2005, 73, 1078–1086. [Google Scholar]
- Ricci, G.; Cacciola, G.; Altucci, L.; Meccariello, R.; Pierantoni, R.; Fasano, S.; Cobellis, G. Endocannabinoid control of sperm motility: the role of epididymus. Gen. Comp. Endocrinol. 2007, 153, 320–322. [Google Scholar] [PubMed]
- Aquila, S.; Guido, C.; Laezza, C.; Santoro, A.; Pezzi, V.; Panza, S.; Andò, S.; Bifulco, M. A new role of anandamide in human sperm: focus on metabolism. J. Cell. Physiol. 2009, 221, 147–153. [Google Scholar]
- Welch, J.E.; Brown, P.L.; O'Brien, D.A.; Magyar, P.L.; Bunch, D.O.; Mori, C.; Eddy, E.M. Human glyceraldehyde 3-phosphate dehydrogenase-2 gene is expressed specifically in spermatogenic cells. J. Androl. 2000, 21, 328–338. [Google Scholar]
- Miki, K.; Qu, W.; Goulding, E.H.; Willis, W.D.; Bunch, D.O.; Strader, L.F.; Perreault, S.D.; Eddy, E.M.; O'Brien, D.A. Glyceraldehyde 3-phosphate dehydrogenase-S, a sperm-specific glycolytic enzyme, is required for sperm motility and male fertility. Proc. Natl. Acad. Sci. USA 2004, 101, 16501–16506. [Google Scholar]
- Windsor, D.P.; White, I.G. Assessment of ram sperm mitochondrial function by quantitative determination of sperm rhodamine 123 accumulation. Mol. Reprod. Dev. 1993, 36, 354–360. [Google Scholar]
- Ballester, J.; Fernández-Novell, J.M.; Rutllant, J.; García-Rocha, M.; Jesús Palomo, M.; Mogas, T.; Peña, A.; Rigau, T.; Guinovart, J.J.; Rodríguez-Gil, J.E. Evidence for a functional glycogen metabolism in mature mammalian spermatozoa. Mol. Reprod. Dev. 2000, 56, 207–219. [Google Scholar]
- Reeve, V.C.; Grant, J.D.; Robertson, W.; Gillespie, H.K.; Hollister, L.E. Plasma concentrations of delta-9-tetrahydrocannabinol and impaired motor function. Drug Alcohol Depend. 1983, 11, 167–175. [Google Scholar]
- Whan, L.B.; West, M.C.; McClure, N.; Lewis, S.E. Effects of delta-9-tetrahydrocannabinol, the primary psychoactive cannabinoid in marijuana, on human sperm function in vitro. Fertil. Steril. 2006, 85, 653–660. [Google Scholar] [PubMed]
- Badawy, Z.S.; Chohan, K.R.; Whyte, D.A.; Penefsky, H.S.; Brown, O.M.; Souid, A.K. Cannabinoids inhibit the respiration of human sperm. Fertil. Steril. 2009, 91, 2471–2476. [Google Scholar]
- Gervasi, M.G.; Rapanelli, M.; Ribeiro, M.L.; Farina, M.; Billi, S.; Franchi, A.M.; Perez, M.S. The endocannabinoid system in bull sperm and bovine oviductal epithelium: role of anandamide in sperm-oviduct interaction. Reproduction 2009, 137, 403–414. [Google Scholar]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; Mechoulam, R.; Pertwee, R.G. International Union of Pharmacology. XVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar]
- Darszon, A.; Labarca, P.; Nishigaki, T.; Espinosa, F. Ion channels in sperm physiology. Physiol Rev. 1999, 79, 481–510. [Google Scholar]
- Battista, N.; Rapino, C.; Di Tommaso, M.; Bari, M.; Pasquariello, N.; Maccarrone, M. Regulation of male fertility by the endocannabinoid system. Mol. Cell. Endocrinol. 2008, 286, S17–S23. [Google Scholar]
- Maykut, M.O. Health consequences of acute and chronic marihuana use. Prog. Neuropsychopharmacol. Biol. Psychiatry. 1985, 9, 209–238. [Google Scholar]
- Pope, H.G., Jr.; Yurgelun-Todd, D. The residual cognitive effects of heavy marijuana use in college students. JAMA 1996, 275, 521–527. [Google Scholar]
- Bäckström, C.T.; McNeilly, A.S.; Leask, R.M.; Baird, D.T. Pulsatile secretion of LH, FSH, prolactin, oestradiol and progesterone during the human menstrual cycle. Clin. Endocrinol. 1982, 17, 29–42. [Google Scholar]
- Bauman, J.E. Comparison of radioimmunoassay results in serum and plasma. Clin. Chem. 1980, 26, 676–677. [Google Scholar]
- Mueller, B.A.; Daling, J.R.; Weiss, N.S.; Moore, D.E. Recreational drug use and the risk of primary infertility. Epidemiology 1990, 1, 195–200. [Google Scholar] [PubMed]
- Klonoff-Cohen, H.S.; Natarajan, L.; Chen, R.V. A prospective study of the effects of female and male marijuana use on in vitro fertilization (IVF) and gamete intrafallopian transfer (GIFT) outcomes. Am. J. Obstet. Gynecol. 2006, 194, 369–376. [Google Scholar]
- Murphy, L.L.; Muñoz, R.M.; Adrian, B.A.; Villanúa, M.A. Function of cannabinoid receptors in the neuroendocrine regulation of hormone secretion. Neurobiol. Dis. 1998, 5, 432–346. [Google Scholar]
- Tyrey, L. delta 9-Tetrahydrocannabinol: a potent inhibitor of episodic luteinizing hormone secretion. J. Pharmacol.Exp. Ther. 1980, 213, 306–308. [Google Scholar]
- Adashi, E.Y.; Jones, P.B.; Hsueh, A.J. Direct antigonadal activity of cannabinoids: suppression of rat granulosa cell functions. Am. J. Physiol. 1983, 244, E177–E185. [Google Scholar]
- Nir, I.; Ayalon, D.; Tsafriri, A.; Cordova, T.; Lindner, H.R. Suppression of the cyclic surge of luteinizing hormone secretion and of ovulation in the rat by delta 1-tetrahydrocannabinol. Nature 1973, 243, 470–471. [Google Scholar] [PubMed]
- Ayalon, D.; Nir, I.; Cordova, T.; Bauminger, S.; Puder, M.; Naor, Z.; Kashi, R.; Zor, U.; Harell, A.; Lindner, H.R. Acute effect of delta1-tetrahydrocannabinol on the hypothalamo-pituitary-ovarian axis in the rat. Neuroendocrinology 1977, 23, 31–42. [Google Scholar]
- Smith, C.G.; Besch, N.F.; Smith, R.G.; Besch, P.K. Effect of tetrahydrocannabinol on the hypothalamic-pituitary axis in the ovariectomized rhesus monkey. Fertil. Steril. 1979, 31, 335–339. [Google Scholar] [PubMed]
- Reich, R.; Laufer, N.; Lewysohn, O.; Cordova, T.; Ayalon, D.; Tsafriri, A. In vitro effects of cannabinoids on follicular function in the rat. Biol. Reprod. 1982, 27, 223–231. [Google Scholar]
- Lewysohn, O.; Cordova, T.; Nimrod, A.; Ayalon, D. The suppressive effect of delta-1-tetrahydrocannabinol on the steroidogenic activity of rat granulosa cells in culture. Horm. Res. 1984, 19, 43–51. [Google Scholar]
- Geber, W.F.; Schramm, L.C. Effect of marihuana extract on fetal hamsters and rabbits. Toxicol. Appl. Pharmacol. 1969, 14, 276–282. [Google Scholar]
- Persaud, T.V.; Ellington, A.C. Cannabis in early pregnancy. Lancet 1967, 2, 1306. [Google Scholar]
- El-Talatini, M.R.; Taylor, A.H.; Elson, J.C.; Brown, L.; Davidson, A.C.; Konje, J.C. Localisation and function of the endocannabinoid system in the human ovary. PLoS One 2009, 4, e4579. [Google Scholar]
- El-Talatini, M.R.; Taylor, A.H.; Konje, J.C. The relationship between plasma levels of the endocannabinoid, anandamide, sex steroids, and gonadotrophins during the menstrual cycle. Fertil. Steril. 2010, 93, 1989–1996. [Google Scholar] [PubMed]
- Wassarman, P.M.; Jovine, L.; Litscher, E.S. A profile of fertilization in mammals. Nat. Cell. Biol. 2001, 3, E59–E64. [Google Scholar]
- Psychoyos, A. Hormonal control of ovoimplantation. Vitam. Horm. 1973, 31, 201–256. [Google Scholar]
- Paria, B.C.; Das, S.K.; Dey, S.K. The preimplantation mouse embryo is a target for cannabinoid ligand-receptor signaling. Proc. Natl. Acad. Sci. USA 1995, 92, 9460–9464. [Google Scholar]
- Paria, B.C.; Ma, W.; Andrenyak, D.M.; Schmid, P.C.; Schmid, H.H.; Moody, D.E.; Deng, H.; Makriyannis, A.; Dey, S.K. Effects of cannabinoids on preimplantation mouse embryo development and implantation are mediated by brain-type cannabinoid receptors. Biol. Reprod. 1998, 58, 1490–1495. [Google Scholar] [PubMed]
- Paria, B.C.; Song, H.; Wang, X.; Schmid, P.C.; Krebsbach, R.J.; Schmid, H.H.; Bonner, T.I.; Zimmer, A.; Dey, S.K. Dysregulated cannabinoid signaling disrupts uterine receptivity for embryo implantation. J. Biol. Chem. 2001, 276, 20523–20538. [Google Scholar]
- Wang, H.; Guo, Y.; Wang, D.; Kingsley, P.J.; Marnett, L.J.; Das, S.K.; DuBois, R.N.; Dey, S.K. Aberrant cannabinoid signaling impairs oviductal transport of embryos. Nat. Med. 2004, 10, 1074–1080. [Google Scholar]
- Buckley, N.E.; McCoy, K.L.; Mezey, E.; Bonner, T.; Zimmer, A.; Felder, C.C.; Glass, M.; Zimmer, A. Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB(2) receptor. Eur. J. Pharmacol. 2000, 396, 141–149. [Google Scholar]
- Heilman, R.D.; Reo, R.R.; Hahn, D.W. Changes in the sensitivity of adrenergic receptors in the oviduct during early gestation in the rabbit. Fertil. Steril. 1976, 27, 426–430. [Google Scholar]
- Horne, A.W.; Phillips, J.A., III; Kane, N.; Lourenco, P.C.; McDonald, S.E.; Williams, A.R.; Simon, C.; Dey, S.K.; Critchley, H.O. CB1 expression is attenuated in Fallopian tube and decidua of women with ectopic pregnancy. PLoS One 2008, 3, e3969. [Google Scholar]
- Paria, B.C.; Huet-Hudson, Y.M.; Dey, S.K. Blastocyst's state of activity determines the "window" of implantation in the receptive mouse uterus. Proc. Natl. Acad. Sci. USA 1993, 90, 10159–10162. [Google Scholar]
- Huet, Y.M.; Andrews, G.K.; Dey, S.K. Modulation of c-myc protein in the mouse uterus during pregnancy and by steroid hormones. Prog. Clin. Biol.Res. 1989, 294, 401–412. [Google Scholar]
- Yoshinaga, K.; Fujino, M. Hormonal control of implantation in the rat: inhibition by luteinizing hormone-releasing hormone and its analogues. Ciba Found. Symp. 1978, 64, 85–110. [Google Scholar]
- Dey, S.K. Focus on implantation. Reproduction 2004, 128, 655–656. [Google Scholar]
- Dey, S.K. Reproductive biology: fatty link to fertility. Nature 2005, 435, 34–35. [Google Scholar]
- Martínez Orgado, J.A.; Fernández López, D.; Bonet Serra, B.; Lizasoain Hernández, I.; Romero Paredes, J. The cannabinoid system and its importance in the perinatal period. An. Pediatr. 2005, 63, 433–440. [Google Scholar]
- Paria, B.C.; Zhao, X.; Wang, J.; Das, S.K.; Dey, S.K. Fatty-acid amide hydrolase is expressed in the mouse uterus and embryo during the periimplantation period. Biol. Reprod. 1999, 60, 1151–1157. [Google Scholar]
- Maccarrone, M.; De Felici, M.; Bari, M.; Klinger, F.; Siracusa, G.; Finazzi-Agrò, A. Down-regulation of anandamide hydrolase in mouse uterus by sex hormones. Eur. J. Biochem. 2000, 267, 2991–2997. [Google Scholar]
- Schmid, P.C.; Paria, B.C.; Krebsbach, R.J.; Schmid, H.H.; Dey, S.K. Changes in anandamide levels in mouse uterus are associated with uterine receptivity for embryo implantation. Proc. Natl. Acad. Sci. USA 1997, 94, 4188–4192. [Google Scholar]
- Guo, Y.; Wang, H.; Okamoto, Y.; Ueda, N.; Kingsley, P.J.; Marnett, L.J.; Schmid, H.H.; Das, S.K.; Dey, S.K. N-acylphosphatidylethanolamine-hydrolyzing phospholipase D is an important determinant of uterine anandamide levels during implantation. J. Biol. Chem. 2005, 280, 23429–234932. [Google Scholar]
- Wang, H.; Xie, H.; Sun, X.; Kingsley, P.J.; Marnett, L.J.; Cravatt, B.F.; Dey, S.K. Differential regulation of endocannabinoid synthesis and degradation in the uterus during embryo implantation. Prost. Other Lipid. Mediat. 2007, 83, 62–74. [Google Scholar]
- Ribeiro, M.L.; Vercelli, C.A.; Sordelli, M.; Farina, M.G.; Cervini, M.; Billi, S.; Franchi, A.M. 17beta-oestradiol and progesterone regulate anandamide synthesis in the rat uterus. Reprod. Biomed. Online 2009, 18, 209–218. [Google Scholar]
- Wang, H.; Matsumoto, H.; Guo, Y.; Paria, B.C.; Roberts, R.L.; Dey, S.K. Differential G protein-coupled cannabinoid receptor signaling by anandamide directs blastocyst activation for implantation. Proc Natl Acad Sci USA 2003, 100, 14914–14919. [Google Scholar]
- El-Talatini, M.R.; Taylor, A.H.; Konje, J.C. Fluctuation in anandamide levels from ovulation to early pregnancy in in-vitro fertilization-embryo transfer women, and its hormonal regulation. Hum. Reprod. 2009, 24, 1989–1998. [Google Scholar]
- Maccarrone, M.; Bisogno, T.; Valensise, H.; Lazzarin, N.; Fezza, F.; Manna, C.; Di Marzo, V.; Finazzi-Agrò, A. Low fatty acid amide hydrolase and high anandamide levels are associated with failure to achieve an ongoing pregnancy after IVF and embryo transfer. Mol. Hum. Reprod. 2002, 8, 188–195. [Google Scholar]
- Piccinni, M.P.; Beloni, L.; Livi, C.; Maggi, E.; Scarselli, G.; Romagnani, S. Defective production of both leukemia inhibitory factor and type 2 T-helper cytokines by decidual T cells in unexplained recurrent abortions. Nat. Med. 1998, 4, 1020–1024. [Google Scholar]
- Stewart, C.L.; Cullinan, E.B. Preimplantation development of the mammalian embryo and its regulation by growth factors. Dev. Genet. 1997, 21, 91–101. [Google Scholar]
- Szekeres-Bartho, J.; Wegmann, T.G. A progesterone-dependent immunomodulatory protein alters the Th1/Th2 balance. J. Reprod. Immunol. 1996, 31, 81–95. [Google Scholar]
- Maccarrone, M.; Valensise, H.; Bari, M.; Lazzarin, N.; Romanini, C.; Finazzi-Agrò, A. Progesterone up-regulates anandamide hydrolase in human lymphocytes: role of cytokines and implications for fertility. J. Immunol. 2001, 166, 7183–7189. [Google Scholar]
- Maccarrone, M.; Bari, M.; Di Rienzo, M.; Finazzi-Agrò, A.; Rossi, A. Progesterone activates fatty acid amide hydrolase (FAAH) promoter in human T lymphocytes through the transcription factor Ikaros. Evidence for a synergistic effect of leptin. J. Biol. Chem. 2003, 278, 32726–32732. [Google Scholar] [PubMed]
- Habayeb, O.M.; Taylor, A.H.; Evans, M.D.; Cooke, M.; Taylor, D.J.; Bell, S.C.; Konje, J.C. Plasma levels of the endocannabinoid anandamide in women--a potential role in pregnancy maintenance and labor? J. Clin. Endocrinol. Metab. 2004, 89, 5482–5487. [Google Scholar] [PubMed]
- Nallendran, V.; Lam, P.M.; Marczylo, T.H.; Bankart, M.J.; Taylor, A.H.; Taylor, D.J.; Konje, J.C. The plasma levels of the endocannabinoid, anandamide, increase with the induction of labour. Intern. J. Obst. Gynaecol. 2010, 117, 863–869. [Google Scholar]
- Habayeb, O.M.; Taylor, A.H.; Finney, M.; Evans, M.D.; Konje, J.C. Plasma anandamide concentration and pregnancy outcome in women with threatened miscarriage. J.A.M.A. 2008, 299, 1135–1136. [Google Scholar]
- Marczylo, T.H.; Lam, P.M.; Amoako, A.A.; Konje, J.C. Anandamide levels in human female reproductive tissues: solid-phase extraction and measurement by ultraperformance liquid chromatography tandem mass spectrometry. Anal. Biochem. 2010, 400, 155–162. [Google Scholar]
- Helliwell, R.J.; Chamley, L.W.; Blake-Palmer, K.; Mitchell, M.D.; Wu, J.; Kearn, C.S.; Glass, M. Characterization of the endocannabinoid system in early human pregnancy. J. Clin. Endocrinol. Metab. 2004, 89, 5168–5174. [Google Scholar]
- Trabucco, E.; Acone, G.; Marenna, A.; Pierantoni, R.; Cacciola, G.; Chioccarelli, T.; Mackie, K.; Fasano, S.; Colacurci, N.; Meccariello, R.; Cobellis, G.; Cobellis, L. Endocannabinoid system in first trimester placenta: low FAAH and high CB1 expression characterize spontaneous miscarriage. Placenta 2009, 30, 516–522. [Google Scholar]
- Park, B.; Gibbons, H.M.; Mitchell, M.D.; Glass, M. Identification of the CB1 cannabinoid receptor and fatty acid amide hydrolase (FAAH) in the human placenta. Placenta 2003, 24, 990–995. [Google Scholar]
- Acone, G.; Trabucco, E.; Colacurci, N.; Cobellis, L.; Mackie, K.; Meccariello, R.; Cacciola, G.; Chioccarelli, T.; Fasano, S.; Pierantoni, R.; Cobellis, G. Low type I cannabinoid receptor levels characterize placental villous in labouring delivery. Placenta 2009, 30, 203–205. [Google Scholar]
- Marletta, M.A. Nitric oxide synthase structure and mechanism. J. Biol. Chem. 1993, 268, 12231–12234. [Google Scholar]
- Fernández Celadilla, L.; Carbajo Rueda, M.; Muñoz Rodríguez, M. Prolonged inhibition of nitric oxide synthesis in pregnant rats: effects on blood pressure, fetal growth and litter size. Arch. Gynecol. Obstet. 2005, 271, 243–248. [Google Scholar]
- Amit, I.; Thaler, Y.; Paz, Y.; Itskovitz-Eldor, J. The effect of a nitric oxide donor on Doppler flow velocity waveforms in the uterine artery during the first trimester of pregnancy. Ultras. Obstet. Gynecol. 1998, 11, 94–98. [Google Scholar]
- Chaudhuri, G.; Cuevas, J.; Buga, G.M.; Ignarro, L.J. NO is more important than PGI2 in maintaining low vascular tone in feto-placental vessels. Am. J. Physiol. 1993, 265, H2036–H2043. [Google Scholar] [PubMed]
- Cella, M.; Leguizamón, G.F.; Sordelli, M.S.; Cervini, M.; Guadagnoli, T.; Ribeiro, M.L.; Franchi, A.M.; Farina, M.G. Dual effect of anandamide on rat placenta nitric oxide synthesis. Placenta 2008, 29, 699–707. [Google Scholar]
- Mitchell, M.D.; Sato, T.A.; Wang, A.; Keelan, J.A.; Ponnampalam, A.P.; Glass, M. Cannabinoids stimulate prostaglandin production by human gestational tissues through a tissue- and CB1-receptor-specific mechanism. Am. J. Physiol. Endocrinol. Metab. 2008, 294, 352–356. [Google Scholar]
- Wang, H.; Xie, H.; Dey, S.K. Loss of cannabinoid receptor CB1 induces preterm birth. PLoS One 2008, 3, e3320. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cacciola, G.; Chianese, R.; Chioccarelli, T.; Ciaramella, V.; Fasano, S.; Pierantoni, R.; Meccariello, R.; Cobellis, G. Cannabinoids and Reproduction: A Lasting and Intriguing History. Pharmaceuticals 2010, 3, 3275-3323. https://doi.org/10.3390/ph3103275
Cacciola G, Chianese R, Chioccarelli T, Ciaramella V, Fasano S, Pierantoni R, Meccariello R, Cobellis G. Cannabinoids and Reproduction: A Lasting and Intriguing History. Pharmaceuticals. 2010; 3(10):3275-3323. https://doi.org/10.3390/ph3103275
Chicago/Turabian StyleCacciola, Giovanna, Rosanna Chianese, Teresa Chioccarelli, Vincenza Ciaramella, Silvia Fasano, Riccardo Pierantoni, Rosaria Meccariello, and Gilda Cobellis. 2010. "Cannabinoids and Reproduction: A Lasting and Intriguing History" Pharmaceuticals 3, no. 10: 3275-3323. https://doi.org/10.3390/ph3103275
APA StyleCacciola, G., Chianese, R., Chioccarelli, T., Ciaramella, V., Fasano, S., Pierantoni, R., Meccariello, R., & Cobellis, G. (2010). Cannabinoids and Reproduction: A Lasting and Intriguing History. Pharmaceuticals, 3(10), 3275-3323. https://doi.org/10.3390/ph3103275