
Hydroxylated Dimeric Naphthoquinones Increase the Generation of Reactive Oxygen Species, Induce Apoptosis of Acute Myeloid Leukemia Cells and Are Not Substrates of the Multidrug Resistance Proteins ABCB1 and ABCG2

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Dimeric Naphthoquinones Decreased Viable Numbers of AML Cell Line and Primary Cells with Favorable Therapeutic Index in Relation to Normal Hematopoietic Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | BiQ-1 (µM) | BiQ-2 (µM) | MonoQ (µM) |
|---|---|---|---|
| MOLM-14 (complex karyotype, FLT3-ITD) | 3.1 ± 0.2 ** | 4.5 ± 1.9 ** | >100 (NE) |
| THP-1 (complex karyotype, FLT3-WT) | 8.5 ± 4.2 * | 8.6 ± 4.3 | >100 (NE) |
| AML-A (46,XY; FLT3-WT) | 0.36 | 3 | >100 (NE) |
| AML-B (46,XY; FLT3-WT) | 3.3 ± 0.3 * | NT | NT |
| AML-C (complex karyotype; relapsed post-transplant; FLT3-ITD) | 5.1 ± 0.7 * | NT | NT |
| Normal BM | 14.6 | 14.1 | NT |
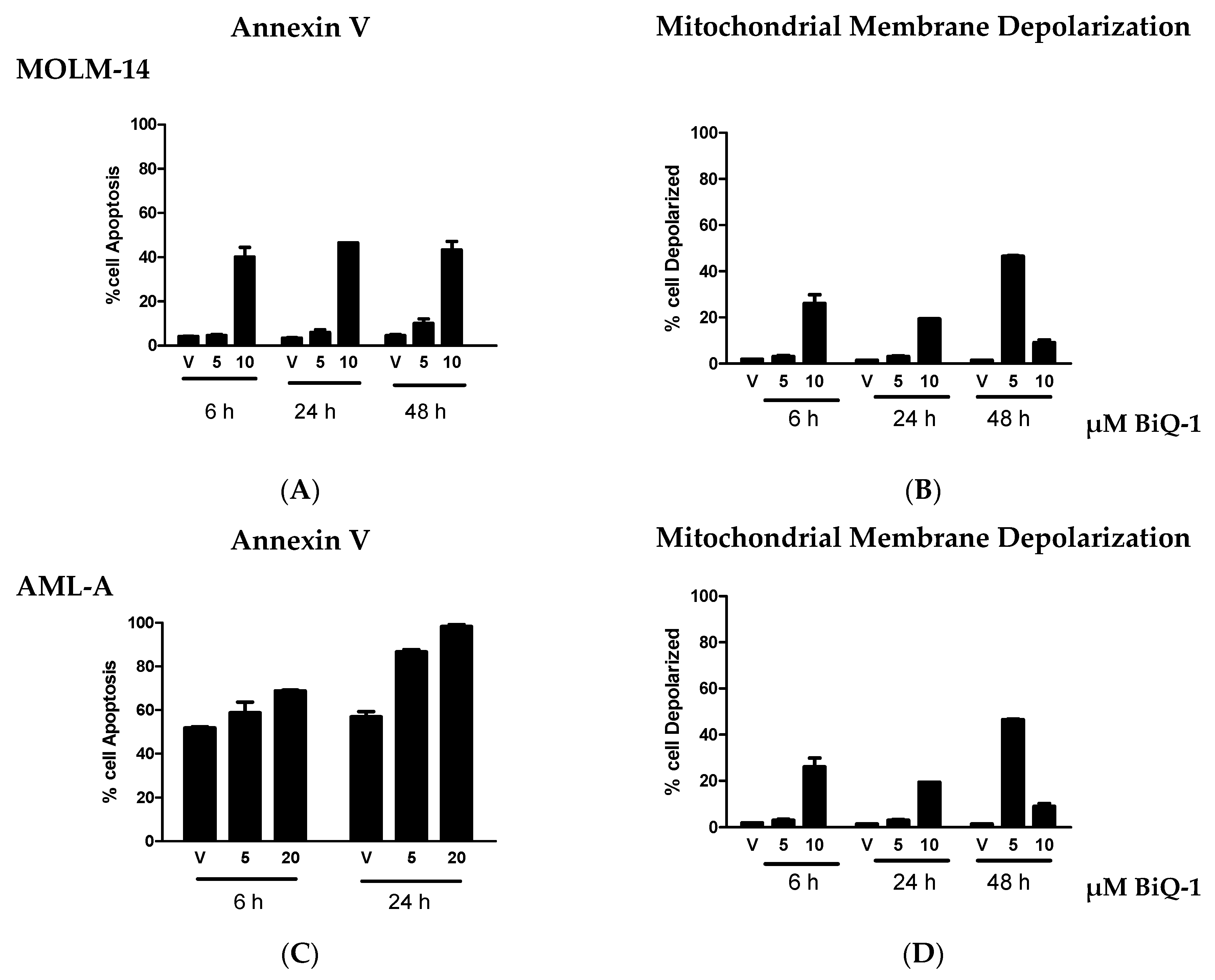
2.2. BiQ-1 Induces Apoptosis of AML Cells


2.3. ROS Induction is Evident after BiQ-1 Exposure

2.4. BiQ-1 Inhibits Clonogenic Growth of AML Cell Lines

2.5. BiQ-1 Is Not a Substrate of the ATP Binding Cassette (ABC) Transporters


2.6. BiQ-1 Was Well Tolerated in Female Swiss Webster Mice

| Vehicle | BiQ-1 (10 mg/kg) | Normal Range | Units | |||
|---|---|---|---|---|---|---|
| ID number of mouse | 1 | 3 | 2 | 4 | ||
| White Blood Cell | 9.46 | 9.68 | 4.80 | 16.73 | 3.2–12.7 | (× 103 cells/µL) |
| Neutrophil | 5.90 | 7.30 | 16.40 | 23.50 | % | |
| Lymphocyte | 86.80 | 82.90 | 61.70 | 62.10 | % | |
| Monocyte | 1.80 | 2.00 | 3.00 | 3.10 | % | |
| Eosinophil | 4.40 | 5.40 | 17.70 | 10.10 | % | |
| Basophil | 0.20 | 0.20 | 0.30 | 0.20 | % | |
| Red Blood Cell | 8.11 | 10.02 | 7.28 | 11.07 | 7.0–10.1 | (× 103 cells/µL) |
| Hemoglobin | 13.50 | 15.90 | 11.50 | 17.50 | 11.8–14.9 | mg/dL |
| Hematocrit | 46.10 | 55.70 | 40.40 | 62.20 | 36.7–46.8 | % |
| Mean corpuscular volume | 56.90 | 55.60 | 55.50 | 56.20 | 42.2–59.2 | fL |
| Platelet | 1168 | 1328 | 1611 | 1944 | 766–1657 | (× 103 cells/µL) |
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Lines and Patient Cells
4.3. Cell Proliferation Assay
4.4. Cell Survival Assay
4.5. Clonogenic Assays
4.6. Western Blotting
4.7. Flow Cytometric Analysis of Apoptosis and Mitochondrial Membrane Potential
4.8. Measurement of Cellular Reactive Oxygen Species (ROS)
4.9. Uptake of Fluorescent ATP-Binding Cassette (ABC) Protein Substrates
4.10. In Vivo Tolerability Studies
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Buchner, T.; Schlenk, R.F.; Schaich, M.; Dohner, K.; Krahl, R.; Krauter, J.; Heil, G.; Krug, U.; Sauerland, M.C.; Heinecke, A.; et al. Acute myeloid leukemia (AML): Different treatment strategies versus a common standard arm—Combined prospective analysis by the german aml intergroup. J. Clin. Oncol. 2012, 30, 3604–3610. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; Karp, J.E. The state of the union on treatment of acute myeloid leukemia. Leuk. Lymphoma 2014, 55, 2423–2425. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Holohan, C.; van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Sallmyr, A.; Fan, J.; Datta, K.; Kim, K.T.; Grosu, D.; Shapiro, P.; Small, D.; Rassool, F. Internal tandem duplication of FLT3 (FLT3/ITD) induces increased ROS production, DNA damage, and misrepair: Implications for poor prognosis in AML. Blood 2008, 111, 3173–3182. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Li, L.; Small, D.; Rassool, F. Cells expressing FLT3/ITD mutations exhibit elevated repair errors generated through alternative nhej pathways: Implications for genomic instability and therapy. Blood 2010, 116, 5298–5305. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; Sadowska, M.; Carter-Cooper, B.; Bhatnagar, V.; van der Merwe, I.; Levis, M.J.; Sausville, E.A.; Lapidus, R.G. Perturbation of cellular oxidative state induced by dichloroacetate and arsenic trioxide for treatment of acute myeloid leukemia. Leuk. Res. 2015, 39, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Pei, S.; Minhajuddin, M.; Callahan, K.P.; Balys, M.; Ashton, J.M.; Neering, S.J.; Lagadinou, E.D.; Corbett, C.; Ye, H.; Liesveld, J.L.; et al. Targeting aberrant glutathione metabolism to eradicate human acute myelogenous leukemia cells. J. Biol. Chem. 2013, 288, 33542–33558. [Google Scholar] [CrossRef] [PubMed]
- Sriskanthadevan, S.; Jeyaraju, D.V.; Chung, T.E.; Prabha, S.; Xu, W.; Skrtic, M.; Jhas, B.; Hurren, R.; Gronda, M.; Wang, X.; et al. Aml cells have low spare reserve capacity in their respiratory chain that renders them susceptible to oxidative metabolic stress. Blood 2015, 125, 2120–2130. [Google Scholar] [CrossRef] [PubMed]
- Powis, G. Free radical formation by antitumor quinones. Free Radic. Biol. Med. 1989, 6, 63–101. [Google Scholar] [CrossRef]
- Driscoll, J.S. Quinone structure-antitumor activity relationships. Cancer Chemother. Rep. Part 2 1974, 4, 3–4. [Google Scholar]
- Keinan, S.; Paquette, W.D.; Skoko, J.J.; Beratan, D.N.; Yang, W.; Shinde, S.; Johnston, P.A.; Lazo, J.S.; Wipf, P. Computational design, synthesis and biological evaluation of para-quinone-based inhibitors for redox regulation of the dual-specificity phosphatase Cdc25B. Org. Biomol. Chem. 2008, 6, 3256–3263. [Google Scholar] [CrossRef] [PubMed]
- Wellington, K.W. Understanding cancer and the anticancer activities of naphthoquinones—A review. R. Soc. Chem. (RSC) Adv. 2015, 5, 20309–20338. [Google Scholar] [CrossRef]
- Decosterd, L.A.; Parsons, I.C.; Gustafson, K.R.; CardellinaII, J.H.; McMahon, J.B.; Cragg, G.M.; Murata, Y.; Pannell, L.K.; Steiner, J.R.; Clardy, J.; et al. Structure, absolute stereochemistry, and synthesis of conocurvone, a potent, novel HIV-inhibitory naphthoquinone trimer from a Conospermum sp. J. Am. Chem. Soc. 1993, 115, 6673–6679. [Google Scholar] [CrossRef]
- Dai, J.R.; Decosterd, L.A.; Gustafson, K.R.; Cardellina, J.H., 2nd; Gray, G.N.; Boyd, M.R. Novel naphthoquinones from Conospermum incurvum. J. Nat. Prod. 1994, 57, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; Harwood, J.S.; Kohanim, S.; Stagliano, K.W. Regiocontrolled synthesis of the trimeric quinone framework of conocurvone. Org. Lett. 2002, 4, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Stagliano, K.W.; Lu, Z.; Emadi, A.; Harwood, J.S.; Harwood, C.A. Effect of methoxyl group position on the regioselectivity of ammonia substitution reactions involving 3,3′-dichloro-2,2′-binaphthoquinones. J. Org. Chem. 2004, 69, 5128–5131. [Google Scholar] [CrossRef] [PubMed]
- Stagliano, K.W.; Emadi, A.; Lu, Z.; Malinakova, H.C.; Twenter, B.; Yu, M.; Holland, L.E.; Rom, A.M.; Harwood, J.S.; Amin, R.; et al. Regiocontrolled synthesis and HIV inhibitory activity of unsymmetrical binaphthoquinone and trimeric naphthoquinone derivatives of conocurvone. Bioorg. Med. Chem. 2006, 14, 5651–5665. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.E.; Emadi, A.; Marchionni, L.; Hurley, P.J.; Simons, B.W.; Schaeffer, E.M.; Vuica-Ross, M. Dimeric naphthoquinones, a novel class of compounds with prostate cancer cytotoxicity. BJU Int. 2011, 108, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; le, A.; Harwood, C.J.; Stagliano, K.W.; Kamangar, F.; Ross, A.E.; Cooper, C.R.; Dang, C.V.; Karp, J.E.; Vuica-Ross, M. Metabolic and electrochemical mechanisms of dimeric naphthoquinones cytotoxicity in breast cancer cells. Bioorg. Med. Chem. 2011, 19, 7057–7062. [Google Scholar] [CrossRef] [PubMed]
- Emadi, A.; Ross, A.E.; Cowan, K.M.; Fortenberry, Y.M.; Vuica-Ross, M. A chemical genetic screen for modulators of asymmetrical 2,2′-dimeric naphthoquinones cytotoxicity in yeast. PLoS ONE 2010, 5, e10846. [Google Scholar] [CrossRef] [PubMed]
- Bey, E.A.; Reinicke, K.E.; Srougi, M.C.; Varnes, M.; Anderson, V.E.; Pink, J.J.; Li, L.S.; Patel, M.; Cao, L.; Moore, Z.; et al. Catalase abrogates β-lapachone-induced PARP1 hyperactivation-directed programmed necrosis in NQO1-positive breast cancers. Mol. Cancer Ther. 2013, 12, 2110–2120. [Google Scholar] [CrossRef] [PubMed]
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2—An update. Free Radic. Biol. Med. 2014, 66, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, J.I.; Haber, M.; Henderson, M.J.; Norris, M.D. ABC transporters in cancer: More than just drug efflux pumps. Nat. Rev. Cancer 2010, 10, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Elangovan, S.; Hsieh, T.C. Control of cellular redox status and upregulation of quinone reductase NQO1 via Nrf2 activation by α-lipoic acid in human leukemia HL-60 cells. Int. J. Oncol. 2008, 33, 833–838. [Google Scholar] [PubMed]
- Irwin, M.E.; Rivera-Del Valle, N.; Chandra, J. Redox control of leukemia: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2013, 18, 1349–1383. [Google Scholar] [CrossRef] [PubMed]
- Duthie, S.J.; Grant, M.H. The role of reductive and oxidative metabolism in the toxicity of mitoxantrone, adriamycin and menadione in human liver derived Hep G2 hepatoma cells. Br. J. Cancer 1989, 60, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Anusevicius, Z.; Nivinskas, H.; Sarlauskas, J.; Sari, M.A.; Boucher, J.L.; Cenas, N. Single-electron reduction of quinone and nitroaromatic xenobiotics by recombinant rat neuronal nitric oxide synthase. Acta Biochim. Pol. 2013, 60, 217–222. [Google Scholar] [PubMed]
- Meyer, M.; Rubsamen, D.; Slany, R.; Illmer, T.; Stabla, K.; Roth, P.; Stiewe, T.; Eilers, M.; Neubauer, A. Oncogenic ras enables DNA damage- and p53-dependent differentiation of acute myeloid leukemia cells in response to chemotherapy. PLoS ONE 2009, 4, e7768. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Natarajan, K.; Bhullar, J.; Shukla, S.; Fang, H.B.; Cai, L.; Chen, Z.S.; Ambudkar, S.V.; Baer, M.R. The novel BCR-ABL and FLT3 inhibitor ponatinib is a potent inhibitor of the MDR-associated ATP-binding cassette transporter ABCG2. Mol. Cancer Ther. 2012, 11, 2033–2044. [Google Scholar] [CrossRef] [PubMed]
- Young, I.T. Proof without prejudice: Use of the Kolmogorov-Smirnov test for the analysis of histograms from flow systems and other sources. J. Histochem. Cytochem. 1977, 25, 935–941. [Google Scholar] [CrossRef]
- Minderman, H.; Suvannasankha, A.; O’Loughlin, K.L.; Scheffer, G.L.; Scheper, R.J.; Robey, R.W.; Baer, M.R. Flow cytometric analysis of breast cancer resistance protein expression and function. Cytometry 2002, 48, 59–65. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lapidus, R.G.; Carter-Cooper, B.A.; Sadowska, M.; Choi, E.Y.; Wonodi, O.; Muvarak, N.; Natarajan, K.; Pidugu, L.S.; Jaiswal, A.; Toth, E.A.; et al. Hydroxylated Dimeric Naphthoquinones Increase the Generation of Reactive Oxygen Species, Induce Apoptosis of Acute Myeloid Leukemia Cells and Are Not Substrates of the Multidrug Resistance Proteins ABCB1 and ABCG2. Pharmaceuticals 2016, 9, 4. https://doi.org/10.3390/ph9010004
Lapidus RG, Carter-Cooper BA, Sadowska M, Choi EY, Wonodi O, Muvarak N, Natarajan K, Pidugu LS, Jaiswal A, Toth EA, et al. Hydroxylated Dimeric Naphthoquinones Increase the Generation of Reactive Oxygen Species, Induce Apoptosis of Acute Myeloid Leukemia Cells and Are Not Substrates of the Multidrug Resistance Proteins ABCB1 and ABCG2. Pharmaceuticals. 2016; 9(1):4. https://doi.org/10.3390/ph9010004
Chicago/Turabian StyleLapidus, Rena G., Brandon A. Carter-Cooper, Mariola Sadowska, Eun Yong Choi, Omasiri Wonodi, Nidal Muvarak, Karthika Natarajan, Lakshmi S. Pidugu, Anil Jaiswal, Eric A. Toth, and et al. 2016. "Hydroxylated Dimeric Naphthoquinones Increase the Generation of Reactive Oxygen Species, Induce Apoptosis of Acute Myeloid Leukemia Cells and Are Not Substrates of the Multidrug Resistance Proteins ABCB1 and ABCG2" Pharmaceuticals 9, no. 1: 4. https://doi.org/10.3390/ph9010004
APA StyleLapidus, R. G., Carter-Cooper, B. A., Sadowska, M., Choi, E. Y., Wonodi, O., Muvarak, N., Natarajan, K., Pidugu, L. S., Jaiswal, A., Toth, E. A., Rassool, F. V., Etemadi, A., Sausville, E. A., Baer, M. R., & Emadi, A. (2016). Hydroxylated Dimeric Naphthoquinones Increase the Generation of Reactive Oxygen Species, Induce Apoptosis of Acute Myeloid Leukemia Cells and Are Not Substrates of the Multidrug Resistance Proteins ABCB1 and ABCG2. Pharmaceuticals, 9(1), 4. https://doi.org/10.3390/ph9010004