
Trio-Drug Combination of Sodium Valproate, Baclofen and Thymoquinone Exhibits Synergistic Anticonvulsant Effects in Rats and Neuro-Protective Effects in HEK-293 Cells

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Drugs and Dosing Schedule
2.3. Establishment and Assessment of MES Rat Model
2.4. Study Design
2.5. Estimation of Hippocampal mTOR, IL-1β and 1L-6 Levels
2.6. Invitro Studies
2.7. Molecular Modeling
2.8. Statistical Analysis
3. Results
3.1. Anticonvulsant Effect of Sodium Valproate (SVP), Baclofen (BFN), Thymoquinone (THQ) Alone and in Combination against MES-Induced Seizures

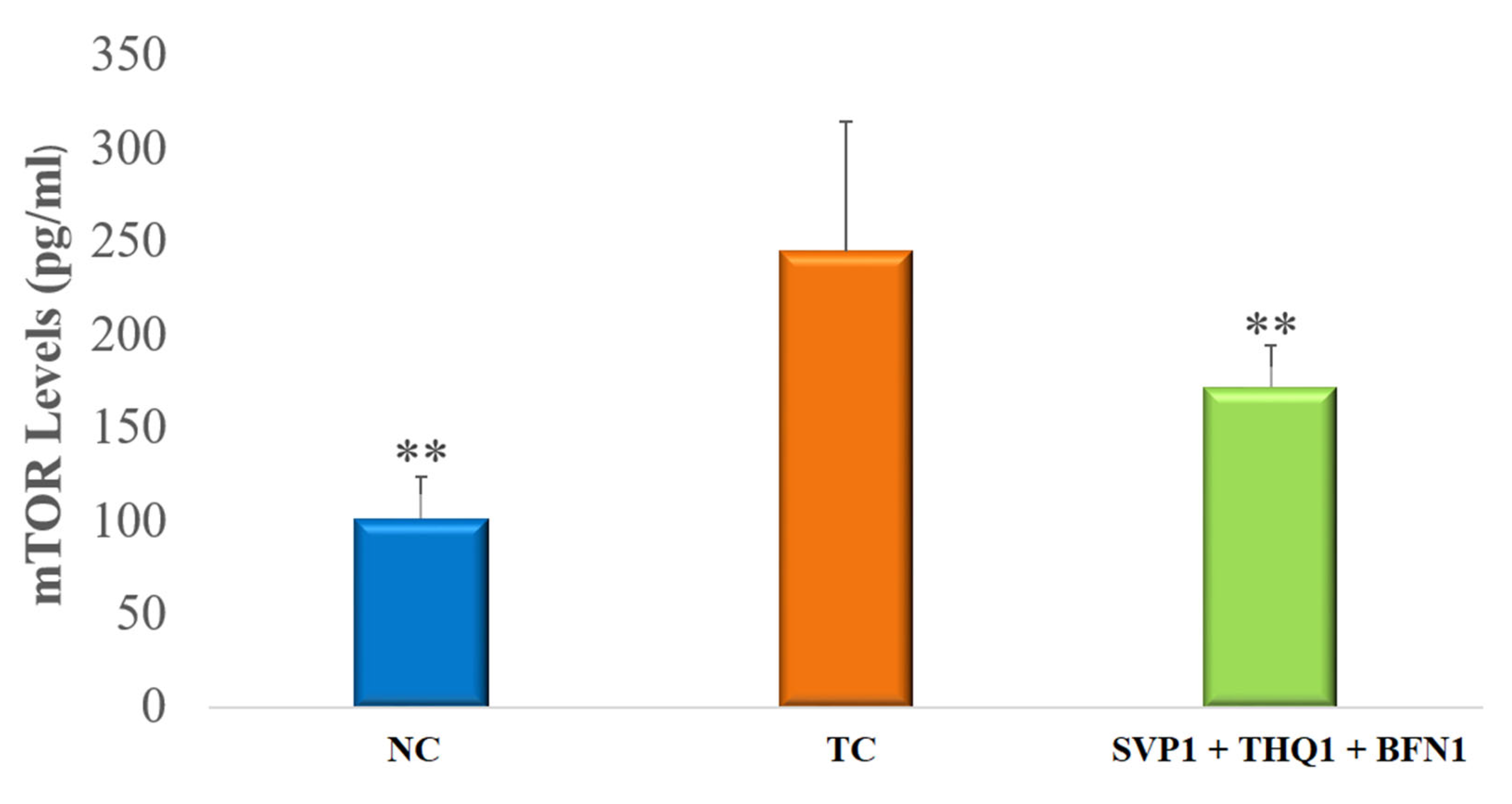
3.2. Hippocampal mTOR Levels

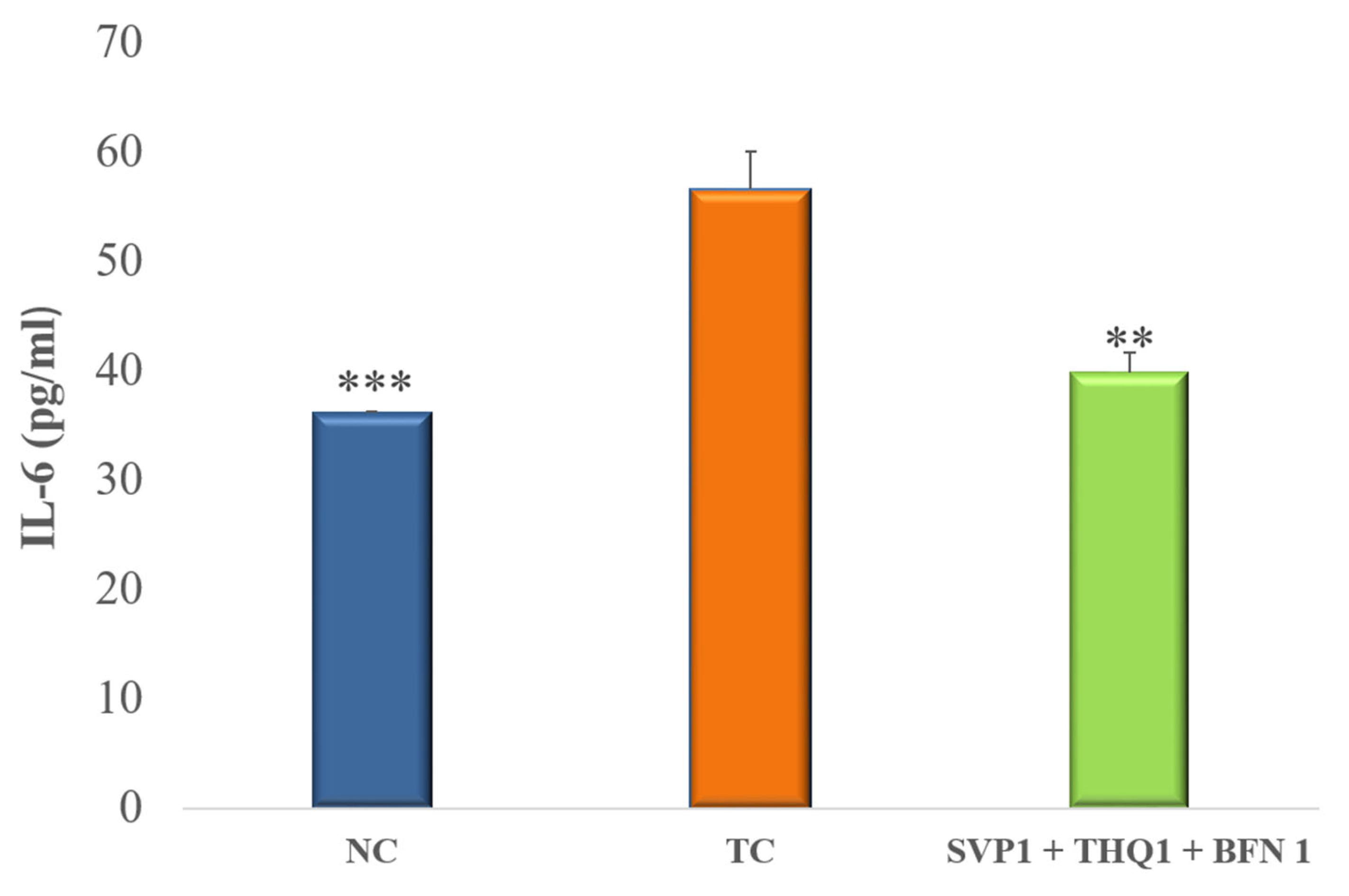
3.3. Pro-Inflammatory Cytokines


3.4. Neuroprotective Effect of Trio-Drug Combination of Sodium Valproate (SVP), Baclofen (BFN), Thymoquinone (THQ) on Rat Hippocampus


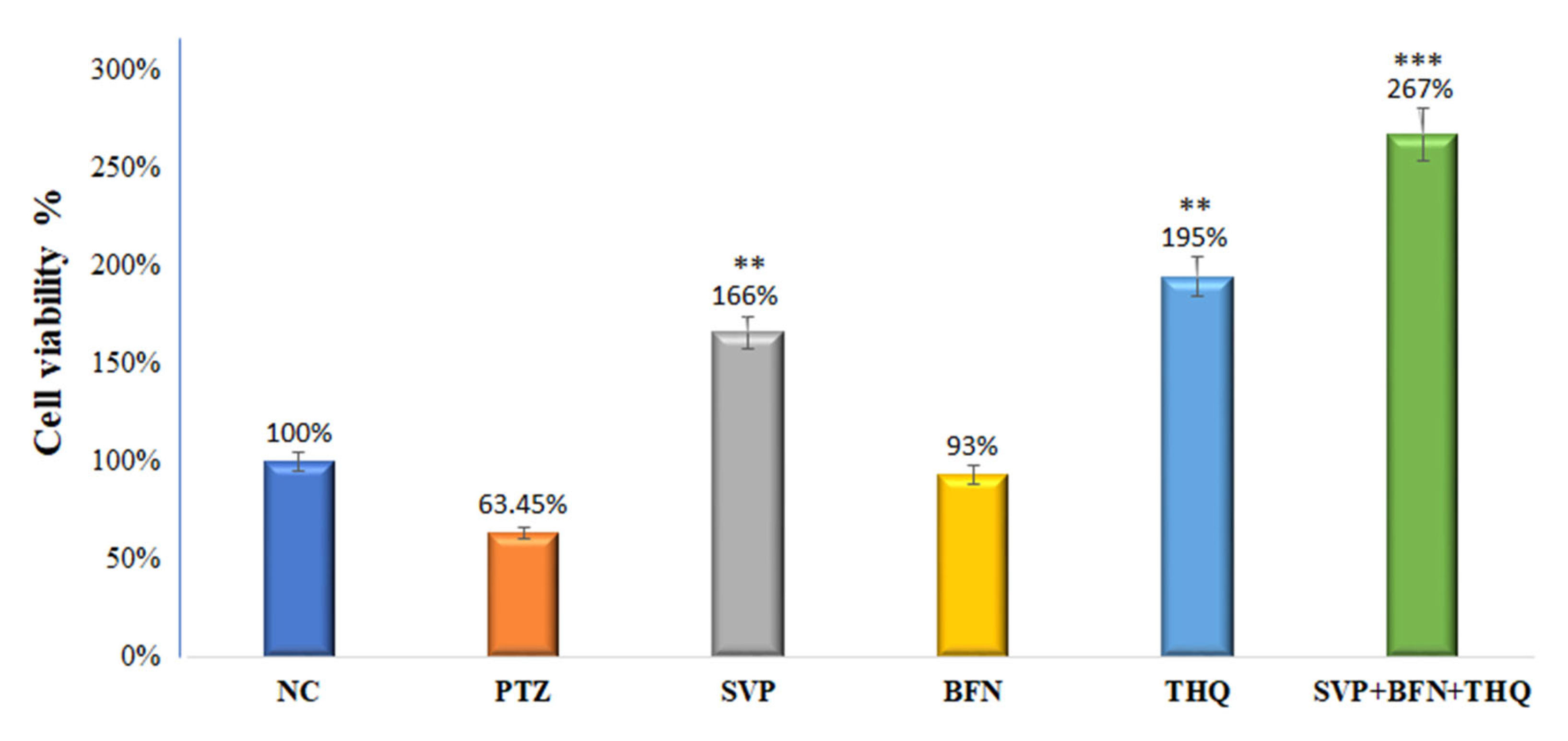
3.5. Effect of Sodium Valproate (SVP), Baclofen (BFN), Thymoquinone (THQ) Alone and in Combination on HEK-293 Cell Viability and Cell Morphology—Invitro

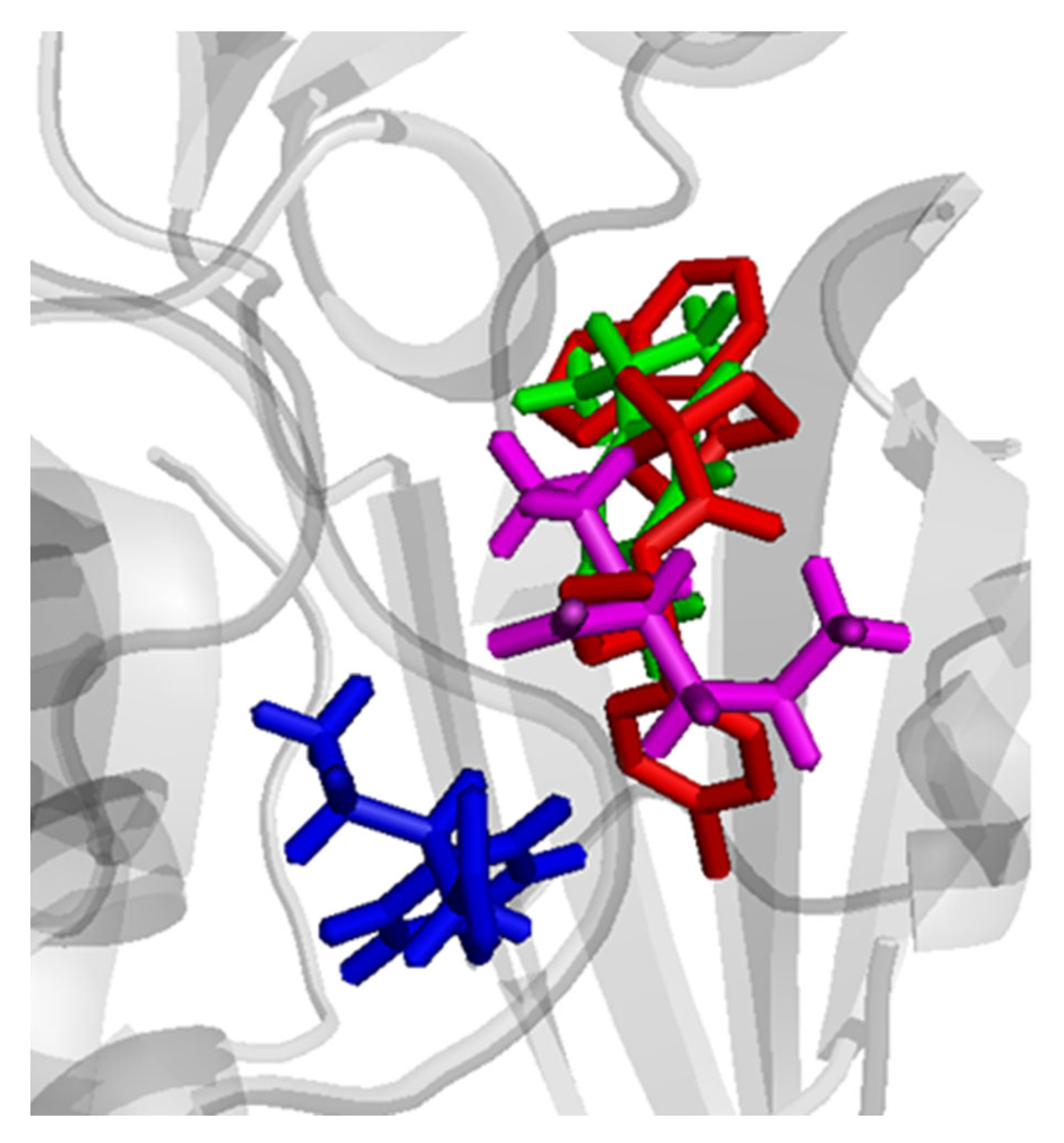
3.6. Molecular Simulation Studies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stafstrom, C.E.; Carmant, L. Seizures and Epilepsy: An Overview for Neuroscientists. Cold Spring Harb. Perspect. Med. 2015, 5, a022426. [Google Scholar] [CrossRef] [PubMed]
- Nakken, E.I.; Grinde, F.; Vaaler, A.; Drange, O.K.; Brodtkorb, E.; Sæther, S.G. Epilepsy and other seizure disorders in acute psychiatric inpatients. BMC Psychiatry 2021, 21, 626. [Google Scholar] [CrossRef] [PubMed]
- Vinti, V.; Dell’Isola, G.B.; Tascini, G.; Mencaroni, E.; Cara, G.D.; Striano, P.; Verrotti, A. Temporal Lobe Epilepsy and Psychiatric Comorbidity. Front. Neurol. 2021, 12, 2155. [Google Scholar] [CrossRef] [PubMed]
- Fiest, K.M.; Sauro, K.M.; Wiebe, S.; Patten, S.B.; Kwon, C.-S.; Dykeman, J.; Pringsheim, T.; Lorenzetti, D.L.; Jetté, N. Prevalence and incidence of epilepsy: A systematic review and meta-analysis of international studies. Neurology 2017, 88, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Nevitt, S.J.; Sudell, M.; Weston, J.; Smith, C.T.; Marson, A.G. Antiepileptic drug monotherapy for epilepsy: A network meta-analysis of individual participant data. Cochrane Database Syst. Rev. 2017, 2017, CD011412. [Google Scholar] [CrossRef]
- Goldenberg, M.M. Overview of Drugs Used for Epilepsy and Seizures. Pharm. Ther. 2010, 35, 392–415. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2912003/ (accessed on 16 February 2019).
- Kandratavicius, L.; Balista, P.A.; Lopes-Aguiar, C.; Ruggiero, R.N.; Umeoka, E.H.; Garcia-Cairasco, N.; Bueno-Junior, L.S.; Leite, J.P. Animal models of epilepsy: Use and limitations. Neuropsychiatr. Dis. Treat. 2014, 10, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- Leclercq, K.; Matagne, A.; Provins, L.; Klitgaard, H.; Kaminski, R.M. Pharmacological Profile of the Novel Antiepileptic Drug Candidate Padsevonil: Characterization in Rodent Seizure and Epilepsy Models. J. Pharmacol. Exp. Ther. 2020, 372, 11–20. [Google Scholar] [CrossRef]
- Rowley, H.L.; Marsden, C.A.; Martin, K.F. Differential effects of phenytoin and sodium valproate on seizure-induced changes in γ-aminobutyric acid and glutamate release in vivo. Eur. J. Pharmacol. 1995, 294, 541–546. [Google Scholar] [CrossRef]
- Gavzan, H.; Sayyah, M.; Sardari, S.; Babapour, V. Synergistic effect of docosahexaenoic acid on anticonvulsant activity of valproic acid and lamotrigine in animal seizure models. Naunyn Schmiedebergs Arch. Pharmacol. 2015, 388, 1029–1038. [Google Scholar] [CrossRef]
- Al-Rafiah, A.R.; Mehdar, K.M. Histopathological and Biochemical Assessment of Neuroprotective Effects of Sodium Valproate and Lutein on the Pilocarpine Albino Rat Model of Epilepsy. Behav. Neurol. 2021, 2021, 5549638. [Google Scholar] [CrossRef]
- Łuszczki, J.J.; Marzeda, P.; Gut-Lepiech, A.; Kondrat-Wróbel, M.W.; Wróblewska-Łuczka, P.; Karwan, S.; Plech, T. New derivative of 1,2,4-triazole-3-thione (TP427) potentiates the anticonvulsant action of valproate, but not that of carbamazepine, phenytoin or phenobarbital in the mouse tonic-clonic seizure model. Pharmacol. Rep. 2019, 71, 299–305. [Google Scholar] [CrossRef]
- Karpova, M.N.; Kuznetsova, L.V.; Zin’kovskii, K.A.; Klishina, N.V. Anticonvulsant Effects of Combined Treatment with Citicoline and Valproate on the Model of Acute Generalized Convulsions Induced by Pentylenetetrazole in Wistar Rats. Bull. Exp. Biol. Med. 2016, 160, 429–431. [Google Scholar] [CrossRef]
- Shakya, R.; Hoque, M.K.; Sapkota, A.S.; Gupta, P.K. Differential Hepatotoxic Effects of Sodium Valproate at Different Doses in Albino Rats. Kathmandu Univ. Med. J. (KUMJ) 2018, 16, 78–82. [Google Scholar] [CrossRef]
- Salim, S.A.; Thomas, L.; Achanti, A.; Gööz, M.B.; Castaneda, J.; Arany, I.; Dreisbach, A.W.; Fülöp, T. Baclofen-induced neurotoxicity in patients with compromised renal function: Review. Int. J. Clin. Pharmacol. Ther. 2018, 56, 467–475. [Google Scholar] [CrossRef]
- Ogata, N.; Matsuo, T.; Inoue, M. Potent depressant action of baclofen on hippocampal epileptiform activity in vitro: Possible use in the treatment of epilepsy. Brain Res. 1986, 377, 362–367. [Google Scholar] [CrossRef]
- Watts, A.E.; Jefferys, J.G. Effects of carbamazepine and baclofen on 4-aminopyridine-induced epileptic activity in rat hippocampal slices. Br. J. Pharmacol. 1993, 108, 819–823. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1908042/ (accessed on 9 January 2022). [CrossRef]
- Sato, K.; Morimoto, K.; Okamoto, M.; Nakamura, Y.; Otsuki, S.; Sato, M. An analysis of anticonvulsant actions of GABA agonists (progabide and baclofen) in the kindling model of epilepsy. Epilepsy Res. 1990, 5, 117–124. [Google Scholar] [CrossRef]
- Velíšková, J.; Velíšek, L.; Moshé, S.L. Age-Specific Effects of Baclofen on Pentylenetetrazol-Induced Seizures in Developing Rats. Epilepsia 1996, 37, 718–722. [Google Scholar] [CrossRef]
- Duthey, B.; Hübner, A.; Diehl, S.; Boehncke, S.; Pfeffer, J.; Boehncke, W.-H. Anti-inflammatory effects of the GABA(B) receptor agonist baclofen in allergic contact dermatitis. Exp. Dermatol. 2010, 19, 661–666. [Google Scholar] [CrossRef]
- Kuhn, S.A.; van Landeghem, F.K.H.; Zacharias, R.; Färber, K.; Rappert, A.; Pavlovic, S.; Hoffmann, A.; Nolte, C.; Kettenmann, H. Microglia express GABA(B) receptors to modulate interleukin release. Mol. Cell Neurosci. 2004, 25, 312–322. [Google Scholar] [CrossRef]
- Lee, M.; Schwab, C.; McGeer, P.L. Astrocytes are GABAergic cells that modulate microglial activity. Glia 2011, 59, 152–165. [Google Scholar] [CrossRef]
- Kaymak, E.; Akin, A.T.; Öztürk, E.; Karabulut, D.; Kuloğlu, N.; Yakan, B. Thymoquinone has a neuroprotective effect against inflammation, oxidative stress, and endoplasmic reticulum stress in the brain cortex, medulla, and hippocampus due to doxorubicin. J. Biochem. Mol. Toxicol. 2021, 35, e22888. [Google Scholar] [CrossRef]
- Samarghandian, S.; Farkhondeh, T.; Samini, F. A Review on Possible Therapeutic Effect of Nigella sativa and Thymoquinone in Neurodegenerative Diseases. CNS Neurol. Disord. Drug Targets 2018, 17, 412–420. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Samarghandian, S.; Shahri, A.M.P.; Samini, F. The Neuroprotective Effects of Thymoquinone: A Review. Dose Response 2018, 16, 1559325818761455. [Google Scholar] [CrossRef]
- Shao, Y.-Y.; Li, B.; Huang, Y.-M.; Luo, Q.; Xie, Y.-M.; Chen, Y.-H. Thymoquinone Attenuates Brain Injury via an Anti-oxidative Pathway in a Status Epilepticus Rat Model. Transl. Neurosci. 2017, 8, 9–14. [Google Scholar] [CrossRef]
- Shao, Y.; Feng, Y.; Xie, Y.; Luo, Q.; Chen, L.; Li, B.; Chen, Y. Protective Effects of Thymoquinone Against Convulsant Activity Induced by Lithium-Pilocarpine in a model of Status Epilepticus. Neurochem. Res. 2016, 41, 3399–3406. [Google Scholar] [CrossRef]
- Ullah, I.; Badshah, H.; Naseer, M.I.; Lee, H.Y.; Kim, M.O. Thymoquinone and Vitamin C Attenuates Pentylenetetrazole-Induced Seizures Via Activation of GABAB1 Receptor in Adult Rats Cortex and Hippocampus. Neuromol. Med. 2015, 17, 35–46. [Google Scholar] [CrossRef]
- Beyazcicek, E.; Ankarali, S.; Beyazcicek, O.; Ankarali, H.; Demir, S.; Ozmerdivenli, R. Effects of thymoquinone, the major constituent of Nigella sativa seeds, on penicillin-induced epileptiform activity in rats. Neurosciences 2016, 21, 131–137. [Google Scholar] [CrossRef]
- Aboubakr, M.; Elshafae, S.M.; Abdelhiee, E.Y.; Fadl, S.E.; Soliman, A.; Abdelkader, A.; Abdel-Daim, M.M.; Bayoumi, K.A.; Baty, R.S.; Elgendy, E.; et al. Antioxidant and Anti-Inflammatory Potential of Thymoquinone and Lycopene Mitigate the Chlorpyrifos-Induced Toxic Neuropathy. Pharmaceuticals 2021, 14, 940. [Google Scholar] [CrossRef]
- Firdaus, F.; Zafeer, M.F.; Ahmad, M.; Afzal, M. Anxiolytic and anti-inflammatory role of thymoquinone in arsenic-induced hippocampal toxicity in Wistar rats. Heliyon 2018, 4, e00650. [Google Scholar] [CrossRef] [PubMed]
- Wong, M. Mammalian Target of Rapamycin (mTOR) Pathways in Neurological Diseases. Biomed. J. 2013, 36, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.; Lee, H.-K. Mammalian/mechanistic target of rapamycin (mTOR) complexes in neurodegeneration. Mol. Neurodegener. 2021, 16, 44. [Google Scholar] [CrossRef] [PubMed]
- Crino, P.B. mTOR Signaling in Epilepsy: Insights from Malformations of Cortical Development. Cold Spring Harb. Perspect. Med. 2015, 5, a022442. [Google Scholar] [CrossRef]
- Griffith, J.L.; Wong, M. The mTOR pathway in treatment of epilepsy: A clinical update. Future Neurol. 2018, 13, 49–58. [Google Scholar] [CrossRef]
- Meng, X.-F.; Yu, J.-T.; Song, J.-H.; Chi, S.; Tan, L. Role of the mTOR signaling pathway in epilepsy. J. Neurol. Sci. 2013, 332, 4–15. [Google Scholar] [CrossRef]
- Zhang, B.; McDaniel, S.S.; Rensing, N.R.; Wong, M. Vigabatrin inhibits seizures and mTOR pathway activation in a mouse model of tuberous sclerosis complex. PLoS ONE 2013, 8, e57445. [Google Scholar] [CrossRef]
- French, J.A.; Faught, E. Rational polytherapy. Epilepsia 2009, 50 (Suppl. S8), 63–68. [Google Scholar] [CrossRef]
- Louis, E.K.S. Truly “rational” polytherapy: Maximizing efficacy and minimizing drug interactions, drug load, and adverse effects. Curr. Neuropharmacol. 2009, 7, 96–105. [Google Scholar] [CrossRef]
- Leppik, I.E. Monotherapy and polypharmacy. Neurology 2000, 55, S25–S29. [Google Scholar]
- Perucca, E. Pharmacological principles as a basis for polytherapy. Acta Neurol. Scand. Suppl. 1995, 162, 31–34. [Google Scholar] [CrossRef]
- Schmidt, D. Modern management of epilepsy: Rational polytherapy. Baillieres Clin. Neurol. 1996, 5, 757–763. [Google Scholar]
- Jefferys, J.G.R. Advances in understanding basic mechanisms of epilepsy and seizures. Seizure 2010, 19, 638–646. [Google Scholar] [CrossRef]
- Fiest, K.M.; Birbeck, G.L.; Jacoby, A.; Jette, N. Stigma in epilepsy. Curr. Neurol. Neurosci. Rep. 2014, 14, 444. [Google Scholar] [CrossRef]
- Beghi, E. The Epidemiology of Epilepsy. Neuroepidemiology 2020, 54, 185–191. [Google Scholar] [CrossRef]
- Fattorusso, A.; Matricardi, S.; Mencaroni, E.; Dell’Isola, G.B.; di Cara, G.; Striano, P.; Verrotti, A. The Pharmacoresistant Epilepsy: An Overview on Existant and New Emerging Therapies. Front. Neurol. 2021, 12, 1030. [Google Scholar] [CrossRef]
- Lee, J.W.; Dworetzky, B. Rational Polytherapy with Antiepileptic Drugs. Pharmaceuticals 2010, 3, 2362–2379. [Google Scholar] [CrossRef]
- Verrotti, A.; Lattanzi, S.; Brigo, F.; Zaccara, G. Pharmacodynamic interactions of antiepileptic drugs: From bench to clinical practice. Epilepsy Behav. 2020, 104, 106939. [Google Scholar] [CrossRef]
- Brodie, M.J.; Sills, G.J. Combining antiepileptic drugs—Rational polytherapy? Seizure 2011, 20, 369–375. [Google Scholar] [CrossRef]
- Macfarlane, A.; Greenhalgh, T. Sodium valproate in pregnancy: What are the risks and should we use a shared decision-making approach? BMC Pregnancy Childbirth 2018, 18, 200. [Google Scholar] [CrossRef]
- Louis, E.S. Minimizing AED Adverse Effects: Improving Quality of Life in the Interictal State in Epilepsy Care. Curr. Neuropharmacol. 2009, 7, 106–114. [Google Scholar] [CrossRef]
- Tyurenkov, I.N.; Borodkina, L.E.; Bagmetova, V.V. Functional aspects of neuroprotective effects of new salts and compositions of baclofen in the convulsive syndrome caused by electroshock. Bull. Exp. Biol. Med. 2012, 153, 710–713. [Google Scholar] [CrossRef]
- Tyurenkov, I.N.; Borodkina, L.E.; Bagmetova, V.V.; Berestovitskaya, V.M.; Vasil’eva, O.S. Comparison of Nootropic and Neuroprotective Features of Aryl-Substituted Analogs of Gamma-Aminobutyric Acid. Bull. Exp. Biol. Med. 2016, 160, 465–469. [Google Scholar] [CrossRef]
- Swartzwelder, H.S.; Bragdon, A.C.; Sutch, C.P.; Ault, B.; Wilson, W.A. Baclofen suppresses hippocampal epileptiform activity at low concentrations without suppressing synaptic transmission. J. Pharmacol. Exp. Ther. 1986, 237, 881–887. [Google Scholar]
- Poorgholam, P.; Yaghmaei, P.; Hajebrahimi, Z. Thymoquinone recovers learning function in a rat model of Alzheimer’s disease. Avicenna J. Phytomed. 2018, 8, 188–197. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5987434/ (accessed on 11 January 2022).
- Hosseinzadeh, H.; Parvardeh, S.; Nassiri-Asl, M.; Mansouri, M.-T. Intracerebroventricular administration of thymoquinone, the major constituent of Nigella sativa seeds, suppresses epileptic seizures in rats. Med. Sci. Monit. 2005, 11, BR106–BR110. [Google Scholar]
- Pottoo, F.H.; Salahuddin, M.; Khan, F.A.; Alomar, F.; al Dhamen, M.A.; Alhashim, A.F.; Alqattan, H.H.; Gomaa, M.S.; Alomary, M.N. Thymoquinone Potentiates the Effect of Phenytoin against Electroshock-Induced Convulsions in Rats by Reducing the Hyperactivation of m-TOR Pathway and Neuroinflammation: Evidence from In Vivo, In Vitro and Computational Studies. Pharmaceuticals 2021, 14, 1132. [Google Scholar] [CrossRef]
- Raza, M.; Alghasham, A.A.; Alorainy, M.S.; El-Hadiyah, T.M. Potentiation of Valproate-induced Anticonvulsant Response by Nigella sativa Seed Constituents: The Role of GABA Receptors. Int. J. Health Sci. 2008, 2, 15–25. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3068708/ (accessed on 11 January 2022).
- Raza, M.; Alghasham, A.A.; Alorainy, M.S.; El-Hadiyah, T.M. Beneficial Interaction of Thymoquinone and Sodium Valproate in Experimental Models of Epilepsy: Reduction in Hepatotoxicity of Valproate. Scientia Pharmaceutica 2006, 74, 159–173. [Google Scholar] [CrossRef][Green Version]
- Ostendorf, A.P.; Wong, M. mTOR Inhibition in Epilepsy: Rationale and Clinical Perspectives. CNS Drugs 2015, 29, 91–99. [Google Scholar] [CrossRef]
- Orlova, K.A.; Crino, P.B. The tuberous sclerosis complex. Ann. N. Y. Acad. Sci. 2010, 1184, 87–105. [Google Scholar] [CrossRef]
- Zeng, L.-H.; Xu, L.; Gutmann, D.H.; Wong, M. Rapamycin prevents epilepsy in a mouse model of tuberous sclerosis complex. Ann. Neurol. 2008, 63, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhang, H.; Yang, J.; Wu, J.; McMahon, J.; Lin, Y.; Cao, Z.; Gruenthal, M.; Huang, Y. Pharmacological inhibition of the mammalian target of rapamycin pathway suppresses acquired epilepsy. Neurobiol. Dis. 2010, 40, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Drion, C.M.; Borm, L.E.; Kooijman, L.; Aronica, E.; Wadman, W.J.; Hartog, A.F.; van Vliet, E.A.; Gorter, J.A. Effects of rapamycin and curcumin treatment on the development of epilepsy after electrically induced status epilepticus in rats. Epilepsia 2016, 57, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Bartfai, T.; Schultzberg, M. Cytokines in neuronal cell types. Neurochem. Int. 1993, 22, 435–444. [Google Scholar] [CrossRef]
- Wong, M. mTOR as a potential treatment target for epilepsy. Future Neurol. 2012, 7, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Kai, J.; Chen, L.; Wu, M.; Dong, J.; Wang, Q.; Zeng, L.-H. Akt Inhibitor Perifosine Prevents Epileptogenesis in a Rat Model of Temporal Lobe Epilepsy. Neurosci. Bull. 2018, 34, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Ravizza, T.; Noé, F.; Zardoni, D.; Vaghi, V.; Sifringer, M.; Vezzani, A. Interleukin Converting Enzyme inhibition impairs kindling epileptogenesis in rats by blocking astrocytic IL-1beta production. Neurobiol. Dis. 2008, 31, 327–333. [Google Scholar] [CrossRef]
- Bianchi, M.E. DAMPs, PAMPs and alarmins: All we need to know about danger. J. Leukoc. Biol. 2007, 81, 1–5. [Google Scholar] [CrossRef]
- Tsan, M.-F.; Gao, B. Endogenous ligands of Toll-like receptors. J. Leukoc. Biol. 2004, 76, 514–519. [Google Scholar] [CrossRef]
- Ulevitch, R.J.; Tobias, P.S. Receptor-dependent mechanisms of cell stimulation by bacterial endotoxin. Annu. Rev. Immunol. 1995, 13, 437–457. [Google Scholar] [CrossRef]
- Vezzani, A.; Maroso, M.; Balosso, S.; Sanchez, M.-A.; Bartfai, T. IL-1 receptor/Toll-like receptor signaling in infection, inflammation, stress and neurodegeneration couples hyperexcitability and seizures. Brain Behav. Immun. 2011, 25, 1281–1289. [Google Scholar] [CrossRef]
- Choi, J.; Min, H.J.; Shin, J.-S. Increased levels of HMGB1 and pro-inflammatory cytokines in children with febrile seizures. J. Neuroinflammation 2011, 8, 135. [Google Scholar] [CrossRef]
- Friedman, A.; Dingledine, R. Molecular cascades that mediate the influence of inflammation on epilepsy. Epilepsia 2011, 52, 33–39. [Google Scholar] [CrossRef]
- Gruol, D.L. IL-6 regulation of synaptic function in the CNS. Neuropharmacology 2015, 96, 42–54. [Google Scholar] [CrossRef]
- Samuelsson, A.-M.; Jennische, E.; Hansson, H.-A.; Holmäng, A. Prenatal exposure to interleukin-6 results in inflammatory neurodegeneration in hippocampus with NMDA/GABA(A) dysregulation and impaired spatial learning. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1345–R1356. [Google Scholar] [CrossRef]
- Rana, A.; Musto, A.E. The role of inflammation in the development of epilepsy. J. Neuroinflammation 2018, 15, 144. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Crowley, T.; Cryan, J.F.; Downer, E.J.; O’Leary, O.F. Inhibiting neuroinflammation: The role and therapeutic potential of GABA in neuro-immune interactions. Brain Behav. Immun. 2016, 54, 260–277. [Google Scholar] [CrossRef]
- Garcia-Oscos, F.; Salgado, H.; Hall, S.; Thomas, F.; Farmer, G.E.; Bermeo, J.; Galindo, L.C.; Ramirez, R.D.; D’Mello, S.; Rose-John, S.; et al. The stress-induced cytokine interleukin-6 decreases the inhibition/excitation ratio in the rat temporal cortex via trans-signaling. Biol. Psychiatry 2012, 71, 574–582. [Google Scholar] [CrossRef]
- Vrinda, M.; Arun, S.; Srikumar, B.N.; Kutty, B.M.; Rao, B.S.S. Temporal lobe epilepsy-induced neurodegeneration and cognitive deficits: Implications for aging. J. Chem. Neuroanat. 2019, 95, 146–153. [Google Scholar] [CrossRef]
- Farrell, J.S.; Wolff, M.D.; Teskey, G.C. Neurodegeneration and Pathology in Epilepsy: Clinical and Basic Perspectives. Adv. Neurobiol. 2017, 15, 317–334. [Google Scholar] [CrossRef]
- Fu, J.; Peng, L.; Wang, W.; He, H.; Zeng, S.; Chen, T.C.; Chen, Y. Sodium Valproate Reduces Neuronal Apoptosis in Acute Pentylenetetrzole-Induced Seizures via Inhibiting ER Stress. Neurochem. Res. 2019, 44, 2517–2526. [Google Scholar] [CrossRef]
- Costa, C.; Martella, G.; Picconi, B.; Prosperetti, C.; Pisani, A.; di Filippo, M.; Pisani, F.; Bernardi, G.; Calabresi, P. Multiple Mechanisms Underlying the Neuroprotective Effects of Antiepileptic Drugs Against In Vitro Ischemia. Stroke 2006, 37, 1319–1326. [Google Scholar] [CrossRef]
- Hleihil, M.; Vaas, M.; Bhat, M.A.; Balakrishnan, K.; Benke, D. Sustained Baclofen-Induced Activation of GABAB Receptors After Cerebral Ischemia Restores Receptor Expression and Function and Limits Progressing Loss of Neurons. Front. Mol. Neurosci. 2021, 14, 188. [Google Scholar] [CrossRef]
- Tavakkoli, A.; Hosseinzadeh, H. Chapter 21—Nigella sativa L. and thymoquinone as neuroprotective antioxidants, In Oxidative Stress and Dietary Antioxidants in Neurological Diseases; Martin, C.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 325–341. [Google Scholar] [CrossRef]
- Xiao, X.-Y.; Zhu, Y.-X.; Bu, J.-Y.; Li, G.-W.; Zhou, J.-H.; Zhou, S.-P. Evaluation of Neuroprotective Effect of Thymoquinone Nanoformulation in the Rodent Cerebral Ischemia-Reperfusion Model. BioMed Res. Int. 2016, 2016, e2571060. [Google Scholar] [CrossRef]
- Babcock, A.M.; Everingham, A.; Paden, C.M.; Kimura, M. Baclofen is neuroprotective and prevents loss of calcium/calmodulin-dependent protein kinase II immunoreactivity in the ischemic gerbil hippocampus. J. Neurosci. Res. 2002, 67, 804–811. [Google Scholar] [CrossRef]
- Liu, L.; Li, C.; Lu, Y.; Zong, X.; Luo, C.; Sun, J.; Guo, L. Baclofen mediates neuroprotection on hippocampal CA1 pyramidal cells through the regulation of autophagy under chronic cerebral hypoperfusion. Sci. Rep. 2015, 5, 14474. [Google Scholar] [CrossRef]
- Sahak, M.K.A.; Kabir, N.; Abbas, G.; Draman, S.; Hashim, N.H.; Adli, D.S.H. The Role of Nigella sativa and Its Active Constituents in Learning and Memory. Evid. Based Complement. Altern. Med. 2016, 2016, e6075679. [Google Scholar] [CrossRef]
- Alhebshi, A.H.; Gotoh, M.; Suzuki, I. Thymoquinone protects cultured rat primary neurons against amyloid β-induced neurotoxicity. Biochem. Biophys. Res. Commun. 2013, 433, 362–367. [Google Scholar] [CrossRef]
- García-Pupo, L.; Zaldo-Castro, A.; Exarchou, V.; Tacoronte-Morales, J.E.; Pieters, L.; Berghe, W.V.; Nuñez-Figueredo, Y.; Delgado-Hernández, R. In Vitro Neuroprotective and Anti-Inflammatory Activities of Natural and Semi-Synthetic Spirosteroid Analogues. Molecules 2016, 21, 992. [Google Scholar] [CrossRef] [PubMed]
- Hening, P.; Auriva, M.B.M.; Wijayanti, N.; Kusindarta, D.L.; Wihadmadyatami, H. The neuroprotective effect of Ocimum sanctum Linn. ethanolic extract on human embryonic kidney-293 cells as in vitro model of neurodegenerative disease. Vet. World 2018, 11, 1237–1243. [Google Scholar] [CrossRef] [PubMed]
- Shivapriya, S.; Ilango, K.; Dubey, G.P. Evaluation of antioxidant and neuroprotective effect of Hippophae rhamnoides (L.) on oxidative stress induced cytotoxicity in human neural cell line IMR32. Saudi J. Biol. Sci. 2015, 22, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-C.; Boone, M.; Meuris, L.; Lemmens, I.; van Roy, N.; Soete, A.; Reumers, J.; Moisse, M.; Plaisance, S.; Drmanac, R.; et al. Genome dynamics of the human embryonic kidney 293 lineage in response to cell biology manipulations. Nat. Commun. 2014, 5, 4767. [Google Scholar] [CrossRef]
- Thomas, P.; Smart, T.G. HEK293 cell line: A vehicle for the expression of recombinant proteins. J. Pharmacol. Toxicol. Methods 2005, 51, 187–200. [Google Scholar] [CrossRef]
- Shaw, G.; Morse, S.; Ararat, M.; Graham, F.L. Preferential transformation of human neuronal cells by human adenoviruses and the origin of HEK 293 cells. FASEB J. 2002, 16, 869–871. [Google Scholar] [CrossRef]
- Wang, L.; Jiang, J.; Zhou, T.; Xue, X.; Cao, Y. Improvement of Cerebral Ischemia-Reperfusion Injury via Regulation of Apoptosis by Exosomes Derived from BDNF-Overexpressing HEK293. Biomed. Res. Int. 2021, 2021, 6613510. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyder Pottoo, F.; Salahuddin, M.; Khan, F.A.; Albaqshi, B.T.; Gomaa, M.S.; Abdulla, F.S.; AlHajri, N.; Alomary, M.N. Trio-Drug Combination of Sodium Valproate, Baclofen and Thymoquinone Exhibits Synergistic Anticonvulsant Effects in Rats and Neuro-Protective Effects in HEK-293 Cells. Curr. Issues Mol. Biol. 2022, 44, 4350-4366. https://doi.org/10.3390/cimb44100299
Hyder Pottoo F, Salahuddin M, Khan FA, Albaqshi BT, Gomaa MS, Abdulla FS, AlHajri N, Alomary MN. Trio-Drug Combination of Sodium Valproate, Baclofen and Thymoquinone Exhibits Synergistic Anticonvulsant Effects in Rats and Neuro-Protective Effects in HEK-293 Cells. Current Issues in Molecular Biology. 2022; 44(10):4350-4366. https://doi.org/10.3390/cimb44100299
Chicago/Turabian StyleHyder Pottoo, Faheem, Mohammed Salahuddin, Firdos Alam Khan, Batool Taleb Albaqshi, Mohamed S. Gomaa, Fatima S. Abdulla, Noora AlHajri, and Mohammad N. Alomary. 2022. "Trio-Drug Combination of Sodium Valproate, Baclofen and Thymoquinone Exhibits Synergistic Anticonvulsant Effects in Rats and Neuro-Protective Effects in HEK-293 Cells" Current Issues in Molecular Biology 44, no. 10: 4350-4366. https://doi.org/10.3390/cimb44100299
APA StyleHyder Pottoo, F., Salahuddin, M., Khan, F. A., Albaqshi, B. T., Gomaa, M. S., Abdulla, F. S., AlHajri, N., & Alomary, M. N. (2022). Trio-Drug Combination of Sodium Valproate, Baclofen and Thymoquinone Exhibits Synergistic Anticonvulsant Effects in Rats and Neuro-Protective Effects in HEK-293 Cells. Current Issues in Molecular Biology, 44(10), 4350-4366. https://doi.org/10.3390/cimb44100299