

Anti-Inflammatory and Hypouricemic Effect of Bioactive Compounds: Molecular Evidence and Potential Application in the Management of Gout
 , , ,
, , ,  and
and 
Abstract
:1. Introduction
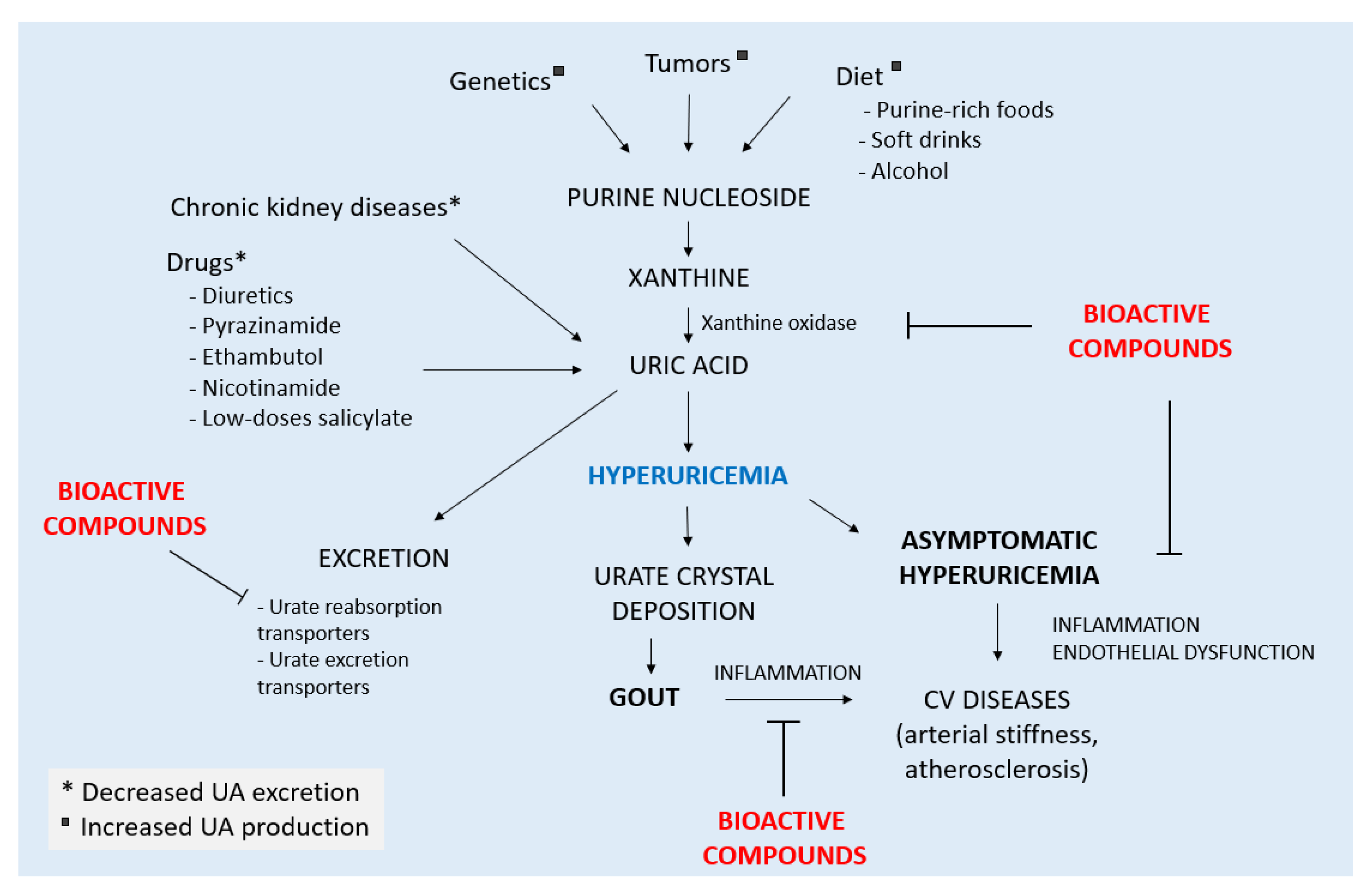
2. Gout and Hyperuricemia
2.1. The Inflammatory Process in Gout
2.2. Hyperuricemia-Associated Inflammation
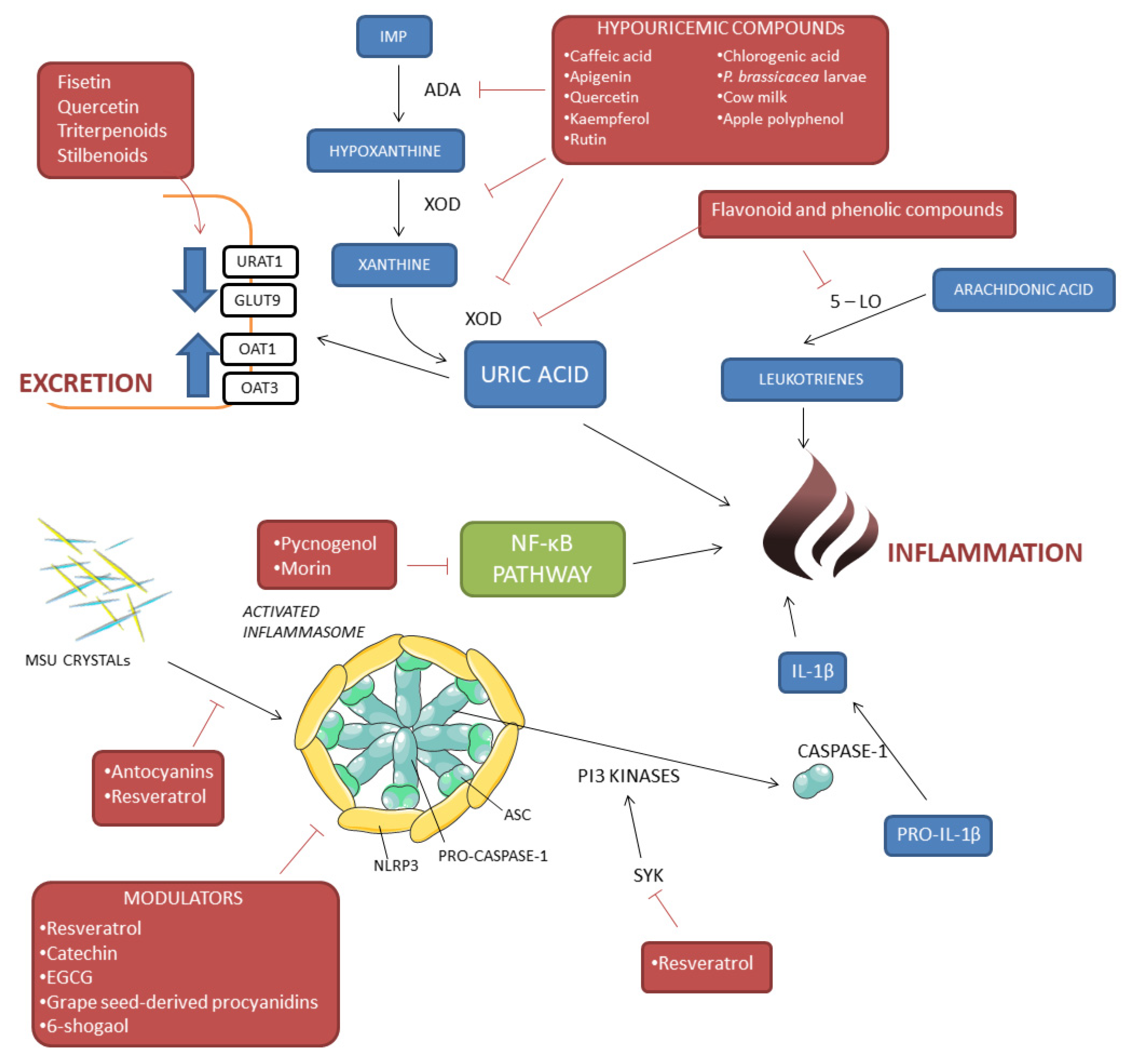
3. Beneficial Effect of Bioactive Compounds in Gout and Hyperuricemia
3.1. Hypouricemic Properties of Bioactive Compounds
3.2. Modulation of MSU Crystal-Induced Inflammation
4. Beneficial Properties of Bioactive Compounds in Animal Models of Gout
5. Clinical Studies on Bioactive Compound in Hyperuricemia and Gout
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuo, C.F.; Grainge, M.J.; Zhang, W.; Doherty, M. Global Epidemiology of Gout: Prevalence, Incidence and Risk Factors. Nat. Rev. Rheumatol. 2015, 11, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wu, Q.; Wang, H.; Zhang, S.; Jiang, Y.; Gong, T.; Xu, X.; Chang, Q.; Niu, K.; Zhao, Y. Global, Regional and National Burden of Gout, 1990–2017: A Systematic Analysis of the Global Burden of Disease Study. Rheumatology 2020, 59, 1529–1538. [Google Scholar] [CrossRef] [PubMed]
- Punzi, L.; Scanu, A.; Galozzi, P.; Luisetto, R.; Spinella, P.; Scirè, C.A.; Oliviero, F. One Year in Review 2020: Gout. Clin. Exp. Rheumatol. 2020, 38, 807–821. [Google Scholar] [PubMed]
- So, A.K.; Martinon, F. Inflammation in Gout: Mechanisms and Therapeutic Targets. Nat. Rev. Rheumatol. 2017, 13, 639–647. [Google Scholar] [CrossRef]
- Renaudin, F.; Orliaguet, L.; Castelli, F.; Fenaille, F.; Prignon, A.; Alzaid, F.; Combes, C.; Delvaux, A.; Adimy, Y.; Cohen-Solal, M.; et al. Gout and Pseudo-Gout-Related Crystals Promote GLUT1-Mediated Glycolysis That Governs NLRP3 and Interleukin-1β Activation on Macrophages. Ann. Rheum. Dis. 2020, 79, 1506–1514. [Google Scholar] [CrossRef] [PubMed]
- Borghi, C.; Agnoletti, D.; Cicero, A.F.G.; Lurbe, E.; Virdis, A. Uric Acid and Hypertension: A Review of Evidence and Future Perspectives for the Management of Cardiovascular Risk. Hypertension 2022, 79, 1927–1936. [Google Scholar] [CrossRef]
- Ponticelli, C.; Podestà, M.A.; Moroni, G. Hyperuricemia as a Trigger of Immune Response in Hypertension and Chronic Kidney Disease. Kidney Int. 2020, 98, 1149–1159. [Google Scholar] [CrossRef]
- Jayachandran, M.; Qu, S. Harnessing Hyperuricemia to Atherosclerosis and Understanding Its Mechanistic Dependence. Med. Res. Rev. 2021, 41, 616–629. [Google Scholar] [CrossRef]
- Crisan, T.O.; Cleophas, M.C.P.; Oosting, M.; Lemmers, H.; Toenhake-Dijkstra, H.; Netea, M.G.; Jansen, T.L.; Joosten, L.A.B. Soluble Uric Acid Primes TLR-Induced Proinflammatory Cytokine Production by Human Primary Cells via Inhibition of IL-1Ra. Ann. Rheum. Dis. 2016, 75, 755–762. [Google Scholar] [CrossRef]
- Biesalski, H.K.; Dragsted, L.O.; Elmadfa, I.; Grossklaus, R.; Müller, M.; Schrenk, D.; Walter, P.; Weber, P. Bioactive Compounds: Definition and Assessment of Activity. Nutrition 2009, 25, 1202–1205. [Google Scholar] [CrossRef]
- Jiang, L.L.; Gong, X.; Ji, M.Y.; Wang, C.C.; Wang, J.H.; Li, M.H. Bioactive Compounds from Plant-Based Functional Foods: A Promising Choice for the Prevention and Management of Hyperuricemia. Foods 2020, 9, 973. [Google Scholar] [CrossRef] [PubMed]
- Masuda, T.; Shingai, Y.; Takahashi, C.; Inai, M.; Miura, Y.; Honda, S.; Masuda, A. Identification of a Potent Xanthine Oxidase Inhibitor from Oxidation of Caffeic Acid. Free. Radic. Biol. Med. 2014, 69, 300–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozyel, B.; le Gall, G.; Needs, P.W.; Kroon, P.A. Anti-Inflammatory Effects of Quercetin on High-Glucose and Pro-Inflammatory Cytokine Challenged Vascular Endothelial Cell Metabolism. Mol. Nutr. Food Res. 2021, 65, e2000777. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.I.; Oyama, M.; Kondo, S.; Yagasaki, K. Comparative Effects of Quercetin, Luteolin, Apigenin and Their Related Polyphenols on Uric Acid Production in Cultured Hepatocytes and Suppression of Purine Bodies-induced Hyperuricemia by Rutin in Mice. Cytotechnology 2021, 73, 343–351. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Pan, J.; Gong, D. Novel Insights into the Inhibitory Mechanism of Kaempferol on Xanthine Oxidase. J. Agric. Food Chem. 2015, 63, 526–534. [Google Scholar] [CrossRef]
- Yoon, I.S.; Park, D.H.; Kim, J.E.; Yoo, J.C.; Bae, M.S.; Oh, D.S.; Shim, J.H.; Choi, C.Y.; An, K.W.; Kim, E.I.; et al. Identification of the Biologically Active Constituents of Camellia Japonica Leaf and Anti-Hyperuricemic Effect in Vitro and in Vivo. Int. J. Mol. Med. 2017, 39, 1613–1620. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, S.; Chiyo, A.; Fukuoka, K.; Ueda, Y.; Tokunaga, Y.; Nishida, Y.; Kinoshita, H.; Matsuda, Y.; Igoshi, K.; Ono, M.; et al. Unique Antioxidant Effects of Herbal Leaf Tea and Stem Tea from Moringa Oleifera L. Especially on Superoxide Anion Radical Generation Systems. Biosci. Biotechnol. Biochem. 2018, 82, 1973–1984. [Google Scholar] [CrossRef] [Green Version]
- Ishaq, M.; Mehmood, A.; Ur Rehman, A.; Dounya Zad, O.; Li, J.; Zhao, L.; Wang, C.; Hossen, I.; Naveed, M.; Lian, Y. Antihyperuricemic Effect of Dietary Polyphenol Sinapic Acid Commonly Present in Various Edible Food Plants. J. Food Biochem. 2020, 44, e13111. [Google Scholar] [CrossRef]
- Yuk, H.J.; Kim, J.Y.; Sung, Y.Y.; Kim, D.S. Phloroglucinol Derivatives from Dryopteris Crassirhizoma as Potent Xanthine Oxidase Inhibitors. Molecules 2020, 26, 122. [Google Scholar] [CrossRef]
- Minh, T.N.; Van, T.M.; Andriana, Y.; Vinh, L.T.; Hau, D.V.; Duyen, D.H.; de Guzman-Gelani, C. Antioxidant, Xanthine Oxidase, α-Amylase and α-Glucosidase Inhibitory Activities of Bioactive Compounds from Rumex Crispus L. Root. Molecules 2019, 24, 3899. [Google Scholar] [CrossRef]
- Spanou, C.; Veskoukis, A.S.; Kerasioti, T.; Kontou, M.; Angelis, A.; Aligiannis, N.; Skaltsounis, A.L.; Kouretas, D. Flavonoid Glycosides Isolated from Unique Legume Plant Extracts as Novel Inhibitors of Xanthine Oxidase. PLoS ONE 2012, 7, e32214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Chen, R.; Zhang, W.; Lai, X.; Sun, L.; Li, Q.; Zhang, Z.; Cao, J.; Wen, S.; Lai, Z.; et al. Tea and Its Components Reduce the Production of Uric Acid by Inhibiting Xanthine Oxidase. Food Nutr. Res. 2022, 66, 8239. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gong, Y.; Li, J.; Fan, L. In Vitro Inhibitory Effects of Polyphenols from Tartary Buckwheat on Xanthine Oxidase: Identification, Inhibitory Activity, and Action Mechanism. Food Chem. 2022, 379, 132100. [Google Scholar] [CrossRef]
- Loh, K.E.; Chin, Y.S.; Safinar Ismail, I.; Tan, H.Y. Rapid Characterisation of Xanthine Oxidase Inhibitors from the Flowers of Chrysanthemum Morifolium Ramat. Using Metabolomics Approach. Phytochem. Anal. 2022, 33, 12–22. [Google Scholar] [CrossRef]
- Li, X.; Jin, W.; Zhang, W.; Zheng, G. The Inhibitory Kinetics and Mechanism of Quercetin-3-O-Rhamnoside and Chlorogenic Acid Derived from Smilax China L. EtOAc Fraction on Xanthine Oxidase. Int. J. Biol. Macromol. 2022, 213, 447–455. [Google Scholar] [CrossRef]
- Abdulhafiz, F.; Mohammed, A.; Kayat, F.; Bhaskar, M.; Hamzah, Z.; Podapati, S.K.; Reddy, L.V. Xanthine Oxidase Inhibitory Activity, Chemical Composition, Antioxidant Properties and GC-MS Analysis of Keladi Candik (Alocasia Longiloba Miq). Molecules 2020, 25, 2658. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, D.; Liu, J.; Li, G.; Zhang, Z.; Chen, C.; Zhang, L.; Li, J. Characterization of Xanthine Oxidase Inhibitory Activities of Phenols from Pickled Radish with Molecular Simulation. Food Chem. X 2022, 14, 100343. [Google Scholar] [CrossRef]
- Sousa, C.; Pereira, D.M.; Valentão, P.; Ferreres, F.; Pereira, J.A.; Seabra, R.M.; Andrade, P.B. Pieris Brassicae Inhibits Xanthine Oxidase. J. Agric. Food Chem. 2009, 57, 2288–2294. [Google Scholar] [CrossRef]
- Nile, S.H.; Keum, Y.S.; Nile, A.S.; Kwon, Y.D.; Kim, D.H. Potential Cow Milk Xanthine Oxidase Inhibitory and Antioxidant Activity of Selected Phenolic Acid Derivatives. J. Biochem. Mol. Toxicol. 2018, 32, 1–6. [Google Scholar] [CrossRef]
- Kohoude, M.J.; Gbaguidi, F.; Agbani, P.; Ayedoun, M.A.; Cazaux, S.; Bouajila, J. Chemical Composition and Biological Activities of Extracts and Essential Oil of Boswellia Dalzielii Leaves. Pharm. Biol. 2017, 55, 33–42. [Google Scholar] [CrossRef]
- Sato, V.H.; Chewchinda, S.; Parichatikanond, W.; Vongsak, B. In Vitro and in Vivo Evidence of Hypouricemic and Anti-Inflammatory Activities of Maclura Cochinchinensis (Lour.) Corner Heartwood Extract. J. Tradit. Complement. Med. 2020, 10, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, J.; Zhang, Y.; Gou, S.; Shi, B.; Shi, D.; Zheng, M.; Yu, M.; Liu, C. Screening Anti-gout Compounds from Phellinus Igniarius by Ultrafiltration Liquid Chromatography Mass Spectrometry Based on Evaluation of an in Vitro Method Combined with Enzymatic Reaction. J. Sep. Sci. 2021, 44, 2868–2874. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Caliceti, C.; Fogacci, F.; Giovannini, M.; Calabria, D.; Colletti, A.; Veronesi, M.; Roda, A.; Borghi, C. Effect of Apple Polyphenols on Vascular Oxidative Stress and Endothelium Function: A Translational Study. Mol. Nutr. Food Res. 2017, 61, 1700373. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Takada, T.; Saito, H.; Hirata, H.; Ota-Kontani, A.; Tsuchiya, Y.; Suzuki, H. Identification of Inhibitory Activities of Dietary Flavonoids against URAT1, a Renal Urate Re-Absorber: In Vitro Screening and Fractional Approach Focused on Rooibos Leaves. Nutrients 2022, 14, 575. [Google Scholar] [CrossRef]
- Wu, X.H.; Wang, C.Z.; Zhang, J.; Wang, S.Q.; Han, L.; Zhang, Y.W.; Yuan, C.S. Effects of Smi-laxchinoside A and Smilaxchinoside C, two steroidal glycosides from Smilax riparia, on hyperuricemia in a mouse model. Phytother. Res. 2014, 28, 1822–1828. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.Y.; Zhao, Y.L.; Xiong, D.S.; He, Y.J.; Zhou, Z.S.; Zhu, P.F.; Wang, Z.J.; Wang, Y.L.; Zhao, L.X.; Luo, X.D. Potent Antihyperuricemic Triterpenoids Based on Two Unprecedented Scaffolds from the Leaves of Alstonia Scholaris. Org. Lett. 2021, 23, 4158–4162. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Fornai, M.; Antonioli, L.; Blandizzi, C.; Calderone, V. Phytochemicals as Novel Therapeutic Strategies for Nlrp3 Inflammasome-Related Neurological, Metabolic, and Inflammatory Diseases. Int. J. Mol. Sci. 2019, 20, 2876. [Google Scholar] [CrossRef] [Green Version]
- Pezzuto, J.M. Resveratrol: Twenty Years of Growth, Development and Controversy. Biomol. Ther. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Misawa, T.; Saitoh, T.; Kozaki, T.; Park, S.; Takahama, M.; Akira, S. Resveratrol Inhibits the Acetylated α-Tubulin-Mediated Assembly of the NLRP3-Inflammasome. Int. Immunol. 2015, 27, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.H.; Kim, H.Y.; Yoon, B.R.; Kang, Y.J.; Lee, W.W. Suppression of Syk Activation by Resveratrol Inhibits MSU Crystal-Induced Inflammation in Human Monocytes. J. Mol. Med. 2019, 97, 369–383. [Google Scholar] [CrossRef]
- Lin, Y.C.; Huang, D.-Y.; Wang, J.-S.; Lin, Y.-L.; Hsieh, S.-L.; Huang, K.-C.; Lin, W.-W. Syk Is Involved in NLRP3 Inflammasome-Mediated Caspase-1 Activation through Adaptor ASC Phosphorylation and Enhanced Oligomerization. J. Leukoc. Biol. 2015, 97, 825–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, I.H.; Scoville, J.P.; Reynolds, D.J.; Simlot, R.; Duncan, P. Substituted Cyclic Imides as Potential Anti-Gout Agents. Life Sci. 1990, 46, 1923–1927. [Google Scholar] [CrossRef]
- Wu, X.H.; Wang, C.Z.; Wang, S.Q.; Mi, C.; He, Y.; Zhang, J.; Zhang, Y.W.; Anderson, S.; Yuan, C.S. Anti-Hyperuricemia Effects of Allopurinol Are Improved by Smilax Riparia, a Traditional Chinese Herbal Medicine. J. Ethnopharmacol. 2015, 162, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.J.; Bakris, G.L.; Borghi, C.; Chonchol, M.B.; Feldman, D.; Lanaspa, M.A.; Merriman, T.R.; Moe, O.W.; Mount, D.B.; Sanchez Lozada, L.G.; et al. Hyperuricemia, Acute and Chronic Kidney Disease, Hypertension, and Cardiovascular Disease: Report of a Scientific Workshop Organized by the National Kidney Foundation. Am. J. Kidney Dis. 2018, 71, 851–865. [Google Scholar] [CrossRef]
- Shi, Y.W.; Wang, C.P.; Liu, L.; Liu, Y.L.; Wang, X.; Hong, Y.; Li, Z.; Kong, L.D. Antihyperuricemic and Nephroprotective Effects of Resveratrol and Its Analogues in Hyperuricemic Mice. Mol. Nutr. Food Res. 2012, 56, 1433–1444. [Google Scholar] [CrossRef]
- Wang, C.P.; Wang, Y.; Wang, X.; Zhang, X.; Ye, J.F.; Hu, L.S.; Kong, L.D. Mulberroside A Possesses Potent Uricosuric and Nephroprotective Effects in Hyperuricemic Mice. Planta. Med. 2011, 77, 786–794. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.X.; Wang, Y.; Kong, L.D.; Yang, C.; Zhang, X. Effects of Biota Orientalis Extract and Its Flavonoid Constituents, Quercetin and Rutin on Serum Uric Acid Levels in Oxonate-Induced Mice and Xanthine Dehydrogenase and Xanthine Oxidase Activities in Mouse Liver. J. Ethnopharmacol. 2004, 93, 133–140. [Google Scholar] [CrossRef]
- Chen, G.; Tan, M.L.; Li, K.K.; Leung, P.C.; Ko, C.H. Green Tea Polyphenols Decreases Uric Acid Level through Xanthine Oxidase and Renal Urate Transporters in Hyperuricemic Mice. J. Ethnopharmacol. 2015, 175, 14–20. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Z.; Li, Y.; Yang, Y.; Li, L.; Jiang, Y.; Lin, C.; Cao, Y.; Zhou, P.; Tian, Y.; et al. Baicalein Alleviates Hyperuricemia by Promoting Uric Acid Excretion and Inhibiting Xanthine Oxidase. Phytomedicine 2021, 80, 153374. [Google Scholar] [CrossRef]
- Hou, C.W.; Lee, Y.C.; Hung, H.F.; Fu, H.W.; Jeng, K.C. Longan Seed Extract Reduces Hyperuricemia via Modulating Urate Transporters and Suppressing Xanthine Oxidase Activity. Am. J. Chin. Med. 2012, 40, 979–991. [Google Scholar] [CrossRef]
- Xu, Y.; Cao, X.; Zhao, H.; Yang, E.; Wang, Y.; Cheng, N.; Cao, W. Impact of Camellia Japonica Bee Pollen Polyphenols on Hyperuricemia and Gut Microbiota in Potassium Oxonate-Induced Mice. Nutrients 2021, 13, 2665. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, L.; Zhang, P.; Zhi, J.; Xing, R.; He, L. Lycium barbarum polysaccharides protect mice from hyperuricaemia through promoting kidney excretion of uric acid and inhibiting liver xanthine oxidase. Pharm. Biol. 2020, 58, 944–949. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, A.; Zhao, L.; Wang, C.; Hossen, I.; Rakaa, R.N.; Zhanga, H. Stevia residue extract increases intestinal uric acid excretion via interactions with intestinal urate transporters in hyperuricemic mice. Food Funct. 2019, 10, 7900. [Google Scholar] [CrossRef]
- Liang, G.; Nie, Y.; Chang, Y.; Zeng, S.; Liang, C.; Zheng, X.; Xiao, D.; Zhan, S.; Zheng, Q. Protective Effects of Rhizoma Smilacis Glabrae Extracts on Potassium Oxonate- and Monosodium Urate-Induced Hyperuricemia and Gout in Mice. Phytomedicine 2019, 59, 152772. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.T.; Chang, L.C.; Liu, C.W.; Wu, P.F. Negative Correlation between Serum Uric Acid and Kidney URAT1 MRNA Expression Caused by Resveratrol in Rats. Mol. Nutr. Food Res. 2017, 61, 1601030. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, L.; Jiang, L.; Wu, Y.; Wei, L.; Wu, X.; Xiao, S.; Liu, Y.; Gao, C.; Cai, J.; et al. Sonneratia apetala seed oil attenuates potassium oxonate/hypoxanthine-induced hyperuricemia and renal injury in mice. Food Funct. 2021, 12, 9416. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Lozada, L.G.; Soto, V.; Tapia, E.; Avila-Casado, C.; Sautin, Y.Y.; Nakagawa, T.; Franco, M.; Rodríguez-Iturbe, B.; Johnson, R.J. Role of Oxidative Stress in the Renal Abnormalities Induced by Experimental Hyperuricemia. Am. J. Physiol. Renal. Physiol. 2008, 295, F1134–F1141. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.W.; Hsieh, S.H.; Chen, J.F.; Liu, C.R.; Chen, C.W.; Huang, Y.F.; Ho, H.H. Lactobacillus Reuteri TSR332 and Lactobacillus Fermentum TSF331 Stabilize Serum Uric Acid Levels and Prevent Hyperuricemia in Rats. PeerJ 2021, 9, e11209. [Google Scholar] [CrossRef]
- Lee, Y.; Werlinger, P.; Suh, J.-W.; Cheng, J. Potential Probiotic Lacticaseibacillus Paracasei MJM60396 Prevents Hyperuricemia in a Multiple Way by Absorbing Purine, Suppressing Xanthine Oxidase and Regulating Urate Excretion in Mice. Microorganisms 2022, 10, 851. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, Y.; Gao, X.; Wu, Y.; El-Seedi, H.R.; Cao, Y.; Zhao, C. Antihyperuricemic Effect of Green Alga Ulva lactuca Ulvan through Regulating Urate Transporters. J. Agric. Food Chem. 2021, 69, 11225–11235. [Google Scholar] [CrossRef]
- Li, H.; Ou, G.; He, Y.; Ren, L.; Yang, X.; Zeng, M. Resveratrol Attenuates the MSU Crystal-Induced Inflammatory Response through the Inhibition of TAK1 Activity. Int. Immunol. 2019, 67, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Jhang, J.J.; Lu, C.C.; Yen, G.C. Epigallocatechin Gallate Inhibits Urate Crystals-Induced Peritoneal Inflammation in C57BL/6 Mice. Mol. Nutr. Food Res. 2016, 60, 2297–2303. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.; Macdonald, L.; Croll, S.D.; Reinhardt, J.; Dore, A.; Stevens, S.; Hylton, D.M.; Rudge, J.S.; Liu-Bryan, R.; Terkeltaub, R.A.; et al. Hyperalgesia, Synovitis and Multiple Biomarkers of Inflammation Are Suppressed by Interleukin 1 Inhibition in a Novel Animal Model of Gouty Arthritis. Ann. Rheum. Dis. 2009, 68, 1602–1608. [Google Scholar] [CrossRef] [Green Version]
- Pineda, C.; Fuentes-Gómez, A.J.; Hernández-Díaz, C.; Zamudio-Cuevas, Y.; Fernández-Torres, J.; López-Macay, A.; Alba-Sánchez, I.; Camacho-Galindo, J.; Ventura, L.; Gómez-Quiróz, L.E.; et al. Animal Model of Acute Gout Reproduces the Inflammatory and Ultrasonographic Joint Changes of Human Gout. Arthritis. Res. Ther. 2015, 17, 37. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.D.; Zhao, Y.L.; Sun, W.J.; He, Y.J.; Liu, Y.P.; Jin, Q.; Yang, X.W.; Luo, X.D. “kidney Tea” and Its Bioactive Secondary Metabolites for Treatment of Gout. J. Agric. Food Chem. 2020, 68, 9131–9138. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Gu, L.; Li, Q.; Ju, Y.; Zhou, X.; Hu, M.; Li, Q. Anti-Gout Effects of the Medicinal Fungus Phellinus Igniarius in Hyperuricaemia and Acute Gouty Arthritis Rat Models. Front. Pharm. 2022, 12, 801910. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Liu, B.; Wang, P.; Li, X.; Li, Y.; Zheng, X.; Tai, Y.; Wang, C.; Liu, B. Eucalyptol Alleviates Inflammation and Pain Responses in a Mouse Model of Gout Arthritis. Br. J. Pharm. 2020, 177, 2042–2057. [Google Scholar] [CrossRef]
- Santos, P.D.; Vieira, T.N.; Couto, A.C.G.; Mesquita Luiz, J.P.; Lopes Saraiva, A.L.; Borges Linhares, C.R.; Barbosa, M.F.; Justino, A.B.; Franco, R.R.; da Silva Brum, E.; et al. Stephalagine, an Aporphinic Alkaloid with Therapeutic Effects in Acute Gout Arthritis in Mice. SSRN Electron. J. 2022, 293, 115291. [Google Scholar] [CrossRef]
- Chen, H.; Zheng, S.; Wang, Y.; Zhu, H.; Liu, Q.; Xue, Y.; Qiu, J.; Zou, H.; Zhu, X. The Effect of Resveratrol on the Recurrent Attacks of Gouty Arthritis. Clin. Rheumatol. 2014, 35, 1189–1195. [Google Scholar] [CrossRef]
- de Souza, M.R.; de Paula, C.A.; Pereira De Resende, M.L.; Grabe-Guimarães, A.; de Souza Filho, J.D.; Saúde-Guimarães, D.A. Pharmacological Basis for Use of Lychnophora Trichocarpha in Gouty Arthritis: Anti-Hyperuricemic and Anti-Inflammatory Effects of Its Extract, Fraction and Constituents. J. Ethnopharmacol. 2012, 142, 845–850. [Google Scholar] [CrossRef]
- Martins de Sá Müller, C.; Coelho, G.B.; Carolina de Paula Michel Araújo, M.; Saúde-Guimarães, D.A. Lychnophora Pinaster Ethanolic Extract and Its Chemical Constituents Ameliorate Hyperuricemia and Related Inflammation. J. Ethnopharmacol. 2019, 242, 112040. [Google Scholar] [CrossRef] [PubMed]
- Iside, C.; Scafuro, M.; Nebbioso, A.; Altucci, L. SIRT1 Activation by Natural Phytochemicals: An Overview. Front. Pharm. 2020, 11, 1225. [Google Scholar] [CrossRef]
- Yang, Q.B.; He, Y.L.; Zhong, X.W.; Xie, W.G.; Zhou, J.G. Resveratrol Ameliorates Gouty Inflammation via Upregulation of Sirtuin 1 to Promote Autophagy in Gout Patients. Inflammopharmacology 2019, 27, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, F.; Zamudio-Cuevas, Y.; Belluzzi, E.; Andretto, L.; Scanu, A.; Favero, M.; Ramonda, R.; Ravagnan, G.; López-Reyes, A.; Spinella, P.; et al. Polydatin and Resveratrol Inhibit the Inflammatory Process Induced by Urate and Pyrophosphate Crystals in THP-1 Cells. Foods 2019, 8, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhang, J.J.; Lu, C.C.; Ho, C.Y.; Cheng, Y.T.; Yen, G.C. Protective Effects of Catechin against Monosodium Urate-Induced Inflammation through the Modulation of NLRP3 Inflammasome Activation. J. Agric. Food Chem. 2015, 63, 7343–7352. [Google Scholar] [CrossRef]
- Oliviero, F.; Sfriso, P.; Scanu, A.; Fiocco, U.; Spinella, P.; Punzi, L. Epigallocatechin-3-Gallate Reduces Inflammation Induced by Calcium Pyrophosphate Crystals in Vitro. Front. Pharm. 2013, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.J.; Pan, X.X.; Liu, B.Q.; Gui, X.; Hu, L.; Jiang, C.Y.; Han, Y.; Fan, Y.X.; Tang, Y.L.; Liu, W.T. Grape Seed-Derived Procyanidins Alleviate Gout Pain via NLRP3 Inflammasome Suppression. J. Neuroinflamm. 2017, 14, 74. [Google Scholar] [CrossRef] [Green Version]
- Vírgen Gen, J.J.; Guzmán-Gerónimo, R.I.; Martínez-Flores, K.; Martínez-Nava, G.A.; Fernández-Torres, J.; Zamudio-Cuevas, Y. Cherry Extracts Attenuate Inflammation and Oxidative Stress Triggered by Monosodium Urate Crystals in THP-1 Cells. J. Food Biochem. 2020, 44, e13403. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, L.; Xu, S.; Ye, C.; Chen, A. Erianin: A Direct NLRP3 Inhibitor with Remarkable Anti-Inflammatory Activity. Front. Immunol. 2021, 12, 739953. [Google Scholar] [CrossRef]
- Ho, S.C.; Chang, Y.H. Comparison of Inhibitory Capacities of 6-, 8- and 10-Gingerols/Shogaols on the Canonical NLRP3 Inflammasome-Mediated IL-1β Secretion. Molecules 2018, 23, 466. [Google Scholar] [CrossRef]
- Peng, Y.J.; Lee, C.H.; Wang, C.C.; Salter, D.M.; Lee, H.S. Pycnogenol Attenuates the Inflammatory and Nitrosative Stress on Joint Inflammation Induced by Urate Crystals. Free Radic. Biol. Med. 2012, 52, 765–774. [Google Scholar] [CrossRef]
- Dhanasekar, C.; Kalaiselvan, S.; Rasool, M. Morin, a Bioflavonoid Suppresses Monosodium Urate Crystal-Induced Inflammatory Immune Response in RAW 264.7 Macrophages through the Inhibition of Inflammatory Mediators, Intracellular ROS Levels and NF-ΚB Activation. PLoS ONE 2015, 10, e0145093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, R.; Kan, J.; Cai, H.; Hong, J.; Jin, C.; Zhang, M. In Vitro and in Vivo Ameliorative Effects of Polyphenols from Purple Potato Leaves on Renal Injury and Associated Inflammation Induced by Hyperuricemia. J. Food Biochem. 2022, 46, e14049. [Google Scholar] [CrossRef] [PubMed]
- Giordo, R.; Cossu, A.; Pasciu, V.; Hoa, P.T.; Posadino, A.M.; Pintus, G. Different Redox Response Elicited by Naturally Occurring Antioxidants in Human Endothelial Cells. Open Biochem. J. 2013, 7, 44–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasongwatana, V.; Woottisin, S.; Sriboonlue, P.; Kukongviriyapan, V. Uricosuric Effect of Roselle (Hibiscus Sabdariffa) in Normal and Renal-Stone Former Subjects. J. Ethnopharmacol. 2008, 117, 491–495. [Google Scholar] [CrossRef]
- Shi, Y.; Williamson, G. Quercetin Lowers Plasma Uric Acid in Pre-Hyperuricaemic Males: A Randomised, Double-Blinded, Placebo-Controlled, Cross-over Trial. Br. J. Nutr. 2016, 115, 800–806. [Google Scholar] [CrossRef] [Green Version]
- Kawakami, Y.; Yasuda, A.; Hayashi, M.; Akiyama, M.; Asai, T.; Hosaka, T.; Arai, H. Acute Effect of Green Tea Catechins on Uric Acid Metabolism after Alcohol Ingestion in Japanese Men. Clin. Rheumatol. 2021, 40, 2881–2888. [Google Scholar] [CrossRef]
- Hokazono, H.; Omori, T.; Yamamoto, T.; Akaoka, I.; Ono, K. Effects of a Fermented Barley Extract on Subjects with Slightly High Serum Uric Acid or Mild Hyperuricemia. Biosci. Biotechnol. Biochem. 2010, 74, 828–834. [Google Scholar] [CrossRef]
- Moriwaki, Y.; Okuda, C.; Yamamoto, A.; Ka, T.; Tsutsumi, Z.; Takahashi, S.; Yamamoto, T.; Kitadate, K.; Wakame, K. Effects of Oligonol®, an Oligomerized Polyphenol Formulated from Lychee Fruit, on Serum Concentration and Urinary Excretion of Uric Acid. J. Funct. Foods 2011, 3, 13–16. [Google Scholar] [CrossRef]
- Blau, L.W. Cherry Diet Control for Gout and Arthritis. Tex. Rep. Biol. Med. 1950, 8, 309–311. [Google Scholar]
- Bell, P.G.; Gaze, D.C.; Davison, G.W.; George, T.W.; Scotter, M.J.; Howatson, G. Montmorency Tart Cherry (Prunus cerasus L.) Concentrate Lowers Uric Acid, Independent of Plasma Cyanidin-3-O-Glucosiderutinoside. J. Funct. Foods 2014, 11, 82–90. [Google Scholar] [CrossRef]
- Jacob, R.A.; Spinozzi, G.M.; Simon, V.A.; Kelley, D.S.; Prior, R.L.; Hess-Piercet, B.; Kadert, A.A. Consumption of Cherries Lowers Plasma Urate in Healthy Women. J. Nutr. 2003, 133, 1826–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, K.R.; Coles, K.M. Consumption of 100% Tart Cherry Juice Reduces Serum Urate in Overweight and Obese Adults. Curr. Dev. Nutr. 2019, 3, nzz011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillman, A.R.; Uhranowsky, K. Acute Ingestion of Montmorency Tart Cherry Reduces Serum Uric Acid but Has No Impact on High Sensitivity C-Reactive Protein or Oxidative Capacity. Plant Foods Hum. Nutr. 2021, 76, 83–89. [Google Scholar] [CrossRef]
- Zhang, Y.; Neogi, T.; Chen, C.; Chaisson, C.; Hunter, D.J.; Choi, H.K. Cherry Consumption and Decreased Risk of Recurrent Gout Attacks. Arthritis Rheum. 2012, 64, 4004–4011. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, N.; Schlesinger, M. Previously Reported Prior Studies of Cherry Juice Concentrate for Gout Flare Prophylaxis: Comment on the Article by Zhang et al. Arthritis Rheum. 2013, 65, 1135–1136. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.A.; Green, C.; Morgan, S.; Willig, A.L.; Darnell, B.; Saag, K.G.; Weiss, R.; Cutter, G.; McGwin, G. A Randomized Internet-Based Pilot Feasibility and Planning Study of Cherry Extract and Diet Modification in Gout. J. Clin. Rheumatol. 2020, 26, 147–156. [Google Scholar] [CrossRef]
- Vu, N.; Thi Tam Nguyen, T.; Haddon Parmenter, B.; Thouas, G.A. Safety, Efficacy and Tolerability of a Combination Micronutrient and Polyherbal Preparation (GoutFighterTM) for Gout: A Single-Arm Open-Label Pilot Study. J. Complement Integr. Med. 2021, 18, 113–121. [Google Scholar] [CrossRef]
- Lamb, K.L.; Lynn, A.; Russell, J.; Barker, M.E. Effect of Tart Cherry Juice on Risk of Gout Attacks: Protocol for a Randomised Controlled Trial. BMJ Open 2020, 10, e035108. [Google Scholar] [CrossRef] [Green Version]
- Stamp, L.K.; Chapman, P.; Frampton, C.; Duffull, S.B.; Drake, J.; Zhang, Y.; Neogi, T. Lack of Effect of Tart Cherry Concentrate Dose on Serum Urate in People with Gout. Rheumatology 2020, 59, 2374–2380. [Google Scholar] [CrossRef]
- Yamanaka, H.; Taniguchi, A.; Tsuboi, H.; Kano, H.; Asami, Y. Hypouricaemic Effects of Yoghurt Containing Lactobacillus Gasseri PA-3 in Patients with Hyperuricaemia and/or Gout: A Randomised, Double-Blind, Placebo-Controlled Study. Mod. Rheumatol. 2019, 29, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Kamatani, N.; Mochii, M.; Hashimoto, M.; Sakurai, K.; Gokita, K.; Yoshihara, J.; Sekine, M. Placebo Controlled Double Blind Parallel Randomized Study to Test the Urate-Lowering Effect of Yogurt Containing Lactobacillus Gasseri PA-3 in Adult Males with Marginal Hyperuricemia. Gout Nucleic Acid Metab. 2016, 40, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Upton, R.A. Pharmacokinetic Interactions Between Theophylline and Other Medication (Part I). Clin. Pharm. 1991, 20, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Razavi, M.; Kraupp, M.; Marz, R. Allopurinol Transport in Human Erythrocytes. Biochem. Pharm. 1993, 45, 893–897. [Google Scholar] [CrossRef]
- Race, T.F.; Paes, I.C.; Faloon, W.W. Intestinal Malabsorption Induced by Oral Colchicine. Comparison with Neomycin and Cathartic Agents. Am. J. Med. Sci. 1970, 259, 32–41. [Google Scholar] [CrossRef]
- Richette, P.; Doherty, M.; Pascual, E.; Barskova, V.; Becce, F.; Castañeda-Sanabria, J.; Coyfish, M.; Guillo, S.; Jansen, T.L.; Janssens, H.; et al. 2016 Updated EULAR Evidence-Based Recommendations for the Management of Gout. Ann. Rheum. Dis. 2017, 76, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Oliviero, F.; Galozzi, P.; Scanu, A.; Galuppini, F.; Lazzarin, V.; Brocco, S.; Ravagnan, G.; Sfriso, P.; Ramonda, R.; Spinella, P.; et al. Polydatin Prevents Calcium Pyrophosphate Crystal-Induced Arthritis in Mice. Nutrients 2021, 13, 929. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Animal Model | Induction | Features | Applications | Species | Reference |
|---|---|---|---|---|---|
| HYPERURICEMIA | Oral administration of a chemical inhibitor of uricase (potassium oxonate) | Increased serum uric acid concentrations. | Study of the influence on the metabolic pathway of uric acid, xanthine oxidase, urate reabsorbtion and excretion transporters | Mouse, rat | [42,43,44,45,46,47,48,49,50,51,52,53,54] |
| Potassium oxonate and uric acid | Increased serum uric acid concentrations. | Study of the effects on urate reabsorbtion and excretion transporters | Mouse | [55] | |
| Potassium oxonate associated to purine rich diet | Increased serum uric acid concentrations | Analyze the association between dietary habits and hyperuricemia | Mouse Rat | [56,57,58,59] | |
| Oral gavage of hypoxanthine and oteracil potassium | Increased serum uric acid concentrations | Study of the influence on xanthine oxidase, urate reabsorbtion and excretion transporters | Mouse | [60] | |
| GOUT | Intraperitoneal injection of MSU crystals | Acute inflammatory process with infiltrating immune cells consisting mainly of neutrophils | Study of therapeutic and prophylactic properties. Molecular pathways. | Mouse, rat | [61,62] |
| Intrarticular injection of MSU crystals (knee, ankle) | Acute inflammatory arthritis with joint swelling, tissue inflammatory infiltrate, synovitis | Study of therapeutic and prophylactic properties. Influence on the development, and the different stages of disease. Molecular pathways. | Mouse, rat, rabbit | [54,61,63,64,65,66,67,68] | |
| Subcutaneous (paw) injection of MSU crystals | Acute inflammatory process with edema and tenosynovitis | Study of the effects on the inflammatory process. Molecular pathways. | Mouse, rat | [69,70,71] |
| Bioactive Compounds | Chemical Nature | Main Sources | Experimental Model | Main Benefits | References |
|---|---|---|---|---|---|
| Baicalein | Flavones | Scutellaria roots, thyme | PO-induced hyperuricemia in mice | ↓sUA | [49] |
| Caffeic acid | Hydroxycinnamic acid | Herbs, spices | XOD inhibition assay HUVECs | ↓XOD ↓intracellular ROS | [12,84] |
| Catechins | Flavonols | Cacao, tea | MSU crystal-activated THP-1 MSU crystals-induced peritonitis | ↓NLRP3 ↓IL-1ß ↓ROS ↓NFkB | [62,75] |
| Chlorogenic acid | Hydroxycinnamic acid | Dark chocolate, herbs, hard wheat | XOD inhibition assay HUVECs PO-induced hyperuricemia in mice | ↓XOD ↓cell apoptosis ↓ROS ↓sUA | [17,25,71,84] |
| Eucalyptol | Terpene | Eucalyptus oil, herbs, spices | MSU crystals-induced arthritis in mice | ↑TRPV-1 | [67] |
| Ferulic acid | Hydroxycinnamic acid | Cereals, fruits | HUVECs | ↓ROS | [84] |
| Morin | Flavonols | Strawberries, almonds, fig | LPS-stimulated RAW 264.7 cells MSU crystal-stimulated RAW 264.7 cells PO-induced hyperuricemia in mice | ↓XOD ↓ROS ↓NFkB ↓sUA | [31,46,82] |
| Pycnogenol | Procyanidins | Pine bark | MSU crystal-stimulated chondrocytes and synoviocytes | ↓COX2 ↓IL-8 ↓NFkB | [81] |
| Quercetin | Flavonols | Red onion, berries | HUVECs treated with high-glucose concentrations Kidney 293A cells PO-induced hyperuricemia in mice | ↓XOD, ↓ADA ↓URAT 1 ↓sUA | [13,14,34,47,71] |
| 6-Shogoal | Monomethoxybenzene | Dried ginger | LPS-stimulated THP-1 cells | ↓IL-1ß | [80] |
| Resveratrol | Stilbenes | Grapes, red wine | MSU crystal-stimulated mouse primary macrophages MSU crystal-stimulated human primary monocytes MSU crystal-stimulated PBMCs MSU-crystal-stimulated THP-1 MSU crystal-induced murine peritonitis PO-induced hyperuricemia in mice | ↓NLRP3 ↓IL-1ß ↓Syk ↓NFkB ↑Sirtuin ↓autophagy ↓TAK ↓Phago ↓sUA | [39,40,45,55,61,73,74] |
| Bioactive Compounds or Plant | Target | Study | Cases, N. | Doses | Intervention Period | Main Results | Reference |
|---|---|---|---|---|---|---|---|
| Hibiscus sabdariffa | Hyperuricemia | nRCT | 18(9t + 9c) | 1.5 g/bid | 2 weeks | ↑UA excretion | [85] |
| Quercetin | Hyperuricemia | R-Crossover | 22(14t + 9c) | 0.5 g/d | 4 weeks | ↓sUA | [45] |
| Catechins | Healthy | R-Crossover | 10t | 617 mg | 5 h | ↑UA excretion | [87] |
| Barley extract | Hyperuricemia | RCT | 111(56t + 55c) | 2 g/d | 12 weeks | ↓sUA,↑UA excretion | [88] |
| Oligonol | Healthy | nRCT | 6t | 0.6 g | 1 h | ↓UA clearance, ↓XOD activity | [89] |
| Apple extract | Overweight | RCT | 62(31t + 31c) | 0.3 g/d | 8 weeks | ↓sUA,↓ER | [33] |
| Cherry concentrate | Healthy | R-Crossover | 60t | 30–60 mL/bid | 48 h | ↓sUA,↑UA excretion | [91] |
| Cherries | Healthy | nRCT | 10t | 280 g | 5 h | ↓sUA,↑UA excretion | [92] |
| Cherry juice | Overweight, obese | R-Crossover | 26t | 240 mL/d | 4 weeks | ↓sUA | [93] |
| Cherries | Gout | Case-crossover | 633 | Not reported | 1 year | ↓Gout flares | [95] |
| Cherry juice | Gout | RCT | Not reported | Not reported | 4–6 months | ↓Gout flares | [96] |
| Cherry extract | Gout | RCT | 84(41t + 43c) | 3.6 g/d | 9 months | ↓Gout flares | [97] |
| Polyherbal preparation | Gout | nRCT | 27t | 4 tablets/d | 45 days | ↓sUA | [98] |
| Cherry juice | Gout | RCT/protocol | 120 | 30 mL/d | 12 months | Outcomes: Gout flares, sUA, UA excretion | [99] |
| Cherry concentrate | Gout | RCT | 50(25t + 25c) | 7.5, 15, 22.5, 30mL/bid | 4 weeks | ↔sUA,↓gout flares | [100] |
| Probiotic/ L.gasseri PA3 | Gout | RCT | 25(13t + 12c) | 100 g/bid | 8 weeks | ↓sUA levels | [101] |
| Probiotic/ L.gasseri PA3 | Hyperuricemia | RCT | 60(20t + 20t + 20c) | 3–30 million CFU/d | 8 weeks | ↓sUA levels | [102] |
| Classes | Bioactive Compounds | Animal Model | Outcomes | Results | Evidence in Humans |
|---|---|---|---|---|---|
| Polyphenols [45,46] | Resveratrol, Mulberroside A | PO-induced hyperuricemia | Expression of urate excretion and reabsorbtion transporters | Increase (OAT1), decrease (URAT1, GLUT9) | Potential benefit on sUA levles through increase excretion [68,86,90,91] |
| Lactobacilli [58,59] | Lactobacillus Reuteri, Lactobacillus Paracasei | PO-induced hyperuricemia + high purine diet | Serum UA, Expression of urate transporters | Increase (OAT1, OAT3), decrease (URAT1) | Potential benefit on sUA levels [100,101] |
| Polyphenols [66,70,71] | Lychnophora Trichocarpha and Pinaster Extract, Fungus Phellinus Igniarius | MSU crystals-induced arthritis | Swelling, IL-1ß, ROS, XOD activity | Decrease | No evidence |
| Alcaloids, terpenes [67,68] | Eucalyptol, Stephalagine | MSU crystals-induced arthritis | Pain | Down-regulation TRPV1, TRPA1 | No evidence |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scanu, A.; Luisetto, R.; Ramonda, R.; Spinella, P.; Sfriso, P.; Galozzi, P.; Oliviero, F. Anti-Inflammatory and Hypouricemic Effect of Bioactive Compounds: Molecular Evidence and Potential Application in the Management of Gout. Curr. Issues Mol. Biol. 2022, 44, 5173-5190. https://doi.org/10.3390/cimb44110352
Scanu A, Luisetto R, Ramonda R, Spinella P, Sfriso P, Galozzi P, Oliviero F. Anti-Inflammatory and Hypouricemic Effect of Bioactive Compounds: Molecular Evidence and Potential Application in the Management of Gout. Current Issues in Molecular Biology. 2022; 44(11):5173-5190. https://doi.org/10.3390/cimb44110352
Chicago/Turabian StyleScanu, Anna, Roberto Luisetto, Roberta Ramonda, Paolo Spinella, Paolo Sfriso, Paola Galozzi, and Francesca Oliviero. 2022. "Anti-Inflammatory and Hypouricemic Effect of Bioactive Compounds: Molecular Evidence and Potential Application in the Management of Gout" Current Issues in Molecular Biology 44, no. 11: 5173-5190. https://doi.org/10.3390/cimb44110352
APA StyleScanu, A., Luisetto, R., Ramonda, R., Spinella, P., Sfriso, P., Galozzi, P., & Oliviero, F. (2022). Anti-Inflammatory and Hypouricemic Effect of Bioactive Compounds: Molecular Evidence and Potential Application in the Management of Gout. Current Issues in Molecular Biology, 44(11), 5173-5190. https://doi.org/10.3390/cimb44110352