Abstract
Candida albicans is the chief etiological agent of candidiasis, a mycosis prevalent in individuals with acquired immunodeficiency syndrome (AIDS). In recent years, the introduction of human immunodeficiency virus (HIV) protease inhibitors (HIV-PI) has reduced the prevalence of candidiasis in these patients. Seeking new therapeutic strategies based on the perspective of drug repositioning, we evaluated the effects of two second-generation HIV-PIs, atazanavir (ATV) and darunavir (DRV), on virulence factors of C. albicans and experimental candidiasis. For this, clinical strains of C. albicans were subjected to in vitro and in vivo treatments with ATV or DRV. As a result, ATV and DRV exhibited antifungal activity against fungal cells at 512 μg/mL, reduced the viability and biomass of biofilms, and inhibited filamentation of C. albicans. In addition, these HIV-PIs downregulated the expression of SAP2 and BRC1 genes of C. albicans. In an in vivo study, prophylactic use of ATV and DRV prolonged the survival rate of Galleria mellonella larvae infected with C. albicans. Therefore, ATV and DRV showed activity against C. albicans by reducing cell growth, biofilm formation, filamentation, and expression of virulence genes. Furthermore, ATV and DRV decreased experimental candidiasis, suggesting the repurposing of HIV-PIs as antifungal treatments for C. albicans infections.
1. Introduction
Candida albicans is an opportunistic fungus that usually colonizes the oral cavity, skin, gastrointestinal, and reproductive tracts [1]. Although it is harmless in healthy individuals, C. albicans can assume a pathogenic feature with rapid proliferation and invasion in the tissues, causing a variety of diseases. C. albicans is the most common causative agent of oral, vaginal, and disseminated candidiasis [2]. The annual incidence of Candida infections is about 4 million cases worldwide, involving an average healthcare cost of USD 65,000.00 per patient [3,4]. Furthermore, candidiasis is frequent among immunodeficient individuals, with oral candidiasis indicative of untreated human immunodeficiency virus (HIV) infection [5]. However, since the introduction of the first generation of HIV protease inhibitors (HIV-PI) in highly active antiretroviral therapy (HAART), the prevalence of candidiasis in HIV-infected individuals has reduced [6].
Despite the initial controversy about the role of HIV-PIs in decreasing the prevalence of candidiasis in HIV-infected individuals, clinical studies have demonstrated that HIV-PIs can exert antifungal activity against C. albicans associated with an improvement in the patient’s immune status [2,7]. It has been suggested that HIV-PIs can influence the virulence factors of C. albicans, including adhesion [8], morphogenesis [9], ability to form biofilm [10,11], and secretion of extracellular hydrolytic enzymes, especially the secreted aspartyl protease (Saps) [10,12].
Currently, the major classes of antifungal drugs used to treat C. albicans infections are polyenes, azoles, and echinocandins. However, these treatments exhibit limitations related to toxicity, cost, and antifungal resistance, especially to azole [13]. In the search for new therapeutic strategies based on drug repositioning, the use of HIV-PIs has become an interesting approach to the treatment of candidiasis [14]. The HIV-PIs inhibit the activity of HIV-1 protease, an enzyme that cleaves viral polyproteins during the virus maturation process [15,16]. The antifungal activity of first-generation HIV-PIs, such as saquinavir, ritonavir, and amprenavir, has been demonstrated in previous studies focusing on Aspergillus candidus [17], Cryptococcus gattii, Cryptococcus neoformans [18], and C. albicans [10,12]. However, the accumulation of drug-resistance mutations stimulated the development of HIV-PIs with molecular modifications. Then, second-generation HIV-PIs have been developed to treat HIV strains resistant to other HIV-PIs, and include darunavir (DRV) and atazanavir (ATV) [15]. In the treatment of HIV infection, DRV and ATV have demonstrated efficacy, safety, reduced toxicity, and low risk of regimen failure [19,20]. However, its antifungal properties have not yet been explored. Therefore, we evaluated the antifungal activity of the HIV-PIs darunavir (DRV) and atazanavir (ATV) on virulence factors of C. albicans and experimental candidiasis in the Galleria mellonella model.
2. Material and Methods
2.1. Microorganisms and Drugs
We used two clinical strains of Candida albicans isolated from oropharyngeal candidiasis lesions of patients diagnosed with AIDS and not yet treated: C. albicans 60 (Ca60) and C. albicans 70 (Ca70) strains [21]. A reference strain of C. albicans (ATCC 18804) was also included. These strains were cultured on Sabouraud dextrose agar (SDA; Difco, Detroit, USA) at 37 °C for 24 h before each assay. The HIV protease inhibitors used were darunavir (DRV; Sigma–Aldrich, St. Louis, Missouri, USA) and atazanavir (ATV; Sigma–Aldrich). The antifungal fluconazole (FCZ; Sigma–Aldrich) was also used in the experiments. The drugs were diluted in dimethyl sulfoxide (DMSO) and stored at −20 °C.
2.2. Susceptibility of C. albicans Planktonic Cells to HIV-PIs
To determine the susceptibility of C. albicans strains to DRV, ATV, and FCZ, we used the broth microdilution method proposed by the Clinical and Laboratory Standards Institute (CLSI) document M27-A2 [22]. Concentrations ranging from 1024 to 1 μg/mL were tested. C. albicans colonies were suspended in PBS, quantified using a hemocytometer, and diluted in Roswell Park Memorial Institute (RPMI) 1640 medium (Sigma–Aldrich) to a concentration of 1 × 103 cells/mL. After the adjusted inoculum was added, the microplates were incubated at 37 °C for 48 h. The minimum inhibitory concentration (MIC) was visually determined as the lowest concentration capable of inhibiting C. albicans growth by 100%. Candida albicans ATCC 18804 was used as quality control. Sterility and growth controls were also included. The susceptibility assay was performed in duplicates.
2.3. Effect of HIV-PIs in Biofilm Formation of C. albicans
Suspension of the Ca60 and Ca70 strains were counted using a hemocytometer and adjusted to 107 viable cells/mL. Initially, 100 μL of the standardized suspension was added to 96-well microplates containing Yeast Nitrogen Peptone (YNB; Difco) with 100 mM glucose and incubated for 90 min at 37 °C with stirring at 75 rpm for the initial adhesion phase. Subsequently, the supernatant was removed, each well was washed twice with PBS to remove non-adherent cells, 200 µL of fresh YNB containing DRV, ATV, and FCZ (3x MIC) was added to the plates, and incubated for 48 h at 37 °C with shaking. The medium was removed and fresh YNB was added after 24 h. A control group treated with PBS was used for each strain. After treatment, each well was washed twice with PBS for the subsequent analysis of cell viability and total biomass.
To evaluate the effect of HIV-PIs on biofilm viability, PBS was added to each well and the biofilms were disrupted using an ultrasonic homogenizer (Sonopuls HD 2200, Bandelin Electronic, Berlin, Germany) at 50 W for 30 s. The suspension was plated on SDA and the plates were incubated at 37 °C for 48 h to determine the number of colony-forming units (CFU/mL) [23].
Biofilm biomass was quantified by crystal violet staining [24]. Following this, the biofilm was fixed with methanol (Sigma–Aldrich), stained with a crystal violet 1% solution, washed with 0.85% NaCl, and discolored with peracetic acid. The supernatant was transferred to new 96-well microplates and the absorbance was measured at 570 nm (AJX-1900 spectrophotometer, Micronal, São Paulo, Brazil). Both assays were performed in triplicates.
2.4. Effect of HIV-PIs on C. albicans Filamentation
The effects of ATV and DRV on the filamentation of Ca60 and Ca70 strains were evaluated. In a 24-well culture plate, 2 mL of deionized water was mixed with 10% fetal bovine serum (FBS) (Sigma-Aldrich) and 100 μL of standardized C. albicans suspension (107 viable cells/mL). DRV (3× MIC) and ATV (3× MIC) were also added to the experimental groups. For the control group, 50 μL of PBS was added to each well. The plates were incubated at 37 °C for 24 h (5% CO2). After incubation, 50 μL of the inoculum was transferred to glass slides with 10 previously demarcated fields on the back of the slide and observed under a light microscope at 400 × magnification in triplicates. Images were analyzed and 10 microscopic fields per slide were chosen for hyphal quantification. A score (0–5) was assigned to each field according to the number of hyphae observed as follows: 0, no hyphae; 1, 1–10 hyphae; 2, 11–20 hyphae; 3, 21–30 hyphae; 4, 31–40 hyphae; and 5, more than 41 hyphae [25].
2.5. Effect of HIV-PIs on SAP2 and BCR1 Gene Expression
Biofilms of the Ca70 strain were formed in 24-well microtiter plates, as previously described [26]. Subsequently, 1 mL TRIzol (Ambion, Carlsbad, CA, USA) was added to each well to extract RNA, according to the manufacturer’s protocol. RNA concentration was measured using a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA). Total extracted RNA (700 ng) was treated with RQ1 RNase-Free DNase (Promega Corporation, Madison, WI, USA) and transcribed to cDNA using the GoScript Reverse Transcription Mix, Random Primers (Promega Corporation), according to the manufacturer’s recommendations. Candida albicans BCR1 and SAP2 primers were used [27,28]. cDNA was amplified to determine the relative quantification of BCR1 and SAP2 expression. The RIP1 reference gene was tested in all experimental groups.
For quantitative polymerase chain reaction (qPCR) reactions, a GoTaq qPCR Master Mix kit (Promega Corporation) was used, according to the manufacturer’s recommendations. The reactions were performed in two biological replicates using the StepOnePlus Real-Time PCR System (Applied Biosystems, Foster, CA, USA). The 2−ΔΔCT method was used to calculate the gene expression levels of BCR1 and SAP2 [29].
2.6. Evaluation of HIV-PI Treatment of Experimental Infection by C. albicans in Galleria Mellonella
G. mellonella larvae were maintained in the laboratory as described by Jorjão et al. [30], and larvae weighing ~330 mg without color alterations were selected. Ten randomly selected G. mellonella larvae were used per group for all the assays. Two controls were included: one group was inoculated with PBS and the second group, as control, received no injection to evaluate general viability. The larvae were pretreated by injecting 10 µL of DRV or ATV (20 mg/kg) through the last left proleg using a Hamilton 10 μL syringe. Suspensions of Ca60 and Ca70 strains were prepared from cultures in 5 mL of YPD liquid medium and incubated at 37 °C for 18 h. Cells were then centrifuged at 2000× g for 10 min and the supernatant was discarded. Cell pellets were dissolved in PBS and homogenized. This procedure was repeated twice. The cell densities were counted using a hemocytometer and adjusted to 10⁸ cells/mL. Two hours after treatment with ATV or DRV, the larvae were infected with 106 cells/larva of C. albicans through the last right proleg (10 µL). Larvae were stored in Petri dishes at 37 °C and monitored daily for seven days to determine the survival curve.
2.7. Statistical Analysis
Student’s t-test was used to compare the CFU/mL results from the in vitro biofilm formation assay and gene expression. The scores obtained from the in vitro filamentation analysis were compared using the Kruskal–Wallis and Dunn’s tests. Percent survival and survival curves of G. mellonella were plotted and statistical analysis was performed using the log-rank test. All analyses were performed using GraphPad Prism 5.0 (GraphPad Inc., San Diego, CA, USA), and a p value ≤ 0.05 was considered significant.
3. Results
3.1. HIV-PIs Showed Antifungal Activity against C. albicans Planktonic Cells
Initially, we evaluated the susceptibility of planktonic cells to HIV-PIs and FCZ using reference and clinical strains of C. albicans. The minimum inhibitory concentration (MIC) of DRV and ATV were 512 µg/mL for C. albicans ATCC 18804, C. albicans 60 (Ca60), and C. albicans 70 (Ca70). The MIC of fluconazole (FCZ) ranged from 1 to >64 µg/mL for C. albicans strains (Table 1).

Table 1.
Minimal inhibitory concentration values (MIC; µg/mL) of atazanavir (ATV), darunavir (DRV), and fluconazole (FCZ) against C. albicans strains ATCC 18804, C. albicans 60 and C. albicans 70.
3.2. HIV-PIs Decreased Viability and Biomass of C. albicans Biofilms
Based on the susceptibility of planktonic C. albicans cells to HIV-PIs, we evaluated the effect of these drugs (10× MIC value) on biofilms formed by Ca60 and Ca70 strains. HIV-PIs reduced the viability of both Ca60 and Ca70 strains. DRV (1536 μg/mL) decreased Ca60 viability (2, 27 log) and led to complete inhibition of Ca70 growth (Figure 1A,B). ATV (1536 μg/mL) completely inhibited fungal growth in both strains (Figure 1A,B). Treatment of biofilms with HIV-PIs also reduced fungal biomass. DRV (1536 μg/mL) reduced the biomass of Ca60 and Ca70 by 81 and 67%, respectively, whereas ATV (1536 μg/mL) decreased fungal biomass by 82% for both strains (Figure 1C,D).
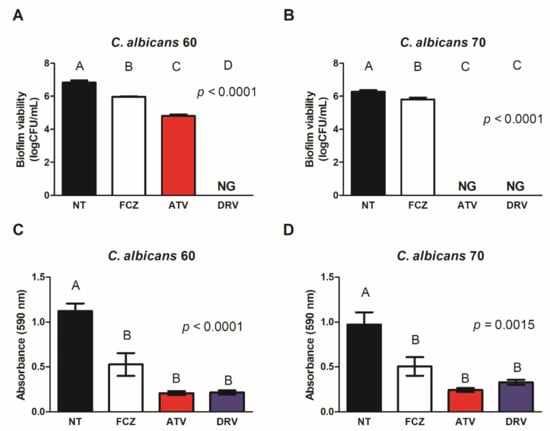
Figure 1.
ATV and DRV reduced the viability and biomass of Candida albicans biofilms. Fungal viability of biofilms of (A) C. albicans 60 (Ca60) and (B) C. albicans 70 (Ca70) not treated (NT) and treated with fluconazole (FCZ), darunavir (DRV), and atazanavir (ATV). Biofilm biomass of (C) Ca60 and (D) Ca70 without treatment (NT) and after treatment with FCZ, DRV, and ATV. Results represent the mean of three replicates. FCZ: fluconazole; DRV: darunavir; ATV: atazanavir; NT = not treated; NG = no growth. Different letters (A, B, C, D) indicate significant differences (p < 0.05).
3.3. HIV-PIs Inhibited the Filamentation of C. albicans
Since HIV-PIs reduced cell viability and biofilm biomass, we evaluated the influence of these drugs on C. albicans filamentation. ATV and DRV decreased the filamentation of both Ca60 and Ca70 strains and total inhibition of hyphal formation was observed after treatment with ATV (Figure 2).
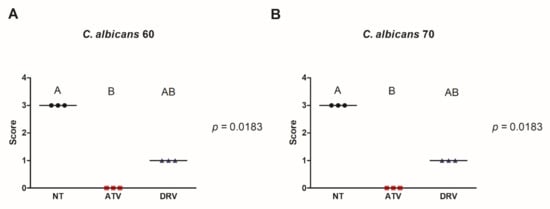
Figure 2.
ATV and DRV decreased the number of C. albicans filaments. Scores attributed for hyphae formation of (A) C. albicans 60 (Ca60) and (B) C. albicans 70 (Ca70) not treated (NT) and treated with darunavir (DRV) and atazanavir (ATV). Results represent the mean of three replicates. DRV: darunavir; ATV: atazanavir; NT = not treated. Different letters (A, B) indicate significant differences (p < 0.05).
3.4. HIV-PIs Downregulated the Expression of SAP2 and BRC1 in C. albicans
To elucidate the possible effects involved in the anti-biofilm action of HIV-PIs, we quantified the expression of two genes related to biofilm formation (BRC1) and proteinase secretion (SAP2) in a fluconazole-resistant C. albicans strain (Ca70). The expression of SAP2 was significantly downregulated by DRV treatment, while BCR1 expression showed a significant downregulation by ATV treatment (Figure 3).
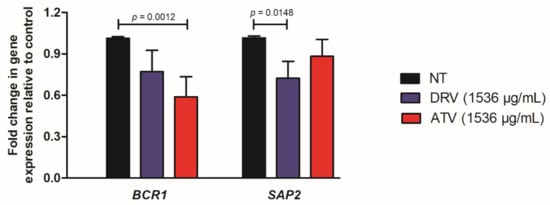
Figure 3.
Relative expression of C. albicans genes. Relative quantification of BCR1 and SAP2 genes for Ca70 biofilms non-treated (NT) and treated with darunavir (DRV) and atazanavir (ATV). Normalization was performed using the RIP1 gene. DRV and ATV treatments were compared with NT using the student’s t-test (p < 0.05).
3.5. HIV-PIs Increased the Survival of G. mellonella Infected with C. albicans
The antifungal effects of ATV and DRV determined in vitro were expanded to an in vivo model of G. mellonella. All uninfected larvae treated with PBS or HIV-PIs survived for seven days. Larvae infected with Ca60 or Ca70 strains and untreated larvae died between 24 and 48 h (Figure 4). In contrast, larvae treated with ATV and infected with the Ca60 and Ca70 strains increased the survival rate by 40 and 10%, respectively (Figure 4A,B). Concerning DRV, the treatment increased the survival of larvae infected with Ca60 by up to 40% and 20% for Ca70 (Figure 4C,D).
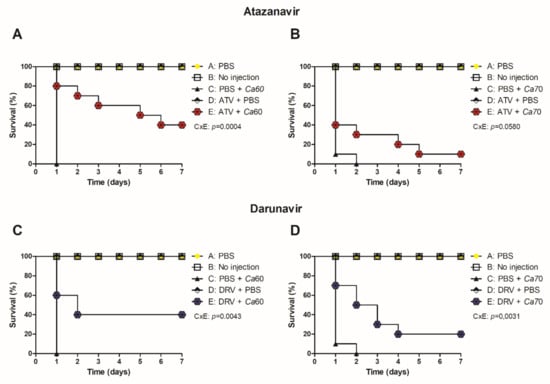
Figure 4.
ATV and DRV increased the survival of G. mellonella infected with C. albicans strains. Survival curve of uninoculated G. mellonella larvae (no injection), infected with (A,C) C. albicans 60 and (B,D) C. albicans 70 and treated with PBS, atazanavir (ATV), or darunavir (DRV). Larvae were pre-treated with ATV or DRV at the last left proleg and infected with C. albicans at the last right proleg. Ca60: C. albicans 60; Ca70: C. albicans 70; DRV: darunavir; ATV: atazanavir. p < 0.05.
4. Discussion
The development of new therapeutic strategies against Candida infections is needed because of the limited arsenal of antifungal compounds, toxic effects, elevated costs, and the emergence of fungal strains resistant to currently available drugs. However, the processes that involve prospecting, screening, and marketing new antifungals are expensive and time-consuming [31]. Therefore, drug repositioning assumes an important role when investigating new uses for existing drugs and has emerged as an interesting and promising tool against fungal infections [14,32]. Thus, HIV-PIs have gained special attention due to their antimicrobial activity in different fungal species, including C. albicans [11,18,33]. In this study, we evaluated the activity of the second-generation HIV-PIs DRV and ATV against C. albicans. To the best of our knowledge, this is the first study with a focus on DRV and ATV as a therapy for C. albicans and candidiasis.
We found that ATV and DRV exhibited antifungal activity against planktonic cells of C. albicans at 512 μg/mL. In previous studies, DRV MIC values ranging from 256 to 512 μg/mL were found for other fungal species, such as Cryptococcus neoformans and C. gattii [18]. Another second-generation HIV-PI, lopinavir, exhibited MIC values greater than 128 μg/mL for Candida auris, an emerging multi-drug resistant yeast [34]. Taken together, these studies indicated the necessity to use a high MIC value of HIV-PIs as monotherapy for inhibiting the growth of fungal cells. These data can suggest the use of HIV-PIs as adjuvants to conventional antifungal drugs. Brilhante et al. [34] demonstrated that DRV had a synergistic effect with amphotericin B against Cryptococcus species. In combined treatment, the MIC values of both DRV and amphotericin B were reduced in comparison to monotherapy. These authors hypothesized that the cellular membrane alterations caused by amphotericin B may have facilitated the penetration of a greater amount of antiretroviral in the cytoplasm of Cryptococcus cells. Therefore, future studies of HIV-PIs ATV and DRV against Candida species should be focused on their association with amphotericin B, azoles, or echinocandins.
To understand the action of ATV and DRV on fungal virulence factors, we evaluated the role of HIV-PIs in the biofilm formation and filamentation of C. albicans. Candida biofilms are important because of their ability to colonize biotic and abiotic surfaces and their reduced susceptibility to antifungal therapy. This reduced susceptibility is due to intrinsic factors in its structure, such as the difficulty of drug diffusion into the extracellular polysaccharide matrix and the expression of genes related to efflux pumps, which can interfere with the response to different drugs. The difficulty in treating biofilms requires an increase in antifungal doses, causing toxicity, which reinforces the limitations of the therapeutic arsenal available [35,36].
Here, ATV and DRV exhibited anti-biofilm effects by reducing biomass and cell viability post-treatment. Previous studies demonstrated the ability of ATV to disrupt mature C. albicans biofilms in association with caspofungin. In contrast, ATV was ineffective in preventing C. albicans biofilm formation [11]. The anti-biofilm action of HIV-PIs amprenavir and lopinavir in monotherapy was demonstrated for C. albicans [9,10]. Brilhante et al. [18] also found a decrease in the biomass and metabolic activity of Cryptococcus gattii and C. neoformans biofilms after treatment with DRV.
In this study, we also evaluated the effect of ATV and DRV treatments on the filamentation of C. albicans, another virulence factor important for fungal pathogenicity. C. albicans hyphae and pseudohyphae are invasive forms capable of penetrating the host tissue, causing tissue damage and eventually organ colonization [37]. Therefore, filamentation represents an attractive target for developing therapeutic approaches against candidiasis [38]. We demonstrated that treatment with DRV reduced and ATV completely inhibited C. albicans filamentation, reinforcing the anti-biofilm effect of these HIV-PIs. Using scanning electron microscopy, Santos et al. [9] demonstrated that although treatment with HIV-PI lopinavir did not fully inhibit C. albicans filamentation, the remaining yeast cells exhibited significant surface injuries associated with decreased fungal viability.
The reduction in C. albicans filamentation, biomass, and biofilm viability was associated with the downregulation of BRC1 and SAP2 genes observed in ATV and DRV treatment. The importance of SAP in the pathogenicity of C. albicans is well established. Infection with C. albicans mutants for the SAP1, SAP2, and SAP3 genes resulted in decreased mortality in mice [39] and reduced tissue damage in the human reconstituted epithelium [40]. Previous studies exploring the relationship between treatment with HIV-PIs and the activity of SAPs demonstrated that lopinavir, saquinavir, indinavir, and amprenavir blocked the hydrolytic activity of the Sap1, Sap2, and Sap3 enzymes [9,10,41]. In addition, indinavir, tipranavir, and ritonavir were potent inhibitors of Sap production in C. albicans culture [12,42,43], while amprenavir and indinavir reduced the expression of Sap1, Sap2, and Sap3 antigens bound to the yeast cell wall [10,44].
Thus, our study is the first to evaluate ATV and DRV action in SAP expression in C. albicans biofilms, demonstrating the inhibitory activity of these HIV-PIs on the gene encoding Sap2, an important extracellular protease in the Saps family. These results are promising since some studies have shown that SAPs play a multimodal role in the morphogenesis, adhesion, and biofilm development of C. albicans, and their inhibition directly affects fungal growth and viability. Furthermore, several studies have demonstrated the importance of SAP2 overexpression in azole-resistant C. albicans strains [10,12]. Here, we observed that DRV treatment reduced SAP2 expression in biofilms of C. albicans 70 (Ca70), a fluconazole-resistant strain.
Likewise, Ca70 biofilms treated with HIV-PIs exhibited downregulation in BRC1 expression, especially with ATV. The transcription factor bcr1 plays a prominent role in regulating C. albicans filamentation and biofilm formation [45,46,47]. C. albicans strains with mutations in the bcr1 alleles were unable to form biofilms and establish the initial adhesion phase. In addition, other studies have demonstrated the participation of this factor in cell filamentation in the opaque state [45]. Thus, ATV and DRV reduced filamentation, biofilm viability, and biomass, and downregulated the expression of genes involved in C. albicans virulence.
Based on the in vitro results, the G. mellonella model was used to evaluate the protective effects of ATV and DRV against experimental candidiasis. Interestingly, ATV and DRV increased the survival of G. mellonella compared with the untreated group. Previous studies demonstrated that the combination of lopinavir and itraconazole increased the survival of the invertebrate Caenorhabditis elegans (90%) infected with C. auris AR0390. In contrast, lopinavir did not increase larval survival in monotherapy [11]. Combination treatment with darunavir and ritonavir in HIV-positive patients elevated immune system function, characterized by a progressive increase in lymphocyte proliferation in response to Candida until reaching normal levels after 48 weeks of therapy [48]. These in vivo data reinforce the antifungal action of HIV-PIs in a manner associated with an improvement in the immune status, which may explain the increased survival of G. mellonella. Further studies are required to confirm this hypothesis.
Finally, it is important to emphasize that our results are limited to a few Candida strains analyzed. Only three C. albicans strains were investigated, including one reference strain and two clinical strains obtained from non-treated patients diagnosed with AIDS. It is known that there are genetic variations among Candida strains that implicate different profiles of virulence and susceptibility to antimicrobial agents. Therefore, further trials with other C. albicans isolates and non-albicans strains, mainly multidrug-resistant strains, are required to strengthen the anti-Candida activity of HIV-PIs.
In summary, the second-generation HIV-PIs DRV and ATV exhibited in vitro antifungal activity against C. albicans in both planktonic and biofilm stages. Furthermore, ATV and DRV reduced filamentation and the expression of virulence-related genes in C. albicans, associated with a protective effect in experimental candidiasis in G. mellonella. Importantly, HIV-PIs also showed inhibitory effects against a fluconazole-resistant strain, suggesting that ATV and DRV warrant more attention in future studies of drug repositioning strategies for Candida infections.
Author Contributions
Conceptualization and methodology, J.d.C.F., R.D.R. and J.C.J.; Validation, J.d.C.F. and P.P.d.B.; Formal analysis, J.d.C.F., P.P.d.B., P.H.F.d.C. and M.T.G.; Writing—original draft preparation, J.d.C.F., P.H.F.d.C. and M.T.G.; Data curation and writing—review and editing, J.d.C.F., P.P.d.B., P.H.F.d.C., M.T.G., R.D.R. and J.C.J.; Supervision and project administration, R.D.R. and J.C.J.; Funding acquisition, J.C.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Mello, T.P.; de Souza Ramos, L.; Braga-Silva, L.A.; Branquinha, M.H.; Dos Santos, A.L.S. Fungal Biofilm-A Real Obstacle against an Efficient Therapy: Lessons from Candida. Curr. Top. Med. Chem. 2017, 17, 1987–2004. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and Mechanisms of Antifungal Resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Wan Ismail, W.N.A.; Jasmi, N.; Khan, T.M.; Hong, Y.H.; Neoh, C.F. The Economic Burden of Candidemia and Invasive Candidiasis: A Systematic Review. Value Health Reg. Issues 2020, 21, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Bongomin, F.; Gago, S.; Oladele, R.O.; Denning, D.W. Global and Multi-National Prevalence of Fungal Diseases-Estimate Precision. J. Fungi 2017, 3, 57. [Google Scholar] [CrossRef]
- Spalanzani, R.N.; Mattos, K.; Marques, L.I.; Barros, P.F.D.; Pereira, P.I.P.; Paniago, A.M.M.; Mendes, R.P.; Chang, M.R. Clinical and Laboratorial Features of Oral Candidiasis in HIV-Positive Patients. Rev. Soc. Bras. Med. Trop. 2018, 51, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Taverne-Ghadwal, L.; Kuhns, M.; Buhl, T.; Schulze, M.H.; Mbaitolum, W.J.; Kersch, L.; Weig, M.; Bader, O.; Groß, U. Epidemiology and Prevalence of Oral Candidiasis in HIV Patients From Chad in the Post-HAART Era. Front. Microbiol. 2022, 13, 844069. [Google Scholar] [CrossRef] [PubMed]
- Selmecki, A.; Forche, A.; Berman, J. Genomic Plasticity of the Human Fungal Pathogen Candida albicans. Eukaryot. Cell 2010, 9, 991–1008. [Google Scholar] [CrossRef]
- Tsang, C.S.P.; Hong, I. HIV Protease Inhibitors Differentially Inhibit Adhesion of Candida albicans to Acrylic Surfaces. Mycoses 2010, 53, 488–494. [Google Scholar] [CrossRef]
- Santos, A.L.S.; Braga-Silva, L.A.; Gonçalves, D.S.; Ramos, L.S.; Oliveira, S.S.C.; Souza, L.O.P.; Oliveira, V.S.; Lins, R.D.; Pinto, M.R.; Muñoz, J.E.; et al. Repositioning Lopinavir, an HIV Protease Inhibitor, as a Promising Antifungal Drug: Lessons Learned from Candida albicans-In Silico, In Vitro and In Vivo Approaches. J. Fungi 2021, 7, 424. [Google Scholar] [CrossRef]
- Braga-Silva, L.A.; Mogami, S.S.V.; Valle, R.S.; Silva-Neto, I.D.; Santos, A.L.S. Multiple Effects of Amprenavir against Candida albicans. FEMS Yeast Res. 2010, 10, 221–224. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Craik, C.S.; Johnson, A.D.; Nobile, C.J. Combination of Antifungal Drugs and Protease Inhibitors Prevent Candida albicans Biofilm Formation and Disrupt Mature Biofilms. Front. Microbiol. 2020, 11, 1027. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Yang, J.; Ma, Y.; Xi, Z.; Zhao, X.; Zhao, X.; Zhao, M. The Effects of Secreted Aspartyl Proteinase Inhibitor Ritonavir on Azoles-Resistant Strains of Candida albicans as Well as Regulatory Role of SAP2 and ERG11. Immun. Inflamm. Dis. 2021, 9, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Roemer, T.; Krysan, D.J. Antifungal Drug Development: Challenges, Unmet Clinical Needs, and New Approaches. Cold Spring Harb. Perspect. Med. 2014, 4, a019703. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.; Lopez-Ribot, J.L. Screening Repurposing Libraries for Identification of Drugs with Novel Antifungal Activity. Antimicrob. Agents Chemother. 2020, 64, e00924-20. [Google Scholar] [CrossRef]
- Lv, Z.; Chu, Y.; Wang, Y. HIV Protease Inhibitors: A Review of Molecular Selectivity and Toxicity. HIV/AIDS 2015, 7, 95–104. [Google Scholar] [CrossRef]
- Berkow, E.L.; Lockhart, S.R. Fluconazole Resistance in Candida Species: A Current Perspective. Infect. Drug Resist. 2017, 10, 237–245. [Google Scholar] [CrossRef]
- Becker, A.; Sifaoui, F.; Gagneux, M.; Desprez, S.; Vignoli, P.; Huguet, D.; Froidure, M.; Leduc, D.; Legout, L. Drug Interactions between Voriconazole, Darunavir/Ritonavir and Tenofovir/Emtricitabine in an HIV-Infected Patient Treated for Aspergillus candidus Lung Abscess. Int. J. Std Aids 2015, 26, 672–675. [Google Scholar] [CrossRef]
- Brilhante, R.S.; Silva, J.A.; dos Santos Araújo, G.; Pereira, V.S.; Gotay, W.J.; de Oliveira, J.S.; de Melo Guedes, G.M.; Perei-ra-Neto, W.A.; Castelo, D.D.; de Aguiar Cordeiro, R.; et al. Darunavir Inhibits Cryptococcus Neoformans/Cryptococcus Gattii Species Complex Growth and Increases the Susceptibility of Biofilms to Antifungal Drugs. J. Med. Microbiol. 2020, 69, 830–837. [Google Scholar] [CrossRef]
- Antoniou, T.; Szadkowski, L.; Walmsley, S.; Cooper, C.; Burchell, A.N.; Bayoumi, A.M.; Montaner, J.S.G.; Loutfy, M.; Klein, M.B.; Machouf, N.; et al. Comparison of Atazanavir/Ritonavir and Darunavir/Ritonavir Based Antiretroviral Therapy for Antiretroviral Naïve Patients. BMC Infect. Dis. 2017, 17, 266. [Google Scholar] [CrossRef]
- Safety of Darunavir and Atazanavir in HIV-Infected Children in Europe and Thailand. Antivir. Ther. 2016, 21, 353–358. [CrossRef]
- Junqueira, J.C.; Vilela, S.F.G.; Rossoni, R.D.; Barbosa, J.O.; Costa, A.C.B.P.; Rasteiro, V.M.C.; Suleiman, J.M.A.H.; Jorge, A.O.C. Oral Colonization by Yeasts in HIV-Positive Patients in Brazil. Rev. Inst. Med. Trop. Sao Paulo 2012, 54, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; Approved Standard Second Edition. CLSI Document M27-A2, 4th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2002. [Google Scholar]
- Costa, A.C.B.P.; Pereira, C.A.; Freire, F.; Junqueira, J.C.; Jorge, A.O.C. Methods for Obtaining Reliable and Reproducible Results in Studies of Candida Biofilms Formed in Vitro. Mycoses 2013, 56, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, J.O.; Rossoni, R.D.; Vilela, S.F.G.; de Alvarenga, J.A.; dos Santos Velloso, M.; de Azevendo Prata, M.C.; Jorge, A.O.C.; Junqueira, J.C. Streptococcus mutans Can Modulate Biofilm Formation and Attenuate the Virulence of Candida albicans. PLoS ONE 2016, 11, e0150457. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.C.; de Barros, P.P.; Rossoni, R.D.; Junqueira, J.C.; Jorge, A.O.C. Lactobacillus rhamnosus Inhibits Candida albicans Virulence Factors in Vitro and Modulates Immune System in Galleria mellonella. J. Appl. Microbiol. 2017, 122, 201–211. [Google Scholar] [CrossRef]
- De Barros, P.P.; Freire, F.; Rossoni, R.D.; Junqueira, J.C.; Jorge, A.O.C. Candida krusei and Candida glabrata Reduce the Filamentation of Candida albicans by Downregulating Expression of HWP1 Gene. Folia Microbiol. 2017, 62, 317–323. [Google Scholar] [CrossRef]
- Nailis, H.; Kucharíková, S.; Ricicová, M.; Van Dijck, P.; Deforce, D.; Nelis, H.; Coenye, T. Real-Time PCR Expression Profiling of Genes Encoding Potential Virulence Factors in Candida albicans Biofilms: Identification of Model-Dependent and -Independent Gene Expression. BMC Microbiol. 2010, 10, 114. [Google Scholar] [CrossRef]
- Hnisz, D.; Bardet, A.F.; Nobile, C.J.; Petryshyn, A.; Glaser, W.; Schöck, U.; Stark, A.; Kuchler, K. A Histone Deacetylase Adjusts Transcription Kinetics at Coding Sequences during Candida albicans Morphogenesis. PLoS Genet. 2012, 8, e1003118. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jorjão, A.L.; Oliveira, L.D.; Scorzoni, L.; Figueiredo-Godoi, L.M.A.; Cristina, A.; Prata, M.; Jorge, A.O.C.; Junqueira, J.C. From Moths to Caterpillars: Ideal Conditions for Galleria mellonella Rearing for in Vivo Microbiological Studies. Virulence 2018, 9, 383–389. [Google Scholar] [CrossRef]
- Tillotson, J.; Tillotson, G.S. The Regulatory Pathway for Antifungal Drugs: A US Perspective. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 61 (Suppl. 6), S678–S683. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug Repurposing: Progress, Challenges and Recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Eldesouky, H.E.; Li, X.; Abutaleb, N.S.; Mohammad, H.; Seleem, M.N. Synergistic Interactions of Sulfamethoxazole and Azole Antifungal Drugs against Emerging Multidrug-Resistant Candida auris. Int. J. Antimicrob. Agents 2018, 52, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Eldesouky, H.E.; Salama, E.A.; Lanman, N.A.; Hazbun, T.R.; Seleem, M.N. Potent Synergistic Interactions between Lopinavir and Azole Antifungal Drugs against Emerging Multidrug-Resistant Candida auris. Antimicrob. Agents Chemother. 2020, 65, e00684-20. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilm Formation: A Clinically Relevant Microbiological Process. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2001, 33, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 62, e1–e50. [Google Scholar] [CrossRef]
- Sudbery, P.; Gow, N.; Berman, J. The Distinct Morphogenic States of Candida albicans. Trends Microbiol. 2004, 12, 317–324. [Google Scholar] [CrossRef]
- Vila, T.; Romo, J.A.; Pierce, C.G.; McHardy, S.F.; Saville, S.P.; Lopez-Ribot, J.L. Targeting Candida albicans Filamentation for Antifungal Drug Development. Virulence 2017, 8, 150–158. [Google Scholar] [CrossRef]
- Hube, B.; Sanglard, D.; Odds, F.C.; Hess, D.; Monod, M.; Schäfer, W.; Brown, A.J.; Gow, N.A. Disruption of Each of the Secreted Aspartyl Proteinase Genes SAP1, SAP2, and SAP3 of Candida albicans Attenuates Virulence. Infect. Immun. 1997, 65, 3529–3538. [Google Scholar] [CrossRef]
- Naglik, J.R.; Moyes, D.; Makwana, J.; Kanzaria, P.; Tsichlaki, E.; Weindl, G.; Tappuni, A.R.; Rodgers, C.A.; Woodman, A.J.; Challacombe, S.J.; et al. Quantitative Expression of the Candida albicans Secreted Aspartyl Proteinase Gene Family in Human Oral and Vaginal Candidiasis. Microbiology 2008, 154, 3266–3280. [Google Scholar] [CrossRef]
- Schaller, M.; Krnjaic, N.; Niewerth, M.; Hamm, G.; Hube, B.; Korting, H.C. Effect of Antimycotic Agents on the Activity of Aspartyl Proteinases Secreted by Candida albicans. J. Med. Microbiol. 2003, 52, 247–249. [Google Scholar] [CrossRef]
- Cassone, A.; De Bernardis, F.; Torosantucci, A.; Tacconelli, E.; Tumbarello, M.; Cauda, R. In Vitro and in Vivo Anticandidal Activity of Human Immunodeficiency Virus Protease Inhibitors. J. Infect. Dis. 1999, 180, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Cenci, E.; Francisci, D.; Belfiori, B.; Pierucci, S.; Baldelli, F.; Bistoni, F.; Vecchiarelli, A. Tipranavir Exhibits Different Effects on Opportunistic Pathogenic Fungi. J. Infect. 2008, 56, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.; Speth, C.; Lukasser-Vogl, E.; Zangerle, R.; Borg-von Zepelin, M.; Dierich, M.P.; Würzner, R. Human Immunodeficiency Virus Type 1 Protease Inhibitor Attenuates Candida albicans Virulence Properties in Vitro. Immunopharmacology 1999, 41, 227–234. [Google Scholar] [CrossRef]
- Guan, G.; Xie, J.; Tao, L.; Nobile, C.J.; Sun, Y.; Cao, C.; Tong, Y.; Huang, G. Bcr1 Plays a Central Role in the Regulation of Opaque Cell Filamentation in Candida Albicans. Mol. Microbiol. 2013, 89, 732–750. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Mitchell, A.P. Regulation of Cell-Surface Genes and Biofilm Formation by the C. albicans Transcription Factor Bcr1p. Curr. Biol. 2005, 15, 1150–1155. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.-I.; Finkel, J.S.; Solis, N.V.; Chaili, S.; Mitchell, A.P.; Yeaman, M.R.; Filler, S.G. Bcr1 Functions Downstream of Ssd1 to Mediate Antimicrobial Peptide Resistance in Candida albicans. Eukaryot. Cell 2013, 12, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Tsoukas, C.; Gilbert, L.; Lewis, T.; Hatzakis, G.; Falcon, R.; Mrus, J. Improvements in Immune Function and Activation with 48-Week Darunavir/Ritonavir-Based Therapy: GRACE Substudy. Isrn Aids 2013, 2013, 358294. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).