

Influence of Technological Stages of Preparation of Rooster Semen for Short-Term and Long-Term Storage on Its Quality Characteristics

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Endorsement
2.2. Animals
2.3. Design of Technological Stages of the Experiment
2.4. Assessment of Semen Quality in the Field
2.5. Reagents and Diluent Composition
2.6. Sperm Cryopreservation and Thawing
2.7. Quantitative Chromatographic Analysis of Spermatozoa Cytosol Carbohydrates
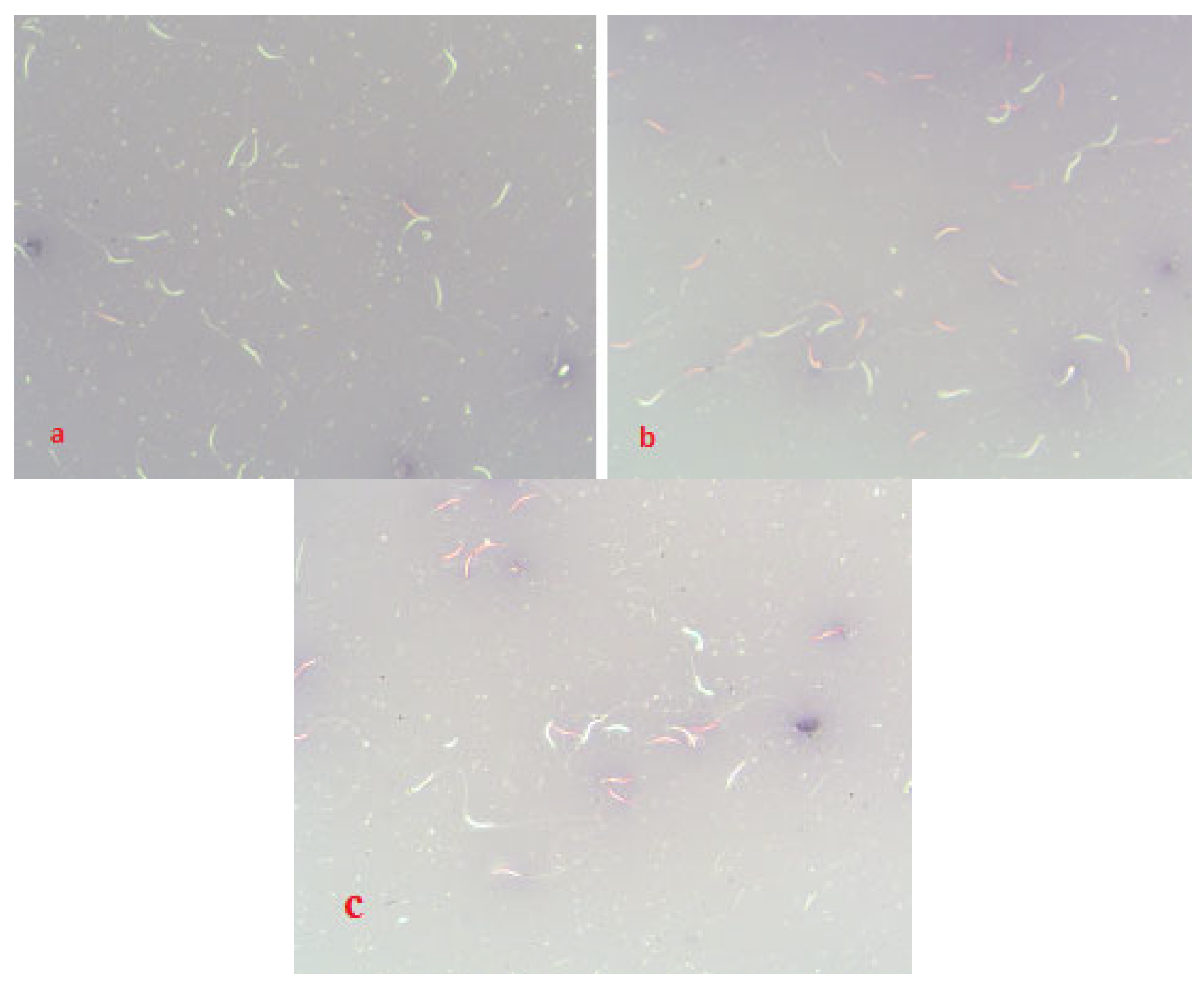
2.8. Viability of Spermatozoa of Native and Frozen/Thawed Semen
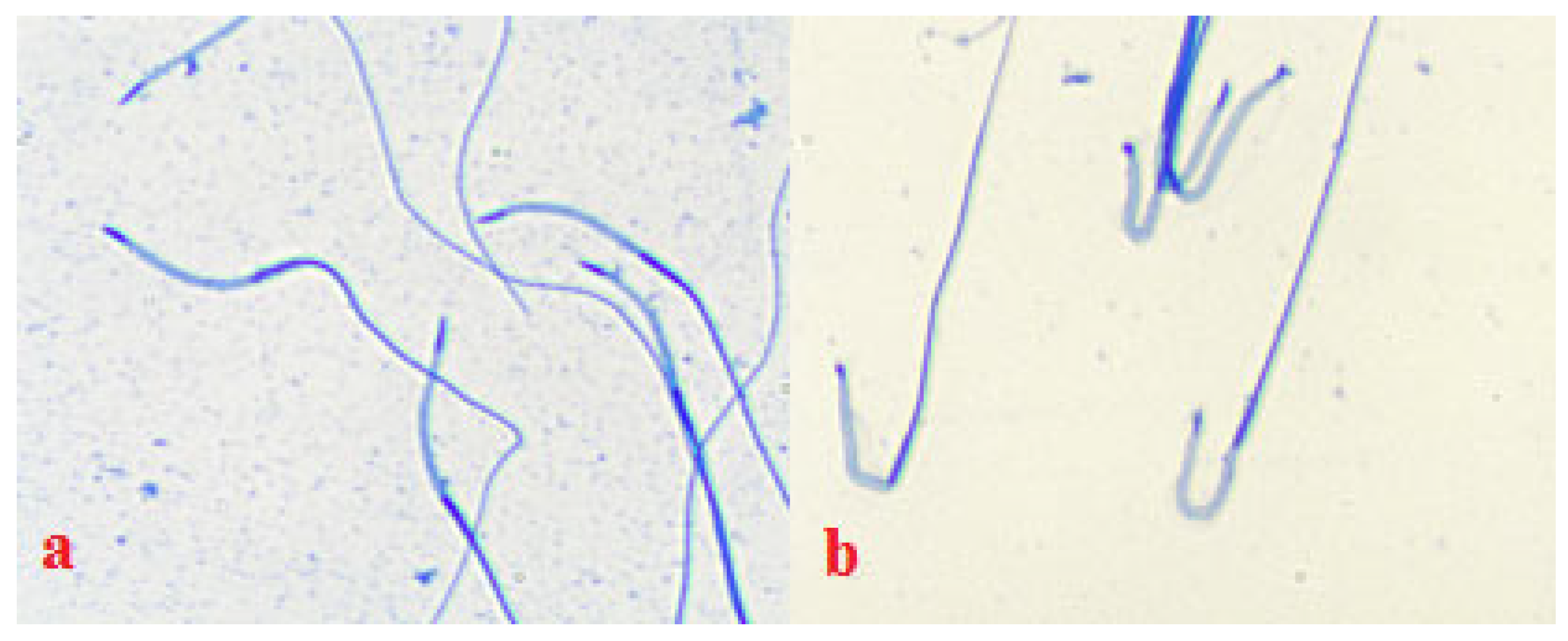
2.9. Sperm Chromatin Integrity of Native and Frozen/Thawed Semen
2.10. Integrity of Spermatozoa Acrosomes of Native and Frozen/Thawed Semen
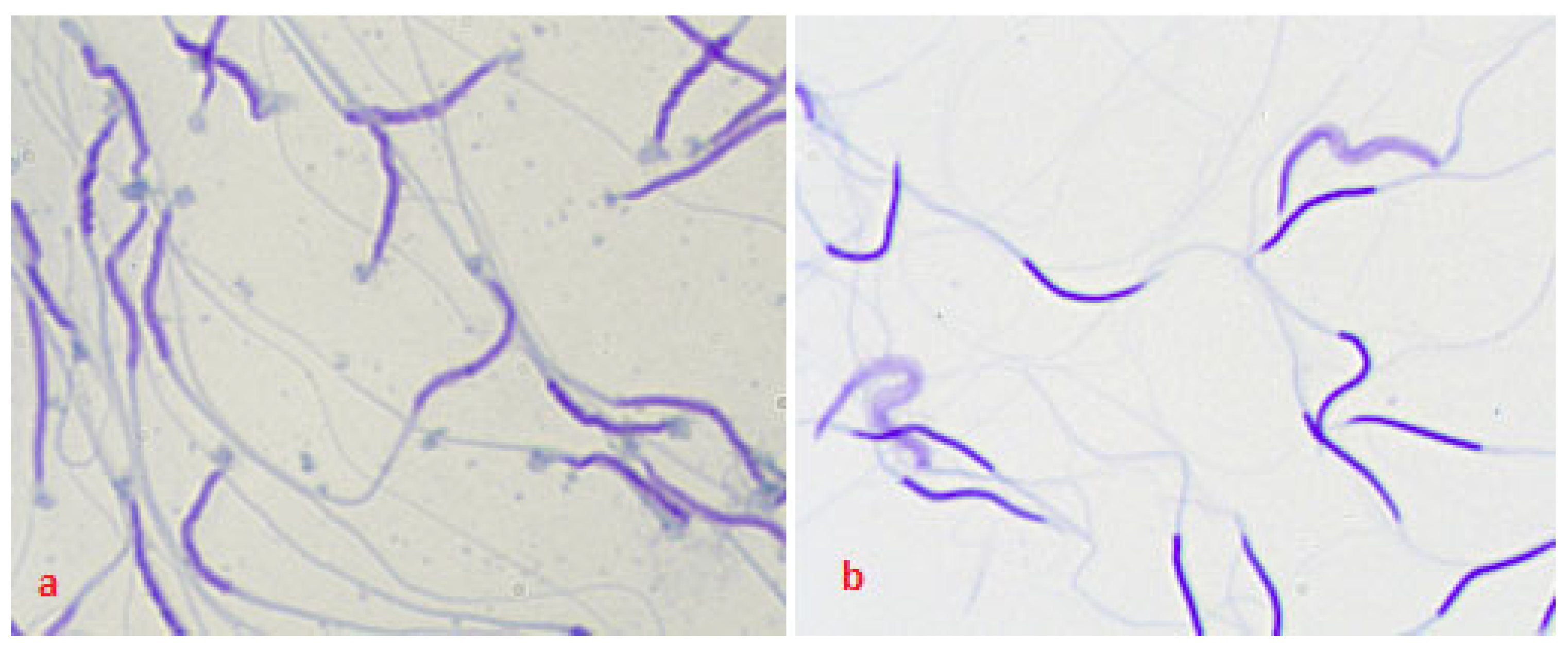
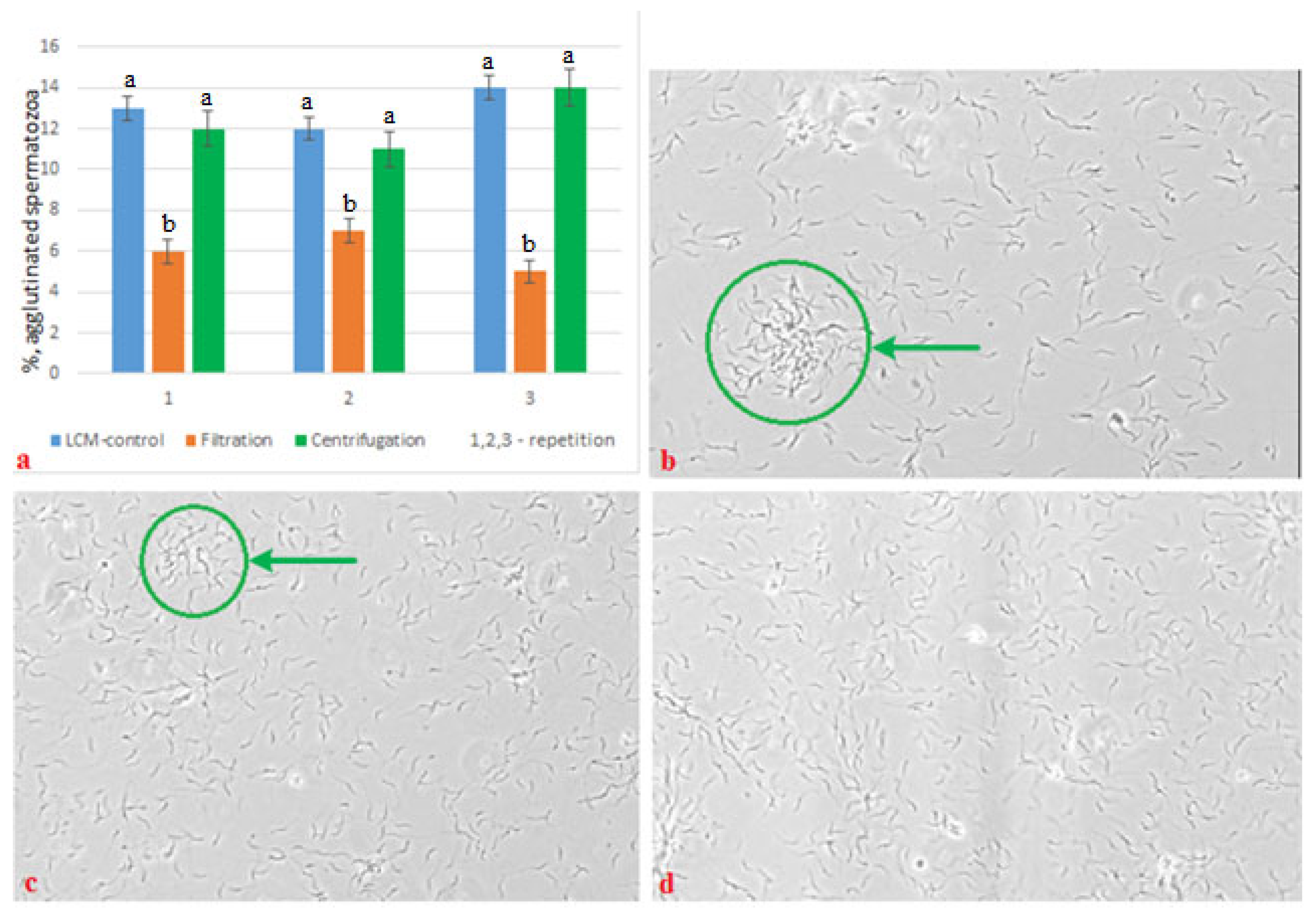
2.11. Degree of Agglutination of Spermatozoa of Native and Frozen/Thawed Semen
2.12. Statistical Analysis
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Shabani, S.; Mehri, M.; Shirmohammad, F.; Sharafi, M. Enhancement of sperm quality and fertility-related parameters in Hubbard grandparent rooster fed diets supplemented with soybean lecithin and vitamin E. Poult. Sci. 2022, 101, 101635. [Google Scholar] [CrossRef] [PubMed]
- Partyka, A.; Łukaszewicz, E.; Niżański, W. Lipid peroxidation and antioxidant enzymes activity in avian semen. Anim. Reprod. Sci. 2012, 134, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Finelli, R.; Agarwal, A.; Henkel, R. Total antioxidant capacity-Relevance, methods and clinical implications. Andrologia 2021, 53, e13624. [Google Scholar] [CrossRef] [PubMed]
- Amini, M.R.; Kohram, H.; Zare-Shahaneh, A.; Zhandi, M.; Sharideh, H.; Nabi, M.M. The effects of different levels of catalase and superoxide dismutase in modified Beltsville extender on rooster post-thawed sperm quality. Cryobiology 2015, 70, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.N.; Ramachandran, R.; Wamsley, K.; Kiess, A.; Mcdaniel, C. The Impact of in vitro inoculation and dietary supplementation with Bacillus subtilis on sperm quality of aged White Leghorn roosters. J. Appl. Poult. Res. 2018, 27, 304–315. [Google Scholar] [CrossRef]
- Pelaez, J.; Bongalhardo, D.C.; Long, J.A. Characterizing the glycocalyx of poultry spermatozoa: III. Semen cryopreservation methods alter the carbohydrate component of rooster sperm membrane glycoconjugates. Poult. Sci. 2011, 90, 435–443. [Google Scholar] [CrossRef]
- Kuroda, S.; Yumura, Y.; Mori, K.; Yasuda, K.; Takeshima, T.; Kawahara, T.; Kondo, Y. Negative correlation between the presence of reactive oxygen species and Sperm Motility Index in whole semen samples of infertile males. Rev. Int. Androl. 2017, 15, 84–89. [Google Scholar] [CrossRef]
- Soler, L.; Labas, V.; Thelie, A.; Grasseau, I.; Teixeira-Gomes, A.P.; Blesbois, E. Intact cell MALDI-TOF MS on sperm: Towards a molecular test for male fertility diagnosis. Mol. Cell Proteom. 2016, 15, 1998–2010. [Google Scholar] [CrossRef] [Green Version]
- Santiago Moreno, J.; Blesbois, E. Functional Aspects of Seminal Plasma in Bird Reproduction. Int. J. Mol. Sci. 2020, 21, 5664. [Google Scholar] [CrossRef]
- Tang, B.; Xie, G.; Hu, X.; Zhang, X.; Hu, S.; Hu, J.; Hu, B.; Li, L.; Wang, J. A comparative proteomic study of high and low semen quality seminal plasma in drakes. Poult. Sci. 2022, 101, 102130. [Google Scholar] [CrossRef]
- Li, Y.; Sun, Y.; Ni, A.; Shi, L.; Wang, P.; Isa, A.M.; Chen, J. Seminal Plasma Proteome as an Indicator of Sperm Dysfunction and Low Sperm Motility Mol. Cell Proteom. 2020, 19, 1035–1046. [Google Scholar] [CrossRef] [Green Version]
- Michailidis, G.; Avdi, M. Transcriptional profiling of gallinacinsantimicrobial peptides in the chicken reproductive tract and embryos. J. Biol. Res. 2010, 14, 211–218. [Google Scholar]
- Marzoni, M.; Castillo, A.; Sagona, S.; Citti, L.; Rocchiccioli, S.; Romboli, I.; Felicioli, A. A proteomic approach to identify seminal plasma proteins in roosters (Gallus gallus domesticus). Anim. Reprod. Sci. 2013, 140, 216–223. [Google Scholar] [CrossRef]
- Stanishevskaya, O.; Silyukova, Y.; Pleshanov, N.; Kurochkin, A. Role of Mono- and Disaccharide Combination in Cryoprotective Medium for Rooster Semen to Ensure Cryoresistance of Spermatozoa. Molecules 2021, 26, 5920. [Google Scholar] [CrossRef]
- Stanishevskaya, O.; Silyukova, Y.; Pleshanov, N.; Kurochkin, A.; Fedorova, E.; Fedorova, Z.; Perinek, O.; Prituzhalova, A.; Meftakh, I. Effects of Saccharides Supplementation in the Extender of Cryopreserved Rooster (Gallus domesticus) Semen on the Fertility of Frozen/Thawed Spermatozoa. Animals 2021, 11, 189. [Google Scholar] [CrossRef]
- GOST 27267-2017. Products for Reproduction. Non-Diluted Fresh Sperm of Cocks and Turkey-Cocks. Specifications. Federal Agency for Technical Regulation and Metrology: Moscow, Russia, 2017.
- Tselutin, K.V.; Tur, B.K. Artificial Insemination and Cryopreservation of Poultry’s Sperm (Roosters, Indica, Gander, Drake); The Russian Academy of Agricultural Sciences: Saint Petersburg, Russia, 2013; p. 88. [Google Scholar]
- Somogui, M. Determination of blood sugar. J. Biol. Chem. 1945, 160, 69. [Google Scholar] [CrossRef]
- Brobst, K.M.; Scobell, H.D. Modern Chromatographic Methods for the Analysis of Carbohydrate Mixtures. Starch-Starke 1982, 34, 117–121. [Google Scholar] [CrossRef]
- Pintado, B.; de la Fuente, J.; Roldan, E.R.S. Permeability of boar and bull spermatozoa to the nucleic acid stains propidium iodide or Hoechst 33258, or to eosin: Accuracy in the assessment of cell viability. Journal of reproduction and fertility. J. Reprod. Fertile 2000, 118, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Beletti, M.E.; Mello, M.L. Comparison between the toluidine blue stain and the Feulgen reaction for evaluation of rabbit sperm chromatin condensation and their relationship with sperm morphology. Theriogenology 2004, 62, 398–402. [Google Scholar] [CrossRef]
- Erenpreiss, J.; Jepson, K.; Giwercman, A.; Tsarev, I.; Erenpreisa, J.; Spano, M. Toluidine blue cytometry test for sperm DNA conformation: Comparison with the flow cytometric sperm chromatin structure and TUNEL assays. Hum Reprod. 2004, 19, 2277–2282. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Sandlow, J.I.; Sandra, A. Expression and Function of the c-kit Proto-Oncogene Protein in Mouse Sperm. Biol. Reprod. 1997, 57, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.L.; Han, Y.B.; Hershlag, A.; Zheng, L.J. Impact of Ca2+ Flux Inhibitors on Acrosome Reaction of Hamster Spermatozoa. J. Androl. 2007, 28, 561–564. [Google Scholar] [CrossRef]
- Sexton, T.J. Influence of seminal plasma on the fecundity of chicken spermatozoa. Theriogenology 1988, 30, 711–720. [Google Scholar] [CrossRef]
- Rad, H.M.; Eslami, M.; Ghanie, A. Palmitoleate enhances quality of rooster semen uring chilled storage. Anim. Reprod. Sci. 2016, 165, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Lenický, M.; Slanina, T.; Kačániová, M.; Galovicová, L.; Petrovičová, M.; Ďuračka, M.; Benko, F.; Kovac, J.; Tvrdá, E. Identification of Bacterial Profiles and Their Interactions with Selected Quality, Oxidative, and Immunological Parameters of Turkey Semen. Animals 2021, 11, 1771. [Google Scholar] [CrossRef]
- Pagliuca, C.; Cariati, F.; Bagnulo, F.; Scaglione, E.; Carotenuto, C.; Farina, F.; D’Argenio, V.; Carraturo, F.; D’Aprile, P.; Vitiello, M.; et al. Microbiological Evaluation and Sperm DNA Fragmentation in Semen Samples of Patients Undergoing Fertility Investigation. Genes 2021, 12, 654. [Google Scholar] [CrossRef]
- Ovchinnikov, R.I. Male infertility associated with oxidative stress of spermatozoa: Pathogenesis and therapeutic approach. Meditsinskiy Counc. Med. Counc. 2022, 5, 46–53. (In Russian) [Google Scholar] [CrossRef]
- DeLuca, M.N.; Colone, M.; Gambioli, R.; Stringaro, A.; Unfer, V. Oxidative Stress and Male Fertility: Role of Antioxidants and Inositols. Antioxidants 2021, 10, 1283. [Google Scholar] [CrossRef]
- Oliva, M.M.; Buonomo, G.; Carra, M.C.; Lippa, A.; Lisi, F. Myo-inositol impact on sperm motility in vagina and evaluation of its effects on foetal development. Eur. Rev. Med. Pharmcol. Sci. 2020, 24, 2704–2709. [Google Scholar]
- Condorelli, R.A.; La Vignera, S.; Di Bari, F.; Unfer, V.; Calogero, A.E. Effects of myoinositol on sperm mitochondrial function in-vitro. Eur. Rev. Med. Pharmcol. Sci. 2011, 15, 129–134. [Google Scholar]
- Condorelli, R.A.; La Vignera, S.; Bellanca, S.; Vicari, E.; Calogero, A.E. Myoinositol: Does it improve sperm mitochondrial function and sperm motility? Urology 2012, 79, 1290–1295. [Google Scholar] [CrossRef]
- Raeeszadeh, M.; Shokrollahi, B.; Khademi, N.; Akbari, A. Superior effect of broccoli methanolic extract on control of oxidative damage of sperm cryopreservation and reproductive performance in rats: A comparison with vitamin C and E antioxidant. Theriogenology 2022, 181, 50–58. [Google Scholar] [CrossRef]
- Raeeszadeh, M.; Khademi, N.; Akbari, A. The effects of broccoli and caraway extracts on serum oxidative markers, testicular structure and function, and sperm quality before and after sperm cryopreservation. Cryobiology 2021, 99, 11–19. [Google Scholar] [CrossRef]
- Sharaf, A.E.; Khalil, W.A.; Khalifa, E.I.; Nassan, M.A.; Swelum, A.A.; El-Harairy, M.A. The Supplementation of Bee Bread Methanolic Extract to Egg Yolk or Soybean Lecithin Extenders Can Improve the Quality of Cryopreserved Ram Semen. Cells 2022, 11, 3403. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
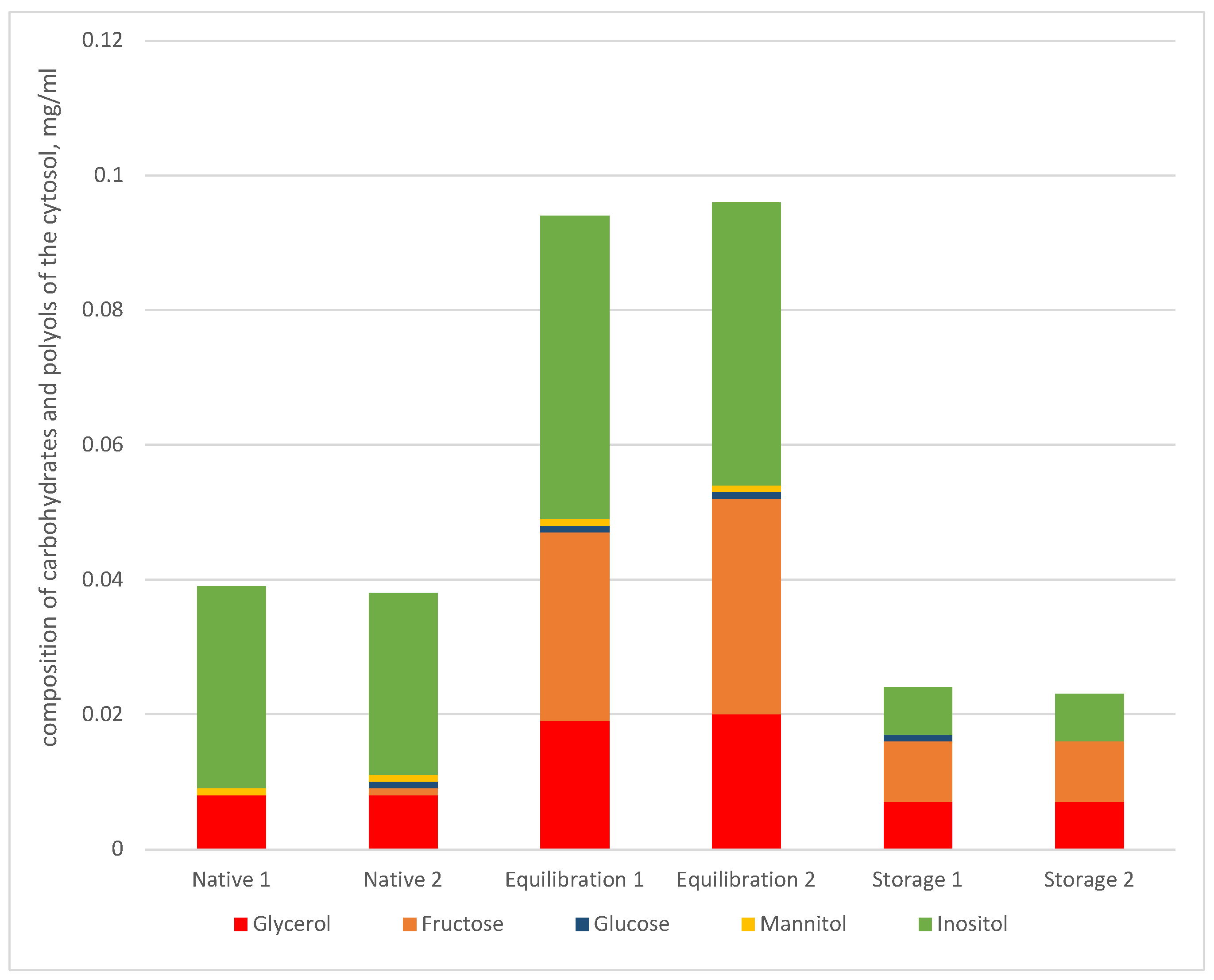{kind=link}
| n ♂ | Ejaculate Volume, mL | Concentration, Billion/mL | Total Motility (TM), % | Progressive Motility (PM), % | Viability, % | Chromatin Integrity, % |
|---|---|---|---|---|---|---|
| 22 | 0.38 ± 0.026 | 2.08 ± 0.18 | 84.3 ± 2.23 | 66.5 ± 2.5 | 72.9 ± 5.6 | 81.1 ± 1.0 |
| Qualitative Indicator | Aliquot I (LCM-Control) | Aliquot II (Filtering) | Aliquot III (Centrifugation) |
|---|---|---|---|
| concentration ppb/ml | 1.82 ± 0.05 | 1.71 ± 0.08 | 1.23 ± 0.11 |
| CV concentration, % | 5.3 | 9.4 | 18.3 |
| decrease in concentration, % | 0 | 6.0 | 33.4 |
| total motility, % | 80.0 ± 1.47 a | 79.8 ± 0.6 a | 69.5 ± 0.79 b |
| CV total motility, % | 3.7 | 1.5 | 2.3 |
| progressive motility, % | 39.2 ± 1.52 a | 41.0 ± 1.56 a | 27.0 ± 1.21 b |
| CV progressive motility, % | 7.8 | 7.6 | 9.0 |
| viability, % | 65.2 ± 0.07 | 58.6 ± 0.08 | 59.2 ± 0.03 |
| CV viability, % | 8.8 | 23.9 | 18.0 |
| chromatin integrity, % | 30.9 ± 4.12 a | 56.6 ± 0.75 b | 33.6 ± 2.85 a |
| CV chromatin integrity, % | 23.1 | 2.3 | 14.7 |
| acrosome integrity, % | 98.5 ± 0.26 | 96.3 ± 0.49 | 98.1 ± 0.34 |
| CV acrosome integrity, % | 0.5 | 0.9 | 0.6 |
| Qualitative Indicator | Aliquot I (LCM Control) | Aliquot II (Filtering) | Aliquot III (Centrifugation) |
|---|---|---|---|
| total motility, % | 44.7 ± 5.2 ab | 52.2 ± 2.3 a | 30.4 ± 3.6 b |
| progressive motility, % | 21.4 ± 0.9 a | 25.5 ± 2.5 a | 5.5 ± 0.4 b |
| viability, % | 26.8 ± 0.4 a | 40.1 ± 1.7 b | 33.4 ± 2.6 b |
| chromatin integrity, % | 90.4 ± 6.3 a | 83.5 ± 7.5 a | 64.4 ± 2.1 b |
| acrosome integrity, % | 68.2 ± 1.1 a | 68.7 ± 3.1 a | 56.9 ± 2.0 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silyukova, Y.; Fedorova, E.; Stanishevskaya, O. Influence of Technological Stages of Preparation of Rooster Semen for Short-Term and Long-Term Storage on Its Quality Characteristics. Curr. Issues Mol. Biol. 2022, 44, 5531-5542. https://doi.org/10.3390/cimb44110374
Silyukova Y, Fedorova E, Stanishevskaya O. Influence of Technological Stages of Preparation of Rooster Semen for Short-Term and Long-Term Storage on Its Quality Characteristics. Current Issues in Molecular Biology. 2022; 44(11):5531-5542. https://doi.org/10.3390/cimb44110374
Chicago/Turabian StyleSilyukova, Yulia, Elena Fedorova, and Olga Stanishevskaya. 2022. "Influence of Technological Stages of Preparation of Rooster Semen for Short-Term and Long-Term Storage on Its Quality Characteristics" Current Issues in Molecular Biology 44, no. 11: 5531-5542. https://doi.org/10.3390/cimb44110374
APA StyleSilyukova, Y., Fedorova, E., & Stanishevskaya, O. (2022). Influence of Technological Stages of Preparation of Rooster Semen for Short-Term and Long-Term Storage on Its Quality Characteristics. Current Issues in Molecular Biology, 44(11), 5531-5542. https://doi.org/10.3390/cimb44110374