

Human Non-Hypertrophic Nonunion Tissue Contains Osteoblast Lineage Cells and E-BMP-2 Activates Osteogenic and Chondrogenic Differentiation

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Characteristics
2.2. Isolation and Culture of NHNCs
2.3. Preparation of E-BMP-2
2.4. Growth Kinetics
2.5. Immunophenotyping of NHNCs Using Flow Cytometry
2.6. Differentiation Studies
2.6.1. Osteogenic Induction
2.6.2. Chondrogenic Induction
2.6.3. Isolation of Control Cells Using a Reamer-Irrigator-Aspirator (RIA)
2.6.4. Real-Time RT-PCR Analysis
2.7. Statistical Analysis
3. Results
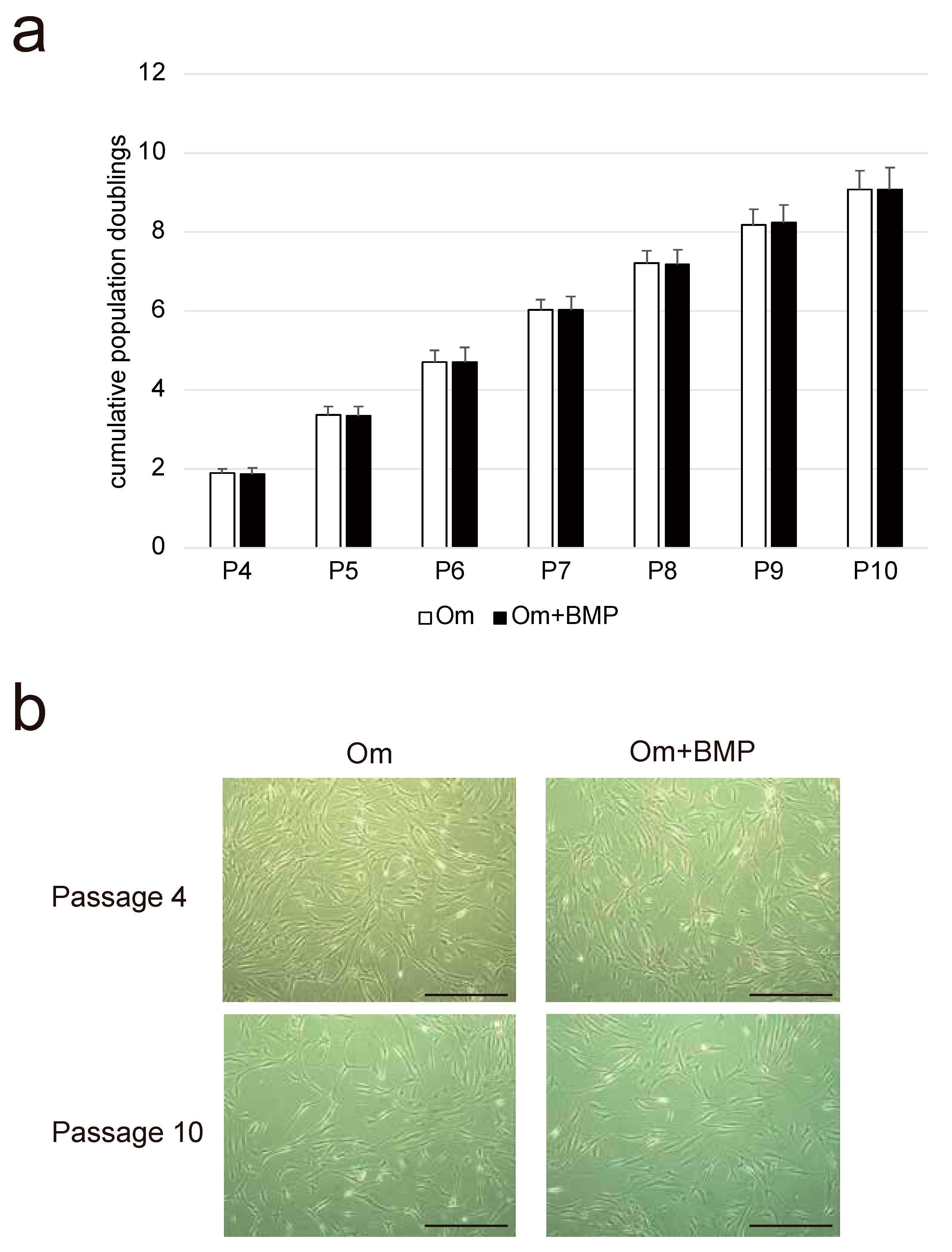
3.1. Growth Kinetics and Morphological Characteristics
3.2. Immunophenotypes
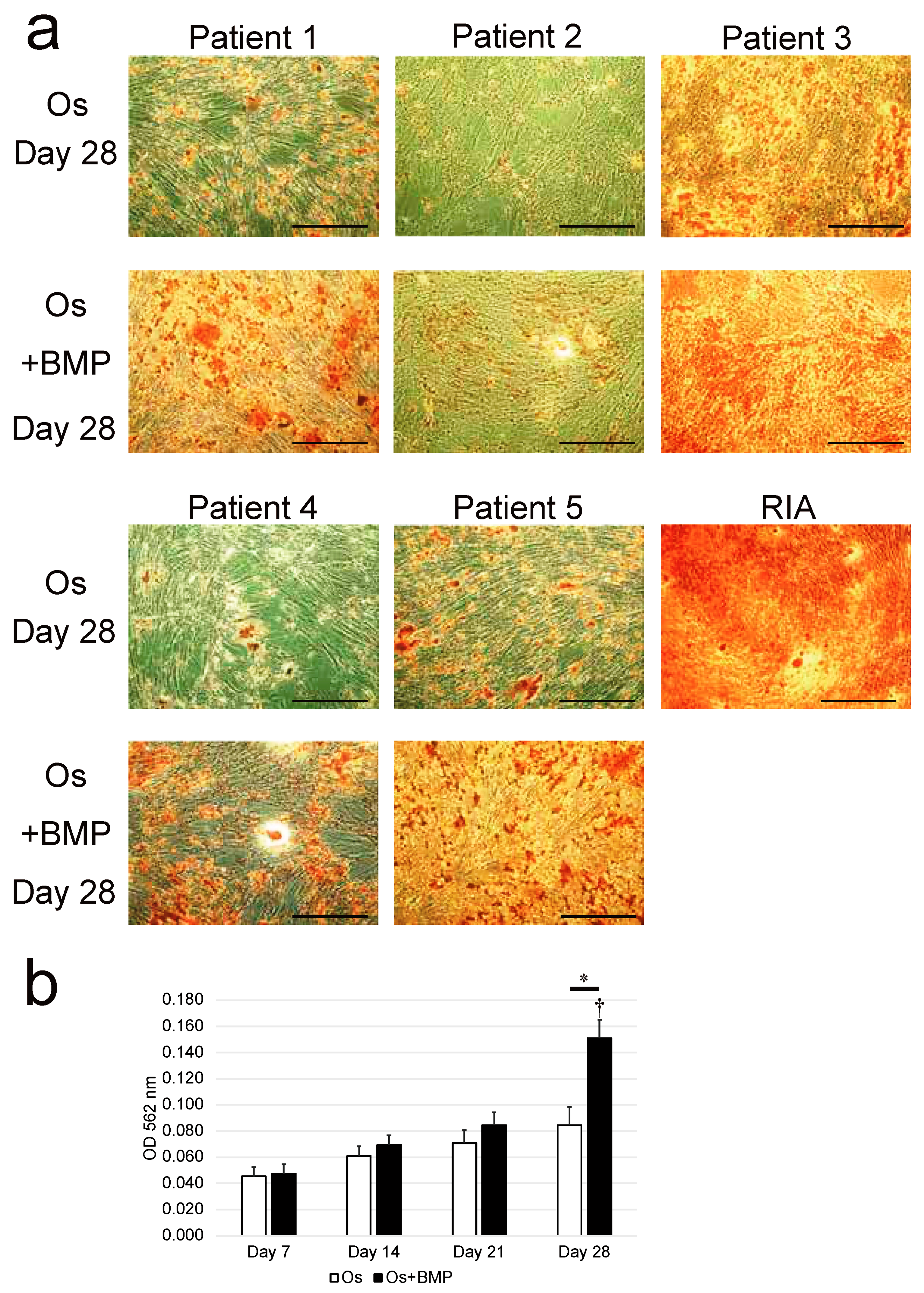
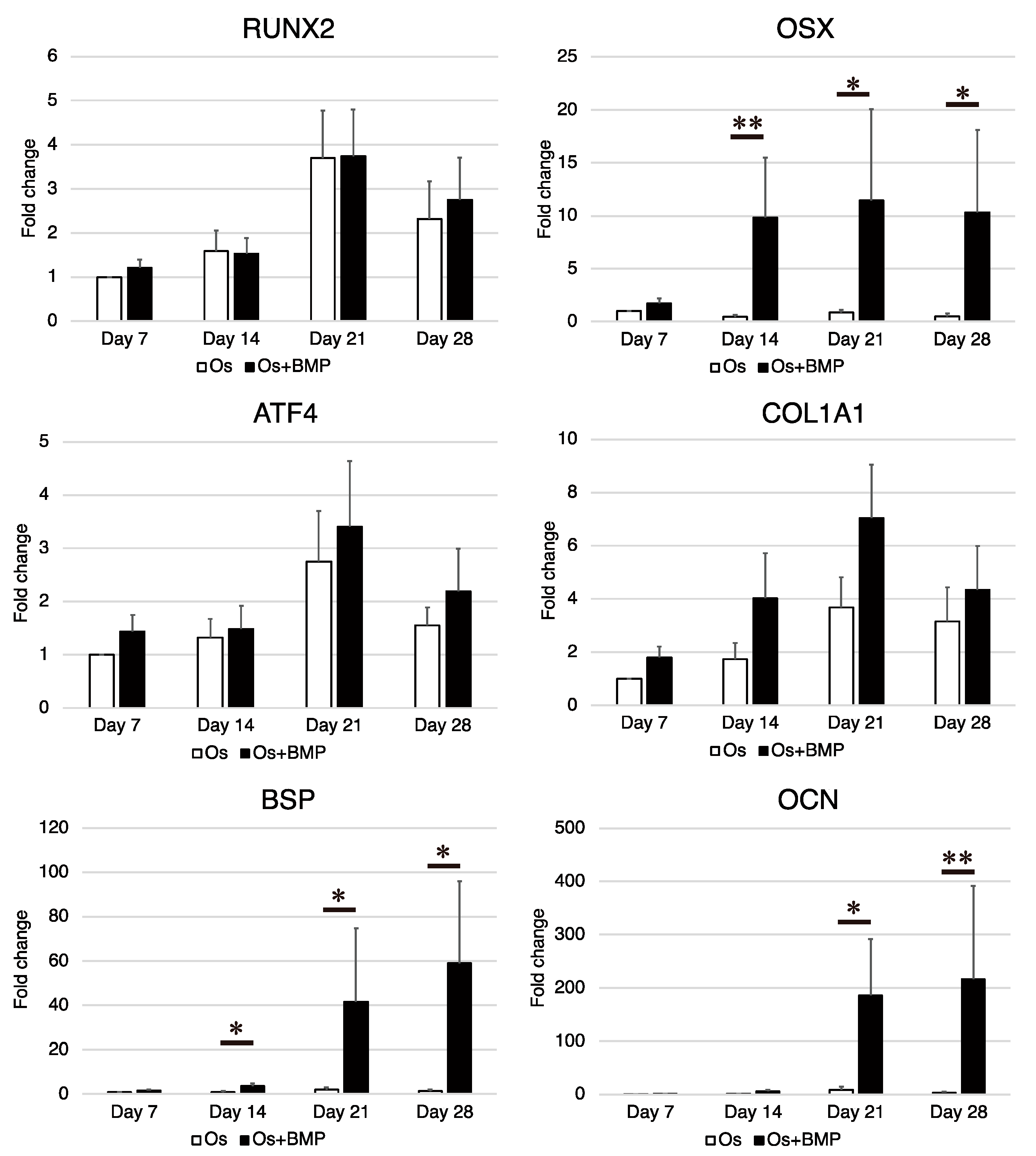
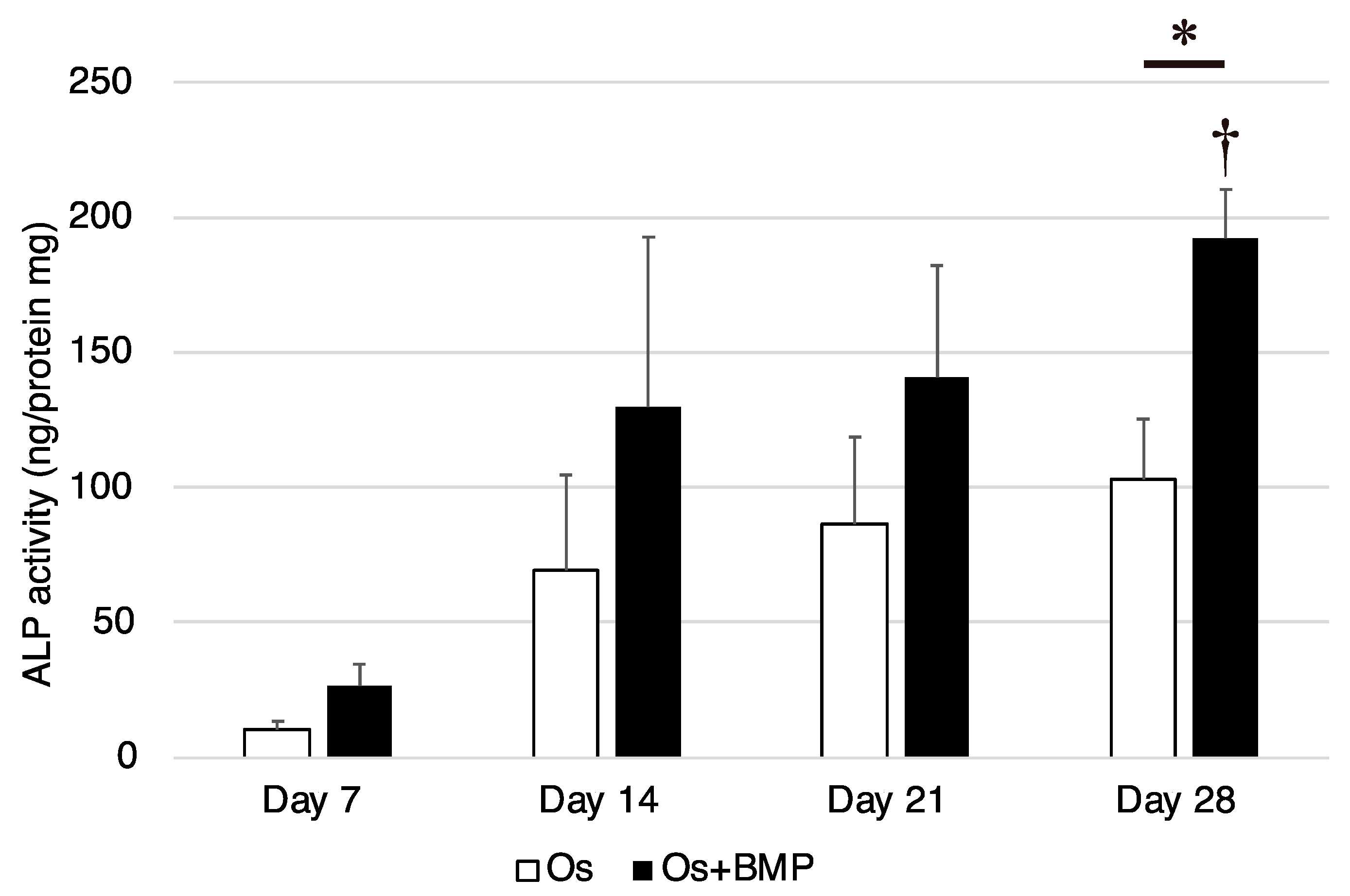
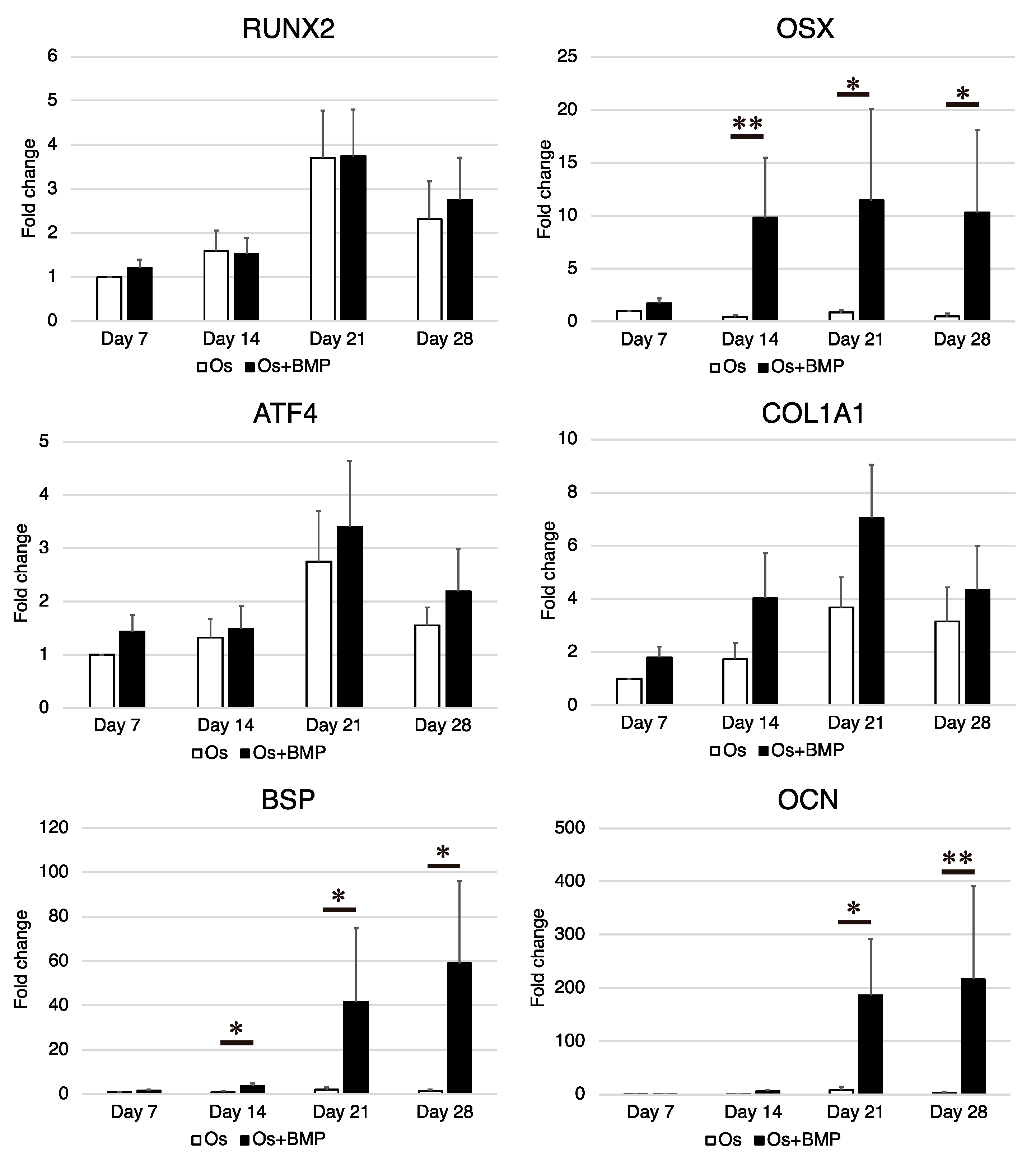
3.3. Osteogenic Differentiation Potential
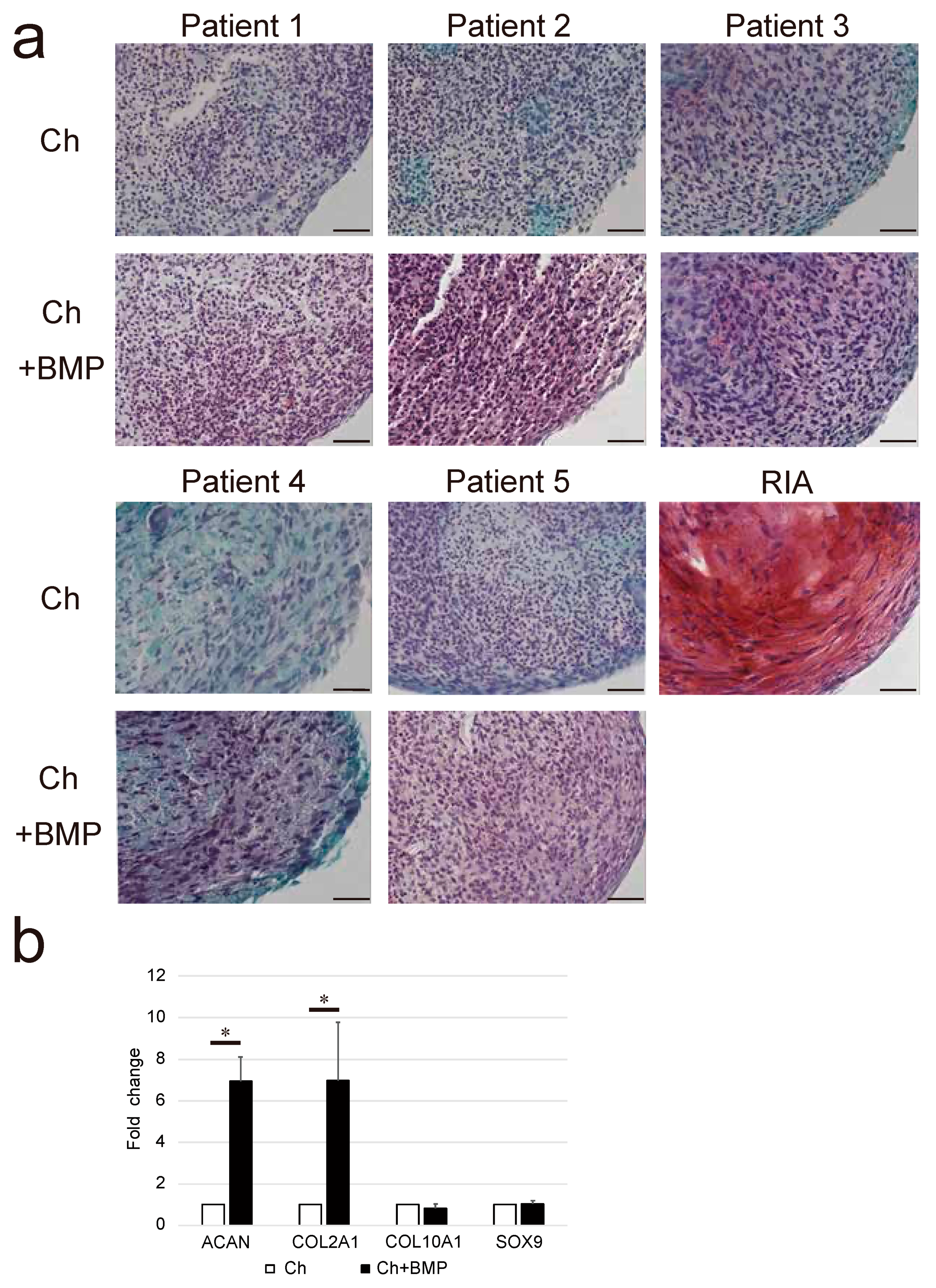
3.4. Chondrogenic Differentiation Potential
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BMP-2 | Bone morphogenetic protein-2 |
| E-BMP-2 | Escherichia coli-derived bone morphogenetic protein-2 |
| NHNCs | Non-hypertrophic nonunion cells |
| RT-PCR | Reverse transcription-polymerase chain reaction |
| rhBMP-2 | Recombinant human bone morphogenetic protein-2 |
| CHO | Chinese hamster ovary |
| PBS | Phosphate-buffered saline |
| Om | Original medium |
| PD | Population doubling |
| Os | Osteogenic medium |
| ALP | Alkaline phosphatase |
| RUNX2 | Runt-related transcription factor 2 |
| OSX | Osterix |
| ATF4 | Activating transcription factor 4 |
| COL1A1 | Collagen type I |
| BSP | Bone sialoprotein |
| OCN | Osteocalcin |
| Ch | Chondrogenic medium |
| ACAN | Aggrecan |
| COL2A1 | Collagen type II |
| COL10A1 | Collagen type X |
| SOX9 | Sry-type high-mobility group box 9 |
| RIA | Reamer-irrigator-aspirator |
| MSC | Mesenchymal stromal cell |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| BMSC | Bone marrow mesenchymal stromal cell |
References
- Bishop, G.B.; Einhorn, T.A. Current and future clinical applications of bone morphogenetic proteins in orthopaedic trauma surgery. Int. Orthop. 2007, 31, 721–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hak, D.J.; Fitzpatrick, D.; Bishop, J.A.; Marsh, J.L.; Tilp, S.; Schnettler, R.; Simpson, H.; Bishop, J.A. Delayed union and nonunions: Epidemiology, clinical issues, and financial aspects. Injury 2014, 45 (Suppl. S2), S3–S7. [Google Scholar] [CrossRef] [PubMed]
- Weber, B.G.C.O. Pseudarthrosis; Grune & Stratton: New York, NY, USA, 1976. [Google Scholar]
- Niikura, T.; Lee, S.Y.; Sakai, Y.; Nishida, K.; Kuroda, R.; Kurosaka, M. Comparison of radiographic appearance and bone scintigraphy in fracture nonunions. Orthopedics 2014, 37, e44–e50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oe, K.; Zeng, F.; Fukui, T.; Nogami, M.; Murakami, T.; Matsumoto, T.; Kuroda, R.; Niikura, T. Quantitative bone single-photon emission computed tomography imaging for uninfected nonunion: Comparison of hypertrophic nonunion and non-hypertrophic nonunion. J. Orthop. Surg. Res. 2021, 16, 125. [Google Scholar] [CrossRef]
- Iwakura, T.; Miwa, M.; Sakai, Y.; Niikura, T.; Lee, S.Y.; Oe, K.; Hasegawa, T.; Kuroda, R.; Fujioka, H.; Doita, M.; et al. Human hypertrophic nonunion tissue contains mesenchymal progenitor cells with multilineage capacity in vitro. J. Orthop. Res. 2009, 27, 208–215. [Google Scholar] [CrossRef] [Green Version]
- Takahara, S.; Niikura, T.; Lee, S.Y.; Iwakura, T.; Okumachi, E.; Kuroda, R.; Kurosaka, M. Human pseudoarthrosis tissue contains cells with osteogenic potential. Injury 2016, 47, 1184–1190. [Google Scholar] [CrossRef]
- Keskin, D.S.; Tezcaner, A.; Korkusuz, P.; Korkusuz, F.; Hasirci, V. Collagen-chondroitin sulfate-based PLLA-SAIB-coated rhBMP-2 delivery system for bone repair. Biomaterials 2005, 26, 4023–4034. [Google Scholar] [CrossRef]
- Zara, J.N.; Siu, R.K.; Zhang, X.; Shen, J.; Ngo, R.; Lee, M.; Li, W.; Chiang, M.; Chung, J.; Kwak, J.; et al. High doses of bone morphogenetic protein 2 induce structurally abnormal bone and inflammation in vivo. Tissue Eng. Part A 2011, 17, 1389–1399. [Google Scholar] [CrossRef] [Green Version]
- Fung, S.L.; Wu, X.; Maceren, J.P.; Mao, Y.; Kohn, J. In vitro evaluation of recombinant bone morphogenetic Protein-2 bioactivity for regenerative medicine. Tissue Eng. Part C Methods 2019, 25, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Israel, D.I.; Nove, J.; Kerns, K.M.; Moutsatsos, I.K.; Kaufman, R.J. Expression and characterization of bone morphogenetic protein-2 in Chinese hamster ovary cells. Growth Factors 1992, 7, 139–150. [Google Scholar] [CrossRef]
- Lee, J.H.; Jang, S.J.; Koo, T.Y.; Suh, C.W.; Lee, E.N.; Lee, K.M.; Lee, H.S.; Baek, H.R. Expression, purification and osteogenic bioactivity of recombinant human BMP-2 derived by Escherichia coli. J. Tissue Eng. Regen. Med. 2011, 8, 8–15. [Google Scholar]
- Long, S.; Truong, L.; Bennett, K.; Phillips, A.; Wong-Staal, F.; Ma, H. Expression, purification, and renaturation of bone morphogenetic protein-2 from Escherichia coli. Protein Expr. Purif. 2006, 46, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Hoshino, M.; Ohta, Y.; Manaka, T.; Naka, Y.; Imai, Y.; Sebald, W.; Takaoka, K. Osteoinductive capacity and heat stability of recombinant human bone morphogenetic protein-2 produced by Escherichia coli and dimerized by biochemical processing. J. Bone Miner. Metab. 2009, 27, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Kuroiwa, Y.; Niikura, T.; Lee, S.Y.; Oe, K.; Iwakura, T.; Fukui, T.; Matsumoto, T.; Matsushita, T.; Nishida, K.; Kuroda, R. Escherichia coli-derived BMP-2-absorbed beta-TCP granules induce bone regeneration in rabbit critical-sized femoral segmental defects. Int. Orthop. 2019, 43, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Megas, P. Classification of non-union. Injury 2005, 36 (Suppl. S4), S30–S37. [Google Scholar] [CrossRef]
- Hernigou, P.; Poignard, A.; Beaujean, F.; Rouard, H. Percutaneous autologous bone-marrow grafting for nonunions. Influence of the number and concentration of progenitor cells. J. Bone Jt. Surg. Am. 2005, 87, 1430–1437. [Google Scholar] [CrossRef]
- Oe, K.; Miwa, M.; Sakai, Y.; Lee, S.Y.; Kuroda, R.; Kurosaka, M. An in vitro study demonstrating that haematomas found at the site of human fractures contain progenitor cells with multilineage capacity. J. Bone Jt. Surg. Br. 2007, 89, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Ruppert, R.; Hoffmann, E.; Sebald, W. Human bone morphogenetic protein 2 contains a heparin-binding site which modifies its biological activity. Eur. J. Biochem. 1996, 237, 295–302. [Google Scholar] [CrossRef]
- Matsumoto, T.; Toyoda, H.; Dohzono, S.; Yasuda, H.; Wakitani, S.; Nakamura, H.; Takaoka, K. Efficacy of interspinous process lumbar fusion with recombinant human bone morphogenetic protein-2 delivered with a synthetic polymer and beta-tricalcium phosphate in a rabbit model. Eur. Spine J. 2012, 21, 1338–1345. [Google Scholar] [CrossRef] [Green Version]
- Cristofalo, V.J.; Allen, R.G.; Pignolo, R.J.; Martin, B.G.; Beck, J.C. Relationship between donor age and the replicative lifespan of human cells in culture: A reevaluation. Proc. Natl. Acad. Sci. USA 1998, 95, 10614–10619. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Matsunuma, A.; Kurahashi, I.; Yanagawa, T.; Yoshida, H.; Horiuchi, N. Induction of osteoblast differentiation indices by statins in MC3T3-E1 cells. J. Cell Biochem. 2004, 92, 458–471. [Google Scholar] [CrossRef]
- Lee, S.Y.; Miwa, M.; Sakai, Y.; Kuroda, R.; Matsumoto, T.; Iwakura, T.; Fujioka, H.; Doita, M.; Kurosaka, M. In vitro multipotentiality and characterization of human unfractured traumatic hemarthrosis-derived progenitor cells: A potential cell source for tissue repair. J. Cell Physiol. 2007, 210, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Bajada, S.; Marshall, M.J.; Wright, K.T.; Richardson, J.B.; Johnson, W.E. Decreased osteogenesis, increased cell senescence and elevated Dickkopf-1 secretion in human fracture non union stromal cells. Bone 2009, 45, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Schmidmaier, G.; Herrmann, S.; Green, J.; Weber, T.; Scharfenberger, A.; Haas, N.P.; Wildemann, B. Quantitative assessment of growth factors in reaming aspirate, iliac crest, and platelet preparation. Bone 2006, 39, 1156–1163. [Google Scholar] [CrossRef]
- Kuehlfluck, P.; Moghaddam, A.; Helbig, L.; Child, C.; Wildemann, B.; Schmidmaier, G.; HTRG-Heidelberg Trauma Research Group. RIA fractions contain mesenchymal stroma cells with high osteogenic potency. Injury 2015, 46 (Suppl. S8), S23–S32. [Google Scholar] [CrossRef]
- Toosi, S.; Naderi-Meshkin, H.; Kalalinia, F.; Peivandi, M.T.; Hossein, K.H.; Bahrami, A.R.; Heirani-Tabasi, A.; Mirahmadi, M.; Behravan, J. Comparative characteristics of mesenchymal stem cells derived from reamer-irrigator-aspirator, iliac crest bone marrow, and adipose tissue. Cell Mol. Biol. 2016, 62, 68–74. [Google Scholar]
- Ismail, H.D.; Phedy, P.; Kholinne, E.; Kusnadi, Y.; Sandhow, L.; Merlina, M. Existence of mesenchymal stem cells in sites of atrophic nonunion. Bone Jt. Res. 2013, 2, 112–115. [Google Scholar] [CrossRef]
- Geiger, M.; Li, R.H.; Friess, W. Collagen sponges for bone regeneration with rhBMP-2. Adv. Drug Deliv. Rev. 2003, 55, 1613–1629. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.H. BMP-2 mediates PGE2-induced reduction of proliferation and osteogenic differentiation of human tendon stem cells. J. Orthop. Res. 2012, 30, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Gromolak, S.; Krawczenko, A.; Antończyk, A.; Buczak, K.; Kiełbowicz, Z.; Klimczak, A. Biological characteristics and osteogenic differentiation of ovine bone marrow derived mesenchymal stem cells stimulated with FGF-2 and BMP-2. Int. J. Mol. Sci. 2020, 21, 9726. [Google Scholar] [CrossRef]
- Garrison, K.R.; Shemilt, I.; Donell, S.; Ryder, J.J.; Mugford, M.; Harvey, I.; Song, F.; Alt, V. Bone morphogenetic protein (BMP) for fracture healing in adults. Cochrane Database Syst. Rev. 2010, 6, CD006950. [Google Scholar] [CrossRef] [PubMed]
- Kämmerer, P.W.; Pabst, A.M.; Dau, M.; Staedt, H.; Al-Nawas, B.; Heller, M. Immobilization of BMP-2, BMP-7 and alendronic acid on titanium surfaces: Adhesion, proliferation and differentiation of bone marrow-derived stem cells. J. Biomed. Mater Res. A 2020, 108, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Regulation of bone development and extracellular matrix protein genes by RUNX2. Cell Tissue Res. 2010, 339, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.; Li, Q.; Lin, X.; Hu, N.; Liao, J.Y.; Lin, L.B.; Zhao, C.; Hu, Z.-M.; Liang, X.; Xu, W.; et al. BMP2 induces chondrogenic differentiation, osteogenic differentiation and endochondral ossification in stem cells. Cell Tissue Res. 2016, 366, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Hino, K.; Saito, A.; Kido, M.; Kanemoto, S.; Asada, R.; Takai, T.; Cui, M.; Cui, X.; Imaizumi, K. Master regulator for chondrogenesis, Sox9, regulates transcriptional activation of the endoplasmic reticulum stress transducer BBF2H7/CREB3L2 in chondrocytes. J. Biol. Chem. 2014, 289, 13810–13820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tall, M.; Bonkoungou, D.; Sawadogo, M.; Da, S.C.; Toe, M.F. Treatment of nonunion in neglected long bone shaft fractures by osteoperiosteal decortication. Orthop. Traumatol. Surg. Res. 2014, 100, S299–S303. [Google Scholar] [CrossRef] [Green Version]
- Sen, M.K.; Miclau, T. Autologous iliac crest bone graft: Should it still be the gold standard for treating nonunions? Injury 2007, 38 (Suppl. S1), S75–S80. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef]
- Gómez-Barrena, E.; Padilla-Eguiluz, N.G.; Rosset, P. Frontiers in non-union research. EFORT Open Rev. 2020, 5, 574–583. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age (Years) | Fracture Site | Initial Treatment | Duration from Fracture (Months) |
|---|---|---|---|---|---|
| 1 | Female | 20 | Clavicle diaphysis | Conservative | 6 |
| 2 | Male | 23 | Tibia diaphysis | Intramedullary locking nail | 10 |
| 3 | Female | 59 | Femoral diaphysis | Intramedullary locking nail | 9.5 |
| 4 | Male | 24 | Femoral diaphysis | Plate-and-screw fixation | 12 |
| 5 | Male | 57 | Femoral diaphysis | Intramedullary locking nail | 12 |
| Gene | Primer Sequences |
|---|---|
| GAPDH | Forward: 5′-CGTCTTCACCACCATGGAGA-3′ Reverse: 5′-CGGCCATCACGCCACAGTTT-3′ |
| RUNX2 | Forward: 5′-ATGCTTCATTCGCCTCACAAAC-3′ Reverse: 5′-CCAAAAGAAGTTTTGCTGACATGG-3′ |
| OSX | Forward: 5′-CGGGACTCAACAACTCT-3′ Reverse: 5′-CCATAGGGGTGTGTCAT-3′ |
| ATF4 | Forward: 5′-CTGACCACGTTGGATGACAC-3′ Reverse: 5′-GGGCTCATACAGATGCCTCT-3′ |
| COL1A1 | Forward: 5′-AGGAATTCGGCTTCGACGTT-3′ Reverse: 5′-GGTTCAGTTTGGGTTGCTTG-3′ |
| BSP | Forward: 5′-AAACGAAGAAAGCGAAGCAGAA-3′ Reverse: 5′-GCTGCCGTTGCCGTTTT-3′ |
| OCN | Forward: 5′-CATGAGAGCCCTCACA-3′ Reverse: 5′-AGAGCGACACCCTAGAC-3′ |
| ACAN | Forward: 5′-TGAGGAGGGCTGGAACAAGTACC-3′ Reverse: 5′-GGAGGTGGTAATTGCAGGGAACA-3′ |
| COL2A1 | Forward: 5′-TTTCCCAGGTCAAGATGGTC-3′ Reverse: 5′-CTTCAGCACCTGTCCACCA-3′ |
| COL10A1 | Forward: 5′-GCCCAAGAGGTGCCCCTGGAATAC-3′ Reverse: 5′-CCTGAGAAAGAGGAGTGGACATAC-3′ |
| SOX9 | Forward: 5′-ATCTGAAGAAGGAGAGCGAG-3′ Reverse: 5′-TCAGAAGTCTCCAGAGCTTG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshikawa, R.; Fukui, T.; Oe, K.; Kumabe, Y.; Oda, T.; Sawauchi, K.; Takase, K.; Yamamoto, Y.; Sakai, Y.; Kuroda, R.; et al. Human Non-Hypertrophic Nonunion Tissue Contains Osteoblast Lineage Cells and E-BMP-2 Activates Osteogenic and Chondrogenic Differentiation. Curr. Issues Mol. Biol. 2022, 44, 5562-5578. https://doi.org/10.3390/cimb44110377
Yoshikawa R, Fukui T, Oe K, Kumabe Y, Oda T, Sawauchi K, Takase K, Yamamoto Y, Sakai Y, Kuroda R, et al. Human Non-Hypertrophic Nonunion Tissue Contains Osteoblast Lineage Cells and E-BMP-2 Activates Osteogenic and Chondrogenic Differentiation. Current Issues in Molecular Biology. 2022; 44(11):5562-5578. https://doi.org/10.3390/cimb44110377
Chicago/Turabian StyleYoshikawa, Ryo, Tomoaki Fukui, Keisuke Oe, Yohei Kumabe, Takahiro Oda, Kenichi Sawauchi, Kyohei Takase, Yuya Yamamoto, Yoshitada Sakai, Ryosuke Kuroda, and et al. 2022. "Human Non-Hypertrophic Nonunion Tissue Contains Osteoblast Lineage Cells and E-BMP-2 Activates Osteogenic and Chondrogenic Differentiation" Current Issues in Molecular Biology 44, no. 11: 5562-5578. https://doi.org/10.3390/cimb44110377
APA StyleYoshikawa, R., Fukui, T., Oe, K., Kumabe, Y., Oda, T., Sawauchi, K., Takase, K., Yamamoto, Y., Sakai, Y., Kuroda, R., & Niikura, T. (2022). Human Non-Hypertrophic Nonunion Tissue Contains Osteoblast Lineage Cells and E-BMP-2 Activates Osteogenic and Chondrogenic Differentiation. Current Issues in Molecular Biology, 44(11), 5562-5578. https://doi.org/10.3390/cimb44110377