
Glutathione and Its Metabolic Enzymes in Gliomal Tumor Tissue and the Peritumoral Zone at Different Degrees of Anaplasia

 , and
, and
Abstract
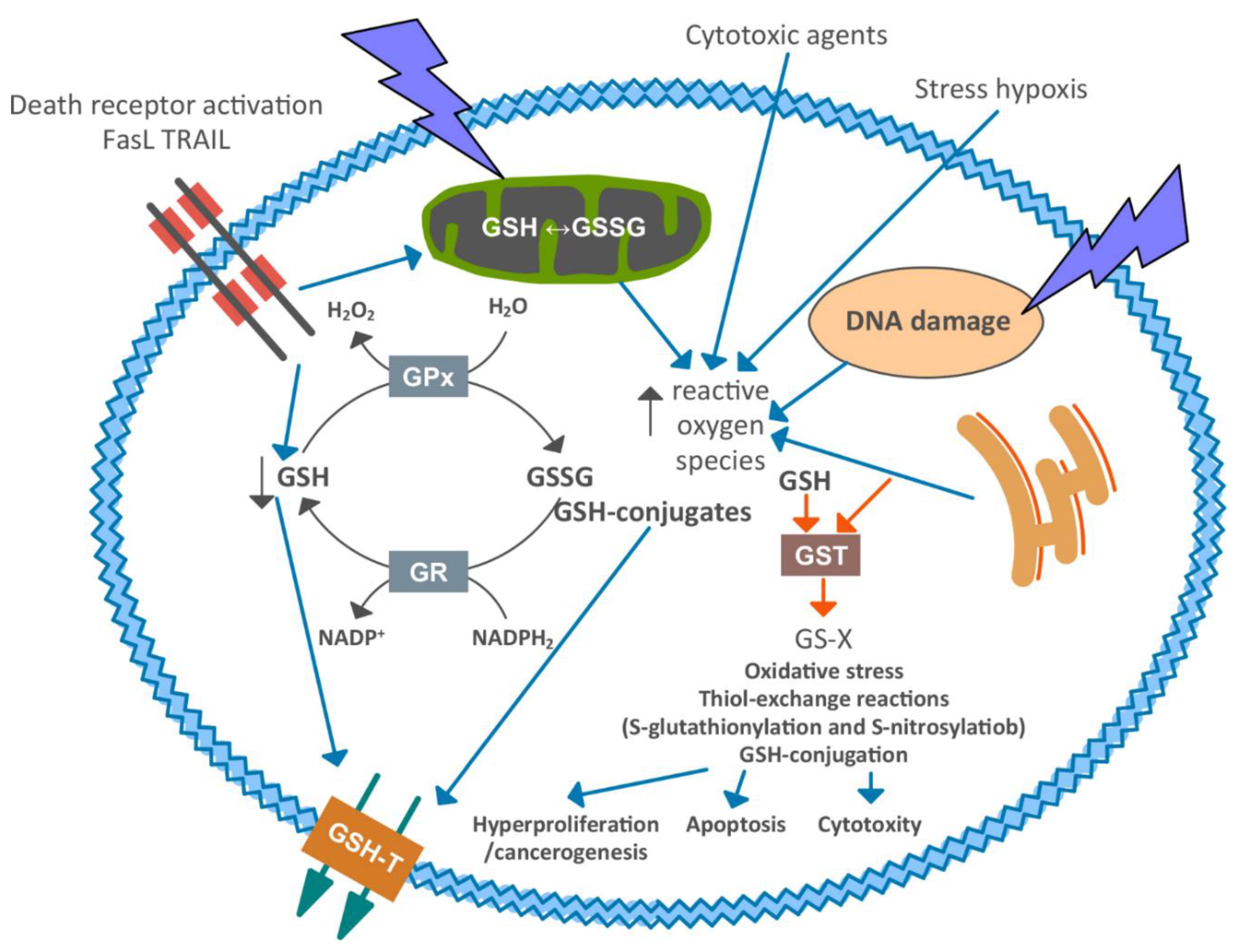
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Tissue Homogenate for Biochemical Research
2.3. Levels of Reactive Oxygen Species
2.4. Analysis of the Parameters of Glutathione Metabolism
2.5. Statistical Analysis
3. Results
4. Discussion
- For Grade I gliomas, there is a classic picture of oxidative stress: glutathione is actively restored in response to the effects of reactive oxygen species and the levels of all the glutathione defense enzymes significantly increase;
- At Grade II, an imbalance of the processes begins, manifested by multidirectional changes in the indicators;
- At Grades III and IV, there is a decrease in all indicators of the glutathione pool. The amounts of reduced glutathione and glutathione disulfide decrease sharply;
- For Low Grade (I, II) and High Grade (III, IV) gliomas, a tendency has been revealed towards opposite changes of all the examined parameters of free radical activity and of glutathione metabolism in the peritumoral zone.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sánchez-Pérez, Y.; Soto-Reyes, E.; García-Cuellar, C.M.; Cacho-Díaz, B.; Santamaría, A.; Rangel-López, E. Role of epigenetics and oxidative stress in gliomagenesis. CNS Neurol. Disord. Drug Targets 2017, 16, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J. Why is the nervous system vulnerable to oxidative stress? In Oxidative Stress and Free Radical Damage in Neurology, Oxidative Stress in Applied Basic Research and Clinical Practice; Gadoth, N., Göbel, H.H., Eds.; Humana Press: New York, NY, USA, 2011; pp. 19–27. [Google Scholar]
- Traverso, N.; Ricciarelli, R.; Nitti, M.; Marengo, B.; Furfaro, A.L.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C. Role of glutathione in cancer progression and chemoresistance. Oxidative Med. Cell. Longev. 2013, 2013, 972913. [Google Scholar] [CrossRef] [PubMed]
- Schumacker, P.T. Reactive oxygen species in cancer: A dance with the devil. Cancer Cell 2015, 27, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Corso, C.R.; Acco, A. Glutathione system in animal model of solid tumors: From regulation to therapeutic target. Crit. Rev. Oncol. Hematol. 2018, 128, 43–57. [Google Scholar] [CrossRef]
- Bansal, A.; Simon, M.C. Glutathione metabolism in cancer progression and treatment resistance. J. Cell Biol. 2018, 217, 2291–2298. [Google Scholar] [CrossRef]
- Kennedy, L.; Sandru, J.K.; Harper, M.E.; Cuperlovic-Culf, M. Role of glutathione in cancer: From mechanisms to therapies. Biomolecules 2020, 10, 1429. [Google Scholar] [CrossRef]
- Tew, K.D. Glutathione-associated enzymes in anticancer drug resistance. Cancer Res. 2016, 76, 7–9. [Google Scholar] [CrossRef]
- Altieri, R.; Barbagallo, D.; Certo, F. Peritumoral microenvironment in high-grade gliomas: From flairectomy to microglia–glioma cross-talk. Brain Sci. 2021, 23, 200. [Google Scholar] [CrossRef]
- Gu, R.; Zhang, X.; Zhang, G.; Tao, T.; Yu, H.; Liu, L.; Dou, Y.; Li, A.; Qin, J. Probing the Bi-directional interaction between microglia and gliomas in a tumor microenvironment on a microdevice. Neurochem. Res. 2017, 42, 1478–1487. [Google Scholar] [CrossRef]
- Basheer, A.S.; Abas, F.; Othman, I.; Naidu, R. Role of Inflammatory mediators, macrophages, and neutrophils in glioma maintenance and progression: Mechanistic understanding and potential therapeutic applications. Cancers 2021, 13, 4226. [Google Scholar] [CrossRef]
- Tamai, S.; Ichinose, T.; Tsutsui, T.; Tanaka, S.; Garaeva, F.; Sabit, H.; Nakada, M. Tumor microenvironment in glioma invasion. Brain Sci. 2022, 12, 505. [Google Scholar] [CrossRef] [PubMed]
- Barthel, L.; Hadamitzky, M.; Dammann, P. Glioma: Molecular signature and crossroads with tumor microenvironment. Cancer Metastasis Rev. 2022, 41, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Radin, D.P.; Tsirka, S.E. Interactions between tumor cells, neurons, and microglia in the glioma microenvironment. Int. J. Mol. Sci. 2020, 21, 8476. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N. WHO Classification of tumours editorial board. Central nervous system tumours. Int. Agency Res. Cancer 2021, 5, 6. [Google Scholar]
- Vladimirov, Y.A.; Proskurnina, E.V. Free radicals and cell chemiluminescence. Biochem. 2009, 74, 1545–1566. [Google Scholar] [CrossRef]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free. Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; Leonart, M.E. Oxidative stress and cancer: An overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radical Bio. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis Int. J. Program. Cell Death 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Halliwell, B. The antioxidant paradox. Lancet 2000, 355, 1179–1180. [Google Scholar] [CrossRef]
- Qu, Y.; Wang, J.; Ray, P.S.; Guo, H.; Huang, J.; Shin-Sim, M.; Bukoye, B.A.; Liu, B.; Lee, A.V.; Lin, X.; et al. Thioredoxin-like 2 regulates human cancer cell growth and metastasis via redox homeostasis and NF-κB signaling. J. Clin. Investig. 2011, 121, 212–225. [Google Scholar] [CrossRef] [PubMed]
- Gamcsik, M.P.; Kasibhatla, M.S.; Teeter, S.D.; Colvin, O.M. Glutathione levels in human tumors. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2012, 17, 671–691. [Google Scholar] [CrossRef]
- Deponte, M. Glutathione catalysis and the reaction mechanisms of glutathione-dependent enzymes. Biochim. Biophys. Acta 2013, 1830, 3217–3266. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Quintana-Cabrera, R.; Bolaños, J.P. Glutathione and γ-glutamylcysteine in hydrogen peroxide detoxification. Methods Enzymol. 2013, 527, 129–144. [Google Scholar] [CrossRef]
- Dawane, J.S.; Pandit, V.A. Understanding redox homeostasis and its role in cancer. J. Clin. Diagn. Res. JCDR 2012, 6, 1796–1802. [Google Scholar] [CrossRef]
- Hatem, E.; El Banna, N.; Huang, M.E. Multifaceted roles of glutathione and glutathione-based systems in carcinogenesis and anticancer drug resistance. Antioxid. Redox Signal. 2017, 27, 1217–1234. [Google Scholar] [CrossRef]
- Martín-Rufián, M.; Nascimento-Gomes, R.; Higuero, A.; Crisma, A.R.; Campos-Sandoval, J.A.; Gómez-García, M.C.; Cardona, C.; Cheng, T.; Lobo, C.; Segura, J.A.; et al. Both GLS silencing and GLS2 overexpression synergize with oxidative stress against proliferation of glioma cells. J. Mol. Med. 2014, 92, 277–290. [Google Scholar] [CrossRef]
- Dokic, I.; Hartmann, C.; Herold-Mende, C.; Régnier-Vigouroux, A. Glutathione peroxidase 1 activity dictates the sensitivity of glioblastoma cells to oxidative stress. Glia 2012, 60, 1785–1800. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Zhu, Z.; Du, S.; Du, Y.; Ren, J.; Ying, G.; Yan, Z. Glutathione reductase mediates drug resistance in glioblastoma cells by regulating redox homeostasis. J. Neurochem. 2018, 144, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Sha, H.; Xu, X.; Hu, T.; Lou, R.; Li, H.; Wu, J.; Dan, C.; Feng, J. Glutathione S-transferase π: A potential role in antitumor therapy. Drug Des. Res. Pract. Med. J. 2021, 8, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Dong, D. Human cytosolic glutathione transferases: Structure, function, and drug discovery. Trends Pharmacol. Sci. 2012, 33, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Allocati, N.; Masulli, M.; Di Ilio, C.; Federici, L. Glutathione transferases: Substrates, inhibitors and pro-drugs in cancer and neurodegenerative diseases. Oncogenesis 2018, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Tikhanovich, I.; Cox, J.; Weinman, S.A. Forkhead box class O transcription factors in liver function and disease. J. Gastroenterol. Hepatol. 2013, 28, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Klaus, A.; Zorman, S.; Berthier, A.; Polge, C.; Ramirez, S.; Michelland, S.; Sève, M.; Vertommen, D.; Rider, M.; Lentze, N.; et al. Glutathione S-transferases interact with AMP-activated protein kinase: Evidence for S-glutathionylation and activation in vitro. PLoS ONE 2013, 8, e62497. [Google Scholar] [CrossRef] [PubMed]
- Markovic, J.; Borras, C.; Ortega, A.; Sastre, J.; Vina, J.; Pallardo, F.V. Glutathione is recruited intothe nucleus in early phases of cell proliferation. J. Biol. Chem. 2007, 282, 20416–20424. [Google Scholar] [CrossRef]
- Garcia-Gimenez, J.L.; Markovic, J.; Dasi, F.; Queval, G.; Schnaubelt, D.; Foyer, C.H.; Pallardo, F.V. Nuclear glutathione. Biochim. Biophy Sica Acta 2013, 1830, 3304–3316. [Google Scholar] [CrossRef]
- Franco, R.; Schoneveld, O.J.; Pappa, A.; Panayiotidis, M.I. The central role of glutathione in the pathophysiology of human diseases. Arch. Physiol. Biochem. 2007, 113, 234–258. [Google Scholar] [CrossRef]
- Tsugawa, S.; Noda, Y.; Tarumi, R.; Mimura, Y.; Yoshida, K.; Iwata, Y.; Elsalhy, M.; Kuromiya, M.; Kurose, S.; Masuda, F.; et al. Glutathione levels and activities of glutathione metabolism enzymes in patients with schizophrenia: A systematic review and meta-analysis. J. Psychopharmacol. 2019, 33, 1199–1214. [Google Scholar] [CrossRef]


{kind=link}
| Adjacent Noncancerous Tissues (Median; Quartiles) | Peritumoral Zone | Tumor | ||||
|---|---|---|---|---|---|---|
| Median; Quartiles | U-Criterion of Mann–Whitney | Median; Quartiles | U-Criterion of Mann–Whitney | |||
| Free radical activity (I max), mV | Grade I (n = 1) | 121 | 97 | 169 | ||
| Grade II, (n = 6) | 265.50 (264.00–290.25) | 173.00 (155.75–190.25) | * p = 0.004 | 227.50 (209.25–341.75 | p = 0.393 | |
| Grade III, (n = 3) | 130.00 (130.00–180.50) | 205.00 (187.50–218.00) | p = 0.275 | 215.00 (189.50–273.00) | p = 0.275 | |
| Grade IV, (n = 10) | 230.50 (163.25–293.25) | 274.00 (221.50–327.25) | p = 0.200 | 357.60 (334.750–381.00) | * p = 0.011 | |
| Oxidized glutathione, μmol/L | Grade I (n = 1) | 12,583.00 | 13,636.00 | 10,532.00 | ||
| Grade I, (n = 6) | 11,514.50 (10,897.465–12,141.75) | 14,359.00 (13,350.75–15,753.50) | * p = 0.054 | 11,430.50 (9772.00–12,059.25) | p = 0.631 | |
| Grade III, (n = 3) | 12,256.00 (11,191.00–12,661.04) | 8137.00 (8124.79–9019.50) | * p = 0.049 | 4013.00 (3635.89–4975.50) | * p = 0.050 | |
| Grade IV, (n = 10) | 11,363.33 (11,146.00–11,853.17) | 9021.00 (8462.97–9837.00) | * p = 0.007 | 6854.65 (6464.75–7006.58) | * p = 0.004 | |
| Reduced glutathione, μmol/L | Grade I (n = 1) | 11,239.00 | 15,210.00 | 1323.00 | ||
| Grade II, (n = 6) | 9342.00 (8783.75–9922.75) | 10,639.50 (7043.55–11,928.00) | p = 0.522 | 14,132.00 (13,294.81–14,675.25) | * p = 0.003 | |
| Grade III, (n = 3) | 10,396.00 (10,033.00–11,347.78) | 6921.00 (5174.08–7725.50) | * p = 0.049 | 7813.00 (7059.00–8641.78) | * p = 0.049 | |
| Grade IV, (n = 10) | 9705.62 (9417.06–9970.00) | 8004.50 (6478.75–8315.22) | p = 0.109 | 8006.50 (7777.00–8636.50) | * p = 0.054 | |
| Glutathione peroxidase, ng/g of protein | Grade I (n = 1) | 8.25 | 24.71 | 21.30 | ||
| Grade II, (n = 6) | 6.84 (5.89–7.44) | 20.00 (19.25–21.50) | * p = 0.004 | 3.79 (3.52–4.73) | * p = 0.010 | |
| Grade III, (n = 3) | 6.43 (4.33–7.06) | 3.88 (2.89–4.07) | p = 0.275 | 3.78 (2.43–5.12) | p = 0.513 | |
| Grade IV, (n = 10) | 7.72 (7.06–12.89) | 3.22 (2.56–4.37) | * p = 0.039 | 2.63 (1.41–3.78) | * p = 0.0002 | |
| Glutathione reductase, pg/g of protein | Grade I (n = 1) | 2105.00 | 3179.00 | 2769.23 | ||
| Grade II, (n = 6) | 1802.00 (1625.25–1872.17) | 3188.50 (3029.00–3452.25) | * p = 0.007 | 1536.49 (1351.56–1698.41) | p = 0.149 | |
| Grade III, (n = 3) | 1750.28 (1657.14–1810.64 | 973.00 (569.84–987.00) | * p = 0.049 | 1134.00 (895.02–1338.36) | * p = 0.049 | |
| Grade IV, (n = 10) | 1894.00 (1482.49–2252.00) | 994.35 (685.42–1739.20) | p = 0.065 | 907.79 (420.39–1374.25) | * p = 0.011 | |
| Glutathione transferase, pg/g of protein | Grade I (n = 1) | 28.25 | 78.19 | 67.31 | ||
| Grade II, (n = 6) | 20.07 (16.60–22.08) | 70.70 (66.60–74.61) | * p = 0.004 | 54.47 (41.82–61.49) | * p = 0.007 | |
| Grade III, (n = 3) | 27.00 (26.02–28.50) | 19.00 (18.52–21.00) | * p = 0.005 | 15.00 (13.48–24.17) | p = 0.513 | |
| Grade IV, (n = 10) | 35.17 (32.89–55.20) | 33.24 (23.21–40.51) | p = 0.064 | 12.09 (7.37–16.32) | * p = 0.001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obukhova, L.; Kopytova, T.; Murach, E.; Shchelchkova, N.; Kontorshchikova, C.; Medyanik, I.; Orlinskaya, N.; Grishin, A.; Kontorshchikov, M.; Badanina, D. Glutathione and Its Metabolic Enzymes in Gliomal Tumor Tissue and the Peritumoral Zone at Different Degrees of Anaplasia. Curr. Issues Mol. Biol. 2022, 44, 6439-6449. https://doi.org/10.3390/cimb44120439
Obukhova L, Kopytova T, Murach E, Shchelchkova N, Kontorshchikova C, Medyanik I, Orlinskaya N, Grishin A, Kontorshchikov M, Badanina D. Glutathione and Its Metabolic Enzymes in Gliomal Tumor Tissue and the Peritumoral Zone at Different Degrees of Anaplasia. Current Issues in Molecular Biology. 2022; 44(12):6439-6449. https://doi.org/10.3390/cimb44120439
Chicago/Turabian StyleObukhova, Larisa, Tatiana Kopytova, Elena Murach, Natalya Shchelchkova, Claudia Kontorshchikova, Igor Medyanik, Natalia Orlinskaya, Artem Grishin, Michael Kontorshchikov, and Dariya Badanina. 2022. "Glutathione and Its Metabolic Enzymes in Gliomal Tumor Tissue and the Peritumoral Zone at Different Degrees of Anaplasia" Current Issues in Molecular Biology 44, no. 12: 6439-6449. https://doi.org/10.3390/cimb44120439
APA StyleObukhova, L., Kopytova, T., Murach, E., Shchelchkova, N., Kontorshchikova, C., Medyanik, I., Orlinskaya, N., Grishin, A., Kontorshchikov, M., & Badanina, D. (2022). Glutathione and Its Metabolic Enzymes in Gliomal Tumor Tissue and the Peritumoral Zone at Different Degrees of Anaplasia. Current Issues in Molecular Biology, 44(12), 6439-6449. https://doi.org/10.3390/cimb44120439