
The Transgene Expression of the Immature Form of the HCV Core Protein (C191) and the LncRNA MEG3 Increases Apoptosis in HepG2 Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Constructs and Cloning
2.2. Cell Culture and Transfection
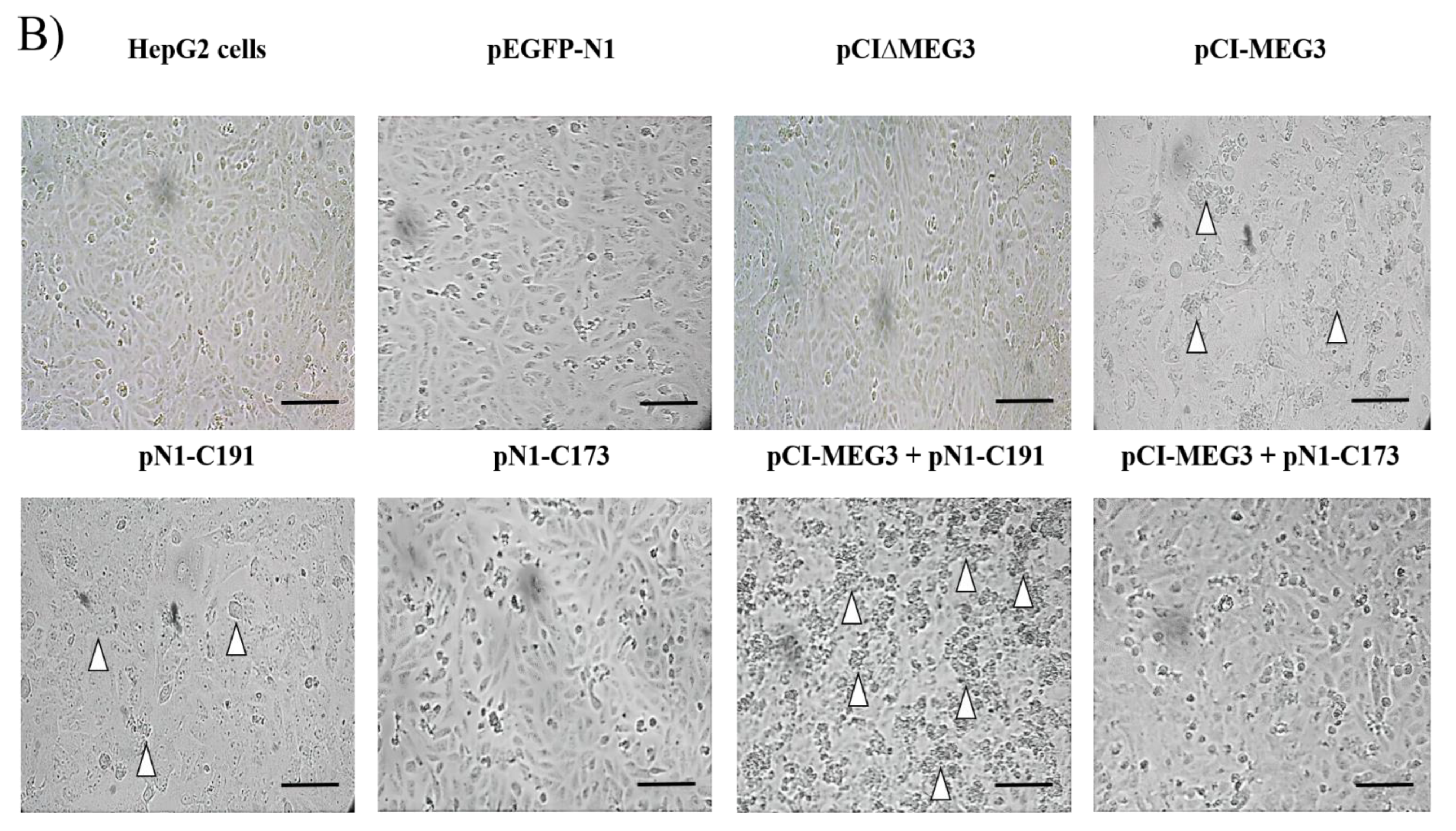
2.3. Examination of Cell Morphology
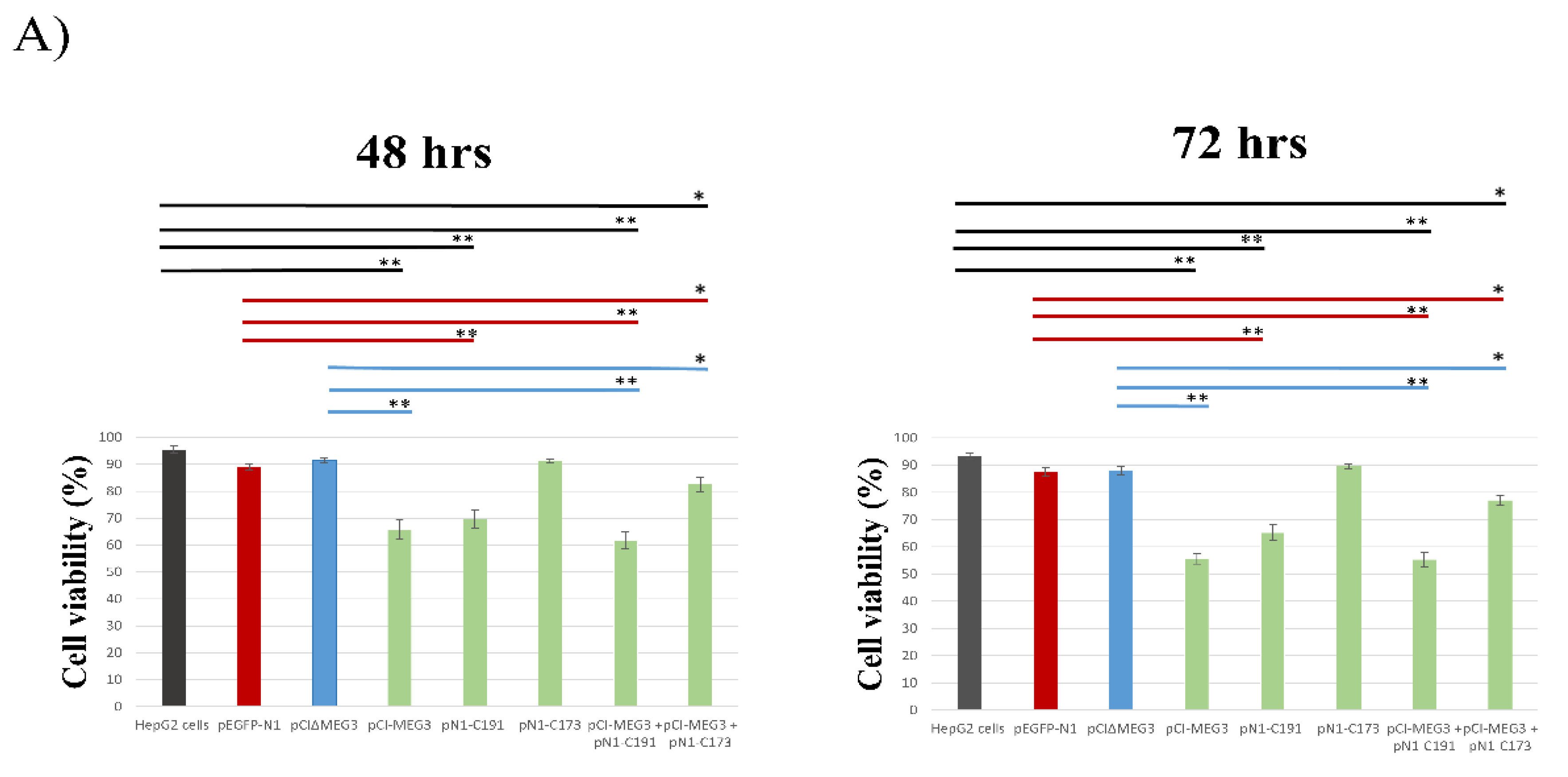
2.4. Cytotoxicity Analysis and Cell Viability by MTT Assay
2.5. Apoptosis Assay
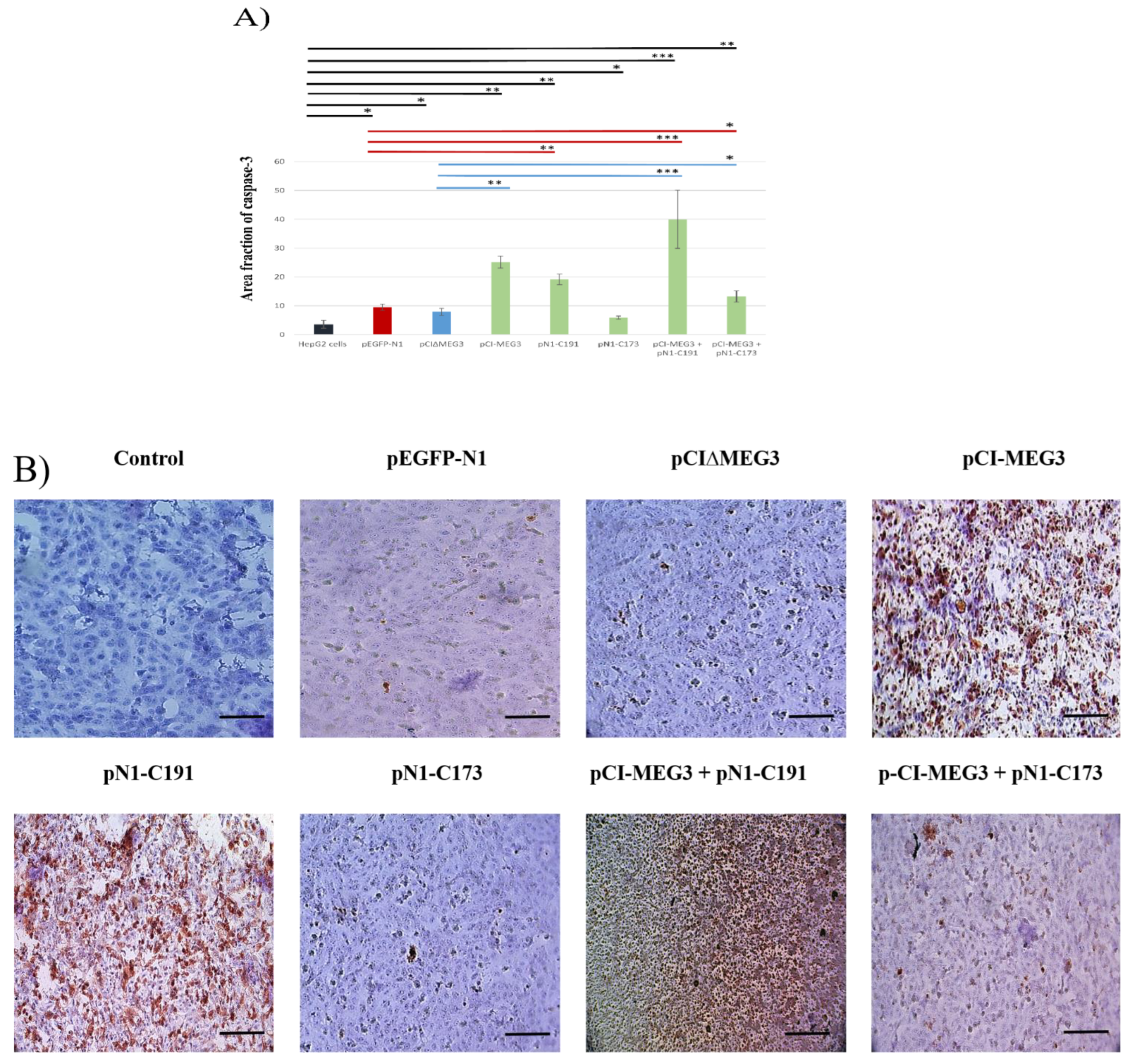
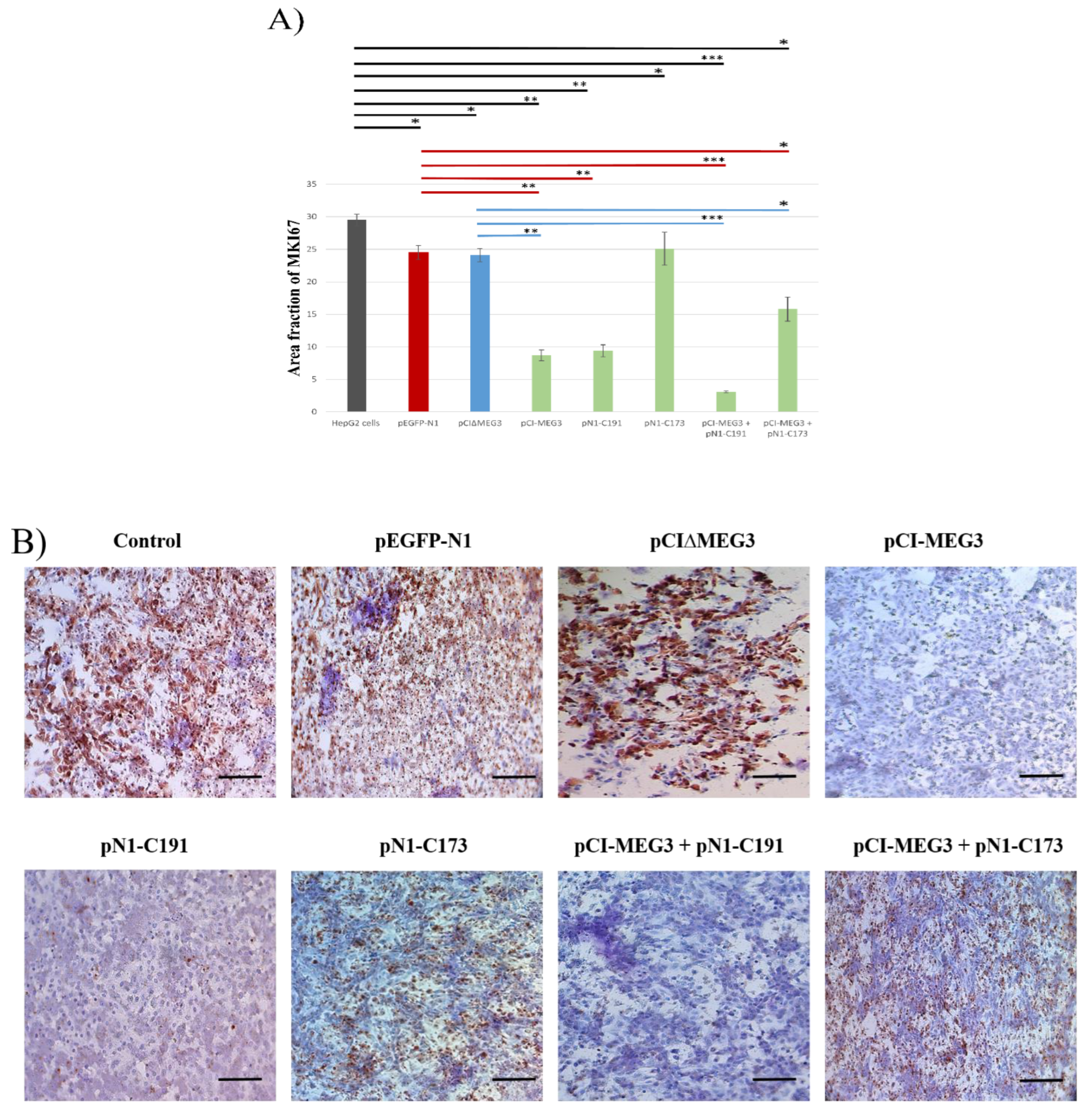
2.6. Immunocytochemistry for Caspase-3 and MKI67
2.7. Total RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
2.8. Statistical Analyses
3. Results
3.1. Overexpression of lncRNA MEG3 and Expression of C191 Reduce the Viability of HepG2 Cells
3.2. Co-Expression of lncRNA MEG3 and C191 Induces Apoptosis in HepG2 Cells
3.3. Co-Expression of lncRNA MEG3 and C191 Increases Caspase-3 and Decreases KI67 in HepG2 Cells
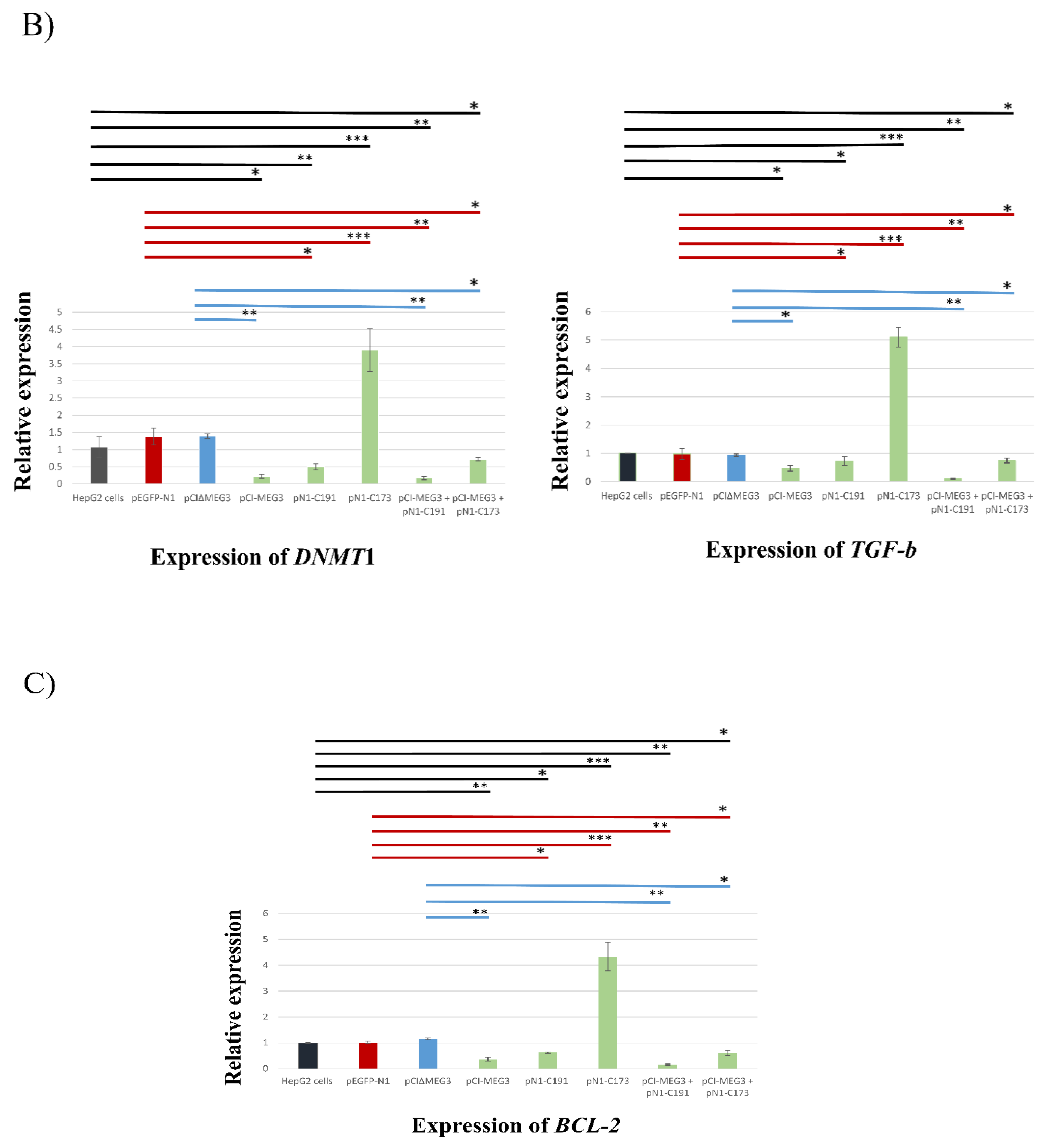
3.4. Co-Expression of lncRNA MEG3 and C191 Suppresses Proliferation of HepG2 Cells by Upregulating p53 and miRNA152 and Downregulation DNMT1, TGF-b, and BCL-2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Projections of Mortality and Causes of Death, 2016 to 2060. 2020. Available online: http://www.who.int/healthinfo/global_burden_disease/projections/en/ (accessed on 12 January 2022).
- Singal, A.; El-Serag, H. Hepatocellular Carcinoma from Epidemiology to Prevention: Translating Knowledge into Practice. Clin. Gastroenterol. Hepatol. 2015, 13, 2140–2151. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Sejima, H.; Ueda, Y.; Mori, K.; Satoh, S.; Dansako, H.; Ikeda, M. Genetic Characterization of Hepatitis C Virus in Long-Term RNA Replication Using Li23 Cell Culture Systems. PLoS ONE 2014, 9, e91156. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudvand, S.; Shokri, S.; Taherkhani, R.; Farshadpour, F. Hepatitis C virus core protein modulates several signaling pathways involved in hepatocellular carcinoma. World J. Gastroenterol. 2019, 25, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Bartosch, B.; Thimme, R.; Blum, H.; Zoulim, F. Hepatitis C virus-induced hepatocarcinogenesis. J. Hepatol. 2009, 51, 810–820. [Google Scholar] [CrossRef]
- Benali-Furet, N.; Chami, M.; Houel, L.; De Giorgi, F.; Vernejoul, F.; Lagorce, D.; Buscail, L.; Bartenschlager, R.; Ichas, F.; Rizzuto, R.; et al. Hepatitis C virus core triggers apoptosis in liver cells by inducing ER stress and ER calcium depletion. Oncogene 2005, 24, 4921–4933. [Google Scholar] [CrossRef]
- Okuda, M.; Li, K.; Beard, M.; Showalter, L.; Scholle, F.; Lemon, S.; Weinman, S.A. Mitochondrial injury, oxidative stress, and antioxidant gene expression are induced by hepatitis C virus core protein. Gastroenterology 2002, 122, 366–375. [Google Scholar] [CrossRef]
- Barba, G.; Harper, F.; Harada, T.; Kohara, M.; Goulinet, S.; Matsuura, Y.; Eder, G.; Schaff, Z.; Chapman, M.J.; Miyamura, T.; et al. Hepatitis C virus core protein shows a cytoplasmic localization and associates to cellular lipid storage droplets. Proc. Natl. Acad. Sci. USA 1997, 4, 1200–1205. [Google Scholar] [CrossRef]
- Santolini, E.; Migliaccio, G.; La Monica, N. Biosynthesis and biochemical properties of the hepatitis C virus core protein. J. Virol. 1994, 6, 3631–3641. [Google Scholar] [CrossRef]
- Pène, V.; Lemasson, M.; Harper, F.; Pierron, G.; Rosenberg, A. Role of cleavage at the core-E1 junction of hepatitis C virus polyprotein in viral morphogenesis. PLoS ONE 2017, 12, e0175810. [Google Scholar] [CrossRef]
- Lemberg, M.; Martoglio, B. Requirements for Signal Peptide Peptidase-Catalyzed Intramembrane Proteolysis. Mol. Cell 2002, 10, 735–744. [Google Scholar] [CrossRef]
- Suzuki, R.; Tamura, K.; Li, J.; Ishii, K.; Matsuura, Y.; Miyamura, T.; Suzuki, T. Ubiquitin-Mediated Degradation of Hepatitis C Virus Core Protein Is Regulated by Processing at Its Carboxyl Terminus. Virology 2001, 280, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Shiu, T.; Huang, S.; Shih, Y.; Chu, H.; Chang, W.; Hsieh, T. Hepatitis C Virus Core Protein Down-Regulates p21Waf1/Cip1 and Inhibits Curcumin-Induced Apoptosis through MicroRNA-345 Targeting in Human Hepatoma Cells. PLoS ONE 2013, 8, e61089. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Adachi, T.; Kitayama, K.; Bungyoku, Y.; Kitazawa, S.; Ishido, S.; Shoji, I.; Hotta, H. Hepatitis C Virus Infection Induces Apoptosis through a Bax-Triggered, Mitochondrion-Mediated, Caspase 3-Dependent Pathway. J. Virol. 2008, 82, 10375–10385. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Xie, Y.; Yang, P.; Chen, P.; Zhang, L. HCV Core Protein-Induced Down-Regulation of microRNA-152 Promoted Aberrant Proliferation by Regulating Wnt1 in HepG2 Cells. PLoS ONE 2014, 9, e81730. [Google Scholar] [CrossRef]
- Park, S.; Lim, J.; Lim, S.; Tiwari, I.; Jang, K. Hepatitis C virus Core protein stimulates cell growth by down-regulating p16 expression via DNA methylation. Cancer Lett. 2011, 310, 61–68. [Google Scholar] [CrossRef]
- Taniguchi, H.; Kato, N.; Otsuka, M.; Goto, T.; Yoshida, H.; Shiratori, Y.; Omata, M. Hepatitis C virus core protein upregulates transforming growth factor-?1 transcription. J. Med. Virol. 2003, 72, 52–59. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Q.; Liu, Y.; Luo, Z.; Kang, L.; Qu, J.; Liu, W.; Xia, X.; Liu, Y.; Wu, K.; et al. Hepatitis C Virus Activates Bcl-2 and MMP-2 Expression through Multiple Cellular Signaling Pathways. J. Virol. 2012, 86, 12531–12543. [Google Scholar] [CrossRef]
- Iyer, M.; Niknafs, Y.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S.; et al. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef]
- Huang, D.; Fang, J.; Luo, G. Roles of long noncoding RNAs in Hepatocellular Carcinoma. Open Life Sci. 2016, 11, 91–97. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Taheri, M. maternally expressed gene 3 (MEG3): A tumor suppressor long non coding RNA. Biomed. Pharmacother. 2019, 118, 109129. [Google Scholar] [CrossRef]
- Braconi, C.; Kogure, T.; Valeri, N.; Huang, N.; Nuovo, G.; Costinean, S.; Negrini, M.; Miotto, E.; Croce, C.M.; Patel, T. microRNA-29 can regulate expression of the long non-coding RNA gene MEG3 in hepatocellular cancer. Oncogene 2011, 30, 4750–4756. [Google Scholar] [CrossRef]
- He, Z.; Yang, D.; Fan, X.; Zhang, M.; Li, Y.; Gu, X.; Yang, M. The Roles and Mechanisms of lncRNAs in Liver Fibrosis. Int. J. Mol. Sci. 2020, 21, 1482. [Google Scholar] [CrossRef]
- Wu, D.; Ma, Z.; Ma, D.; Li, Q. Long non-coding RNA maternally expressed gene 3 affects cell proliferation, apoptosis and migration by targeting the microRNA-9-5p/midkine axis and activating the phosphoinositide-dependent kinase/AKT pathway in hepatocellular carcinoma. Oncol. Lett. 2021, 21, 345. [Google Scholar] [CrossRef]
- Tao, X.; Zeng, L.; Wang, H.; Liu, H. LncRNA MEG3 ameliorates respiratory syncytial virus infection by suppressing TLR4 signaling. Mol. Med. Rep. 2017, 2, 23–35. [Google Scholar] [CrossRef]
- Turjya, R.; Khan, M.A.; Khademul, M.; Islam, A.B. Perversely expressed long noncoding RNAs can alter host response and viral proliferation in SARS-CoV-2 infection. Future Virol. 2020, 15, 577–593. [Google Scholar] [CrossRef]
- Huang, J.; Wang, Y.; Guo, Y.; Sun, S. Down-regulated microRNA-152 induces aberrant DNA methylation in hepatitis B virus-related hepatocellular carcinoma by targeting DNA methyltransferase 1. Hepatology 2010, 52, 60–70. [Google Scholar] [CrossRef]
- He, Y.; Dan, Y.; Gao, X.; Huang, L.; Lv, H.; Chen, J. DNMT1-mediated lncRNA MEG3 methylation accelerates endothelial-mesenchymal transition in diabetic retinopathy through the PI3K/Akt/mTOR signaling pathway. Am. J. Physiol. Endocrinol. Metab. 2021, 20, E598–E608. [Google Scholar] [CrossRef]
- Huang, Z.; Chen, R.; Zhou, X.; Zhan, H.; Hu, M.; Liu, B.; Wu, G.; Wu, L. Long non-coding RNA MEG3 induces cell apoptosis in esophageal cancer through endoplasmic reticulum stress. Oncol. Rep. 2017, 37, 3093–3099. [Google Scholar] [CrossRef]
- Aladdin, A.; Sahly, N.; Faty, R.; Youssef, M.M.; Salem, T.Z. The Baculovirus Promoter Opie2 Sequence Has Inhibitory Effect on The Activity of The Cytomegalovirus (CMV) Promoter in Hela and HEK-293 T Cells. Gene 2021, 781, 145541. [Google Scholar] [CrossRef]
- Ahmed, W.; Mofed, D.; Zekri, A.; El-Sayed, N.; Rahouma, M.; Sabet, S. Antioxidant activity and apoptotic induction as mechanisms of action of Withania somnifera (Ashwagandha) against a hepatocellular carcinoma cell line. J. Int. Med. Res. 2018, 46, 1358–1369. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Bashmail, H.; Alamoudi, A.; Noorwali, A.; Hegazy, G.; AJabnoor, G.; Choudhry, H.; Al-Abd, A.M. Thymoquinone synergizes gemcitabine anti-breast cancer activity via modulating its apoptotic and autophagic activities. Sci. Rep. 2018, 8, 11674. [Google Scholar] [CrossRef] [PubMed]
- Baiomy, A.A.; Ahmed, A.M. Genetic and Histopathological Responses to Cadmium Toxicity in Rabbit’S Kidney and Liver: Protection by Ginger (Zingiber Officinale). Biol. Trace Elem. Res. 2015, 170, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Rizzardi, A.; Johnson, A.; Vogel, R.; Pambuccian, S.; Henriksen, J.; Skubitz, A.; Metzger, G.J.; Schmechel, S.C. Quantitative comparison of immunohistochemical staining measured by digital image analysis versus pathologist visual scoring. Diagn. Pathol. 2012, 7, 42. [Google Scholar] [CrossRef]
- Cuykendall, T.; Rubin, M.; Khurana, E. Non-coding genetic variation in cancer. Curr. Opin. Syst. Biol. 2017, 1, 9–15. [Google Scholar] [CrossRef]
- Chen, R.; Huang, Z.; Liu, L.; Xiang, M.; Li, G.; Feng, J.; Liu, B.; Wu, L.-F. Involvement of endoplasmic reticulum stress and p53 in lncRNA MEG3-induced human hepatoma HepG2 cell apoptosis. Oncol. Rep. 2016, 36, 1649–1657. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, S.; Ye, F.; Shen, Y.; Tie, Y.; Zhu, J.; Wei, L.; Jin, Y.; Fu, H.; Wu, Y.; et al. Long Noncoding RNA MEG3 Interacts with p53 Protein and Regulates Partial p53 Target Genes in Hepatoma Cells. PLoS ONE 2015, 10, e0139790. [Google Scholar] [CrossRef]
- Shen, S.; Li, C.; Dai, M.; Yan, X. Induction of Huh 7 cell apoptosis by HCV core proteins via CK1α p53 Bid signaling pathway. Mol. Med. Rep. 2018, 23, 123–135. [Google Scholar] [CrossRef]
- Herzer, K.; Weyer, S.; Krammer, P.; Galle, P.; Hofmann, T. Hepatitis C Virus Core Protein Inhibits Tumor Suppressor Protein Promyelocytic Leukemia Function in Human Hepatoma Cells. Cancer Res. 2005, 65, 10830–10837. [Google Scholar] [CrossRef]
- Vilchez, V.; Turcios, L.; Marti, F.; Gedaly, R. Targeting Wnt/β-catenin pathway in hepatocellular carcinoma treatment. World J. Gastroenterol. 2016, 22, 823–832. [Google Scholar] [CrossRef]
- Behesti, H.; Marino, S. Cerebellar granule cells: Insights into proliferation, differentiation, and role in medulloblastoma pathogenesis. Int. J. Biochem. Cell Biol. 2009, 41, 435–445. [Google Scholar] [CrossRef]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.R.; Hoberg, E.; et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA–DNA triplex structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Lv, Y.; Zhang, C.; Guo, S. LncRNA meg3 suppresses hepatocellular carcinoma in vitro and vivo studies. Am. J. Transl. Res. 2019, 11, 4089–4099. [Google Scholar]
- Li, L.T.; Jiang, G.; Chen, Q.; Zheng, J.N. Ki67 Is a Promising Molecular Target in yhe Diagnosis of Cancer (Review). Mol. Med. Rep. 2014, 11, 1566–1572. [Google Scholar] [CrossRef]
- Liu, J.; Ding, X.; Tang, J.; Cao, Y.; Hu, P.; Zhou, F.; Shan, X.; Cai, X.; Chen, Q.; Ling, N.; et al. Enhancement of Canonical Wnt/β-Catenin Signaling Activity by HCV Core Protein Promotes Cell Growth of Hepatocellular Carcinoma Cells. PLoS ONE 2011, 6, e27496. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| MEG3 | 5′CCACTCCCAGTTCAATTACAGCTC3′ | 5′TAGTGCCCTCGTGAGGTGTAG3′ |
| P53 | 5′CAGCACATGACGGAGGTTGT3′ | 5′TCATCCAAATACTCCACACG3′ |
| DNMT1 | 5′CCATCAGGCATTCTACCA3′ | 5′CGTTCTCCTTGTCTTCTCT3′ |
| miRNA152 | 5′CCCAGGTTCTGTGATACACTCC3′ | 5′CTTCCGGGCCCAAGTTCTG3′ |
| TGF-B | 5′AAGAAGTCACCCGCGTGCTA3′ | 5′TGTGTGATGTCTTTGGTTTTGTCA3′ |
| BCL-2 | 5′GGTGGGGTCATGTGTGTGG3′ | 5′CGGTTCAGGTACTCAGTCATCC3′ |
| HCVcore 191 or 173 | 5′GATACATCCCGCTCGTAGGC3′ | 5′TTGATCCCGTCCTCCAAAGC3′ |
| β-Actin | 5′CATGGAGTCCTGTGGCATC3′ | 5′CAGGGCAGTGATCTCCTTCT3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mofed, D.; Sabet, S.; Baiomy, A.A.; Salem, T.Z. The Transgene Expression of the Immature Form of the HCV Core Protein (C191) and the LncRNA MEG3 Increases Apoptosis in HepG2 Cells. Curr. Issues Mol. Biol. 2022, 44, 3632-3647. https://doi.org/10.3390/cimb44080249
Mofed D, Sabet S, Baiomy AA, Salem TZ. The Transgene Expression of the Immature Form of the HCV Core Protein (C191) and the LncRNA MEG3 Increases Apoptosis in HepG2 Cells. Current Issues in Molecular Biology. 2022; 44(8):3632-3647. https://doi.org/10.3390/cimb44080249
Chicago/Turabian StyleMofed, Dina, Salwa Sabet, Ahmed A. Baiomy, and Tamer Z. Salem. 2022. "The Transgene Expression of the Immature Form of the HCV Core Protein (C191) and the LncRNA MEG3 Increases Apoptosis in HepG2 Cells" Current Issues in Molecular Biology 44, no. 8: 3632-3647. https://doi.org/10.3390/cimb44080249