
Neuroinflammation in the Evolution of Motor Function in Stroke and Trauma Patients: Treatment and Potential Biomarkers

 , , , and
, , , and
Abstract
:
1. Introduction
2. Neuroinflammation
2.1. Stroke
- -
- Ischemic stroke (80%): Occurs because of a decrease and insufficiency of blood supply to the CNS, causing a circumscribed area of cerebral infarction. Depending on their etiology, strokes can be subclassified as thrombotic due to the formation of a blood clot in an area of the brain and embolic because of the formation of a blood clot in another cerebral artery that subsequently travels to the brain. When the symptoms last less than 24 h, it is called a transient ischemic attack (TIA [35,36]).
- -
- Hemorrhagic stroke (20%): Is due to parenchymal and/or subarachnoid bleeding. Generally, they are caused by arterial hypertension (AHT), aneurysm ruptures, and or arteriovenous malformations [37].
Neuroinflammation and Stroke
2.2. Spinal Cord Injury
- -
- Cause: Traumatic or non-traumatic.
- -
- Mechanism of injury: Hyperflexion, flexion with rotation, hyperextension, or compression.
- -
- Level of injury: Cervical, dorsal, or/and lumbosacral.
- -
- Extension: Complete or incomplete.
Neuroinflammation and Spinal Cord Injury
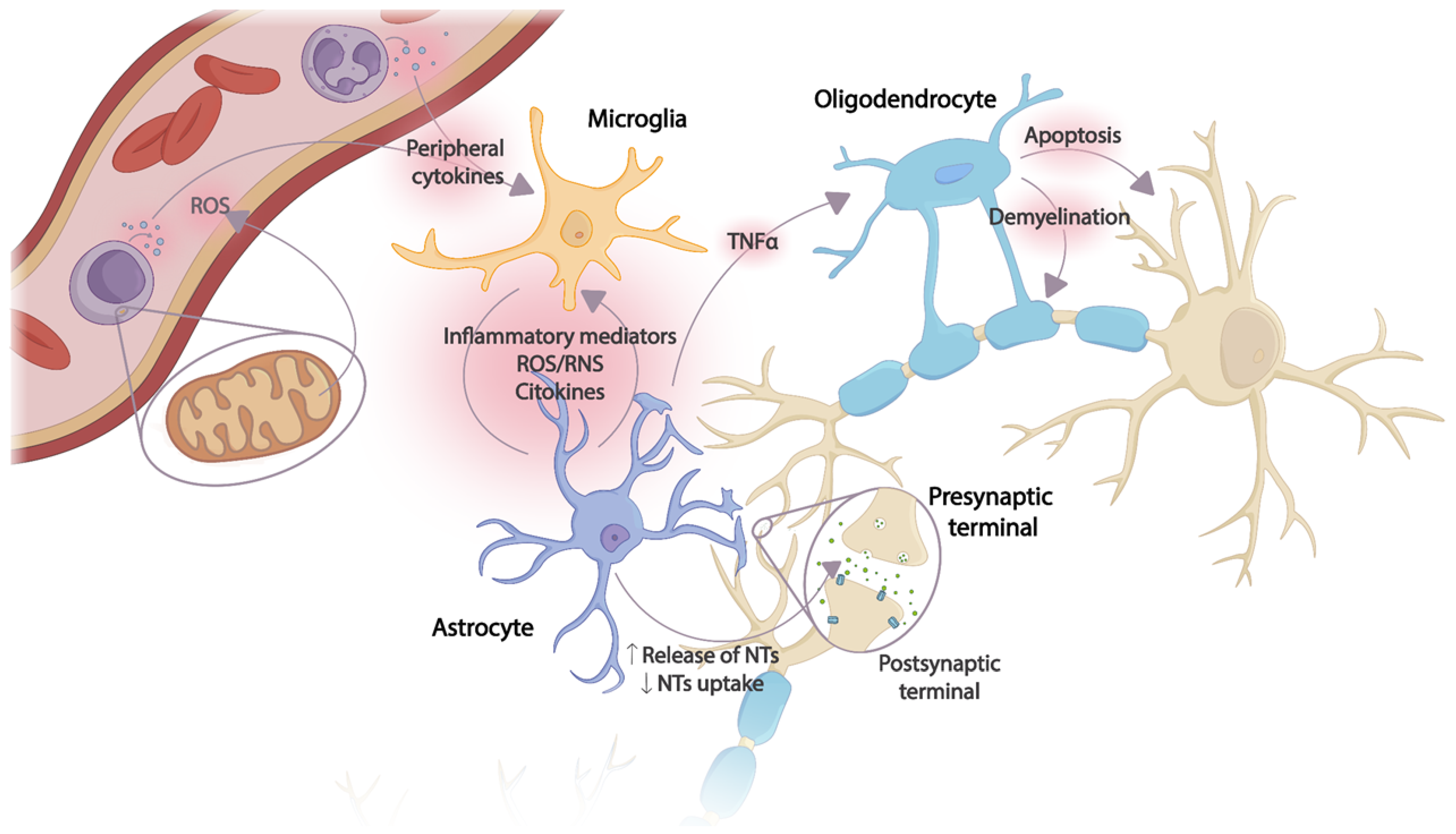
2.3. Neuroinflammation and Mitochondrial Activity
2.4. Cytokines and Chemokines Involved in Neuroinflammation
3. Diagnostic Techniques
3.1. Biomarkers in Neuroinflammation
3.1.1. Biological Markers
3.1.2. Techniques for Biomarkers Evaluation
3.2. Neuroimaging Technologies
3.2.1. Positron Emission Tomography
3.2.2. Magnetic Resonance Imaging
3.2.3. Cerebral Vascular Permeability Magnetic Resonance Imaging
3.2.4. Computed Tomography
3.2.5. Contrast-Enhanced Ultrasound
4. Treatment Techniques
4.1. Electrical Stimulation
4.2. Bionanomaterials
- -
- Drug loading: takes place in the interior or membrane, depending on the solubility of the drugs. It involves the encapsulation of hydrophobic or hydrophilic therapeutic agents. For this, it is necessary to select the most suitable nanoparticle for the application, which depends on factors such as the chemical naturalization of the drug, the desired release, and the mechanism of administration [230]. The loading method chosen depends on the solubility and properties of the therapeutic agent:
- o
- Dissolution and diffusion method: For hydrophobic drugs. Dissolution occurs in the lipid core or polymer matrix during the manufacture of the nanoparticle [231].
- o
- Encapsulation method: For hydrophilic drugs, which are encapsulated in the aqueous core. For this, the drug and the nanoparticle material are emulsified in an organic solvent, and then the solvent is removed [232].
- o
- Surface adsorption: For small molecules or substances with an affinity for the surface of the nanoparticle material. The drug is adsorbed directly on the surface [232].
- o
- Physical methods: Co-precipitation involves the simultaneous formation of nanoparticles and drug precipitates within the nanoparticles during a physical or chemical process [233]. Freeze-drying is based on rapid freezing followed by the removal of the solvent by vacuum sublimation. It is especially useful in thermosensitive drugs as it avoids exposure to high temperatures and produces nanoparticles with high stability and long shelf life [234].
- -
- Administration: Generally, they are administered via intravenous or local injection into the area affected by neuroinflammation [19].
- -
- Targeting of inflamed areas: They are specifically designed to target an area of the brain affected by neuroinflammation to increase treatment efficacy and reduce potential side effects. To this end, the nanoparticle surface is functionalized with specific markers that may include proteins, receptors, or adhesion molecules that are expressed in greater quantities on inflammatory cells [19]. For this purpose, the nanoparticle surface is modified with ligands or antibodies that recognize receptors expressed on inflammatory cells or BBB vessel endothelium. After binding to inflammatory cells, they are internalized through endocytosis processes [235].
- -
- Controlled release: Drug release is performed gradually to prolong the therapeutic effect and reduce the need for frequent dosing. It is accomplished via modification of the nanoparticle matrix, selection of specific polymers, or surface engineering [236]. The release can be sustained or targeted to a specific response in the body. In the case of sustained release, the aim is to maintain a constant and effective concentration of therapeutic agents at the site of action over a prolonged period [237]. In targeted release, the specific delivery of a therapeutic agent to a precise and selective target in the body is sought [236,237].
- -
- Anti-inflammatory action: Released drugs act on inflammatory cells, reducing the response. They can inhibit the production of proinflammatory cytokines using anti-inflammatory drugs or specific molecules that block cytokine signaling [238], such as TNF-α inhibitors [238]. Free radicals can also be neutralized by encapsulating antioxidants or free radical scavengers, such as vitamin E or vitamin C, reducing oxidative stress and protecting cells and tissues [239]. In turn, the activity of immune cells can also be modulated by immunomodulating agents that regulate the immune response, such as corticosteroids [240].
- -
- Biodegradation: They are broken down into non-toxic products and eliminated from the body naturally. Biodegradation can take place using different mechanisms: hydrolysis, hepatic metabolism, phagocytosis by phagocytic cells, and lipid exchange [241].
Nanoparticle Types
- -
- Lipid nanoparticles: are a type of bionanomaterial composed of a lipid bilayer surrounding an aqueous or lipid core, forming a membrane-like structure surrounding the drug core. They are usually between 0.05 and 5 nanometers in size, allowing them to be administered at the cellular and subcellular levels [242]. The advantage of this type of nanoparticles is that they are insoluble in water, so they can be encapsulated in the lipid core of the nanoparticles, increasing solubility and bioavailability [243]. In addition, degradation in the biological environment is avoided, and their stability is improved [229].
- o
- Liposomes: Lipid vesicles that are composed of a lipid bilayer surrounding an internal water cavity. The bilayer is composed of two layers of lipid molecules, with lipid tail structures towards the center and the heads towards the outside. This type of amphiphilic structure allows liposomes to be compatible with hydrophobic and hydrophilic substances. Depending on the manufacturing conditions and composition, different types of liposomes can be obtained. Unimamellar liposomes (LUV) are used for gene therapy, multi-mamellar liposomes (MLV) are used for research and pharmaceutical applications with high drug loads, and finally, miscellaneous liposomes are used for the release of drugs at different rates or locations within the organism [244,245].
- o
- Lipid micelles: Nanometric structures formed by lipid molecules arranged in the form of micelles. They do not have an internal aqueous cavity since the lipid heads are oriented outward and the lipid tails inward in a spherical structure [246].
- -
- Polymeric nanoparticles: Colloidal systems composed of polymers. They are manufactured using different methods, such as emulsification, solvent evaporation, emulsion polymerization, and nanoprecipitation [229,247]. Their size usually does not exceed 100 nanometers. Polyethylene glycol (PEG) nanoparticles are used to improve the stability and bioavailability of neuroinflammation-related drugs [248]. The advantage they offer is the ability to synthesize them with precise and controlled sizes, ensuring uniform size distribution, as well as their high biocompatibility.
- o
- PEG-coated gold nanoparticles: Ability to cross the BBB [248].
- o
- PEG-coated dendrimer nanoparticles: Dendrimers are branched polymers that, when coated with PEG, improve their circulation properties and reduce immunogenicity, facilitating their arrival in the brain [249].
- o
- Poly(lactide-co-glycolic) (PLGA) nanoparticles coated with PEG: PLGA has a great capacity to degrade into biocompatible products and be eliminated naturally by the body, reducing toxicity and gradual release of the encapsulated drug. Being PEG-coated increases their stability and time in circulation [250].
- o
- PEG-coated liposomes and micelles: Lipid vesicles that the PEG coating makes them more stable [245].
- o
- Hydrogel nanoparticles: Composed of hydrophilic polymers such as PEG or arginine, which are chemically cross-linked to form a three-dimensional network structure. Significant amounts of water are retained within the network, conferring gel-like properties [251].
- -
- Iron oxide nanoparticles (NPOH): Composed of iron oxide crystals, generally magnetite (FE3O4). Their diameter can vary depending on the desired application but never exceeds the nanometer scale [252]. Due to their high magnetic susceptibility, they can interact with external magnetic fields, which makes them useful in medical imaging applications by generating intense MRI signals, as well as in magnetic hyperthermia therapy, inducing inflammatory cell death, or activating therapies in combination with controlled drug release [253]. In addition, they can also be functionalized with antibodies or specific molecules to detect biomarkers of neuroinflammation, thus enabling early identification of inflammation and monitoring of the response to such treatments [253,254].
- o
- Super magnetic iron oxide nanoparticles (SPIONs): Used in MR techniques to visualize areas of brain inflammation and diagnosis of neuroinflammatory diseases [255].
- o
- Multifunctional iron oxide nanoparticles: Controlled release of anti-inflammatory drugs or magnetic hyperthermia [256].
- o
- o
- Iron oxide nanoparticles for gene transport: Delivery of therapeutic genes to areas of brain inflammation. Functionalized with DNA or RNA sequences that regulate the expression of genes involved in neuroinflammation [256].
- -
- Silica nanoparticles (SNP): Nanoparticles composed mainly of silicon dioxide (SiO2) that can be synthesized as spheres, nanocapsules, nanotubes, and/or complex structures. The internal structure of NPS can be porous or non-porous, depending on the desired application. Their main advantage is the ability to combine diagnostics and therapy by being able to function as contrast agents and therapy delivery simultaneously [257,258].
- o
- Mesoporous silica nanoparticles (MSN): They present a porous structure with internal channels that allow for greater drug loading capacity and sustained release [259].
- o
- Silica nanoparticles functionalized with antibodies or peptides: Allows specific binding to biomarkers or cells [260].
- o
- Magnetic silica nanoparticles: Contain an iron oxide core coated with silica, which allows them to be guided to areas of the brain by means of external magnetic fields [260].
- o
- Antioxidant-loaded silica nanoparticles: To combat oxidative damage.
- o
- Silica nanoparticles with imaging agents: Incorporate fluor surfactants or contrast agents for MRI [261].
- o
- Multifunctional silica nanoparticles: Combine several features [257].
- -
- Protein nanoparticles: Composed mainly of proteins or peptides that can be natural or specifically designed for the application. Their size generally ranges from 1 to 100 nanometers, and their morphology can be spherical, nanotubes, or vesicles [262].
- o
- Albumin nanoparticles: Biocompatible and long circulation half-life [263].
- o
- High-density lipoprotein (HDL) nanoparticles: Cross the BBB [264].
- o
- Functionalized peptide nanoparticles: Precise delivery of therapeutic agent and reduction in side effects [262].
- o
- Immunoglobulin G (IgG) nanoparticles: Modulation of immune response in CSN [265].
5. Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Breda, C.N.d.S.; Davanzo, G.G.; Basso, P.J.; Saraiva Câmara, N.O.; Moraes-Vieira, P.M.M. Mitochondria as Central Hub of the Immune System. Redox Biol. 2019, 26, 101255. [Google Scholar] [CrossRef] [PubMed]
- Lukacova, N.; Kisucka, A.; Bimbova, K.K.; Bacova, M.; Ileninova, M.; Kuruc, T.; Galik, J. Glial-Neuronal Interactions in Pathogenesis and Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2021, 22, 13577. [Google Scholar] [CrossRef]
- Silva, N.A.; Sousa, N.; Reis, R.L.; Salgado, A.J. From Basics to Clinical: A Comprehensive Review on Spinal Cord Injury. Prog. Neurobiol. 2014, 114, 25–57. [Google Scholar] [CrossRef]
- Anderson, M.A.; Ao, Y.; Sofroniew, M.V. Heterogeneity of Reactive Astrocytes. Neurosci. Lett. 2014, 565, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, M.A.; Lo, E.H.; Iadecola, C. The Science of Stroke: Mechanisms in Search of Treatments. Neuron 2010, 67, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Kamel, H.; Iadecola, C. Brain-Immune Interactions and Ischemic Stroke: Clinical Implications. Arch. Neurol. 2012, 69, 576–581. [Google Scholar] [CrossRef]
- Anthony, S.; Cabantan, D.; Monsour, M.; Borlongan, C.V. Neuroinflammation, Stem Cells, and Stroke. Stroke 2022, 53, 1460–1472. [Google Scholar] [CrossRef]
- Sillerud, L.O.; Yang, Y.; Yang, L.Y.; Duval, K.B.; Thompson, J.; Yang, Y. Longitudinal Monitoring of Microglial/Macrophage Activation in Ischemic Rat Brain Using Iba-1-Specific Nanoparticle-Enhanced Magnetic Resonance Imaging. J. Cereb. Blood Flow. Metab. 2020, 40, S117–S133. [Google Scholar] [CrossRef]
- Campbell, B.C.V. Advances in Stroke Medicine. Med. J. Aust. 2019, 210, 367–374. [Google Scholar] [CrossRef]
- Campbell, B.C.V.; Mitchell, P.J.; Kleinig, T.J.; Dewey, H.M.; Churilov, L.; Yassi, N.; Yan, B.; Dowling, R.J.; Parsons, M.W.; Oxley, T.J.; et al. Endovascular Therapy for Ischemic Stroke with Perfusion-Imaging Selection. N. Engl. J. Med. 2015, 372, 1009–1018. [Google Scholar] [CrossRef]
- Kunte, H.; Schmidt, C.; Harms, L.; Rückert, R.I.; Grigoryev, M.; Fischer, T. Contrast-Enhanced Ultrasound and Detection of Carotid Plaque Neovascularization. Neurology 2012, 79, 2081. [Google Scholar] [CrossRef]
- Candelario-Jalil, E.; Dijkhuizen, R.M.; Magnus, T. Neuroinflammation, Stroke, Blood-Brain Barrier Dysfunction, and Imaging Modalities. Stroke 2022, 53, 1473–1486. [Google Scholar] [CrossRef] [PubMed]
- Sonawane, M.D.; Nimse, S.B. C-Reactive Protein: A Major Inflammatory Biomarker. Anal. Methods 2017, 9, 3400–3413. [Google Scholar] [CrossRef]
- Strimbu, K.; Tavel, J.A. What Are Biomarkers? Curr. Opin. HIV AIDS 2010, 5, 463–466. [Google Scholar] [CrossRef]
- Simats, A.; García-Berrocoso, T.; Montaner, J. Neuroinflammatory Biomarkers: From Stroke Diagnosis and Prognosis to Therapy. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1862, 411–424. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, M.; Rodríguez-Agirretxe, I.; Vecino, E.; Astigarraga, E.; Acera, A.; Barreda-Gómez, G. Elevation of Tear MMP-9 Concentration as a Biomarker of Inflammation in Ocular Pathology by Antibody Microarray Immunodetection Assays. Int. J. Mol. Sci. 2022, 23, 5639. [Google Scholar] [CrossRef]
- Lennikov, A.; Yang, M.; Chang, K.; Pan, L.; Saddala, M.S.; Lee, C.; Ashok, A.; Cho, K.S.; Utheim, T.P.; Chen, D.F. Direct Modulation of Microglial Function by Electrical Field. Front. Cell Dev. Biol. 2022, 10, 980775. [Google Scholar] [CrossRef]
- Schuhmann, M.K.; Papp, L.; Stoll, G.; Blum, R.; Volkmann, J.; Fluri, F. Mesencephalic Electrical Stimulation Reduces Neuroinflammation after Photothrombotic Stroke in Rats by Targeting the Cholinergic Anti-Inflammatory Pathway. Int. J. Mol. Sci. 2021, 22, 1254. [Google Scholar] [CrossRef]
- Lee, C.Y.P.; Chooi, W.H.; Ng, S.Y.; Chew, S.Y. Modulating Neuroinflammation through Molecular, Cellular and Biomaterial-Based Approaches to Treat Spinal Cord Injury. Bioeng. Transl. Med. 2022, 8, e10389. [Google Scholar] [CrossRef]
- Song, G.; Zhao, M.; Chen, H.; Lenahan, C.; Zhou, X.; Ou, Y.; He, Y. The Role of Nanomaterials in Stroke Treatment: Targeting Oxidative Stress. Oxid. Med. Cell. Longev. 2021, 2021, 8857486. [Google Scholar] [CrossRef]
- Kim, T.H.; Kang, M.S.; Mandakhbayar, N.; El-Fiqi, A.; Kim, H.W. Anti-Inflammatory Actions of Folate-Functionalized Bioactive Ion-Releasing Nanoparticles Imply Drug-Free Nanotherapy of Inflamed Tissues. Biomaterials 2019, 207, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Shcharbina, N.; Shcharbin, D.; Bryszewska, M. Nanomaterials in Stroke Treatment: Perspectives. Stroke 2013, 44, 2351–2355. [Google Scholar] [CrossRef] [PubMed]
- Pottorf, T.S.; Rotterman, T.M.; McCallum, W.M.; Haley-Johnson, Z.A.; Alvarez, F.J. The Role of Microglia in Neuroinflammation of the Spinal Cord after Peripheral Nerve Injury. Cells 2022, 11, 2083. [Google Scholar] [CrossRef]
- Miller, A.H.; Maletic, V.; Raison, C.L. Inflammation and Its Discontents: The Role of Cytokines in the Pathophysiology of Major Depression. Biol. Psychiatry 2009, 65, 732–741. [Google Scholar] [CrossRef]
- Kölliker-Frers, R.; Udovin, L.; Otero-Losada, M.; Kobiec, T.; Herrera, M.I.; Palacios, J.; Razzitte, G.; Capani, F. Neuroinflammation: An Integrating Overview of Reactive-Neuroimmune Cell Interactions in Health and Disease. Mediat. Inflamm. 2021, 2021, 9999146. [Google Scholar] [CrossRef] [PubMed]
- Shastri, A.; Bonifati, D.M.; Kishore, U. Innate Immunity and Neuroinflammation. Mediators Inflamm. 2013, 2013, 342931. [Google Scholar] [CrossRef]
- Wang, D.S.; Zurek, A.A.; Lecker, I.; Yu, J.; Abramian, A.M.; Avramescu, S.; Davies, P.A.; Moss, S.J.; Lu, W.Y.; Orser, B.A. Memory Deficits Induced by Inflammation Are Regulated by A5-Subunit-Containing GABAA Receptors. Cell Rep. 2012, 2, 488–496. [Google Scholar] [CrossRef]
- Leboyer, M.; Soreca, I.; Scott, J.; Frye, M.; Henry, C.; Tamouza, R.; Kupfer, D.J. Can Bipolar Disorder Be Viewed as a Multi-System Inflammatory Disease? J. Affect. Disord. 2012, 141, 1–10. [Google Scholar] [CrossRef]
- Muller, N.; Myint, A.-M.J.; Schwarz, M. Kynurenine Pathway in Schizophrenia: Pathophysiological and Therapeutic Aspects. Curr. Pharm. Des. 2011, 17, 130–136. [Google Scholar] [CrossRef]
- McNally, L.; Bhagwagar, Z.; Hannestad, J. Inflammation, Glutamate, and Glia in Depression: A Literature Review. CNS Spectr. 2008, 13, 501–510. [Google Scholar] [CrossRef]
- O’Connor, J.C.; André, C.; Wang, Y.; Lawson, M.A.; Szegedi, S.S.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Interferon-Gamma and Tumor Necrosis Factor-Alpha Mediate the Upregulation of Indoleamine 2,3-Dioxygenase and the Induction of Depressive-like Behavior in Mice in Response to Bacillus Calmette-Guerin. J. Neurosci. 2009, 29, 4200–4209. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, C.C.; Ushakova, A.; Skogseth, R.E.; Alves, G.; Tysnes, O.B.; Aarsland, D.; Lange, J.; Maple-Grødem, J. Inflammatory Biomarkers in Newly Diagnosed Patients With Parkinson Disease and Related Neurodegenerative Disorders. Neurol. Neuroimmunol. Neuroinflamm. 2023, 10, e200132. [Google Scholar] [CrossRef] [PubMed]
- Inicio|NINDS Español. Available online: https://espanol.ninds.nih.gov/es (accessed on 5 September 2023).
- Berru Loayza, K.F. Factores Pronósticos de Morbilidad y Secuelas Del Accidente Cerebrovascular En Adultos Mayores; Universidad Católica de Cuenca: Cuenca, Ecuador, 2021. [Google Scholar]
- Ruiz-Mejía, A.F.; Pérez-Romero, G.E.; Ángel-Macías, M.A.; Ruiz-Mejía, A.F.; Pérez-Romero, G.E.; Ángel-Macías, M.A. Ataque Cerebrovascular Isquémico: Fisiopatología Desde El Sistema Biomédico y Su Equivalente En La Medicina Tradicional China. Rev. Fac. Med. 2017, 65, 137–144. [Google Scholar] [CrossRef]
- Chaves Sell, F. Accidente Vascular Cerebral: ¿es El Accidente Vascular Cerebral Una Enfermedad Tratable? Rev. Costarric. Cardiol. 2000, 2, 27–33. [Google Scholar]
- Harrison. Principios de Medicina Interna, 21e|AccessMedicina|McGraw Hill Medical. Available online: https://accessmedicina.mhmedical.com/book.aspx?bookID=3118 (accessed on 5 September 2023).
- Teresa, P.; Ribeiro, C.; Rio, S. Manual de Patologia Bucal; FAPERJ: Rio de Janeiro, Brazil, 2013. [Google Scholar]
- Gelderblom, M.; Leypoldt, F.; Steinbach, K.; Behrens, D.; Choe, C.U.; Siler, D.A.; Arumugam, T.V.; Orthey, E.; Gerloff, C.; Tolosa, E.; et al. Temporal and Spatial Dynamics of Cerebral Immune Cell Accumulation in Stroke. Stroke 2009, 40, 1849–1857. [Google Scholar] [CrossRef]
- Yilmaz, G.; Arumugam, T.V.; Stokes, K.Y.; Granger, D.N. Role of T Lymphocytes and Interferon-Gamma in Ischemic Stroke. Circulation 2006, 113, 2105–2112. [Google Scholar] [CrossRef]
- Walsh, J.T.; Hendrix, S.; Boato, F.; Smirnov, I.; Zheng, J.; Lukens, J.R.; Gadani, S.; Hechler, D.; Gölz, G.; Rosenberger, K.; et al. MHCII-Independent CD4+ T Cells Protect Injured CNS Neurons via IL-4. J. Clin. Investig. 2015, 125, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, U.M.; Ujas, T.A.; Kong, X.; Kumar, A.; Plautz, E.J.; Zhang, S.; Xing, C.; Sudduth, T.L.; Wilcock, D.M.; Turchan-Cholewo, J.; et al. Delayed Diapedesis of CD8 T Cells Contributes to Long-Term Pathology after Ischemic Stroke in Male Mice. Brain Behav. Immun. 2021, 95, 502–513. [Google Scholar] [CrossRef]
- Polazzi, E.; Monti, B. Microglia and Neuroprotection: From in Vitro Studies to Therapeutic Applications. Prog. Neurobiol. 2010, 92, 293–315. [Google Scholar] [CrossRef]
- Naqvi, I.; Hitomi, E.; Leigh, R. Sustained Opening of the Blood-Brain Barrier with Progressive Accumulation of White Matter Hyperintensities Following Ischemic Stroke. Brain Sci. 2019, 9, 16. [Google Scholar] [CrossRef]
- Bernardo-Castro, S.; Sousa, J.A.; Martins, E.; Donato, H.; Nunes, C.; d’Almeida, O.C.; Castelo-Branco, M.; Abrunhosa, A.; Ferreira, L.; Sargento-Freitas, J. The Evolution of Blood–Brain Barrier Permeability Changes after Stroke and Its Implications on Clinical Outcome: A Systematic Review and Meta-Analysis. Int. J. Stroke 2023, 18, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Arba, F.; Leigh, R.; Inzitari, D.; Warach, S.J.; Luby, M.; Lees, K.R. Blood-Brain Barrier Leakage Increases with Small Vessel Disease in Acute Ischemic Stroke. Neurology 2017, 89, 2143–2150. [Google Scholar] [CrossRef]
- Maynard, F.M., Jr.; Bracken, M.B.; Creasey, G.; Ditunno, J.F., Jr.; Donovan, W.H.; Ducker, T.B.; Garber, S.L.; Marino, R.J.; Stover, S.L.; Tator, C.H.; et al. International Standards for Neurological and Functional Classification of Spinal Cord Injury. Spinal Cord 1997, 35, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Hakimi, K.N.; Massagli, T.L. Anterior Spinal Artery Syndrome in Two Children with Genetic Thrombotic Disorders. J. Spinal Cord. Med. 2005, 28, 69–73. [Google Scholar] [CrossRef]
- Roth, E.J.; Park, T.; Pang, T.; Yarkony, G.M.; Lee, M.Y. Traumatic Cervical Brown-Sequard and Brown-Sequard-plus Syndromes: The Spectrum of Presentations and Outcomes. Paraplegia 1991, 29, 582–589. [Google Scholar] [CrossRef] [PubMed]
- McKinley, W.; Hills, A.; Sima, A. Posterior Cord Syndrome: Demographics and Rehabilitation Outcomes. J. Spinal Cord. Med. 2021, 44, 241. [Google Scholar] [CrossRef] [PubMed]
- Cauda Equina Syndrome—Symptoms, Causes, Diagnosis and Treatments. Available online: https://www.aans.org/en/Patients/Neurosurgical-Conditions-and-Treatments/Cauda-Equina-Syndrome (accessed on 6 September 2023).
- Little, J.W.; Ditunno, J.F.; Stiens, S.A.; Harris, R.M. Incomplete Spinal Cord Injury: Neuronal Mechanisms of Motor Recovery and Hyperreflexia. Arch. Phys. Med. Rehabil. 1999, 80, 587–599. [Google Scholar] [CrossRef]
- Singhal, V.; Aggarwal, R. Chapter 11—Spinal Shock. In Complications in Neuroanesthesia; Academic Press: Cambridge, MA, USA, 2017; pp. 89–94. [Google Scholar] [CrossRef]
- Ditunno, J.F.; Little, J.W.; Tessler, A.; Burns, A.S. Spinal Shock Revisited: A Four-Phase Model. Spinal Cord. 2004, 42, 383–395. [Google Scholar] [CrossRef]
- Smith, P.M.; Jeffery, N.D. Spinal Shock—Comparative Aspects and Clinical Relevance. J. Vet. Intern. Med. 2005, 19, 788–793. [Google Scholar] [CrossRef]
- Shik, M.L.; Orlovsky, G.N. Neurophysiology of Locomotor Automatism. Physiol. Rev. 1976, 56, 465–501. [Google Scholar] [CrossRef]
- Min, K.J.; Jeong, H.K.; Kim, B.; Hwang, D.H.; Shin, H.Y.; Nguyen, A.T.; Kim, J.H.; Jou, I.; Kim, B.G.; Joe, E. hye Spatial and Temporal Correlation in Progressive Degeneration of Neurons and Astrocytes in Contusion-Induced Spinal Cord Injury. J. Neuroinflamm. 2012, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.A.; Yang, M.S.; Jeong, H.K.; Min, K.J.; Kang, S.H.; Jou, I.; Joe, E.H. Resident Microglia Die and Infiltrated Neutrophils and Monocytes Become Major Inflammatory Cells in Lipopolysaccharide-Injected Brain. Glia 2007, 55, 1577–1588. [Google Scholar] [CrossRef]
- Pineau, I.; Sun, L.; Bastien, D.; Lacroix, S. Astrocytes Initiate Inflammation in the Injured Mouse Spinal Cord by Promoting the Entry of Neutrophils and Inflammatory Monocytes in an IL-1 Receptor/MyD88-Dependent Fashion. Brain Behav. Immun. 2010, 24, 540–553. [Google Scholar] [CrossRef] [PubMed]
- Stirling, D.P.; Yong, V.W. Dynamics of the Inflammatory Response after Murine Spinal Cord Injury Revealed by Flow Cytometry. J. Neurosci. Res. 2008, 86, 1944–1958. [Google Scholar] [CrossRef]
- Fleming, J.C.; Norenberg, M.D.; Ramsay, D.A.; Dekaban, G.A.; Marcillo, A.E.; Saenz, A.D.; Pasquale-Styles, M.; Dietrich, W.D.; Weaver, L.C. The Cellular Inflammatory Response in Human Spinal Cords after Injury. Brain 2006, 129, 3249–3269. [Google Scholar] [CrossRef]
- Hawkins, B.T.; Davis, T.P. The Blood-Brain Barrier/Neurovascular Unit in Health and Disease. Pharmacol. Rev. 2005, 57, 173–185. [Google Scholar] [CrossRef]
- TIMPL, R. Structure and Biological Activity of Basement Membrane Proteins. Eur. J. Biochem. 1989, 180, 487–502. [Google Scholar] [CrossRef] [PubMed]
- Scholz, M.; Cinatl, J.; Schädel-Höpfner, M.; Windolf, J. Neutrophils and the Blood-Brain Barrier Dysfunction after Trauma. Med. Res. Rev. 2007, 27, 401–416. [Google Scholar] [CrossRef]
- Lee, S.M.; Rosen, S.; Weinstein, P.; Van Rooijen, N.; Noble-Haeusslein, L.J. Prevention of Both Neutrophil and Monocyte Recruitment Promotes Recovery after Spinal Cord Injury. J. Neurotrauma 2011, 28, 1893–1907. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Integrins: Versatility, Modulation, and Signaling in Cell Adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Yang, L.; Jin, P.; Wang, X.; Zhou, Q.; Lin, X.; Xi, S. Fluoride Activates Microglia, Secretes Inflammatory Factors and Influences Synaptic Neuron Plasticity in the Hippocampus of Rats. Neurotoxicology 2018, 69, 108–120. [Google Scholar] [CrossRef]
- Nishimura, Y.; Onoe, H.; Morichika, Y.; Perfiliev, S.; Tsukada, H.; Isa, T. Time-Dependent Central Compensatory Mechanisms of Finger Dexterity after Spinal Cord Injury. Science 2007, 318, 1150–1155. [Google Scholar] [CrossRef]
- Beck, H.; Yaari, Y. Plasticity of Intrinsic Neuronal Properties in CNS Disorders. Nat. Rev. Neurosci. 2008, 9, 357–369. [Google Scholar] [CrossRef]
- Barbizan, R.; Oliveira, A.L.R. Impact of Acute Inflammation on Spinal Motoneuron Synaptic Plasticity Following Ventral Root Avulsion. J. Neuroinflamm. 2010, 7, 29. [Google Scholar] [CrossRef]
- Chen, T.; Yu, Y.; Tang, L.J.; Kong, L.; Zhang, C.H.; Chu, H.Y.; Yin, L.W.; Ma, H.Y. Neural Stem Cells Over-Expressing Brain-Derived Neurotrophic Factor Promote Neuronal Survival and Cytoskeletal Protein Expression in Traumatic Brain Injury Sites. Neural Regen. Res. 2017, 12, 433. [Google Scholar] [CrossRef]
- Chandhok, G.; Lazarou, M.; Neumann, B. Structure, Function, and Regulation of Mitofusin-2 in Health and Disease. Biol. Rev. Camb. Philos. Soc. 2017, 93, 933–949. [Google Scholar] [CrossRef]
- Duann, P.; Lin, P.H. Mitochondria Damage and Kidney Disease. Adv. Exp. Med. Biol. 2017, 982, 529–551. [Google Scholar] [CrossRef] [PubMed]
- Krauss, S. Mitochondria: Structure and Role in Respiration. In eLS (Encyclopedia of Life Sciences); John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001. [Google Scholar] [CrossRef]
- Marchi, S.; Patergnani, S.; Missiroli, S.; Morciano, G.; Rimessi, A.; Wieckowski, M.R.; Giorgi, C.; Pinton, P. Mitochondrial and Endoplasmic Reticulum Calcium Homeostasis and Cell Death. Cell Calcium 2017, 69, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Abate, M.; Festa, A.; Falco, M.; Lombardi, A.; Luce, A.; Grimaldi, A.; Zappavigna, S.; Sperlongano, P.; Irace, C.; Caraglia, M.; et al. Mitochondria as Playmakers of Apoptosis, Autophagy and Senescence. Semin. Cell Dev. Biol. 2020, 98, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Palomo, A.; Dodd, S.; Anmella, G.; Carvalho, A.F.; Scaini, G.; Quevedo, J.; Pacchiarotti, I.; Vieta, E.; Berk, M. The Role of Mitochondria in Mood Disorders: From Physiology to Pathophysiology and to Treatment. Front. Psychiatry 2021, 12, 546801. [Google Scholar] [CrossRef]
- Van Der Bliek, A.M.; Sedensky, M.M.; Morgan, P.G. Cell Biology of the Mitochondrion. Genetics 2017, 207, 843–871. [Google Scholar] [CrossRef]
- Yang, B.; Chen, Y.; Shi, J. Reactive Oxygen Species (ROS)-Based Nanomedicine. Chem. Rev. 2019, 119, 4881–4985. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Buttgereit, F.; Burmester, G.R.; Brand, M.D. Bioenergetics of Immune Functions: Fundamental and Therapeutic Aspects. Immunol. Today 2000, 21, 194–199. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in Neurodegenerative Disorders: The Roles of Microglia and Astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Felger, J.C.; Lotrich, F.E. Inflammatory Cytokines in Depression: Neurobiological Mechanisms and Therapeutic Implications. Neuroscience 2013, 246, 199–229. [Google Scholar] [CrossRef]
- Blanco, L.P.; Grazioli, S.; Pugin, J. Mitochondrial Damage-Associated Molecular Patterns: From Inflammatory Signaling to Human Diseases. Front. Immunol. 2018, 9, 832. [Google Scholar] [CrossRef]
- Casaril, A.M.; Dantzer, R.; Bas-Orth, C. Neuronal Mitochondrial Dysfunction and Bioenergetic Failure in Inflammation-Associated Depression. Front. Neurosci. 2021, 15, 725547. [Google Scholar] [CrossRef]
- Wang, X.J.; Kong, K.M.; Qi, W.L.; Ye, W.L.; Song, P.S. Interleukin-1 Beta Induction of Neuron Apoptosis Depends on P38 Mitogen-Activated Protein Kinase Activity after Spinal Cord Injury. Acta Pharmacol. Sin. 2005, 26, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Hellenbrand, D.J.; Quinn, C.M.; Piper, Z.J.; Morehouse, C.N.; Fixel, J.A.; Hanna, A.S. Inflammation after Spinal Cord Injury: A Review of the Critical Timeline of Signaling Cues and Cellular Infiltration. J. Neuroinflamm. 2021, 18, 284. [Google Scholar] [CrossRef] [PubMed]
- Hamza, T.; Barnett, J.B.; Li, B. Interleukin 12 a Key Immunoregulatory Cytokine in Infection Applications. Int. J. Mol. Sci. 2010, 11, 789–806. [Google Scholar] [CrossRef] [PubMed]
- Hill, F.; Kim, C.F.; Gorrie, C.A.; Moalem-Taylor, G. Interleukin-17 Deficiency Improves Locomotor Recovery and Tissue Sparing after Spinal Cord Contusion Injury in Mice. Neurosci. Lett. 2011, 487, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Onishi, R.M.; Gaffen, S.L. Interleukin-17 and Its Target Genes: Mechanisms of Interleukin-17 Function in Disease. Immunology 2010, 129, 311–321. [Google Scholar] [CrossRef]
- Ousman, S.S.; David, S. MIP-1alpha, MCP-1, GM-CSF, and TNF-α Control the Immune Cell Response That Mediates Rapid Phagocytosis of Myelin from the Adult Mouse Spinal Cord. J. Neurosci. 2001, 21, 4649–4656. [Google Scholar] [CrossRef]
- Shen, H.; Xu, B.; Yang, C.; Xue, W.; You, Z.; Wu, X.; Ma, D.; Shao, D.; Leong, K.; Dai, J. A DAMP-Scavenging, IL-10-Releasing Hydrogel Promotes Neural Regeneration and Motor Function Recovery after Spinal Cord Injury. Biomaterials 2022, 280, 121279. [Google Scholar] [CrossRef] [PubMed]
- Junttila, I.S. Tuning the Cytokine Responses: An Update on Interleukin (IL)-4 and IL-13 Receptor Complexes. Front. Immunol. 2018, 9, 338745. [Google Scholar] [CrossRef]
- Ferrante, C.J.; Pinhal-Enfield, G.; Elson, G.; Cronstein, B.N.; Hasko, G.; Outram, S.; Leibovich, S.J. The Adenosine-Dependent Angiogenic Switch of Macrophages to an M2-like Phenotype Is Independent of Interleukin-4 Receptor Alpha (IL-4Rα) Signaling. Inflammation 2013, 36, 921–931. [Google Scholar] [CrossRef]
- Lin, S.; Xu, C.; Lin, J.; Hu, H.; Zhang, C.; Mei, X. Regulation of Inflammatory Cytokines for Spinal Cord Injury Recovery. Histol. Histopathol. 2021, 36, 137–142. [Google Scholar] [CrossRef]
- Yang, L.; Blumbergs, P.C.; Jones, N.R.; Manavis, J.; Sarvestani, G.T.; Ghabriel, M.N. Early Expression and Cellular Localization of Proinflammatory Cytokines Interleukin-1beta, Interleukin-6, and Tumor Necrosis Factor-Alpha in Human Traumatic Spinal Cord Injury. Spine 2004, 29, 966–971. [Google Scholar] [CrossRef]
- Garcia, E.; Aguilar-Cevallos, J.; Silva-Garcia, R.; Ibarra, A. Cytokine and Growth Factor Activation In Vivo and In Vitro after Spinal Cord Injury. Mediat. Inflamm. 2016, 2016, 9476020. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, C.; Lee, S.J. Alpha-Synuclein Stimulation of Astrocytes: Potential Role for Neuroinflammation and Neuroprotection. Oxid. Med. Cell. Longev. 2010, 3, 283–287. [Google Scholar] [CrossRef]
- Ransohoff, R.M. The Chemokine System in Neuroinflammation: An Update. J. Infect. Dis. 2002, 186, S152–S156. [Google Scholar] [CrossRef]
- Kiguchi, N.; Kobayashi, Y.; Kishioka, S. Chemokines and Cytokines in Neuroinflammation Leading to Neuropathic Pain. Curr. Opin. Pharmacol. 2012, 12, 55–61. [Google Scholar] [CrossRef]
- Ubogu, E.E.; Callahan, M.K.; Tucky, B.H.; Ransohoff, R.M. Determinants of CCL5-Driven Mononuclear Cell Migration across the Blood–Brain Barrier. Implications for Therapeutically Modulating Neuroinflammation. J. Neuroimmunol. 2006, 179, 132–144. [Google Scholar] [CrossRef]
- Jickling, G.C.; Sharp, F.R. Biomarker Panels in Ischemic Stroke. Stroke 2015, 46, 915. [Google Scholar] [CrossRef]
- Gkantzios, A.; Tsiptsios, D.; Karatzetzou, S.; Kitmeridou, S.; Karapepera, V.; Giannakou, E.; Vlotinou, P.; Aggelousis, N.; Vadikolias, K. Stroke and Emerging Blood Biomarkers: A Clinical Prospective. Neurol. Int. 2022, 14, 784–803. [Google Scholar] [CrossRef]
- Kamtchum-Tatuene, J.; Jickling, G.C. Blood Biomarkers for Stroke Diagnosis and Management. Neuromol. Med. 2019, 21, 344–368. [Google Scholar] [CrossRef] [PubMed]
- Babuin, L.; Jaffe, A.S. Troponin: The Biomarker of Choice for the Detection of Cardiac Injury. CMAJ 2005, 173, 1191–1202. [Google Scholar] [CrossRef]
- Moradi, A.; Srinivasan, S.; Clements, J.; Batra, J. Beyond the Biomarker Role: Prostate-Specific Antigen (PSA) in the Prostate Cancer Microenvironment. Cancer Metastasis Rev. 2019, 38, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Levinson, T.; Wasserman, A. C-Reactive Protein Velocity (CRPv) as a New Biomarker for the Early Detection of Acute Infection/Inflammation. Int. J. Mol. Sci. 2022, 23, 8100. [Google Scholar] [CrossRef] [PubMed]
- Najmi, E.; Bahbah, E.I.; Negida, A.; Afifi, A.M.; Baratloo, A. Diagnostic Value of Serum Neuron-Specific Enolase Level in Patients With Acute Ischemic Stroke; A Systematic Review and Meta-Analysis. Int. Clin. Neurosci. J. 2019, 6, 36–41. [Google Scholar] [CrossRef]
- Dolati, S.; Soleymani, J.; Kazem Shakouri, S.; Mobed, A. The Trends in Nanomaterial-Based Biosensors for Detecting Critical Biomarkers in Stroke. Clin. Chim. Acta 2021, 514, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Lasek-Bal, A.; Jedrzejowska-Szypulka, H.; Student, S.; Warsz-Wianecka, A.; Zareba, K.; Puz, P.; Bal, W.; Pawletko, K.; Lewin-Kowalik, J. The Importance of Selected Markers of Inflammation and Blood-Brain Barrier Damage for Short-Term Ischemic Stroke Prognosis. J. Physiol. Pharmacol. 2019, 70, 209–217. [Google Scholar] [CrossRef]
- Allard, L.; Burkhard, P.R.; Lescuyer, P.; Burgess, J.A.; Walter, N.; Hocnstrasser, D.F.; Sanchez, J.C. PARK7 and Nucleoside Diphosphate Kinase A as Plasma Markers for the Early Diagnosis of Stroke. Clin. Chem. 2005, 51, 2043–2051. [Google Scholar] [CrossRef]
- Montaner, J.; Mendioroz, M.; Ribó, M.; Delgado, P.; Quintana, M.; Penalba, A.; Chacón, P.; Molina, C.; Fernández-Cadenas, I.; Rosell, A.; et al. A Panel of Biomarkers Including Caspase-3 and D-Dimer May Differentiate Acute Stroke from Stroke-Mimicking Conditions in the Emergency Department. J. Intern. Med. 2011, 270, 166–174. [Google Scholar] [CrossRef]
- Falcione, S.; Kamtchum-Tatuene, J.; Sykes, G.; Jickling, G.C. RNA Expression Studies in Stroke: What Can They Tell Us about Stroke Mechanism? Curr. Opin. Neurol. 2020, 33, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Barr, T.L.; Conley, Y.; Ding, J.; Dillman, A.; Warach, S.; Singleton, A.; Matarin, M. Genomic Biomarkers and Cellular Pathways of Ischemic Stroke by RNA Gene Expression Profiling. Neurology 2010, 75, 1009. [Google Scholar] [CrossRef]
- García-Berrocoso, T.; Palà, E.; Consegal, M.; Piccardi, B.; Negro, A.; Gill, N.; Penalba, A.; Huerga Encabo, H.; Fernández-Cadenas, I.; Meisel, A.; et al. Cardioembolic Ischemic Stroke Gene Expression Fingerprint in Blood: A Systematic Review and Verification Analysis. Transl. Stroke Res. 2020, 11, 326–336. [Google Scholar] [CrossRef]
- Jickling, G.C.; Stamova, B.; Ander, B.P.; Zhan, X.; Tian, Y.; Liu, D.; Xu, H.; Johnston, S.C.; Verro, P.; Sharp, F.R. Profiles of Lacunar and Nonlacunar Stroke. Ann. Neurol. 2011, 70, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Dykstra-Aiello, C.; Jickling, G.C.; Ander, B.P.; Shroff, N.; Zhan, X.; Liu, D.; Hull, H.; Orantia, M.; Stamova, B.S.; Sharp, F.R. Altered Expression of Long Noncoding RNAs in Blood Following Ischemic Stroke and Proximity to Putative Stroke Risk Loci. Stroke 2016, 47, 2896. [Google Scholar] [CrossRef]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Böck, J.; Northoff, B.H.; Klein, M.; et al. RNA-Seq Identifies Circulating MiR-125a-5p, MiR-125b-5p, and MiR-143-3p as Potential Biomarkers for Acute Ischemic Stroke. Circ. Res. 2017, 121, 970–980. [Google Scholar] [CrossRef] [PubMed]
- Herpich, F.; Rincon, F. Management of Acute Ischemic Stroke. Crit. Care Med. 2020, 48, 1654–1663. [Google Scholar] [CrossRef]
- Pop, N.; Tit, D.; Diaconu, C.; Munteanu, M.; Babes, E.; Stoicescu, M.; Popescu, M.; Bungau, S. The Alberta Stroke Program Early CT Score (ASPECTS): A Predictor of Mortality in Acute Ischemic Stroke. Exp. Ther. Med. 2021, 22, 1371. [Google Scholar] [CrossRef] [PubMed]
- Kijima, K.; Kubota, K.; Hara, M.; Kobayakawa, K.; Yokota, K.; Saito, T.; Yoshizaki, S.; Maeda, T.; Konno, D.; Matsumoto, Y.; et al. The Acute Phase Serum Zinc Concentration Is a Reliable Biomarker for Predicting the Functional Outcome after Spinal Cord Injury. EBioMedicine 2019, 41, 659–669. [Google Scholar] [CrossRef]
- Du, W.; Li, H.; Sun, J.; Xia, Y.; Zhu, R.; Zhang, X.; Tian, R. The Prognostic Value of Serum Neuron Specific Enolase (NSE) and S100B Level in Patients of Acute Spinal Cord Injury. Med. Sci. Monit. 2018, 24, 4510. [Google Scholar] [CrossRef] [PubMed]
- Khetani, S.; Aburashed, R.; Singh, A.; Sen, A.; Sanati-Nezhad, A. Immunosensing of S100β Biomarker for Diagnosis of Spinal Cord Injuries (SCI). Sens. Actuators B Chem. 2017, 247, 163–169. [Google Scholar] [CrossRef]
- Lubieniecka, J.M.; Streijger, F.; Lee, J.H.T.; Stoynov, N.; Liu, J.; Mottus, R.; Pfeifer, T.; Kwon, B.K.; Coorssen, J.R.; Foster, L.J.; et al. Biomarkers for Severity of Spinal Cord Injury in the Cerebrospinal Fluid of Rats. PLoS ONE 2011, 6, e19247. [Google Scholar] [CrossRef]
- Ding, H.; Yu, J.; Chang, W.; Liu, F.; He, Z. Searching for Differentially Expressed Proteins in Spinal Cord Injury Based on the Proteomics Analysis. Life Sci. 2020, 242, 117235. [Google Scholar] [CrossRef]
- Tong, B.; Jutzeler, C.R.; Cragg, J.J.; Grassner, L.; Schwab, J.M.; Casha, S.; Geisler, F.; Kramer, J.L.K. Serum Albumin Predicts Long-Term Neurological Outcomes After Acute Spinal Cord Injury. Neurorehabil. Neural Repair. 2018, 32, 7–17. [Google Scholar] [CrossRef]
- Albayar, A.A.; Roche, A.; Swiatkowski, P.; Antar, S.; Ouda, N.; Emara, E.; Smith, D.H.; Ozturk, A.K.; Awad, B.I. Biomarkers in Spinal Cord Injury: Prognostic Insights and Future Potentials. Front. Neurol. 2019, 10, 425947. [Google Scholar] [CrossRef]
- Lieu, A.; Tenorio, G.; Kerr, B.J. Protein Kinase C Gamma (PKCγ) as a Novel Marker to Assess the Functional Status of the Corticospinal Tract in Experimental Autoimmune Encephalomyelitis (EAE). J. Neuroimmunol. 2013, 256, 43–48. [Google Scholar] [CrossRef]
- Zhang, B.; Li, Z.; Zhang, R.; Hu, Y.; Jiang, Y.; Cao, T.; Wang, J.; Gong, L.; Ji, L.; Mu, H.; et al. PKCγ Promotes Axonal Remodeling in the Cortico-Spinal Tract via GSK3β/β-Catenin Signaling after Traumatic Brain Injury. Sci. Rep. 2019, 9, 17078. [Google Scholar] [CrossRef] [PubMed]
- Kamencic, H.; Griebel, R.W.; Lyon, A.W.; Paterson, P.G.; Juurlink, B.H.J. Promoting Glutathione Synthesis after Spinal Cord Trauma Decreases Secondary Damage and Promotes Retention of Function. FASEB J. 2001, 15, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.N.; Glaser, E.P.; Mott, C.A.; Bailey, W.M.; Sullivan, P.G.; Patel, S.P.; Gensel, J.C. Advanced Age and Neurotrauma Diminish Glutathione and Impair Antioxidant Defense after Spinal Cord Injury. J. Neurotrauma 2022, 39, 1075–1089. [Google Scholar] [CrossRef]
- Lucas, J.H.; Wheeler, D.G.; Emery, D.G.; Mallery, S.R. The Endogenous Antioxidant Glutathione as a Factor in the Survival of Physically Injured Mammalian Spinal Cord Neurons. J. Neuropathol. Exp. Neurol. 1998, 57, 937–954. [Google Scholar] [CrossRef] [PubMed]
- Stukas, S.; Cooper, J.; Gill, J.; Fallah, N.; Skinnider, M.A.; Belanger, L.; Ritchie, L.; Tsang, A.; Dong, K.; Streijger, F.; et al. Association of CSF and Serum Neurofilament Light and Glial Fibrillary Acidic Protein, Injury Severity, and Outcome in Spinal Cord Injury. Neurology 2023, 100, E1221–E1233. [Google Scholar] [CrossRef]
- Xu, J.E.X.; Liu, H.; Li, F.; Cao, Y.; Tian, J.; Yan, J. Tumor Necrosis Factor-Alpha Is a Potential Diagnostic Biomarker for Chronic Neuropathic Pain after Spinal Cord Injury. Neurosci. Lett. 2015, 595, 30–34. [Google Scholar] [CrossRef]
- Smith, R.; Chepisheva, M.; Cronin, T.; Seemungal, B.M. Chapter 16—Diagnostic Approaches Techniques in Concussion/Mild Traumatic Brain Injury: Where Are We? In Neurosensory Disorders in Mild Traumatic Brain Injury; Academic Press: Cambridge, MA, USA, 2019; pp. 247–277. [Google Scholar] [CrossRef]
- Planz, O. Development of Cellular Signaling Pathway Inhibitors as New Antivirals against Influenza. Antiviral. Res. 2013, 98, 457–468. [Google Scholar] [CrossRef]
- Saito, N.; Shirai, Y. Protein Kinase C Gamma (PKC Gamma): Function of Neuron Specific Isotype. J. Biochem. 2002, 132, 683–687. [Google Scholar] [CrossRef]
- Kuhle, J.; Gaiottino, J.; Leppert, D.; Petzold, A.; Bestwick, J.P.; Malaspina, A.; Lu, C.H.; Dobson, R.; Disanto, G.; Norgren, N.; et al. Serum Neurofilament Light Chain Is a Biomarker of Human Spinal Cord Injury Severity and Outcome. J. Neurol. Neurosurg. Psychiatry 2015, 86, 273–279. [Google Scholar] [CrossRef]
- Ding, S.Q.; Chen, J.; Wang, S.N.; Duan, F.X.; Chen, Y.Q.; Shi, Y.J.; Hu, J.G.; Lü, H.Z. Identification of Serum Exosomal MicroRNAs in Acute Spinal Cord Injured Rats. Exp. Biol. Med. 2019, 244, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.Q.; Chen, Y.Q.; Chen, J.; Wang, S.N.; Duan, F.X.; Shi, Y.J.; Hu, J.G.; Lü, H.Z. Serum Exosomal MicroRNA Transcriptome Profiling in Subacute Spinal Cord Injured Rats. Genomics 2020, 112, 5086–5100. [Google Scholar] [CrossRef] [PubMed]
- Salah, S.M.M.; Matboli, M.; Nasser, H.E.T.; Abdelnaiem, I.A.; Shafei, A.E.S.; EL-Asmer, M.F. Dysregulation in the Expression of (LncRNA-TSIX, TP53INP2 MRNA, MiRNA-1283) in Spinal Cord Injury. Genomics 2020, 112, 3315–3321. [Google Scholar] [CrossRef]
- Freund, P.; Seif, M.; Weiskopf, N.; Friston, K.; Fehlings, M.G.; Thompson, A.J.; Curt, A. MRI in Traumatic Spinal Cord Injury: From Clinical Assessment to Neuroimaging Biomarkers. Lancet Neurol. 2019, 18, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Seif, M.; Gandini Wheeler-Kingshott, C.A.; Cohen-Adad, J.; Flanders, A.E.; Freund, P. Guidelines for the Conduct of Clinical Trials in Spinal Cord Injury: Neuroimaging Biomarkers. Spinal Cord. 2019, 57, 717–728. [Google Scholar] [CrossRef]
- Lin, T.H.; Sun, P.; Hallman, M.; Hwang, F.C.; Wallendorf, M.; Ray, W.Z.; Spees, W.M.; Song, S.K. Noninvasive Quantification of Axonal Loss in the Presence of Tissue Swelling in Traumatic Spinal Cord Injury Mice. J. Neurotrauma 2019, 36, 2308. [Google Scholar] [CrossRef]
- Hu, R.; Hotta, M.; Maruyama, T.; Fujisawa, M.; Sou, K.; Takeoka, S. Temperature-Responsive Liposome-Linked Immunosorbent Assay for the Rapid Detection of SARS-CoV-2 Using Immunoliposomes. ACS Omega 2022, 7, 26936–26944. [Google Scholar] [CrossRef]
- Paulie, S.; Perlmann, P.; Perlmann, H. Enzyme Linked Immunosorbent Assay. In Cell Biology: A Laboratory Handbook; Academic Press: Cambridge, MA, USA, 2023; pp. 533–538. [Google Scholar] [CrossRef]
- Hornbeck, P.; Winston, S.E.; Fuller, S.A. Enzyme-Linked Immunosorbent Assays (ELISA). Curr. Protoc. Mol. Biol. 1991, 15, 11.2.1–11.2.22. [Google Scholar] [CrossRef]
- Landry, V.; Coburn, P.; Kost, K.; Liu, X.; Li-Jessen, N.Y.K. Diagnostic Accuracy of Liquid Biomarkers in Airway Diseases: Toward Point-of-Care Applications. Front. Med. 2022, 9, 855250. [Google Scholar] [CrossRef]
- Sørensen, S.S.; Nygaard, A.B.; Carlsen, A.L.; Heegaard, N.H.H.; Bak, M.; Christensen, T. Elevation of Brain-Enriched MiRNAs in Cerebrospinal Fluid of Patients with Acute Ischemic Stroke. Biomark. Res. 2017, 5, 24. [Google Scholar] [CrossRef]
- Toor, S.M.; Aldous, E.K.; Parray, A.; Akhtar, N.; Al-Sarraj, Y.; Abdelalim, E.M.; Arredouani, A.; El-Agnaf, O.; Thornalley, P.J.; Pananchikkal, S.V.; et al. Identification of Distinct Circulating MicroRNAs in Acute Ischemic Stroke Patients with Type 2 Diabetes Mellitus. Front. Cardiovasc. Med. 2022, 9, 1024790. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.L. Polymerase Chain Reaction (PCR). In Encyclopedic Reference of Immunotoxicology; Springer: New York, NY, USA, 2005; pp. 519–522. [Google Scholar] [CrossRef]
- Bachman, J. Reverse-Transcription PCR (RT-PCR). Methods Enzymol. 2013, 530, 67–74. [Google Scholar] [CrossRef]
- Heid, C.A.; Stevens, J.; Livak, K.J.; Williams, P.M. Real Time Quantitative PCR. Genome Res. 1996, 6, 986–994. [Google Scholar] [CrossRef] [PubMed]
- Dymond, J.S. Explanatory Chapter: Quantitative PCR. Methods Enzymol. 2013, 529, 279–289. [Google Scholar] [CrossRef]
- Filer, J.E.; Channon, R.B.; Henry, C.S.; Geiss, B.J. A Nuclease Protection ELISA Assay for Colorimetric and Electrochemical Detection of Nucleic Acids. Anal. Methods 2019, 11, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.; Mi, T.; Yu, Z. Detection of Several Harmful Algal Species by Sandwich Hybridization Integrated with a Nuclease Protection Assay. Harmful Algae 2009, 8, 651–657. [Google Scholar] [CrossRef]
- Crapnell, R.D.; Ferrari, A.G.-M.; Dempsey, N.C.; Banks, C.E. Electroanalytical Overview: Screen-Printed Electrochemical Sensing Platforms for the Detection of Vital Cardiac, Cancer and Inflammatory Biomarkers. Sens. Diagn. 2022, 1, 405–428. [Google Scholar] [CrossRef]
- Kim, S.-H.; Weiß, C.; Hoffmann, U.; Borggrefe, M.; Akin, I.; Behnes, M. Advantages and Limitations of Current Biomarker Research: From Experimental Research to Clinical Application. Curr. Pharm. Biotechnol. 2017, 18, 445–455. [Google Scholar] [CrossRef]
- Bittner, T. What Are the Remaining Challenges before Blood-Based Biomarkers for Alzheimer’s Disease Can Be Used in Clinical Practice? J. Prev. Alzheimer’s Dis. 2022, 9, 567–568. [Google Scholar] [CrossRef]
- Muehllehner, G.; Karp, J.S. Positron Emission Tomography. Phys. Med. Biol. 2006, 51, R117–R137. [Google Scholar] [CrossRef]
- Maisey, M.N. Positron Emission Tomography in Clinical Medicine. In Positron Emission Tomography; Springer: New York, NY, USA, 2006; pp. 1–12. [Google Scholar] [CrossRef]
- Shen, Z.; Bao, X.; Wang, R. Clinical PET Imaging of Microglial Activation: Implications for Microglial Therapeutics in Alzheimer’s Disease. Front. Aging Neurosci. 2018, 10, 314. [Google Scholar] [CrossRef] [PubMed]
- Zammit, M.; Tao, Y.; Olsen, M.E.; Metzger, J.; Vermilyea, S.C.; Bjornson, K.; Slesarev, M.; Block, W.F.; Fuchs, K.; Phillips, S.; et al. [18F]FEPPA PET Imaging for Monitoring CD68-Positive Microglia/Macrophage Neuroinflammation in Nonhuman Primates. EJNMMI Res. 2020, 10, 93. [Google Scholar] [CrossRef] [PubMed]
- Cumming, P.; Burgher, B.; Patkar, O.; Breakspear, M.; Vasdev, N.; Thomas, P.; Liu, G.J.; Banati, R. Sifting through the Surfeit of Neuroinflammation Tracers. J. Cereb. Blood Flow. Metab. 2018, 38, 204. [Google Scholar] [CrossRef] [PubMed]
- Van Camp, N.; Lavisse, S.; Roost, P.; Gubinelli, F.; Hillmer, A.; Boutin, H. TSPO Imaging in Animal Models of Brain Diseases. Eur. J. Nucl. Med. Mol. Imaging 2021, 49, 77–109. [Google Scholar] [CrossRef]
- Palleis, C.; Sauerbeck, J.; Beyer, L.; Harris, S.; Schmitt, J.; Morenas-Rodriguez, E.; Finze, A.; Nitschmann, A.; Ruch-Rubinstein, F.; Eckenweber, F.; et al. In Vivo Assessment of Neuroinflammation in 4-Repeat Tauopathies. Mov. Disord. 2021, 36, 883–894. [Google Scholar] [CrossRef]
- Hosoya, T.; Fukumoto, D.; Kakiuchi, T.; Nishiyama, S.; Yamamoto, S.; Ohba, H.; Tsukada, H.; Ueki, T.; Sato, K.; Ouchi, Y. In Vivo TSPO and Cannabinoid Receptor Type 2 Availability Early in Post-Stroke Neuroinflammation in Rats: A Positron Emission Tomography Study. J. Neuroinflamm. 2017, 14, 69. [Google Scholar] [CrossRef]
- Backes, H.; Walberer, M.; Ladwig, A.; Rueger, M.A.; Neumaier, B.; Endepols, H.; Hoehn, M.; Fink, G.R.; Schroeter, M.; Graf, R. Glucose Consumption of Inflammatory Cells Masks Metabolic Deficits in the Brain. Neuroimage 2016, 128, 54–62. [Google Scholar] [CrossRef]
- Maresz, K.; Carrier, E.J.; Ponomarev, E.D.; Hillard, C.J.; Dittel, B.N. Modulation of the Cannabinoid CB2 Receptor in Microglial Cells in Response to Inflammatory Stimuli. J. Neurochem. 2005, 95, 437–445. [Google Scholar] [CrossRef]
- Zhang, M.; Martin, B.R.; Adler, M.W.; Razdan, R.J.; Kong, W.; Ganea, D.; Tuma, R.F. Modulation of Cannabinoid Receptor Activation as a Neuroprotective Strategy for EAE and Stroke. J. Neuroimmune Pharmacol. 2009, 4, 249–259. [Google Scholar] [CrossRef]
- Amenta, P.S.; Jallo, J.I.; Tuma, R.F.; Craig Hooper, D.; Elliott, M.B. Cannabinoid Receptor Type-2 Stimulation, Blockade, and Deletion Alter the Vascular Inflammatory Responses to Traumatic Brain Injury. J. Neuroinflamm. 2014, 11, 191. [Google Scholar] [CrossRef]
- Christian, N.; Lee, J.A.; Bol, A.; De Bast, M.; Jordan, B.; Grégoire, V. The Limitation of PET Imaging for Biological Adaptive-IMRT Assessed in Animal Models. Radiother. Oncol. 2009, 91, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Weis, S. Morphometry and Magnetic Resonance Imaging of the Human Brain in Normal Controls and Down’s Syndrome. Anat. Rec. 1991, 231, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.-P.; Lauterbur, P.C. Principles of Magnetic Resonance Imaging: A Signal Processing Perspective; IEEE Press: Piscataway, NY, USA, 2000. [Google Scholar]
- Sourbron, S.P.; Buckley, D.L. Classic Models for Dynamic Contrast-Enhanced MRI. NMR Biomed. 2013, 26, 1004–1027. [Google Scholar] [CrossRef] [PubMed]
- Cuenod, C.A.; Balvay, D. Perfusion and Vascular Permeability: Basic Concepts and Measurement in DCE-CT and DCE-MRI. Diagn. Interv. Imaging 2013, 94, 1187–1204. [Google Scholar] [CrossRef] [PubMed]
- Desestret, V.; Brisset, J.C.; Moucharrafie, S.; Devillard, E.; Nataf, S.; Honnorat, J.; Nighoghossian, N.; Berthezène, Y.; Wiart, M. Early-Stage Investigations of Ultrasmall Superparamagnetic Iron Oxide-Induced Signal Change after Permanent Middle Cerebral Artery Occlusion in Mice. Stroke 2009, 40, 1834–1841. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Kim, J.H.; Kim, E.; Choi, B.S.; Bae, Y.J.; Bae, H.J. Early Stage of Hyperintense Acute Reperfusion Marker on Contrast-Enhanced FLAIR Images in Patients with Acute Stroke. Am. J. Roentgenol. 2016, 206, 1272–1275. [Google Scholar] [CrossRef]
- Gustafsson, B.; Youens, S.; Louie, A.Y. Development of Contrast Agents Targeted to Macrophage Scavenger Receptors for MRI of Vascular Inflammation. Bioconjug. Chem. 2006, 17, 538–547. [Google Scholar] [CrossRef]
- Taylor, A.; Herrmann, A.; Moss, D.; Sée, V.; Davies, K.; Williams, S.R.; Murray, P. Assessing the Efficacy of Nano- and Micro-Sized Magnetic Particles as Contrast Agents for MRI Cell Tracking. PLoS ONE 2014, 9, e100259. [Google Scholar] [CrossRef]
- De Temmerman, M.L.; Soenen, S.J.; Symens, N.; Lucas, B.; Vandenbroucke, R.E.; Libert, C.; Demeester, J.; De Smedt, S.C.; Himmelreich, U.; Rejman, J. Magnetic Layer-by-Layer Coated Particles for Efficient MRI of Dendritic Cells and Mesenchymal Stem Cells. Nanomedicine 2014, 9, 1363–1376. [Google Scholar] [CrossRef]
- Faraj, A.A.; Shaik, A.S.; Afzal, S.; Sayed, B.A.; Halwani, R. MR Imaging and Targeting of a Specific Alveolar Macrophage Subpopulation in LPS-Induced COPD Animal Model Using Antibody-Conjugated Magnetic Nanoparticles. Int. J. Nanomed. 2014, 9, 1491–1503. [Google Scholar] [CrossRef]
- Chagnot, A.; Barnes, S.R.; Montagne, A. Magnetic Resonance Imaging of Blood–Brain Barrier Permeability in Dementia. Neuroscience 2021, 474, 14–29. [Google Scholar] [CrossRef]
- Ewing, J.R.; Brown, S.L.; Lu, M.; Panda, S.; Ding, G.; Knight, R.A.; Cao, Y.; Jiang, Q.; Nagaraja, T.N.; Churchman, J.L.; et al. Model Selection in Magnetic Resonance Imaging Measurements of Vascular Permeability: Gadomer in a 9L Model of Rat Cerebral Tumor. J. Cereb. Blood Flow. Metab. 2006, 26, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.D.; Paudel, R.; Liu, J.; Ma, C.; Zhang, Z.S.; Zhou, S.K. MRI Contrast Agents: Classification and Application (Review). Int. J. Mol. Med. 2016, 38, 1319–1326. [Google Scholar] [CrossRef] [PubMed]
- Iliff, J.J.; Lee, H.; Yu, M.; Feng, T.; Logan, J.; Nedergaard, M.; Benveniste, H. Brain-Wide Pathway for Waste Clearance Captured by Contrast-Enhanced MRI. J. Clin. Investig. 2013, 123, 1299–1309. [Google Scholar] [CrossRef] [PubMed]
- Plank, J.R.; Morgan, C.A.; Smith, A.K.; Sundram, F.; Hoeh, N.R.; Muthukumaraswamy, S.; Lin, J.C. Detection of Neuroinflammation Induced by Typhoid Vaccine Using Quantitative Magnetization Transfer <scp>MR</Scp>: A Randomized Crossover Study. J. Magn. Reson. Imaging 2023. [Google Scholar] [CrossRef]
- Su, M.-Y.; Jao, J.-C.; Nalcioglu, O. Measurement of Vascular Volume Fraction and Blood-tissue Permeability Constants with a Pharmacokinetic Model: Studies in Rat Muscle Tumors with Dynamic Gd-DTPA Enhanced MRI. Magn. Reson. Med. 1994, 32, 714–724. [Google Scholar] [CrossRef]
- Buzug, T.M. Computed Tomography. In Handbook of Medical Technology; Springer: New York, NY, USA, 2011; pp. 311–342. [Google Scholar] [CrossRef]
- Fleischmann, D.; Boas, F.E. Computed Tomography—Old Ideas and New Technology. Eur. Radiol. 2011, 21, 510–517. [Google Scholar] [CrossRef]
- Brooks, S.L. COMPUTED TOMOGRAPHY. Dent. Clin. N. Am. 1993, 37, 575–590. [Google Scholar] [CrossRef]
- Adebayo, O.D.; Culpan, G. Diagnostic Accuracy of Computed Tomography Perfusion in the Prediction of Haemorrhagic Transformation and Patient Outcome in Acute Ischaemic Stroke: A Systematic Review and Meta-Analysis. Eur. Stroke J. 2020, 5, 4–16. [Google Scholar] [CrossRef]
- Gaberel, T.; Gakuba, C.; Goulay, R.; De Lizarrondo, S.M.; Hanouz, J.L.; Emery, E.; Touze, E.; Vivien, D.; Gauberti, M. Impaired Glymphatic Perfusion after Strokes Revealed by Contrast-Enhanced MRI: A New Target for Fibrinolysis? Stroke 2014, 45, 3092–3096. [Google Scholar] [CrossRef]
- Freeze, W.M.; van der Thiel, M.; de Bresser, J.; Klijn, C.J.M.; van Etten, E.S.; Jansen, J.F.A.; van der Weerd, L.; Jacobs, H.I.L.; Backes, W.H.; van Veluw, S.J. CSF Enhancement on Post-Contrast Fluid-Attenuated Inversion Recovery Images; a Systematic Review. Neuroimage Clin. 2020, 28, 102456. [Google Scholar] [CrossRef]
- Harrison, I.F.; Siow, B.; Akilo, A.B.; Evans, P.G.; Ismail, O.; Ohene, Y.; Nahavandi, P.; Thomas, D.L.; Lythgoe, M.F.; Wells, J.A. Non-Invasive Imaging of CSF-Mediated Brain Clearance Pathways via Assessment of Perivascular Fluid Movement with Diffusion Tensor MRI. eLife 2018, 7, e34028. [Google Scholar] [CrossRef]
- Greer, D.M.; Koroshetz, W.J.; Cullen, S.; Gonzalez, R.G.; Lev, M.H. Magnetic Resonance Imaging Improves Detection of Intracerebral Hemorrhage over Computed Tomography after Intra-Arterial Thrombolysis. Stroke 2004, 35, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Ajmal, S. Contrast-Enhanced Ultrasonography: Review and Applications. Cureus 2021, 13, e18243. [Google Scholar] [CrossRef]
- Prada, F.; Ciocca, R.; Corradino, N.; Gionso, M.; Raspagliesi, L.; Vetrano, I.G.; Doniselli, F.; Del Bene, M.; DiMeco, F. Multiparametric Intraoperative Ultrasound in Oncological Neurosurgery: A Pictorial Essay. Front. Neurosci. 2022, 16, 881661. [Google Scholar] [CrossRef] [PubMed]
- Bruce, M.; Hannah, A.; Hammond, R.; Khaing, Z.Z.; Tremblay-Darveau, C.; Burns, P.N.; Hofstetter, C.P. High-Frequency Nonlinear Doppler Contrast-Enhanced Ultrasound Imaging of Blood Flow. IEEE Trans. Ultrason. Ferroelectr. Freq. Control 2020, 67, 1776–1784. [Google Scholar] [CrossRef]
- Zhou, H.; Meng, L.; Xia, X.; Lin, Z.; Zhou, W.; Pang, N.; Bian, T.; Yuan, T.; Niu, L.; Zheng, H. Transcranial Ultrasound Stimulation Suppresses Neuroinflammation in a Chronic Mouse Model of Parkinson’s Disease. IEEE Trans. Biomed. Eng. 2021, 68, 3375–3387. [Google Scholar] [CrossRef] [PubMed]
- Hosny, A.; Parmar, C.; Quackenbush, J.; Schwartz, L.H.; Aerts, H.J.W.L. Artificial Intelligence in Radiology. Nat. Rev. Cancer 2018, 18, 500. [Google Scholar] [CrossRef]
- Ponce, F.A. Electrostimulation. In Encyclopedia of the Neurological Sciences, 2nd ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 1110–1111. [Google Scholar] [CrossRef]
- Aal, G.A.; Atekwana, E.; Radzikowski, S.; Rossbach, S. Effect of Bacterial Adsorption on Low Frequency Electrical Properties of Clean Quartz Sands and Iron-Oxide Coated Sands. Geophys. Res. Lett. 2009, 36, L04403. [Google Scholar] [CrossRef]
- Jakobs, M.; Fomenko, A.; Lozano, A.M.; Kiening, K.L. Cellular, Molecular, and Clinical Mechanisms of Action of Deep Brain Stimulation—A Systematic Review on Established Indications and Outlook on Future Developments. EMBO Mol. Med. 2019, 11, e9575. [Google Scholar] [CrossRef]
- Yaoita, M.; Aizawa, M.; Ikariyama, Y. Electrically Regulated Cellular Morphological and Cytoskeletal Changes on an Optically Transparent Electrode. Exp. Cell Biol. 1989, 57, 43–51. [Google Scholar] [CrossRef]
- Enayati, S.; Chang, K.; Achour, H.; Cho, K.S.; Xu, F.; Guo, S.Z.; Enayati, K.; Xie, J.; Zhao, E.; Turunen, T.; et al. Electrical Stimulation Induces Retinal Müller Cell Proliferation and Their Progenitor Cell Potential. Cells 2020, 9, 781. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, B.; Gradus Pery, T.; Ophir, E.; Blumenfeld-Katzir, T.; Sheinin, A.; Alon, Y.; Danino, N.; Perlson, E.; Nevo, U. Neuronal Activity in the Sciatic Nerve Is Accompanied by Immediate Cytoskeletal Changes. Front. Mol. Neurosci. 2021, 14, 757264. [Google Scholar] [CrossRef]
- Yu, H.; Enayati, S.; Chang, K.; Cho, K.; Lee, S.W.; Talib, M.; Zihlavnikova, K.; Xie, J.; Achour, H.; Fried, S.I.; et al. Noninvasive Electrical Stimulation Improves Photoreceptor Survival and Retinal Function in Mice with Inherited Photoreceptor Degeneration. Investig. Ophthalmol. Vis. Sci. 2020, 61, 5. [Google Scholar] [CrossRef] [PubMed]
- Binkofski, F.; Loebig, M.; Jauch-Chara, K.; Bergmann, S.; Melchert, U.H.; Scholand-Engler, H.G.; Schweiger, U.; Pellerin, L.; Oltmanns, K.M. Brain Energy Consumption Induced by Electrical Stimulation Promotes Systemic Glucose Uptake. Biol. Psychiatry 2011, 70, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Griffin, L.; Decker, M.J.; Hwang, J.Y.; Wang, B.; Kitchen, K.; Ding, Z.; Ivy, J.L. Functional Electrical Stimulation Cycling Improves Body Composition, Metabolic and Neural Factors in Persons with Spinal Cord Injury. J. Electromyogr. Kinesiol. 2009, 19, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Hutber, C.A.; Hardie, D.G.; Winder, W.W. Electrical Stimulation Inactivates Muscle Acetyl-CoA Carboxylase and Increases AMP-Activated Protein Kinase. Am. J. Physiol.-Endocrinol. Metab. 1997, 272, E262–E266. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Vegas, A.; Campos, C.A.; Contreras-Ferrat, A.; Casas, M.; Buvinic, S.; Jaimovich, E.; Espinosa, A. ROS Production via P2Y1-PKC-NOX2 Is Triggered by Extracellular ATP after Electrical Stimulation of Skeletal Muscle Cells. PLoS ONE 2015, 10, e0129882. [Google Scholar] [CrossRef]
- Bertagna, F.; Lewis, R.; Silva, S.R.P.; McFadden, J.; Jeevaratnam, K. Effects of Electromagnetic Fields on Neuronal Ion Channels: A Systematic Review. Ann. N. Y. Acad. Sci. 2021, 1499, 82–103. [Google Scholar] [CrossRef]
- Clarke, D.; Beros, J.; Bates, K.A.; Harvey, A.R.; Tang, A.D.; Rodger, J. Low Intensity Repetitive Magnetic Stimulation Reduces Expression of Genes Related to Inflammation and Calcium Signalling in Cultured Mouse Cortical Astrocytes. Brain Stimul. 2021, 14, 183–191. [Google Scholar] [CrossRef]
- Yang, H.; Datta-Chaudhuri, T.; George, S.J.; Haider, B.; Wong, J.; Hepler, T.D.; Andersson, U.; Brines, M.; Tracey, K.J.; Chavan, S.S. High-Frequency Electrical Stimulation Attenuates Neuronal Release of Inflammatory Mediators and Ameliorates Neuropathic Pain. Bioelectron. Med. 2022, 8, 16. [Google Scholar] [CrossRef]
- Saddala, M.S.; Lennikov, A.; Mukwaya, A.; Yang, Y.; Hill, M.A.; Lagali, N.; Huang, H. Discovery of Novel L-Type Voltage-Gated Calcium Channel Blockers and Application for the Prevention of Inflammation and Angiogenesis. J. Neuroinflamm. 2020, 17, 132. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.R.; Wolfe, S.A.; Bajo, M.; Abeynaike, S.; Pahng, A.; Borgonetti, V.; D’Ambrosio, S.; Nikzad, R.; Edwards, S.; Paust, S.; et al. IL-10 Normalizes Aberrant Amygdala GABA Transmission and Reverses Anxiety-like Behavior and Dependence-Induced Escalation of Alcohol Intake. Prog. Neurobiol. 2021, 199, 101952. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.T.; Lan, T.H.; Yang, F.Y. Low-Intensity Pulsed Ultrasound Attenuates LPS-Induced Neuroinflammation and Memory Impairment by Modulation of TLR4/NF-ΚB Signaling and CREB/BDNF Expression. Cereb. Cortex 2019, 29, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Rojas, B.; Gallego, B.I.; Ramírez, A.I.; Salazar, J.J.; de Hoz, R.; Valiente-Soriano, F.J.; Avilés-Trigueros, M.; Villegas-Perez, M.P.; Vidal-Sanz, M.; Triviño, A.; et al. Microglia in Mouse Retina Contralateral to Experimental Glaucoma Exhibit Multiple Signs of Activation in All Retinal Layers. J. Neuroinflamm. 2014, 11, 133. [Google Scholar] [CrossRef]
- Lucin, K.M.; Wyss-Coray, T. Immune Activation in Brain Aging and Neurodegeneration: Too Much or Too Little? Neuron 2009, 64, 110–122. [Google Scholar] [CrossRef]
- Schatz, A.; Pach, J.; Gosheva, M.; Naycheva, L.; Willmann, G.; Wilhelm, B.; Peters, T.; Bartz-Schmidt, K.U.; Zrenner, E.; Messias, A.; et al. Transcorneal Electrical Stimulation for Patients With Retinitis Pigmentosa: A Prospective, Randomized, Sham-Controlled Follow-up Study Over 1 Year. Investig. Ophthalmol. Vis. Sci. 2017, 58, 257–269. [Google Scholar] [CrossRef]
- Dorrian, R.M.; Berryman, C.F.; Lauto, A.; Leonard, A.V. Electrical Stimulation for the Treatment of Spinal Cord Injuries: A Review of the Cellular and Molecular Mechanisms That Drive Functional Improvements. Front. Cell. Neurosci. 2023, 17, 1095259. [Google Scholar] [CrossRef]
- Lee, M.; Kiernan, M.C.; Macefield, V.G.; Lee, B.B.; Lin, C.S.Y. Short-Term Peripheral Nerve Stimulation Ameliorates Axonal Dysfunction after Spinal Cord Injury. J. Neurophysiol. 2015, 113, 3209–3218. [Google Scholar] [CrossRef]
- Fang, C.Y.; Lien, A.S.Y.; Tsai, J.L.; Yang, H.C.; Chan, H.L.; Chen, R.S.; Chang, Y.J. The Effect and Dose-Response of Functional Electrical Stimulation Cycling Training on Spasticity in Individuals With Spinal Cord Injury: A Systematic Review With Meta-Analysis. Front. Physiol. 2021, 12, 756200. [Google Scholar] [CrossRef]
- Bergles, D.E.; Richardson, W.D. Oligodendrocyte Development and Plasticity. Cold Spring Harb. Perspect. Biol. 2016, 8, a020453. [Google Scholar] [CrossRef] [PubMed]
- Pekna, M.; Pekny, M.; Nilsson, M. Modulation of Neural Plasticity as a Basis for Stroke Rehabilitation. Stroke 2012, 43, 2819–2828. [Google Scholar] [CrossRef]
- Nowak, D.A.; Grefkes, C.; Ameli, M.; Fink, G.R. Interhemispheric Competition after Stroke: Brain Stimulation to Enhance Recovery of Function of the Affected Hand. Neurorehabil. Neural Repair 2009, 23, 641–656. [Google Scholar] [CrossRef]
- Buma, F.; Kwakkel, G.; Ramsey, N. Understanding Upper Limb Recovery after Stroke. Restor. Neurol. Neurosci. 2013, 31, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Mitragotri, S.; Samir Mitragotri, C. Nanoparticles in the Clinic. Bioeng. Transl. Med. 2016, 1, 10–29. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, G.; Jin, S.; Xu, L.; Zhao, C.X. Development of High-Drug-Loading Nanoparticles. Chempluschem 2020, 85, 2143–2157. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Burgess, D.J. In Vitro Dissolution Testing Strategies for Nanoparticulate Drug Delivery Systems: Recent Developments and Challenges. Drug Deliv. Transl. Res. 2013, 3, 409. [Google Scholar] [CrossRef]
- Tarhini, M.; Greige-Gerges, H.; Elaissari, A. Protein-Based Nanoparticles: From Preparation to Encapsulation of Active Molecules. Int. J. Pharm. 2017, 522, 172–197. [Google Scholar] [CrossRef]
- Ahn, T.; Kim, J.H.; Yang, H.M.; Lee, J.W.; Kim, J.D. Formation Pathways of Magnetite Nanoparticles by Coprecipitation Method. J. Phys. Chem. C 2012, 116, 6069–6076. [Google Scholar] [CrossRef]
- Abdelwahed, W.; Degobert, G.; Stainmesse, S.; Fessi, H. Freeze-Drying of Nanoparticles: Formulation, Process and Storage Considerations. Adv. Drug Deliv. Rev. 2006, 58, 1688–1713. [Google Scholar] [CrossRef]
- Iversen, T.G.; Skotland, T.; Sandvig, K. Endocytosis and Intracellular Transport of Nanoparticles: Present Knowledge and Need for Future Studies. Nano Today 2011, 6, 176–185. [Google Scholar] [CrossRef]
- Verdun, C.; Couvreur, P.; Vranckx, H.; Lenaerts, V.; Roland, M. Development of a Nanoparticle Controlled-Release Formulation for Human Use. J. Control. Release 1986, 3, 205–210. [Google Scholar] [CrossRef]
- Kamaly, N.; Yameen, B.; Wu, J.; Farokhzad, O.C. Degradable Controlled-Release Polymers and Polymeric Nanoparticles: Mechanisms of Controlling Drug Release. Chem. Rev. 2016, 116, 2602–2663. [Google Scholar] [CrossRef] [PubMed]
- Teo, P.; Wang, X.; Chen, B.; Zhang, H.; Yang, X.; Huang, Y.; Tang, J. Complex of TNF-α and Modified Fe3O4 Nanoparticles Suppresses Tumor Growth by Magnetic Induction Hyperthermia. Cancer Biother. Radiopharm. 2017, 32, 379–386. [Google Scholar] [CrossRef]
- Afshari-Kaveh, M.; Abbasalipourkabir, R.; Nourian, A.; Ziamajidi, N. The Protective Effects of Vitamins A and E on Titanium Dioxide Nanoparticles (nTiO2)-Induced Oxidative Stress in the Spleen Tissues of Male Wistar Rats. Biol. Trace Elem. Res. 2021, 199, 3677–3687. [Google Scholar] [CrossRef]
- Jiao, Q.; Li, L.; Mu, Q.; Zhang, Q. Immunomodulation of Nanoparticles in Nanomedicine Applications. Biomed. Res. Int. 2014, 2014, 426028. [Google Scholar] [CrossRef]
- Vlasova, I.I.; Kapralov, A.A.; Michael, Z.P.; Burkert, S.C.; Shurin, M.R.; Star, A.; Shvedova, A.A.; Kagan, V.E. Enzymatic Oxidative Biodegradation of Nanoparticles: Mechanisms, Significance and Applications. Toxicol. Appl. Pharmacol. 2016, 299, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Wang, H.; Wu, B.; Zhang, X. Lipid Nanoparticles for the Delivery of Active Natural Medicines. Curr. Pharm. Des. 2017, 23, 6705–6713. [Google Scholar] [CrossRef] [PubMed]
- Basso, J.; Mendes, M.; Cova, T.; Sousa, J.; Pais, A.; Fortuna, A.; Vitorino, R.; Vitorino, C. A Stepwise Framework for the Systematic Development of Lipid Nanoparticles. Biomolecules 2022, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Al-Jamal, W.T.; Kostarelos, K. Liposome-Nanoparticle Hybrids for Multimodal Diagnostic and Therapeutic Applications. Nanomedicine 2007, 2, 85–98. [Google Scholar] [CrossRef]
- Al-Jamal, W.T.; Kostarelos, K. Liposomes: From a Clinically Established Drug Delivery System to a Nanoparticle Platform for Theranostic Nanomedicine. Acc. Chem. Res. 2011, 44, 1094–1104. [Google Scholar] [CrossRef] [PubMed]
- Mehnert, W.; Mäder, K. Solid Lipid Nanoparticles: Production, Characterization and Applications. Adv. Drug Deliv. Rev. 2012, 64, 83–101. [Google Scholar] [CrossRef]
- Jackson, A.W.; Fulton, D.A. Making Polymeric Nanoparticles Stimuli-Responsive with Dynamic Covalent Bonds. Polym. Chem. 2012, 4, 31–45. [Google Scholar] [CrossRef]
- Zhang, W.; Mehta, A.; Tong, Z.; Esser, L.; Voelcker, N.H. Development of Polymeric Nanoparticles for Blood-Brain Barrier Transfer-Strategies and Challenges. Adv. Sci. 2021, 8, 2003937. [Google Scholar] [CrossRef]
- Sun, W.; Mignani, S.; Shen, M.; Shi, X. Dendrimer-Based Magnetic Iron Oxide Nanoparticles: Their Synthesis and Biomedical Applications. Drug Discov. Today 2016, 21, 1873–1885. [Google Scholar] [CrossRef]
- Avgoustakis, K. Pegylated Poly(Lactide) and Poly(Lactide-Co-Glycolide) Nanoparticles: Preparation, Properties and Possible Applications in Drug Delivery. Curr. Drug Deliv. 2004, 1, 321–333. [Google Scholar] [CrossRef]
- Gonçalves, C.; Pereira, P.; Gama, M. Self-Assembled Hydrogel Nanoparticles for Drug Delivery Applications. Materials 2010, 3, 1420. [Google Scholar] [CrossRef]
- Cornell, R.M.; Schwertmann, U. The Iron Oxides. In The Iron Oxides; Wiley: Hoboken, NY, USA, 2003. [Google Scholar] [CrossRef]
- Pankhurst, Q.A.; Connolly, J.; Jones, S.K.; Dobson, J. Applications of Magnetic Nanoparticles in Biomedicine. J. Phys. D Appl. Phys. 2003, 36, R167–R181. [Google Scholar] [CrossRef]
- Hasany, S.; Abdurahman, N.; Sunarti, A.; Jose, R. Magnetic Iron Oxide Nanoparticles: Chemical Synthesis and Applications Review. Curr. Nanosci. 2013, 9, 561–575. [Google Scholar] [CrossRef]
- Cordova, G.; Attwood, S.; Gaikwad, R.; Gu, F.; Leonenko, Z. Magnetic Force Microscopy Characterization of Superparamagnetic Iron Oxide Nanoparticles (SPIONs). Nano Biomed. Eng. 2014, 6, 31–39. [Google Scholar] [CrossRef]
- Yen, S.K.; Padmanabhan, P.; Selvan, S.T. Multifunctional Iron Oxide Nanoparticles for Diagnostics, Therapy and Macromolecule Delivery. Theranostics 2013, 3, 986. [Google Scholar] [CrossRef] [PubMed]
- Shukur, A.; Azzawi, M.; Farooq, A.; Whitehead, D. Chapter 11—Synthesis of Silica Nanoparticles for Biological Applications. In Nanoparticle Therapeutics: Production Technologies, Types of Nanoparticles, and Regulatory Aspects; Academic Press: Cambridge, MA, USA, 2022; pp. 377–412. [Google Scholar] [CrossRef]
- Zhong, C.; He, M.; Lou, K.; Gao, F. Chapter 10—The Application, Neurotoxicity, and Related Mechanism of Silica Nanoparticles. In Neurotoxicity of Nanomaterials and Nanomedicine; Academic Press: Cambridge, MA, USA, 2017; pp. 227–257. [Google Scholar] [CrossRef]
- Mitran, R.A.; Deaconu, M.; Matei, C.; Berger, D. Chapter 11—Mesoporous Silica as Carrier for Drug-Delivery Systems. In Nanocarriers for Drug Delivery: Nanoscience and Nanotechnology in Drug Delivery—Micro and Nano Technologies; Elsevier: Amsterdam, The Netherlands, 2019; pp. 51–374. [Google Scholar] [CrossRef]
- Esim, O.; Kurbanoglu, S.; Savaser, A.; Ozkan, S.A.; Ozkan, Y. Chapter 9—Nanomaterials for Drug Delivery Systems. In New Developments in Nanosensors for Pharmaceutical Analysis; Academic Press: Cambridge, MA, USA, 2019; pp. 273–301. [Google Scholar] [CrossRef]
- Peng, X.; Lin, G.; Zeng, Y.; Lei, Z.; Liu, G. Mesoporous Silica Nanoparticle-Based Imaging Agents for Hepatocellular Carcinoma Detection. Front. Bioeng. Biotechnol. 2021, 9, 749381. [Google Scholar] [CrossRef] [PubMed]
- Jahanshahi, M.; Babaei, Z. Protein Nanoparticle: A Unique System as Drug Delivery Vehicles. Afr. J. Biotechnol. 2008, 7, 4926–4934. [Google Scholar] [CrossRef]
- Smith, A.A.A.; Zuwala, K.; Pilgram, O.; Johansen, K.S.; Tolstrup, M.; Dagnæs-Hansen, F.; Zelikin, A.N. Albumin-Polymer-Drug Conjugates: Long Circulating, High Payload Drug Delivery Vehicles. ACS Macro Lett. 2016, 5, 1089–1094. [Google Scholar] [CrossRef]
- Hawkins, M.J.; Soon-Shiong, P.; Desai, N. Protein Nanoparticles as Drug Carriers in Clinical Medicine. Adv. Drug Deliv. Rev. 2008, 60, 876–885. [Google Scholar] [CrossRef]
- Dacoba, T.G.; Olivera, A.; Torres, D.; Crecente-Campo, J.; Alonso, M.J. Modulating the Immune System through Nanotechnology. Semin. Immunol. 2017, 34, 78–102. [Google Scholar] [CrossRef]
- Cerqueira, S.R.; Ayad, N.G.; Lee, J.K. Neuroinflammation Treatment via Targeted Delivery of Nanoparticles. Front. Cell. Neurosci. 2020, 14, 576037. [Google Scholar] [CrossRef]
- Teleanu, D.M.; Chircov, C.; Grumezescu, A.M.; Teleanu, R.I. Neurotoxicity of Nanomaterials: An Up-to-Date Overview. Nanomaterials 2019, 9, 96. [Google Scholar] [CrossRef]
- Keller, A.; Linko, V. Challenges and Perspectives of DNA Nanostructures in Biomedicine. Angew. Chem. Int. Ed. 2020, 59, 15818–15833. [Google Scholar] [CrossRef]
- Perry, J.C.; Da Cunha, C.; Anselmo-Franci, J.; Andreatini, R.; Miyoshi, E.; Tufik, S.; Vital, M.A.B.F. Behavioural and Neurochemical Effects of Phosphatidylserine in MPTP Lesion of the Substantia Nigra of Rats. Eur. J. Pharmacol. 2004, 484, 225–233. [Google Scholar] [CrossRef]
- He, X.; Zhu, Y.; Ma, B.; Xu, X.; Huang, R.; Cheng, L.; Zhu, R. Bioactive 2D Nanomaterials for Neural Repair and Regeneration. Adv. Drug Deliv. Rev. 2022, 187, 114379. [Google Scholar] [CrossRef] [PubMed]
- Boverhof, D.R.; David, R.M. Nanomaterial Characterization: Considerations and Needs for Hazard Assessment and Safety Evaluation. Anal. Bioanal. Chem. 2010, 396, 953–961. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokines and Chemokines | Detrimental Effects | Beneficial Effects | Secretory Cell | Refs. |
|---|---|---|---|---|
| Interleukin 1β (IL-1β) | Increased secondary brain damage Increased BBB permeability | Tissue recovery Apoptosis inhibition | Microglia Macrophages | [86] |
| Interleukin 1α (IL-1α) | Chronic inflammation Damage of tissue Autoimmune pathologies | Tissue recovery Activation immune system | Microglia | [2,18] |
| Interleukin 1F1 (IL-1F1) | Increased inflammatory response, hypersensitivity, and autoimmune diseases | Regulation of the immune response Tissue recovery. Neuroprotective function. | Neutrophils | [87] |
| Interleukin 1F2 (IL-1F2) | Increased prostaglandins, cyclooxygenase 2, and phospholipase A2 | Regulation of the immune response. Tissue recovery | Dendritic cells, macrophages, endothelial, and T cells | [87] |
| Interleukin 12 (IL-12) | Increased immune response Difficulty axonal regeneration | Activation immune system Elimination death cells | Dendritic cells, macrophages, monocytes, neutrophils, microglia, and T-cells. | [88] |
| Interleukin 17 (IL-17) | Damage of BBB Increased immune response | Antipathogenic response Decontrol immune cells | T helper, dendritic cells, and macrophages | [89,90] |
| Tumor Necrosis Factor α (TNF-α) | Neurotoxicity Increased BBB permeability | Tissue recovery Antipathogenic response | Microglia, neurons, astrocytes, monocytes, and oligodendrocytes | [2,18,91] |
| Interferon γ (IFN-γ) | Neurotoxicity Difficulty neuroplasticity | Antipathogenic response Tissue recovery | γδ T-cells | [2] |
| Interleukin 5 (IL-5) | Allergic response Decreased immune response | Regulation of allergic pathologies Antipathogenic response | Hematopoietic and non-hematopoietic cells, granulocytes, T, and natural helper cells | [88,92] |
| Interleukin 10 (IL-10) | Neurotoxicity Increased inflammatory response | Inhibition TNF-α; IL-1; IL-6 Limitation inflammatory response | T and B cells, monocytes, dendritic, and natural killer cells | [92] |
| Interleukin 4 (IL-4) | Immunosuppression | Inhibition TNF-α; IL-1; IL-6 Limitation inflammatory response | T helper cells, eosinophils, and eosinophils | [93,94] |
| Interleukin 6 (IL-6) | Neurotoxicity Increased inflammatory response | Antipathogenic response Increased axonal regeneration | Astrocytes, microglia, and neurons | [95,96] |
| Interleukin 8 (IL-8) | Chronic inflammation Cardiovascular and pulmonary diseases | Tissue recovery Neutrophills quimiotaxis | Monocytes, endothelial cells, macrophages, and T cells. | [15] |
| C-C Motif Chemokine Ligand 2 (CCL 2) | Chronic inflammation Autoimmune diseases Increased cancer cell migration | Regulation of immune response Angiogenesis Monocyte chemoattraction | Activated T cells, astrocytes, microglia, and monocytes | [2,97] |
| C-C Motif Chemokine Ligand 3 (CCL 3) | Increased production of proinflammatory cytokines | Regulation of inflammatory response | Monocytes, macrophages, and dendritic cells | [98,99,100] |
| C-C Motif Chemokine Ligand 5 (CCL 5) | Chronic inflammation Cardiovascular diseases Neurological disorders | Immune cells quimiotaxis Regulation of immune response Antiviral response | IL-1 and macrophage migration inhibitory factor | [98,101] |
| Diagnosis | Protein/RNA/Parameter | Biomarker |
|---|---|---|
| Stroke | Proteins | CRP, MMP9, cTni, NSE, BNP, GFAP, S100B, Lp-PLA2, NDKA, PARK7, AQP4, LDH, and Hb |
| Proteins | Panel: D-dimer and caspase-3 | |
| Proteins and parameters | Panel: BNP, gial GFAP, MMP9, AQP4, RDW and NLR | |
| Parameters | ASPECTS | |
| Ischemic stroke | mRNAs | ARG1, LY96, MMP9, 100a12 and CCR7 |
| lncRNAs | linc-SLC22A2 and linc-luo-1172 | |
| miRNAs | miR-125a-5p, miR-125b-5p, and miR-143-3p microRNAs | |
| Cardioembolic stroke and atrial fibrillation | mRNAs | CREM, ZAK, PEI1 |
| Differentiate lacunar between non-lacunar stroke | mRNAs | CCL3, CCL4, HLA-DRB3, IGHA1 and IL8 |
| Functional prognosis in SCI | Proteins | Zinc concentration in serum, TNF-α, and PKCγ |
| Evaluate the degree of SCI | Proteins | NSE, S100B, ITIH4, ApoA1, ApoA4, HSPB1, HIST1H1C, HIST1H1E, albumin, MBP, NF-H |
| Acute SCI | Proteins | NF-L |
| miRNAs | miR-130a-3p, miR-152-3p, miR-125b-5p, miR-30b-5p, and miR-124-3p, | |
| SCI | mRNA, lncRNA, and miRNA | TP53INP2 mRNA and lncRNA-TSIX with decreased miRNA-1283 |
| SCI prognosis | Proteins | TNF-α, MBP, and GSH |
| Parameters | Iron deposition across the neuraxis, demyelination, microstructural changes, and brain volume changes |
| Neuroinflammation | |
|---|---|
| Strengths | Opportunities |
| Present in many neurological diseases such as stroke and SCI. Detectable peripheral biomarkers aid in the diagnosis. Neuroimaging techniques can be employed in its diagnosis. Specific therapeutic options. | Development of new diagnostics techniques, such as AbMAs or SPEs. Development of new therapies, such as ES therapy. Development of new treatments, such as biomaterials. Research in this field is growing. |
| Weaknesses | Threats |
| Research in specific fields is required. A complex process involving many cell types and signaling pathways. Limited treatment options. Invasive techniques may be required. Late diagnosticated. | Limited fundings. Studies with humans may provide ethical dilemmas. Studies with animals may not be extrapolated. Regulatory agencies can slow down treatment and therapies development. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larrea, A.; Elexpe, A.; Díez-Martín, E.; Torrecilla, M.; Astigarraga, E.; Barreda-Gómez, G. Neuroinflammation in the Evolution of Motor Function in Stroke and Trauma Patients: Treatment and Potential Biomarkers. Curr. Issues Mol. Biol. 2023, 45, 8552-8585. https://doi.org/10.3390/cimb45110539
Larrea A, Elexpe A, Díez-Martín E, Torrecilla M, Astigarraga E, Barreda-Gómez G. Neuroinflammation in the Evolution of Motor Function in Stroke and Trauma Patients: Treatment and Potential Biomarkers. Current Issues in Molecular Biology. 2023; 45(11):8552-8585. https://doi.org/10.3390/cimb45110539
Chicago/Turabian StyleLarrea, Ane, Ane Elexpe, Eguzkiñe Díez-Martín, María Torrecilla, Egoitz Astigarraga, and Gabriel Barreda-Gómez. 2023. "Neuroinflammation in the Evolution of Motor Function in Stroke and Trauma Patients: Treatment and Potential Biomarkers" Current Issues in Molecular Biology 45, no. 11: 8552-8585. https://doi.org/10.3390/cimb45110539