
Development of a Novel Anti-CD44 Variant 4 Monoclonal Antibody C44Mab-108 for Immunohistochemistry
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Peptides
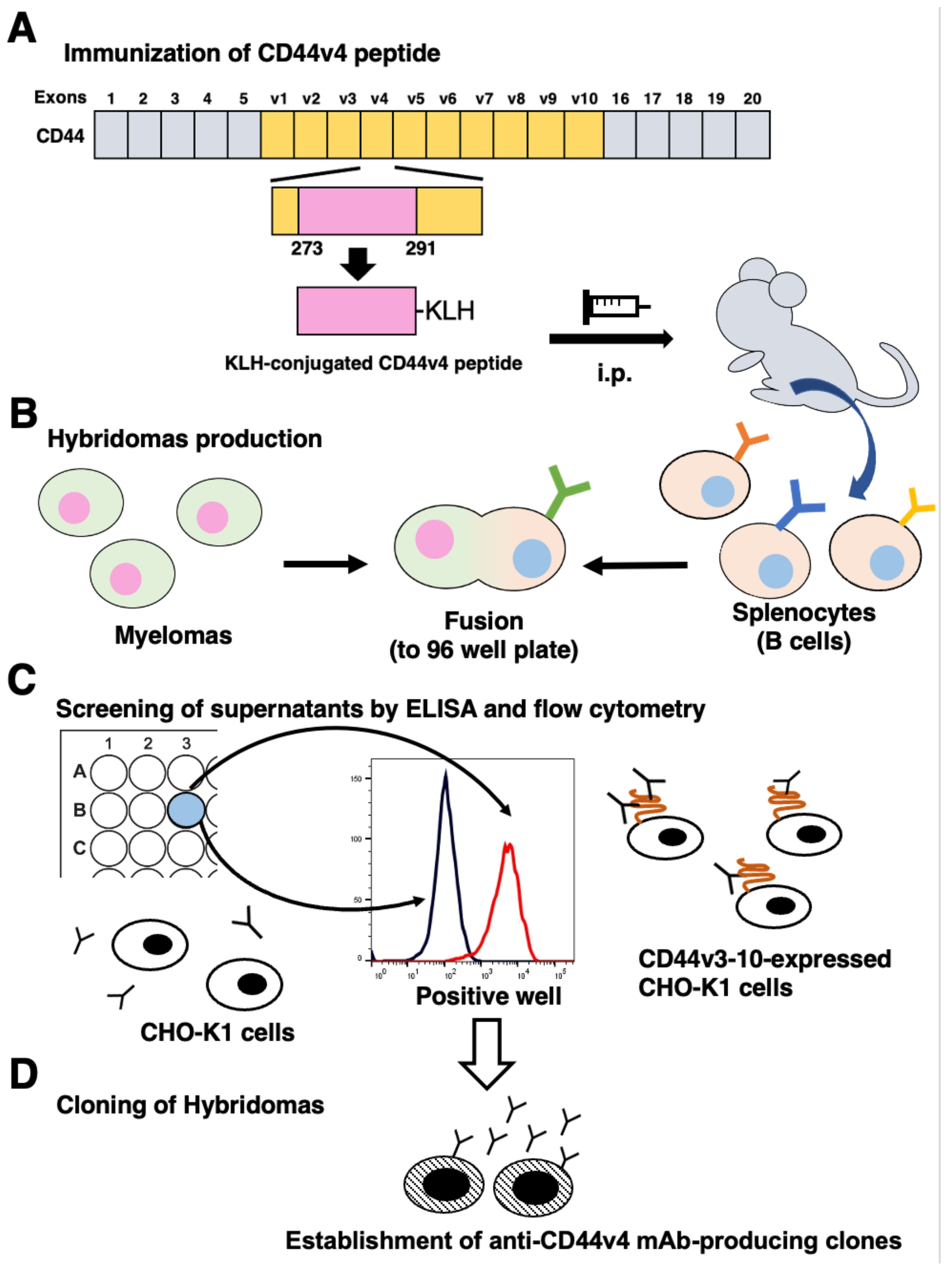
2.3. Hybridoma Production
2.4. ELISA
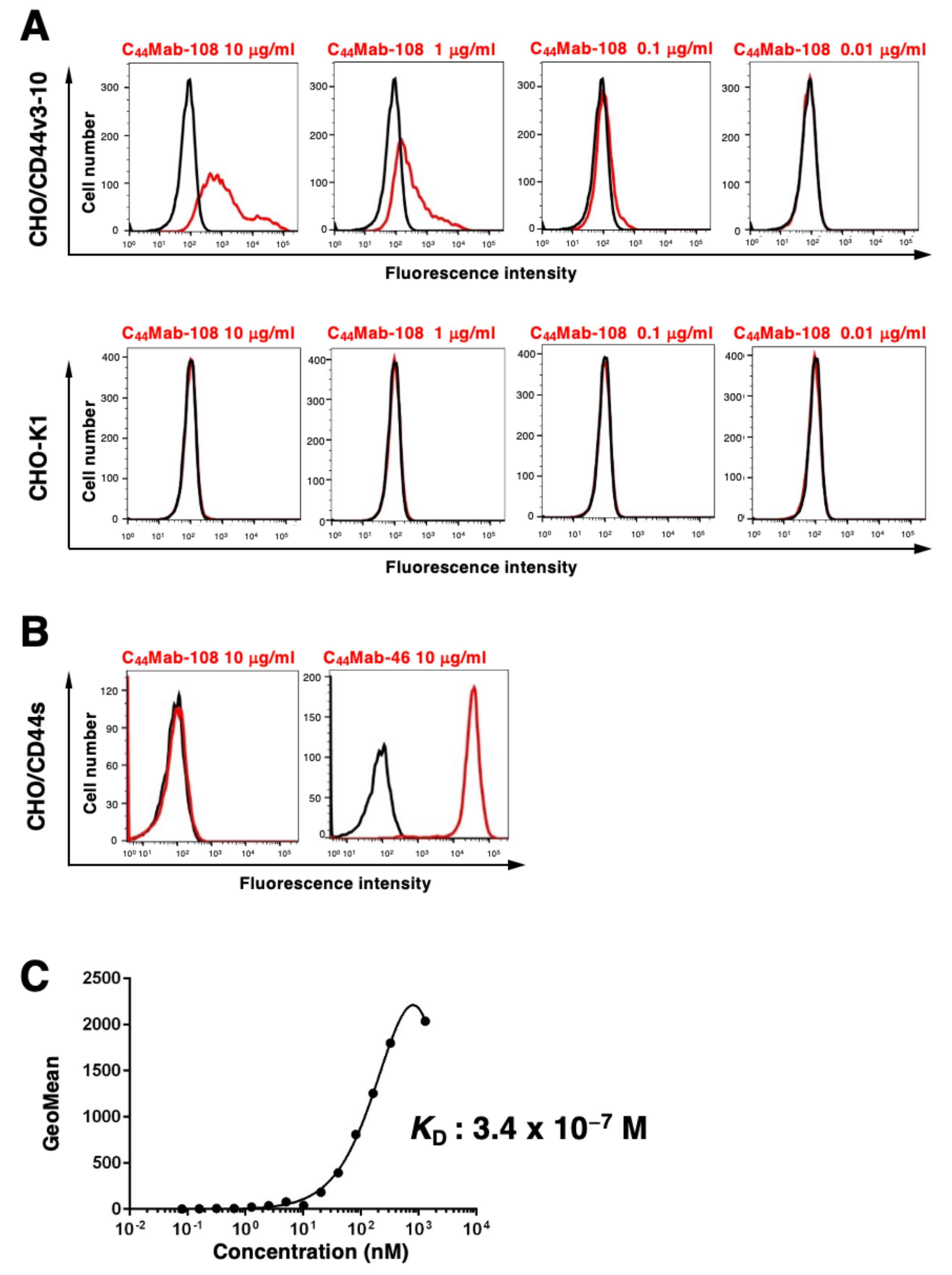
2.5. Flow Cytometry
2.6. Determination of Dissociation Constant (KD) via Flow Cytometry
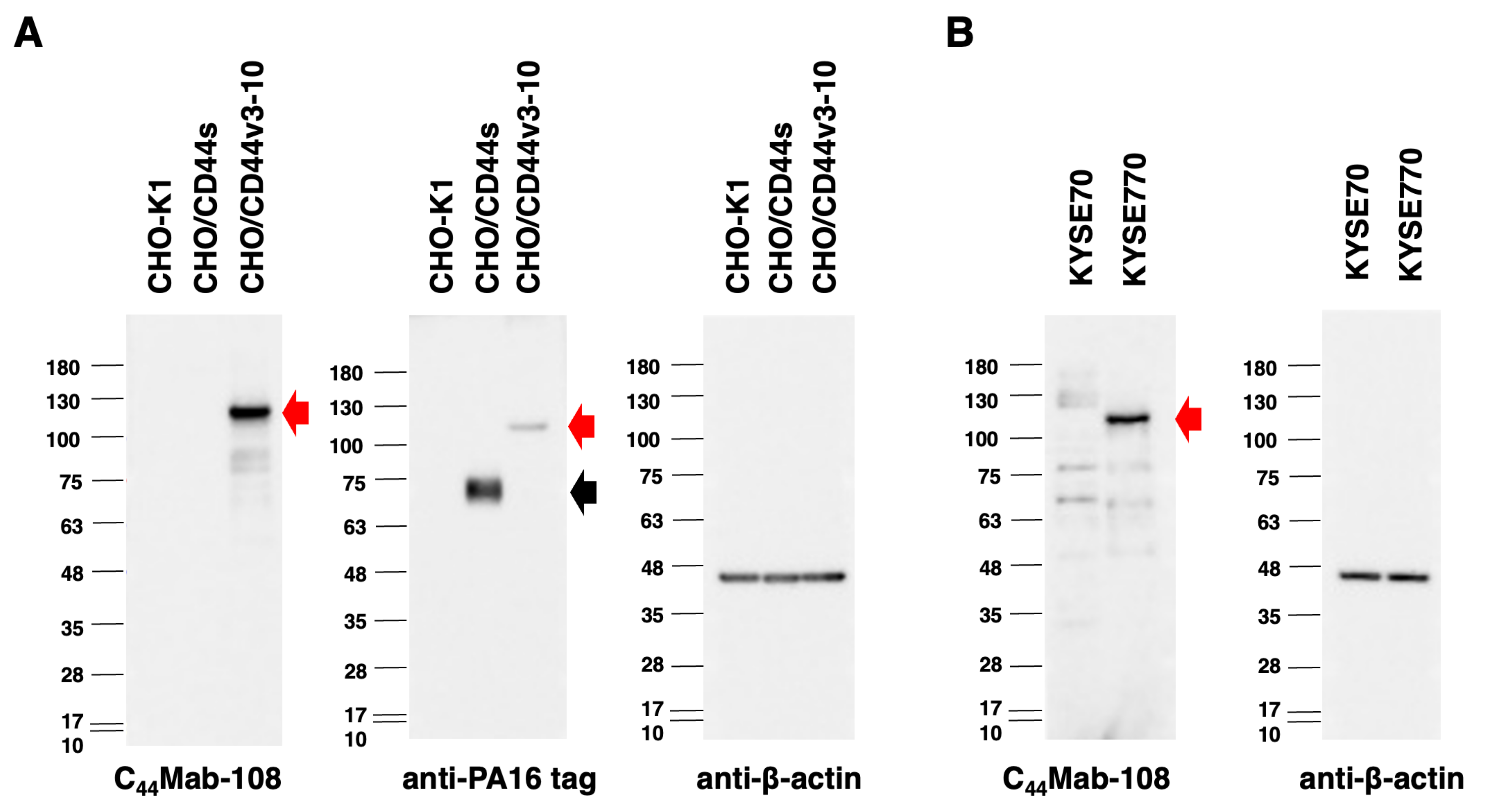
2.7. Western Blot Analysis
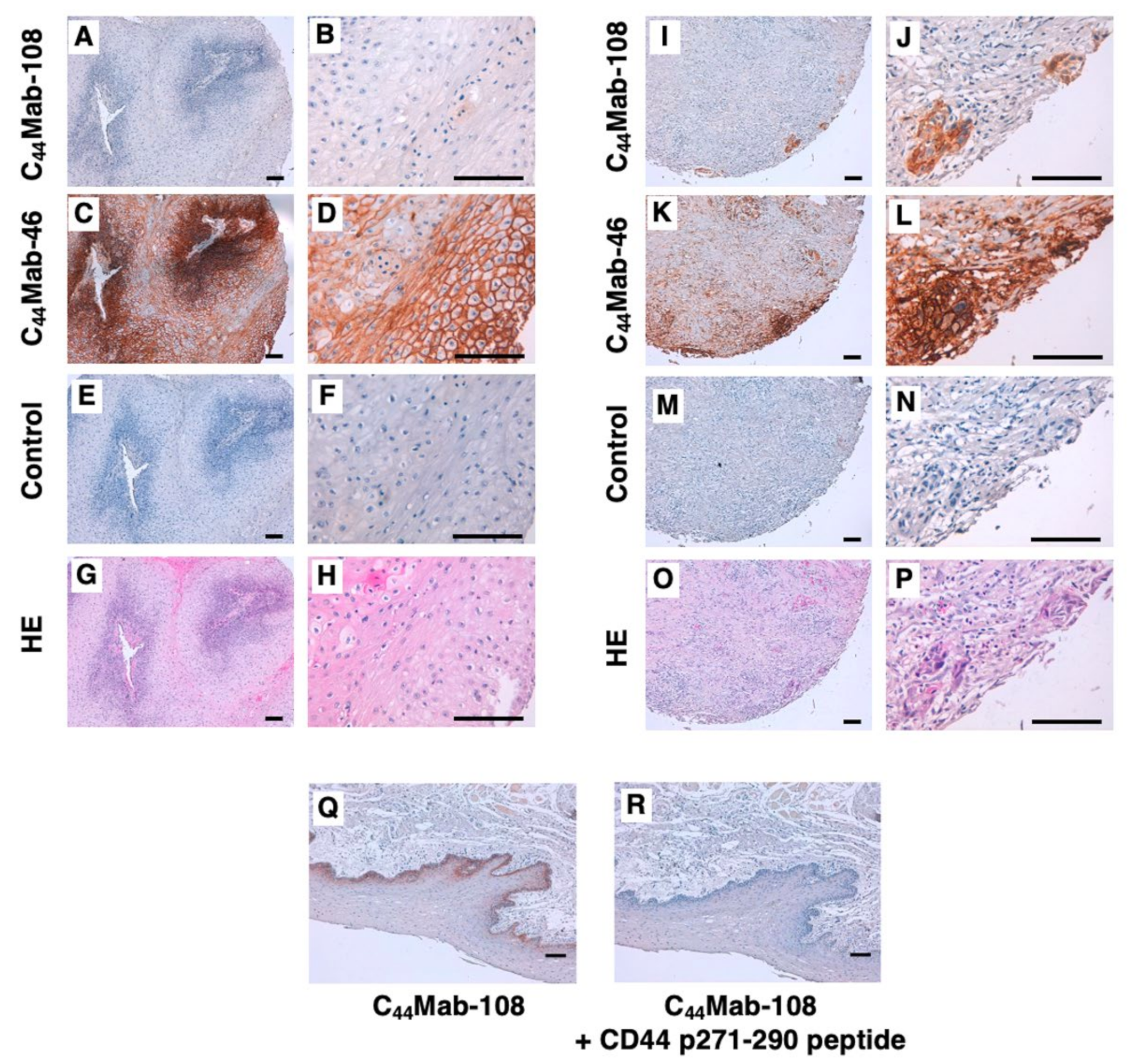
2.8. Immunohistochemical Analysis
3. Results
3.1. Development of Anti-CD44v4 mAbs by Peptide Immunization
3.2. Flow Cytometric Analysis
3.3. Western Blot Analysis
3.4. Immunohistochemical Analysis Using C44Mab-108 against OSCC Tissues
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fox, S.; Fawcett, J.; Jackson, D.G.; Collins, I.; Gatter, K.C.; Harris, A.; Gearing, A.; Simmons, D.L. Normal human tissues, in addition to some tumors, express multiple different CD44 isoforms. Cancer Res. 1994, 54, 4539–4546. [Google Scholar] [PubMed]
- Yan, Y.; Zuo, X.; Wei, D. Concise Review: Emerging Role of CD44 in Cancer Stem Cells: A Promising Biomarker and Therapeutic Target. Stem Cells Transl. Med. 2015, 4, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: Therapeutic implications. J. Hematol. Oncol. 2018, 11, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Günthert, U.; Hofmann, M.; Rudy, W.; Reber, S.; Zöller, M.; Hauβmann, I.; Matzku, S.; Wenzel, A.; Ponta, H.; Herrlich, P. A new variant of glycoprotein CD44 confers metastatic potential to rat carcinoma cells. Cell 1991, 65, 13–24. [Google Scholar] [CrossRef]
- Naor, D.; Wallach-Dayan, S.B.; Zahalka, M.A.; Sionov, R.V. Involvement of CD44, a molecule with a thousand faces, in cancer dissemination. Semin. Cancer Biol. 2008, 18, 260–267. [Google Scholar] [CrossRef]
- Guo, Q.; Yang, C.; Gao, F. The state of CD44 activation in cancer progression and therapeutic targeting. FEBS J. 2021, 289, 7970–7986. [Google Scholar] [CrossRef]
- Slevin, M.; Krupinski, J.; Gaffney, J.; Matou, S.; West, D.; Delisser, H.; Savani, R.C.; Kumar, S. Hyaluronan-mediated angiogenesis in vascular disease: Uncovering RHAMM and CD44 receptor signaling pathways. Matrix Biol. 2007, 26, 58–68. [Google Scholar] [CrossRef]
- Uchino, M.; Kojima, H.; Wada, K.; Imada, M.; Onoda, F.; Satofuka, H.; Utsugi, T.; Murakami, Y. Nuclear β-catenin and CD44 upregulation characterize invasive cell populations in non-aggressive MCF-7 breast cancer cells. BMC Cancer 2010, 10, 414–415. [Google Scholar] [CrossRef]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Mesrati, M.H.; Syafruddin, S.E.; Mohtar, M.A.; Syahir, A. CD44: A Multifunctional Mediator of Cancer Progression. Biomolecules 2021, 11, 1850. [Google Scholar] [CrossRef]
- Morath, I.; Hartmann, T.; Orian-Rousseau, V. CD44: More than a mere stem cell marker. Int. J. Biochem. Cell Biol. 2016, 81, 166–173. [Google Scholar] [CrossRef]
- Bennett, K.L.; Jackson, D.G.; Simon, J.C.; Tanczos, E.; Peach, R.; Modrell, B.; Stamenkovic, I.; Plowman, G.; Aruffo, A. CD44 isoforms containing exon V3 are responsible for the presentation of heparin-binding growth factor. J. Cell Biol. 1995, 128, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Orian-Rousseau, V.; Chen, L.; Sleeman, J.P.; Herrlich, P.; Ponta, H. CD44 is required for two consecutive steps in HGF/c-Met signaling. Genes Dev. 2002, 16, 3074–3086. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, T.; Nagano, O.; Yae, T.; Tamada, M.; Motohara, T.; Oshima, H.; Oshima, M.; Ikeda, T.; Asaba, R.; Yagi, H.; et al. CD44 Variant Regulates Redox Status in Cancer Cells by Stabilizing the xCT Subunit of System xc− and Thereby Promotes Tumor Growth. Cancer Cell 2011, 19, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Itai, S.; Nakamura, T.; Yanaka, M.; Kaneko, M.K.; Kato, Y. Detection of high CD44 expression in oral cancers using the novel monoclonal antibody, C44Mab-5. Biochem. Biophys. Rep. 2018, 14, 64–68. [Google Scholar] [CrossRef]
- Goto, N.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti−CD44 Monoclonal Antibody for Multiple Applications against Esophageal Squamous Cell Carcinomas. Int. J. Mol. Sci. 2022, 23, 5535. [Google Scholar] [CrossRef]
- Takei, J.; Asano, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C44Mab-46) Using Alanine-Scanning Mutagenesis and Surface Plasmon Resonance. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 219–226. [Google Scholar] [CrossRef]
- Asano, T.; Kaneko, M.K.; Takei, J.; Tateyama, N.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C44Mab-46) Using the REMAP Method. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 156–161. [Google Scholar] [CrossRef]
- Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Epitope Mapping System: RIEDL Insertion for Epitope Mapping Method. Monoclon. Antibodies Immunodiagn. Immunother. 2021, 40, 162–167. [Google Scholar] [CrossRef]
- Takei, J.; Kaneko, M.K.; Ohishi, T.; Hosono, H.; Nakamura, T.; Yanaka, M.; Sano, M.; Asano, T.; Sayama, Y.; Kawada, M.; et al. A defucosylated anti-CD44 monoclonal antibody 5-mG2a-f exerts antitumor effects in mouse xenograft models of oral squamous cell carcinoma. Oncol. Rep. 2020, 44, 1949–1960. [Google Scholar] [CrossRef]
- Furusawa, Y.; Yamada, S.; Itai, S.; Nakamura, T.; Takei, J.; Sano, M.; Harada, H.; Fukui, M.; Kaneko, M.K.; Kato, Y. Establishment of a monoclonal antibody PMab-233 for immunohistochemical analysis against Tasmanian devil podoplanin. Biochem. Biophys. Rep. 2019, 18, 100631. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Yamada, S.; Itai, S.; Nakamura, T.; Yanaka, M.; Sano, M.; Harada, H.; Fukui, M.; Kaneko, M.K.; Kato, Y. PMab-219: A monoclonal antibody for the immunohistochemical analysis of horse podoplanin. Biochem. Biophys. Rep. 2019, 18, 100616. [Google Scholar] [CrossRef]
- Furusawa, Y.; Yamada, S.; Itai, S.; Sano, M.; Nakamura, T.; Yanaka, M.; Fukui, M.; Harada, H.; Mizuno, T.; Sakai, Y.; et al. PMab-210: A Monoclonal Antibody Against Pig Podoplanin. Monoclon. Antibodies Immunodiagn. Immunother. 2019, 38, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Yamada, S.; Furusawa, Y.; Itai, S.; Nakamura, T.; Yanaka, M.; Sano, M.; Harada, H.; Fukui, M.; Kaneko, M.K. PMab-213: A Monoclonal Antibody for Immunohistochemical Analysis Against Pig Podoplanin. Monoclon. Antibodies Immunodiagn. Immunother. 2019, 38, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Kaneko, M.K.; Kuno, A.; Uchiyama, N.; Amano, K.; Chiba, Y.; Hasegawa, Y.; Hirabayashi, J.; Narimatsu, H.; Mishima, K.; et al. Inhibition of tumor cell-induced platelet aggregation using a novel anti-podoplanin antibody reacting with its platelet-aggregation-stimulating domain. Biochem. Biophys. Res. Commun. 2006, 349, 1301–1307. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Kunita, A.; Abe, S.; Tsujimoto, Y.; Fukayama, M.; Goto, K.; Sawa, Y.; Nishioka, Y.; Kato, Y. Chimeric anti-podoplanin antibody suppresses tumor metastasis through neutralization and antibody-dependent cellular cytotoxicity. Cancer Sci. 2012, 103, 1913–1919. [Google Scholar] [CrossRef]
- Kato, Y.; Kaneko, M.K.; Kunita, A.; Ito, H.; Kameyama, A.; Ogasawara, S.; Matsuura, N.; Hasegawa, Y.; Suzuki-Inoue, K.; Inoue, O.; et al. Molecular analysis of the pathophysiological binding of the platelet aggregation-inducing factor podoplanin to the C-type lectin-like receptor CLEC-2. Cancer Sci. 2007, 99, 54–61. [Google Scholar] [CrossRef]
- Ogasawara, S.; Kaneko, M.K.; Price, J.E.; Kato, Y. Characterization of Anti-podoplanin Monoclonal Antibodies: Critical Epitopes for Neutralizing the Interaction Between Podoplanin and CLEC-2. Hybridoma 2008, 27, 259–267. [Google Scholar] [CrossRef]
- Kato, Y.; Vaidyanathan, G.; Kaneko, M.K.; Mishima, K.; Srivastava, N.; Chandramohan, V.; Pegram, C.; Keir, S.T.; Kuan, C.-T.; Bigner, D.D.; et al. Evaluation of anti-podoplanin rat monoclonal antibody NZ-1 for targeting malignant gliomas. Nucl. Med. Biol. 2010, 37, 785–794. [Google Scholar] [CrossRef]
- Chandramohan, V.; Bao, X.; Kaneko, M.K.; Kato, Y.; Keir, S.T.; Szafranski, S.E.; Kuan, C.-T.; Pastan, I.H.; Bigner, D.D. Recombinant anti-podoplanin (NZ-1) immunotoxin for the treatment of malignant brain tumors. Int. J. Cancer 2012, 132, 2339–2348. [Google Scholar] [CrossRef]
- Abe, S.; Morita, Y.; Kaneko, M.K.; Hanibuchi, M.; Tsujimoto, Y.; Goto, H.; Kakiuchi, S.; Aono, Y.; Huang, J.; Sato, S.; et al. A Novel Targeting Therapy of Malignant Mesothelioma Using Anti-Podoplanin Antibody. J. Immunol. 2013, 190, 6239–6249. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Kaneko, M.; Neyazaki, M.; Nogi, T.; Kato, Y.; Takagi, J. PA tag: A versatile protein tagging system using a super high affinity antibody against a dodecapeptide derived from human podoplanin. Protein Expr. Purif. 2014, 95, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Matsunaga, Y.; Arimori, T.; Kitago, Y.; Ogasawara, S.; Kaneko, M.K.; Kato, Y.; Takagi, J. Tailored placement of a turn-forming PA tag into the structured domain of a protein to probe its conformational state. J. Cell Sci. 2016, 129, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Tamura, R.; Oi, R.; Akashi, S.; Kaneko, M.K.; Kato, Y.; Nogi, T. Application of the NZ-1 Fab as a crystallization chaperone for PA tag-inserted target proteins. Protein Sci. 2019, 28, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Tamura-Sakaguchi, R.; Aruga, R.; Hirose, M.; Ekimoto, T.; Miyake, T.; Hizukuri, Y.; Oi, R.; Kaneko, M.K.; Kato, Y.; Akiyama, Y.; et al. Moving toward generalizable NZ-1 labeling for 3D structure determination with optimized epitope-tag insertion. Acta Crystallogr. Sect. D Struct. Biol. 2021, 77, 645–662. [Google Scholar] [CrossRef] [PubMed]
- Itai, S.; Ohishi, T.; Kaneko, M.K.; Yamada, S.; Abe, S.; Nakamura, T.; Yanaka, M.; Chang, Y.-W.; Ohba, S.-I.; Nishioka, Y.; et al. Anti-podocalyxin antibody exerts antitumor effects via antibody-dependent cellular cytotoxicity in mouse xenograft models of oral squamous cell carcinoma. Oncotarget 2018, 9, 22480–22497. [Google Scholar] [CrossRef] [PubMed]
- Zöller, M. CD44: Can a cancer-initiating cell profit from an abundantly expressed molecule? Nat. Rev. Cancer 2011, 11, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Orian-Rousseau, V.; Morrison, H.; Matzke, A.; Kastilan, T.; Pace, G.; Herrlich, P.; Ponta, H. Hepatocyte Growth Factor-induced Ras Activation Requires ERM Proteins Linked to Both CD44v6 and F-Actin. Mol. Biol. Cell 2007, 18, 76–83. [Google Scholar] [CrossRef]
- Nagano, O.; Okazaki, S.; Saya, H. Redox regulation in stem-like cancer cells by CD44 variant isoforms. Oncogene 2013, 32, 5191–5198. [Google Scholar] [CrossRef]
- Yu, X.; Orr, C.M.; Chan, H.T.C.; James, S.; Penfold, C.A.; Kim, J.; Inzhelevskaya, T.; Mockridge, C.I.; Cox, K.L.; Essex, J.W.; et al. Reducing affinity as a strategy to boost immunomodulatory antibody agonism. Nature 2023, 614, 539–547. [Google Scholar] [CrossRef]
- Mereiter, S.; Martins, A.M.; Gomes, C.; Balmaña, M.; Macedo, J.A.; Polom, K.; Roviello, F.; Magalhães, A.; Reis, C.A. O-glycan truncation enhances cancer-related functions of CD 44 in gastric cancer. FEBS Lett. 2019, 593, 1675–1689. [Google Scholar] [CrossRef]
- Orian-Rousseau, V.; Ponta, H. Perspectives of CD44 targeting therapies. Arch. Toxicol. 2014, 89, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Riechelmann, H.; Sauter, A.; Golze, W.; Hanft, G.; Schroen, C.; Hoermann, K.; Erhardt, T.; Gronau, S. Phase I trial with the CD44v6-targeting immunoconjugate bivatuzumab mertansine in head and neck squamous cell carcinoma. Oral Oncol. 2008, 44, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Tijink, B.M.; Buter, J.; de Bree, R.; Giaccone, G.; Lang, M.S.; Staab, A.; Leemans, C.R.; van Dongen, G.A. A Phase I Dose Escalation Study with Anti-CD44v6 Bivatuzumab Mertansine in Patients with Incurable Squamous Cell Carcinoma of the Head and Neck or Esophagus. Clin. Cancer Res. 2006, 12, 6064–6072. [Google Scholar] [CrossRef] [PubMed]
- Oordt, C.W.M.-V.D.H.V.; Gomez-Roca, C.; van Herpen, C.; Coveler, A.L.; Mahalingam, D.; Verheul, H.M.W.; van der Graaf, W.T.A.; Christen, R.; Rüttinger, D.; Weigand, S.; et al. First-in-human phase I clinical trial of RG7356, an anti-CD44 humanized antibody, in patients with advanced, CD44-expressing solid tumors. Oncotarget 2016, 7, 80046–80058. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Yamada, S.; Nakamura, T.; Abe, S.; Nishioka, Y.; Kunita, A.; Fukayama, M.; Fujii, Y.; Ogasawara, S.; Kato, Y. Antitumor activity of chLpMab-2, a human–mouse chimeric cancer-specific antihuman podoplanin antibody, via antibody-dependent cellular cytotoxicity. Cancer Med. 2017, 6, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.K.; Nakamura, T.; Kunita, A.; Fukayama, M.; Abe, S.; Nishioka, Y.; Yamada, S.; Yanaka, M.; Saidoh, N.; Yoshida, K.; et al. ChLpMab-23: Cancer-Specific Human–Mouse Chimeric Anti-Podoplanin Antibody Exhibits Antitumor Activity via Antibody-Dependent Cellular Cytotoxicity. Monoclon. Antibodies Immunodiagn. Immunother. 2017, 36, 104–112. [Google Scholar] [CrossRef]
- Kato, Y.; Kaneko, M.K. A Cancer-specific Monoclonal Antibody Recognizes the Aberrantly Glycosylated Podoplanin. Sci. Rep. 2014, 4, srep05924. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Ohishi, T.; Kawada, M.; Kato, Y. A cancer-specific anti-podocalyxin monoclonal antibody (60-mG2a-f) exerts antitumor effects in mouse xenograft models of pancreatic carcinoma. Biochem. Biophys. Rep. 2020, 24, 100826. [Google Scholar] [CrossRef]
- Suzuki, H.; Kaneko, M.K.; Kato, Y. Roles of Podoplanin in Malignant Progression of Tumor. Cells 2022, 11, 575. [Google Scholar] [CrossRef]
- Kato, Y.; Kunita, A.; Fukayama, M.; Abe, S.; Nishioka, Y.; Uchida, H.; Tahara, H.; Yamada, S.; Yanaka, M.; Nakamura, T.; et al. Antiglycopeptide Mouse Monoclonal Antibody LpMab-21 Exerts Antitumor Activity Against Human Podoplanin Through Antibody-Dependent Cellular Cytotoxicity and Complement-Dependent Cytotoxicity. Monoclon. Antibodies Immunodiagn. Immunother. 2017, 36, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, S.; Kaneko, M.K.; Kato, Y.; Fujii, Y.; Honma, R.; Oki, H.; Takagi, M.; Suzuki, H.; Konnai, S.; Nakamura, T.; et al. LpMab-19 Recognizes Sialylated O-Glycan on Thr76 of Human Podoplanin. Monoclon. Antibodies Immunodiagn. Immunother. 2016, 35, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, S.; Kaneko, M.K.; Honma, R.; Oki, H.; Fujii, Y.; Takagi, M.; Suzuki, H.; Kato, Y.; Suzuki, T.; Takakubo, Y.; et al. Establishment of Mouse Monoclonal Antibody LpMab-13 Against Human Podoplanin. Monoclon. Antibodies Immunodiagn. Immunother. 2016, 35, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Ogasawara, S.; Oki, H.; Honma, R.; Takagi, M.; Fujii, Y.; Nakamura, T.; Saidoh, N.; Kanno, H.; Umetsu, M.; et al. Novel Monoclonal Antibody LpMab-17 Developed by CasMab Technology Distinguishes Human Podoplanin from Monkey Podoplanin. Monoclon. Antibodies Immunodiagn. Immunother. 2016, 35, 109–116. [Google Scholar] [CrossRef]
- Kato, Y.; Ogasawara, S.; Oki, H.; Goichberg, P.; Honma, R.; Fujii, Y.; Kaneko, M.K. LpMab-12 Established by CasMab Technology Specifically Detects Sialylated O-Glycan on Thr52 of Platelet Aggregation-Stimulating Domain of Human Podoplanin. PLoS ONE 2016, 11, e0152912. [Google Scholar] [CrossRef]
- Oki, H.; Ogasawara, S.; Kaneko, M.K.; Takagi, M.; Yamauchi, M.; Kato, Y. Characterization of Monoclonal Antibody LpMab-3 Recognizing Sialylated Glycopeptide of Podoplanin. Monoclon. Antibodies Immunodiagn. Immunother. 2015, 34, 44–50. [Google Scholar] [CrossRef]
- Kato, Y.; Kunita, A.; Abe, S.; Ogasawara, S.; Fujii, Y.; Oki, H.; Fukayama, M.; Nishioka, Y.; Kaneko, M.K. The chimeric antibody chLpMab-7 targeting human podoplanin suppresses pulmonary metastasis via ADCC and CDC rather than via its neutralizing activity. Oncotarget 2015, 6, 36003–36018. [Google Scholar] [CrossRef]
- Kaneko, M.K.; Oki, H.; Hozumi, Y.; Liu, X.; Ogasawara, S.; Takagi, M.; Goto, K.; Kato, Y. Monoclonal Antibody LpMab-9 Recognizes O-glycosylated N-Terminus of Human Podoplanin. Monoclon. Antibodies Immunodiagn. Immunother. 2015, 34, 310–317. [Google Scholar] [CrossRef]
- Chalise, L.; Kato, A.; Ohno, M.; Maeda, S.; Yamamichi, A.; Kuramitsu, S.; Shiina, S.; Takahashi, H.; Ozone, S.; Yamaguchi, J.; et al. Efficacy of cancer-specific anti-podoplanin CAR-T cells and oncolytic herpes virus G47Δ combination therapy against glioblastoma. Mol. Ther. Oncolytics 2022, 26, 265–274. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Age | Sex | Organ/Anatomic Site | Pathology Diagnosis | TNM | C44Mab-108 | C44Mab-46 |
|---|---|---|---|---|---|---|---|
| 1 | 78 | M | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | - | + |
| 2 | 40 | M | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | - | ++ |
| 3 | 75 | F | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | ++ | + |
| 4 | 35 | F | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | + | +++ |
| 5 | 61 | M | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | + | +++ |
| 6 | 41 | F | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | + | ++ |
| 7 | 64 | M | Tongue | Squamous cell carcinoma of right tongue | T2N2M0 | - | + |
| 8 | 76 | M | Tongue | Squamous cell carcinoma of tongue | T1N0M0 | - | ++ |
| 9 | 50 | F | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | + | ++ |
| 10 | 44 | M | Tongue | Squamous cell carcinoma of tongue | T2N1M0 | - | +++ |
| 11 | 53 | F | Tongue | Squamous cell carcinoma of tongue | T1N0M0 | - | +++ |
| 12 | 46 | F | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | - | ++ |
| 13 | 50 | M | Tongue | Squamous cell carcinoma of root of tongue | T3N1M0 | + | ++ |
| 14 | 36 | F | Tongue | Squamous cell carcinoma of tongue | T1N0M0 | + | +++ |
| 15 | 63 | F | Tongue | Squamous cell carcinoma of tongue | T1N0M0 | - | ++ |
| 16 | 46 | M | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | + | ++ |
| 17 | 58 | M | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | + | ++ |
| 18 | 64 | M | Lip | Squamous cell carcinoma of lower lip | T1N0M0 | - | +++ |
| 19 | 57 | M | Lip | Squamous cell carcinoma of lower lip | T2N0M0 | - | +++ |
| 20 | 61 | M | Lip | Squamous cell carcinoma of lower lip | T1N0M0 | - | +++ |
| 21 | 60 | M | Gum | Squamous cell carcinoma of gum | T3N0M0 | + | ++ |
| 22 | 60 | M | Gum | Squamous cell carcinoma of gum | T1N0M0 | + | +++ |
| 23 | 69 | M | Gum | Squamous cell carcinoma of upper gum | T3N0M0 | - | +++ |
| 24 | 53 | M | Bucca cavioris | Squamous cell carcinoma of bucca cavioris | T2N0M0 | - | + |
| 25 | 55 | M | Bucca cavioris | Squamous cell carcinoma of bucca cavioris | T1N0M0 | - | ++ |
| 26 | 58 | M | Tongue | Squamous cell carcinoma of base of tongue | T1N0M0 | - | ++ |
| 27 | 63 | M | Oral cavity | Squamous cell carcinoma | T1N0M0 | - | ++ |
| 28 | 48 | F | Tongue | Squamous cell carcinoma of tongue | T1N0M0 | - | ++ |
| 29 | 80 | M | Lip | Squamous cell carcinoma of lower lip | T1N0M0 | - | +++ |
| 30 | 77 | M | Tongue | Squamous cell carcinoma of base of tongue | T2N0M0 | - | +++ |
| 31 | 59 | M | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | - | ++ |
| 32 | 77 | F | Tongue | Squamous cell carcinoma of tongue | T1N0M0 | + | +++ |
| 33 | 56 | M | Tongue | Squamous cell carcinoma of root of tongue | T2N1M0 | - | ++ |
| 34 | 60 | M | Tongue | Squamous cell carcinoma of tongue | T2N1M0 | + | +++ |
| 35 | 62 | M | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | ++ | +++ |
| 36 | 67 | F | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | ++ | +++ |
| 37 | 47 | F | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | + | +++ |
| 38 | 37 | M | Tongue | Squamous cell carcinoma of tongue | T2N1M0 | - | - |
| 39 | 55 | F | Tongue | Squamous cell carcinoma of tongue | T2N0M0 | - | ++ |
| 40 | 56 | F | Bucca cavioris | Squamous cell carcinoma of bucca cavioris | T2N0M0 | + | +++ |
| 41 | 49 | M | Bucca cavioris | Squamous cell carcinoma of bucca cavioris | T1N0M0 | - | - |
| 42 | 45 | M | Bucca cavioris | Squamous cell carcinoma of bucca cavioris | T2N0M0 | - | ++ |
| 43 | 42 | M | Bucca cavioris | Squamous cell carcinoma of bucca cavioris | T3N0M0 | - | +++ |
| 44 | 44 | M | Jaw | Squamous cell carcinoma of right drop jaw | T1N0M0 | + | +++ |
| 45 | 40 | F | Tongue | Squamous cell carcinoma of base of tongue | T2N0M0 | - | +++ |
| 46 | 49 | M | Bucca cavioris | Squamous cell carcinoma of bucca cavioris | T1N0M0 | + | +++ |
| 47 | 56 | F | Tongue | Squamous cell carcinoma of base of tongue | T2N0M0 | - | + |
| 48 | 42 | M | Bucca cavioris | Squamous cell carcinoma of bucca cavioris | T3N0M0 | + | +++ |
| 49 | 87 | F | Face | Squamous cell carcinoma of left face | T2N0M0 | - | + |
| 50 | 50 | M | Gum | Squamous cell carcinoma of gum | T2N0M0 | - | ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, H.; Tanaka, T.; Goto, N.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 4 Monoclonal Antibody C44Mab-108 for Immunohistochemistry. Curr. Issues Mol. Biol. 2023, 45, 1875-1888. https://doi.org/10.3390/cimb45030121
Suzuki H, Tanaka T, Goto N, Kaneko MK, Kato Y. Development of a Novel Anti-CD44 Variant 4 Monoclonal Antibody C44Mab-108 for Immunohistochemistry. Current Issues in Molecular Biology. 2023; 45(3):1875-1888. https://doi.org/10.3390/cimb45030121
Chicago/Turabian StyleSuzuki, Hiroyuki, Tomohiro Tanaka, Nohara Goto, Mika K. Kaneko, and Yukinari Kato. 2023. "Development of a Novel Anti-CD44 Variant 4 Monoclonal Antibody C44Mab-108 for Immunohistochemistry" Current Issues in Molecular Biology 45, no. 3: 1875-1888. https://doi.org/10.3390/cimb45030121
APA StyleSuzuki, H., Tanaka, T., Goto, N., Kaneko, M. K., & Kato, Y. (2023). Development of a Novel Anti-CD44 Variant 4 Monoclonal Antibody C44Mab-108 for Immunohistochemistry. Current Issues in Molecular Biology, 45(3), 1875-1888. https://doi.org/10.3390/cimb45030121