
The Role of Intravesicular Proteins and the Protein Corona of Extracellular Vesicles in the Development of Drug-Induced Polyneuropathy
 , and
, and
Abstract
:1. Proteins and miRNAs of Extracellular Vesicles (EVs) as Biomarkers of Diabetic Polyneuropathy
2. Chemotherapy-Induced Polyneuropathy as a Clinical Challenge
3. Pathogenesis of Polyneuropathy Caused by the Use of Anticancer Drugs
- -
- disruption of microtubule dynamics and axon transport;
- -
- oxidative stress;
- -
- induction of apoptosis by mechanisms associated with microtubule dynamics;
- -
- effect on the endothelium;
- -
- mitochondrial dysfunction;
- -
- proteolytic stress;
- -
- changes in the functioning of the glutamatergic system in the central nervous system (CNS).
3.1. Disruption of Microtubule Dynamics and Axon Transport
3.2. Oxidative Stress
3.3. Apoptosis
3.4. Mitochondrial Damage
3.5. Proteolytic Stress and Autophagy
3.6. Changes in the Functioning of the Glutamatergic System in the Central Nervous System
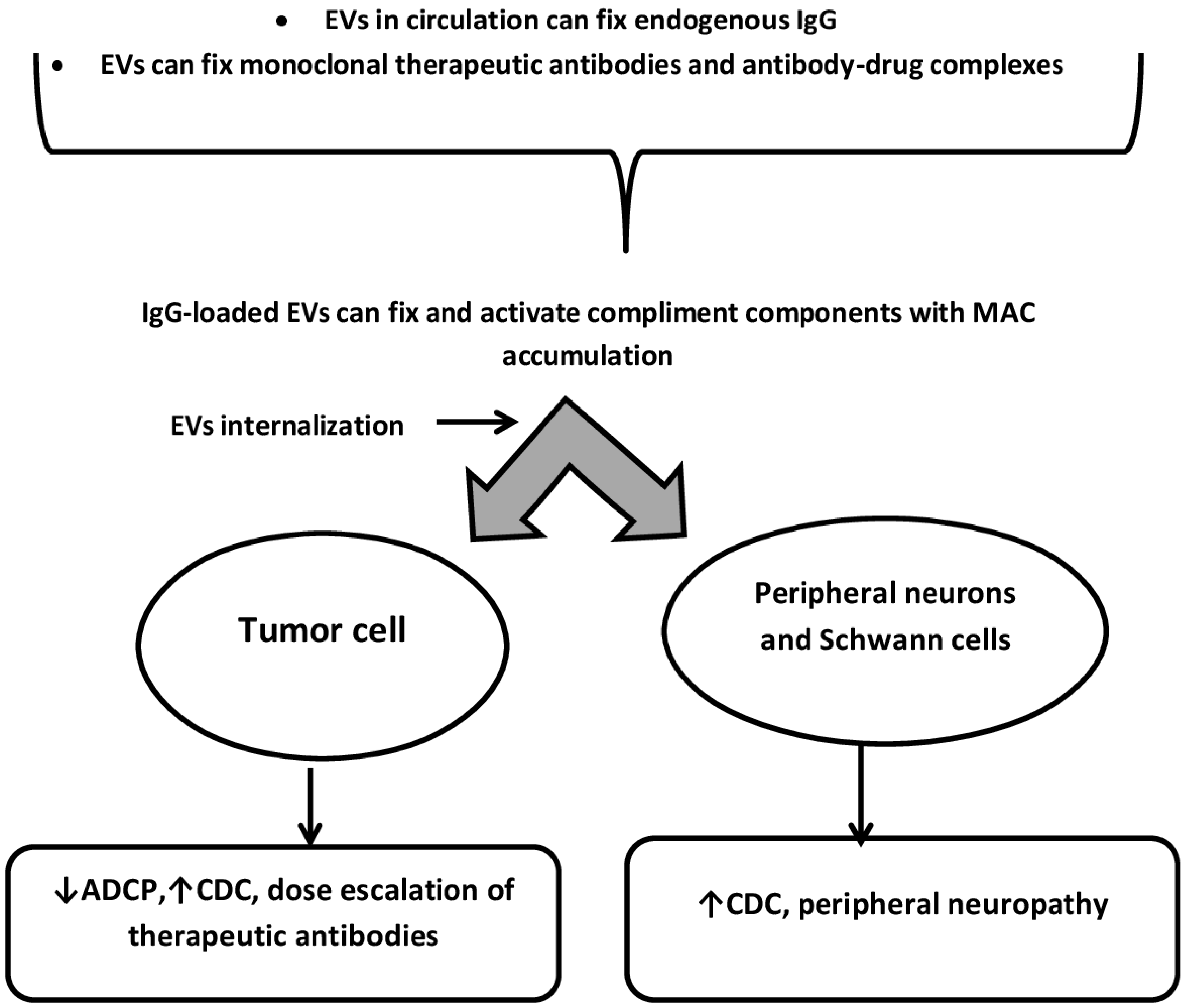
4. Protein-Protein Interactions on the Surface of EVs as Modulators of CIPN and the Effectiveness of Anticancer Therapy with Monoclonal Antibodies and Antibody-Drug Complexes (ADC)
5. Internalization of Extracellular Vesicles by Neurons and Neuroglial Cells: Implications for the Treatment of CIPN
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
References
- Yunusova, N.V.; Dandarova, E.E.; Svarovsky, D.A.; Denisov, N.S.; Kostromitsky, D.N.; Patysheva, M.R.; Cheremisina, O.V.; Spirina, L.V. Production and internalization of extracellular vesicules in normal and under conditions of hyperglycemia and insulin resistance. Biomed. Khim. 2021, 67, 465–474. [Google Scholar] [CrossRef]
- Freeman, D.W.; Noren Hooten, N.; Eitan, E.; Green, J.; Mode, N.A.; Bodogai, M.; Zhang, Y.; Lehrmann, E.; Zonderman, A.B.; Biragyn, A.; et al. Altered Extracellular Vesicle Concentration, Cargo, and Function in Diabetes. Diabetes 2018, 67, 2377–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Marca, V.; Fierabracci, A. Insights into the Diagnostic Potential of Extracellular Vesicles and Their miRNA Signature from Liquid Biopsy as Early Biomarkers of Diabetic Micro/Macrovascular Complications. Int. J. Mol. Sci. 2017, 18, 1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, A.; Benameur, T.; Korashy, H.M.; Zeidan, A.; Agouni, A. Interplay between Endoplasmic Reticulum Stress and Large Extracellular Vesicles (Microparticles) in Endothelial Cell Dysfunction. Biomedicines 2020, 8, 409. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating adipocyte-derived extracellular vesicles are novel markers of metabolic stress. J. Mol. Med. 2016, 94, 1241–1253. [Google Scholar] [CrossRef] [Green Version]
- Kranendonk, M.E.; de Kleijn, D.P.; Kalkhoven, E.; Kanhai, D.A.; Uiterwaal, C.S.; van der Graaf, Y.; Pasterkamp, G.; Visseren, F.L.; SMART Study Group. Extracellular vesicle markers in relation to obesity and metabolic complications in patients with manifest cardiovascular disease. Cardiovasc. Diabetol. 2014, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.D.; Han, L.; Lan, H.T.; Qin, R.R.; Song, M.; Zhang, W.; Zhong, M.; Wang, Z.H. Endothelial microparticle-associated protein disulfide isomerase increases platelet activation in diabetic coronary heart disease. Aging 2021, 13, 18718–18739. [Google Scholar] [CrossRef]
- Osipova, J.; Fischer, D.C.; Dangwal, S.; Volkmann, I.; Widera, C.; Schwarz, K.; Lorenzen, J.M.; Schreiver, C.; Jacoby, U.; Heimhalt, M.; et al. Diabetes-associated microRNAs in pediatric patients with type 1 diabetes mellitus: A cross-sectional cohort study. J. Clin. Endocrinol. Metab. 2014, 99, E1661-5. [Google Scholar] [CrossRef] [Green Version]
- Saenz-Pipaon, G.; Echeverria, S.; Orbe, J.; Roncal, C. Urinary Extracellular Vesicles for Diabetic Kidney Disease Diagnosis. J. Clin. Med. 2021, 10, 2046. [Google Scholar] [CrossRef]
- Kim, H.; Bae, Y.U.; Jeon, J.S.; Noh, H.; Park, H.K.; Byun, D.W.; Han, D.C.; Ryu, S.; Kwon, S.H. The circulating exosomal microRNAs related to albuminuria in patients with diabetic nephropathy. J. Transl. Med. 2019, 17, 236. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, J.; Qian, H.; Wu, Y.; Zhang, Z.; Hu, Z.; Xie, P. Serum Exosomal Circular RNA Expression Profile and Regulative Role in Proliferative Diabetic Retinopathy. Front. Genet. 2021, 12, 719312. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, N.P.; Yan, Y.; Ulrichsen, M.; Venø, M.T.; Poulsen, E.T.; Enghild, J.J.; Kjems, J.; Vægter, C.B. Modulation of Small RNA Signatures in Schwann-Cell-Derived Extracellular Vesicles by the p75 Neurotrophin Receptor and Sortilin. Biomedicines. 2020, 8, 450. [Google Scholar] [CrossRef] [PubMed]
- Kasimu, A.; Apizi, X.; Talifujiang, D.; Ma, X.; Fang, L.; Zhou, X. miR-125a-5p in astrocytes attenuates peripheral neuropathy in type 2 diabetic mice through targeting TRAF6. Endocrinol. Diabetes. Nutr. Engl. Ed. 2021, 69, 43–51. [Google Scholar]
- Cui, X.; Zhu, L.; Zhai, R.; Zhang, B.; Zhang, F. Mesenchymal stem cell-derived exosomes: A promising vector in treatment for diabetes and its microvascular complications. Am. J. Transl. Res. 2021, 13, 3942–3953. [Google Scholar]
- Fujita, Y.; Murakami, T.; Nakamura, A. Recent advances in biomarkers and regenerative medicine for diabetic neuropathy. Int. J. Mol. Sci. 2021, 22, 2301. [Google Scholar] [CrossRef]
- Fan, B.; Chopp, M.; Zhang, Z.; Liu, X.S. Emerging Roles of microRNAs as Biomarkers and Therapeutic Targets for Diabetic Neuropathy. Front. Neurol. 2020, 11, 558758. [Google Scholar] [CrossRef]
- Yan, W.; Wu, Z.; Zhang, Y.; Hong, D.; Dong, X.; Liu, L.; Rao, Y.; Huang, L.; Zhang, X.; Wu, J. The molecular and cellular insight into the toxicology of bortezomib-induced peripheral neuropathy. Biomed. Pharmacother. 2021, 142, 112068. [Google Scholar] [CrossRef]
- Yamamoto, S.; Egashira, N. Pathological Mechanisms of Bortezomib-Induced Peripheral Neuropathy. Int. J. Mol. Sci. 2021, 22, 888. [Google Scholar] [CrossRef]
- Cavaletti, G.; Marmiroli, P. Chemotherapy-induced peripheral neurotoxicity. Nat. Rev. Neurol. 2010, 6, 657–666. [Google Scholar] [CrossRef]
- Bhagra, A.; Rao, R.D. Chemotherapy-induced neuropathy. Curr. Oncol. Rep. 2007, 9, 290–299. [Google Scholar] [CrossRef]
- Cavaletti, G.; Bogliun, G.; Marzorati, L.; Zincone, A.; Piatti, M.; Colombo, N.; Franchi, D.; La Presa, M.T.; Lissoni, A.; Buda, A.; et al. Early predictors of peripheral neurotoxicity in cisplatin and paclitaxel combination chemotherapy. Ann. Oncol. 2004, 15, 1439–1442. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Goldstein, D.; Krishnan, A.; Lin, C.S.; Friedlander, M.L.; Cassidy, J.; Koltzenburg, M.; Kiernan, M. Chemotherapy-induced peripheral neurotoxicity: A critical analysis. CA Cancer J. Clin. 2013, 63, 419–437. [Google Scholar] [CrossRef]
- Kannarkat, G.; Lasher, E.E.; Schiff, D. Neurologic complications of chemotherapy agents. Curr. Opin. Neurol. 2007, 20, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Kus, T.; Aktas, G.; Kalender, M.E.; Sevinc, A.; Kul, S.; Suner, A.; Ulker, E.; Camci, C. Taxane-induced peripheral sensorial neuropathy in cancer patients is associated with duration of diabetes mellitus: A single-center retrospective study. Support Care Cancer 2016, 24, 1175–1179. [Google Scholar] [CrossRef]
- Lee, J.J.; Swain, S.M. Peripheral neuropathy induced by microtubule-stabilizing agents. J. Clin. Oncol. 2006, 24, 1633–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaPointe, N.E.; Morfini, G.; Brady, S.T.; Feinstein, S.C.; Wilson, L.; Jordan, M.A. Effects of eribulin, vincristine, paclitaxel and ixabepilone on fast axonal transport and kinesin-1 driven microtubule gliding: Implications for chemotherapy-induced peripheral neuropathy. Neurotoxicology 2013, 37, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.M.; Pang, H.; Cirrincione, C.; Fleishman, S.; Paskett, E.D.; Ahles, T.; Bressler, L.R.; Fadul, C.E.; Knox, C.; Le-Lindqwister, N.; et al. Alliance for Clinical Trials in Oncology. Effect of duloxetine on pain, function, and quality of life among patients with chemotherapyinduced painful peripheral neuropathy: A randomized clinical trial. JAMA 2013, 309, 1359–1367. [Google Scholar] [CrossRef]
- Argyriou, A.A.; Koltzenburg, M.; Polychronopoulos, P.; Papapetropoulos, S.; Kalofonos, H.P. Peripheral nerve damage associated with administration of taxanes in patients with cancer. Crit. Rev. Oncol. Hematol. 2008, 66, 218–228. [Google Scholar] [CrossRef]
- Argyriou, A.A.; Bruna, J.; Marmiroli, P. Chemotherapyinduced peripheral neurotoxicity (CIPN): An update. Crit. Rev. Oncol. Hematol. 2012, 82, 51–77. [Google Scholar] [CrossRef]
- Goodson, H.V.; Jonasson, E.M. Microtubules and Microtubule-Associated Proteins. Cold Spring Harb. Perspect. Biol. 2018, 10, a022608. [Google Scholar] [CrossRef]
- Hara, M.; Fukagawa, T. Where is the right path heading from the centromere to spindle microtubules? Cell Cycle 2018, 18, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Staff, N.P.; Podratz, J.L.; Grassner, L.; Bader, M.; Paz, J.; Knight, A.M.; Loprinzi, C.L.; Trushina, E.; Windebank, A.J. Bortezomib alters microtubule polymerization and axonal transport in rat dorsal root ganglion neurons. Neurotoxicology 2013, 39, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starobova, H.; Vetter, I. Pathophysiology of Chemotherapy-Induced Peripheral Neuropathy. Front. Mol. Neurosci. 2017, 31, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.Y.; Hur, E.M. A Role of Microtubules in Oligodendrocyte Differentiation. Int. J. Mol. Sci. 2020, 21, 1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meregalli, C.; Marjanovic, I.; Scali, C.; Monza, L.; Spinoni, N.; Galliani, C.; Brivio, R.; Chiorazzi, A.; Ballarini, E.; Rodriguez-Menendez, V.; et al. High-dose intravenous immunoglobulins reduce nerve macrophage infiltration and the severity of bortezomib-induced peripheral neurotoxicity in rats. J. Neuroinflammation 2018, 15, 232. [Google Scholar] [CrossRef] [Green Version]
- Ta, L.E.; Espeset, L.; Podratz, J.; Windebank, A.J. Neurotoxicity of oxaliplatin and cisplatin for dorsal root ganglion neurons correlates with platinum-DNA binding. Neurotoxicology 2006, 27, 992–1002. [Google Scholar] [CrossRef]
- Poitelon, Y.; Kopec, A.M.; Belin, S. Myelin Fat Facts: An Overview of Lipids and Fatty Acid Metabolism. Cells 2020, 9, 812. [Google Scholar] [CrossRef] [Green Version]
- Ermakov, E.A.; Dmitrieva, E.M.; Parshukova, D.A.; Kazantseva, D.V.; Vasilieva, A.R.; Smirnova, L.P. Oxidative Stress-Related Mechanisms in Schizophrenia Pathogenesis and New Treatment Perspectives. Oxid. Med. Cell. Longev. 2021, 2021, 8881770. [Google Scholar] [CrossRef]
- Old, E.A.; Nadkarni, S.; Grist, J.; Gentry, C.; Bevan, S.; Kim, K.W.; Mogg, A.J.; Perretti, M.; Malcangio, M. Monocytes expressing CX3CR1 orchestrate the development of vincristine-induced pain. J. Clin. Investig. 2014, 124, 2023–2036. [Google Scholar] [CrossRef] [Green Version]
- Islam, B.; Lustberg, M.; Staff, N.P.; Kolb, N.; Alberti, P.; Argyriou, A.A. Vinca alkaloids, thalidomide and eribulin-induced peripheral neurotoxicity: From pathogenesis to treatment. J. Peripher. Nerv. Syst. 2019, 24, 63–73. [Google Scholar] [CrossRef]
- Marchi, S.; Patergnani, S.; Missiroli, S.; Morciano, G.; Rimessi, A.; Wieckowski, M.R.; Giorgi, C.; Pinton, P. Mitochondrial and endoplasmic reticulum calcium homeostasis and cell death. Cell Calcium 2018, 69, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Xiao, W.H.; Bennett, G.J. Mitotoxicity and bortezomib-induced chronic painful peripheral neuropathy. Exp. Neurol. 2012, 238, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Meregalli, C.; Canta, A.; Carozzi, V.A.; Chiorazzi, A.; Oggioni, N.; Gilardini, A.; Ceresa, C.; Avezza, F.; Crippa, L.; Marmiroli, P.; et al. Bortezomib-induced painful neuropathy in rats: A behavioral, neurophysiological and pathological study in rats. Eur. J. Pain. 2010, 14, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Janes, K.; Doyle, T.; Bryant, L.; Esposito, E.; Cuzzocrea, S.; Ryerse, J.; Bennett, G.J.; Salvemini, D. Bioenergetic deficits in peripheral nerve sensory axons during chemotherapy-induced neuropathic pain resulting from peroxynitrite-mediated post-translational nitration of mitochondrial superoxide dismutase. Pain 2013, 154, 2432–2440. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Tang, B.; Yu, P.W.; Tang, B.; Hao, Y.X.; Lei, X.; Luo, H.X.; Zeng, D.Z. Autophagy protects against oxaliplatin-induced cell death via ER stress and ROS in Caco-2 cells. PLoS ONE 2012, 7, e51076. [Google Scholar] [CrossRef]
- Meregalli, C.; Chiorazzi, A.; Carozzi, V.A.; Canta, A.; Sala, B.; Colombo, M.; Oggioni, N.; Ceresa, C.; Foudah, D.; La Russa, F.; et al. Evaluation of tubulin polymerization and chronic inhibition of proteasome as citotoxicity mechanisms in bortezomib-induced peripheral neuropathy. Cell Cycle 2014, 13, 612–621. [Google Scholar] [CrossRef] [Green Version]
- Ghelardini, C.; Menicacci, C.; Cerretani, D.; Bianchi, E. Spinal administration of mGluR5 antagonist prevents the onset of bortezomib induced neuropathic pain in rat. Neuropharmacology 2014, 86, 294–300. [Google Scholar] [CrossRef]
- Robinson, C.R.; Dougherty, P.M. Spinal astrocyte gap junction and glutamate transporter expression contributes to a rat model of bortezomib-induced peripheral neuropathy. Neuroscience 2015, 285, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Warrington, A.E.; Rodriguez, M. Remyelination-promoting human IgMs: Developing a therapeutic reagent for demyelinating disease. Curr. Top. Microbiol. Immunol. 2008, 318, 213–239. [Google Scholar]
- Belogurov, A.A., Jr.; Kurkova, I.N.; Friboulet, A.; Thomas, D.; Misikov, V.K.; Zakharova, M.Y.; Suchkov, S.V.; Kotov, S.V.; Alehin, A.I.; Avalle, B.; et al. Recognition and degradation of myelin basic protein peptides by serum autoantibodies: Novel biomarker for multiple sclerosis. J. Immunol. 2008, 180, 1258–1267. [Google Scholar] [CrossRef] [Green Version]
- Doronin, V.B.; Parkhomenko, T.A.; Castellazzi, M.; Padroni, M.; Pastore, M.; Buneva, V.N.; Granieri, E.; Nevinsky, G.A. Comparison of antibodies hydrolyzing myelin basic protein from the cerebrospinal fluid and serum of patients with multiple sclerosis. PLoS ONE 2014, 9, e107807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ermakov, E.A.; Kabirova, E.M.; Buneva, V.N.; Nevinsky, G.A. IgGs-Abzymes from the Sera of Patients with Multiple Sclerosis Recognize and Hydrolyze miRNAs. Int. J. Mol. Sci. 2021, 22, 2812. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, L.P.; Mednova, I.A.; Krotenko, N.M.; Alifirova, V.M.; Ivanova, S.A. IgG-Dependent Dismutation of Superoxide in Patients with Different Types of Multiple Sclerosis and Healthy Subjects. Oxid. Med. Cell. Longev. 2020, 2020, 8171020. [Google Scholar] [CrossRef] [PubMed]
- Tolmacheva, A.S.; Buneva, V.N.; Nevinsky, G.A. Substrate specificity of IgGs with peroxidase and oxidoreductase activities from sera of patients with systemic lupus erythematosus and multiple sclerosis. J. Mol. Recognit. 2019, 32, e2807. [Google Scholar] [CrossRef]
- Parshukova, D.A.; Smirnova, L.P.; Kornetova, E.G.; Semke, A.V.; Buneva, V.N.; Ivanova, S.A. IgG-Dependent Hydrolysis of Myelin Basic Protein of Patients with Different Courses of Schizophrenia. J. Immunol. Res. 2020, 2020, 8986521. [Google Scholar] [CrossRef] [PubMed]
- Toth, E.A.; Turiak, L.; Visnovitz, T.; Cserep, C.; Mazlo, A.; Sodar, B.W.; Försönits, A.I.; Petovari, G.; Sebestyen, A.; Komlosi, Z.; et al. Formation of a protein corona on the surface of extracellular vesicles in blood plasma. J. Extracell. Vesicles 2021, 10, e12140. [Google Scholar] [CrossRef] [PubMed]
- Economopoulos, T.; Papageorgiou, S.; Pappa, V.; Papageorgiou, E.; Valsami, S.; Kalantzis, D.; Xiros, N.; Dervenoulas, J.; Raptis, S. Monoclonal gammopathies in B-cell non-Hodgkin’s lymphomas. Leuk. Res. 2003, 27, 505–508. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, Z.; Li, J.; Gao, R.; Liu, P. Monoclonal gammopathies regardless of subtypes are associated with poor prognosis of diffuse large B-cell lymphoma. Medicine 2018, 97, e11719. [Google Scholar] [CrossRef]
- Seki, M.; Kitano, S.; Suzuki, S. Neurological disorders associated with immune checkpoint inhibitors: An association with autoantibodies. Cancer Immunol. Immunother. 2022, 71, 769–775. [Google Scholar] [CrossRef]
- Elkoshi, Z. Cancer and Autoimmune Diseases: A Tale of Two Immunological Opposites? Front. Immunol. 2022, 13, 821598. [Google Scholar] [CrossRef]
- Khan, S.A.; Pruitt, S.L.; Xuan, L.; Gerber, D.E. Prevalence of Autoimmune Disease among Patients with Lung Cancer: Implications for Immunotherapy Treatment Options. JAMA Oncol. 2016, 2, 1507–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolmacheva, A.S.; Nevinsky, G.A. Essential Protective Role of Catalytically Active Antibodies (Abzymes) with Redox Antioxidant Functions in Animals and Humans. Int. J. Mol. Sci 2022, 23, 3898. [Google Scholar] [CrossRef] [PubMed]
- Wootla, B.; Lacroix-Desmazes, S.; Warrington, A.E.; Bieber, A.J.; Kaveri, S.V.; Rodriguez, M. Autoantibodies with enzymatic properties in human autoimmune diseases. J. Autoimmun. 2011, 37, 144–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barok, M.; Puhka, M.; Yazdi, N.; Joensuu, H. Extracellular vesicles as modifiers of antibody-drug conjugate efficacy. J. Extracell. Vesicles 2021, 10, e12070. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.L.; Liu, J.Y.; Chen, G. Small extracellular vesicle PD-L1 in cancer: The knowns and unknowns. NPJ Precis. Oncol. 2022, 6, 42. [Google Scholar] [CrossRef]
- Oksvold, M.P.; Kullmann, A.; Forfang, L.; Kierulf, B.; Li, M.; Brech, A.; Vlassov, A.V.; Smeland, E.B.; Neurauter, A.; Pedersen, K.W. Expression of B-cell surface antigens in subpopulations of exosomes released from B-cell lymphoma cells. Clin. Ther. 2014, 36, 847–862. [Google Scholar] [CrossRef] [Green Version]
- Drucker, A.; Yoo, B.H.; Khan, I.A.; Choi, D.; Montermini, L.; Liu, X.; Jovanovic, S.; Younis, T.; Rosen, K.V. Trastuzumab-induced upregulation of a protein set in extracellular vesicles emitted by ErbB2-positive breast cancer cells correlates with their trastuzumab sensitivity. Breast Cancer Res. 2020, 22, 105. [Google Scholar] [CrossRef]
- Karasu, E.; Eisenhardt, S.U.; Harant, J.; Huber-Lang, M. Extracellular Vesicles: Packages Sent with Complement. Front. Immunol. 2018, 9, 721. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Fisher, K.P.; Hammer, S.S.; Busik, J.V. Extracellular Vesicle-Induced Classical Complement Activation Leads to Retinal Endothelial Cell Damage via MAC Deposition. Int. J. Mol. Sci. 2020, 21, 1693. [Google Scholar] [CrossRef] [Green Version]
- Nogueras-Ortiz, C.J.; Mahairaki, V.; Delgado-Peraza, F.; Das, D.; Avgerinos, K.; Eren, E.; Hentschel, M.; Goetzl, E.J.; Mattson, M.P.; Kapogiannis, D. Astrocyte- and Neuron-Derived Extracellular Vesicles from Alzheimer’s Disease Patients Effect Complement-Mediated Neurotoxicity. Cells 2020, 9, 1618. [Google Scholar] [CrossRef]
- Zappulli, V.; Friis, K.P.; Fitzpatrick, Z.; Maguire, C.A.; Breakefield, X.O. Extracellular vesicles and intercellular communication within the nervous system. J. Clin. Invest. 2016, 126, 1198–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chine, V.B.; Au, N.P.B.; Ma, C.H.E. Therapeutic benefits of maintaining mitochondrial integrity and calcium homeostasis by forced expression of Hsp27 in chemotherapy-induced peripheral neuropathy. Neurobiol. Dis. 2019, 130, 104492. [Google Scholar] [CrossRef] [PubMed]
- Korngut, L.; Ma, C.H.; Martinez, J.A.; Toth, C.C.; Guo, G.F.; Singh, V.; Woolf, C.J.; Zochodne, D.W. Overexpression of human HSP27 protects sensory neurons from diabetes. Neurobiol. Dis. 2012, 47, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, A.V.; Kim, E.R.; Ovchinnikov, L.P. Proteasome system of protein degradation and processing. Biochemistry 2009, 74, 1411–1442. [Google Scholar] [CrossRef] [PubMed]
- Njomen, E.; Osmulski, P.A.; Jones, C.L.; Gaczynska, M.; Tepe, J.J. Small Molecule Modulation of Proteasome Assembly. Biochemistry 2018, 57, 4214–4224. [Google Scholar] [CrossRef]
- Yunusova, N.; Kolegova, E.; Sereda, E.; Kolomiets, L.; Villert, A.; Patysheva, M.; Rekeda, I.; Grigor’eva, A.; Tarabanovskaya, N.; Kondakova, I.; et al. Plasma Exosomes of Patients with Breast and Ovarian Tumors Contain an Inactive 20S Proteasome. Molecules 2021, 26, 6965. [Google Scholar] [CrossRef] [PubMed]
- Prada, I.; Meldolesi, J. Binding and Fusion of Extracellular Vesicles to the Plasma Membrane of Their Cell Targets. Int. J. Mol. Sci. 2016, 17, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droste, M.; Thakur, B.K.; Eliceiri, B.P. Tumor-Derived Extracellular Vesicles and the Immune System-Lessons From Immune-Competent Mouse-Tumor Models. Front. Immunol. 2020, 11, 606859. [Google Scholar] [CrossRef]
- Kamerkar, S.; Le Bleu, V.S.; Sugimoto, H.; Yang, S.; Ruivo, C.F.; Melo, S.A.; Lee, J.J.; Kalluri, R. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature 2017, 546, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Oh, J.M.; Gangadaran, P.; Kalimuthu, S.; Baek, S.H.; Jeong, S.Y.; Lee, S.W.; Lee, J.; Ahn, B.C. Targeting and Therapy of Glioblastoma in a Mouse Model Using Exosomes Derived from Natural Killer Cells. Front. Immunol. 2018, 9, 824. [Google Scholar] [CrossRef]
- Wang, L.; Chopp, M.; Szalad, A.; Lu, X.; Zhang, Y.; Wang, X.; Cepparulo, P.; Lu, M.; Li, C.; Zhang, Z.G. Exosomes Derived from Schwann Cells Ameliorate Peripheral Neuropathy in Type 2 Diabetic Mice. Diabetes 2020, 69, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chopp, M.; Liu, X.S.; Katakowski, M.; Wang, X.; Tian, X.; Wu, D.; Zhang, Z.G. Exosomes Derived from Mesenchymal Stromal Cells Promote Axonal Growth of Cortical Neurons. Mol. Neurobiol. 2017, 54, 2659–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, C.; Qin, Y.; Cepparulo, P.; Millman, M.; Chopp, M.; Kemper, A.; Szalad, A.; Lu, X.; Wang, L.; et al. Small extracellular vesicles ameliorate peripheral neuropathy and enhance chemotherapy of oxaliplatin on ovarian cancer. J. Extracell. Vesicles 2021, 10, e12073. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Substance Group | Chemotherapy Drugs | Drug Indications | References |
|---|---|---|---|
| Vinca alkaloids | Vincristine Vinblastine Vinorelbine | Acute leukemias, lymphogranulomatosis, non-Hodgkin’s lymphomas, multiple myeloma, Ewing’s sarcoma, osteogenic sarcoma, soft tissue sarcomas, breast cancer, small cell lung cancer, melanoma, bladder cancer, Wilms tumor, neuroblastoma, cervical cancer, uterine sarcoma | [23,24] |
| Platinum-based drugs | Cisplatin Carboplatin Oxaliplatin | Small and non-small cell lung cancer; squamous cell carcinoma of various localizations (brain, stomach, esophagus, bladder, cervix); ovarian cancer; osteosarcoma, colorectal cancer | [19,21] |
| Proteasome inhibitors | Bortezomib | Multiple myeloma | [17,18] |
| Taxanes | Paclitaxel, docetaxel | Breast cancer, ovarian cancer, gastric cancer | [20,21,22,23,24] |
| Non-taxane microtubule dynamics inhibitor | Eribulin | Breast cancer, liposarcomas, leiomyosarcomas | [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yunusova, N.V.; Popova, N.O.; Udintseva, I.N.; Klyushina, T.S.; Kazantseva, D.V.; Smirnova, L.P. The Role of Intravesicular Proteins and the Protein Corona of Extracellular Vesicles in the Development of Drug-Induced Polyneuropathy. Curr. Issues Mol. Biol. 2023, 45, 3302-3314. https://doi.org/10.3390/cimb45040216
Yunusova NV, Popova NO, Udintseva IN, Klyushina TS, Kazantseva DV, Smirnova LP. The Role of Intravesicular Proteins and the Protein Corona of Extracellular Vesicles in the Development of Drug-Induced Polyneuropathy. Current Issues in Molecular Biology. 2023; 45(4):3302-3314. https://doi.org/10.3390/cimb45040216
Chicago/Turabian StyleYunusova, Natalia V., Natalia O. Popova, Irina N. Udintseva, Tatyana S. Klyushina, Daria V. Kazantseva, and Liudmila P. Smirnova. 2023. "The Role of Intravesicular Proteins and the Protein Corona of Extracellular Vesicles in the Development of Drug-Induced Polyneuropathy" Current Issues in Molecular Biology 45, no. 4: 3302-3314. https://doi.org/10.3390/cimb45040216