

Transcriptome Analysis and VIGS Identification of Key Genes Regulating Citric Acid Metabolism in Citrus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Measurement of Total of Soluble Sugar (TSS) Titratable Acid (TA) in Citrus Flesh
2.3. Quantification of Organic Acids and Soluble Sugars in Citrus Fruit Flesh
2.4. RNA Extraction and Transcriptome Sequencing Data Comparison
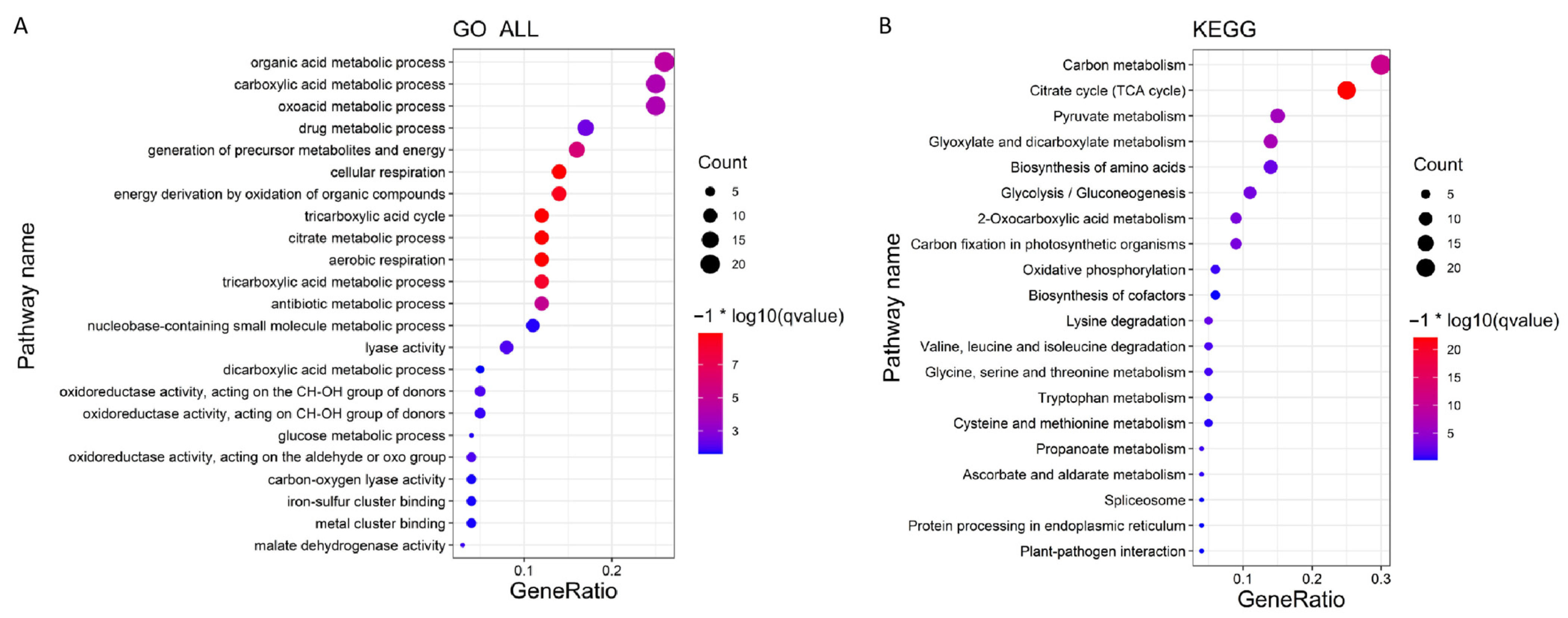
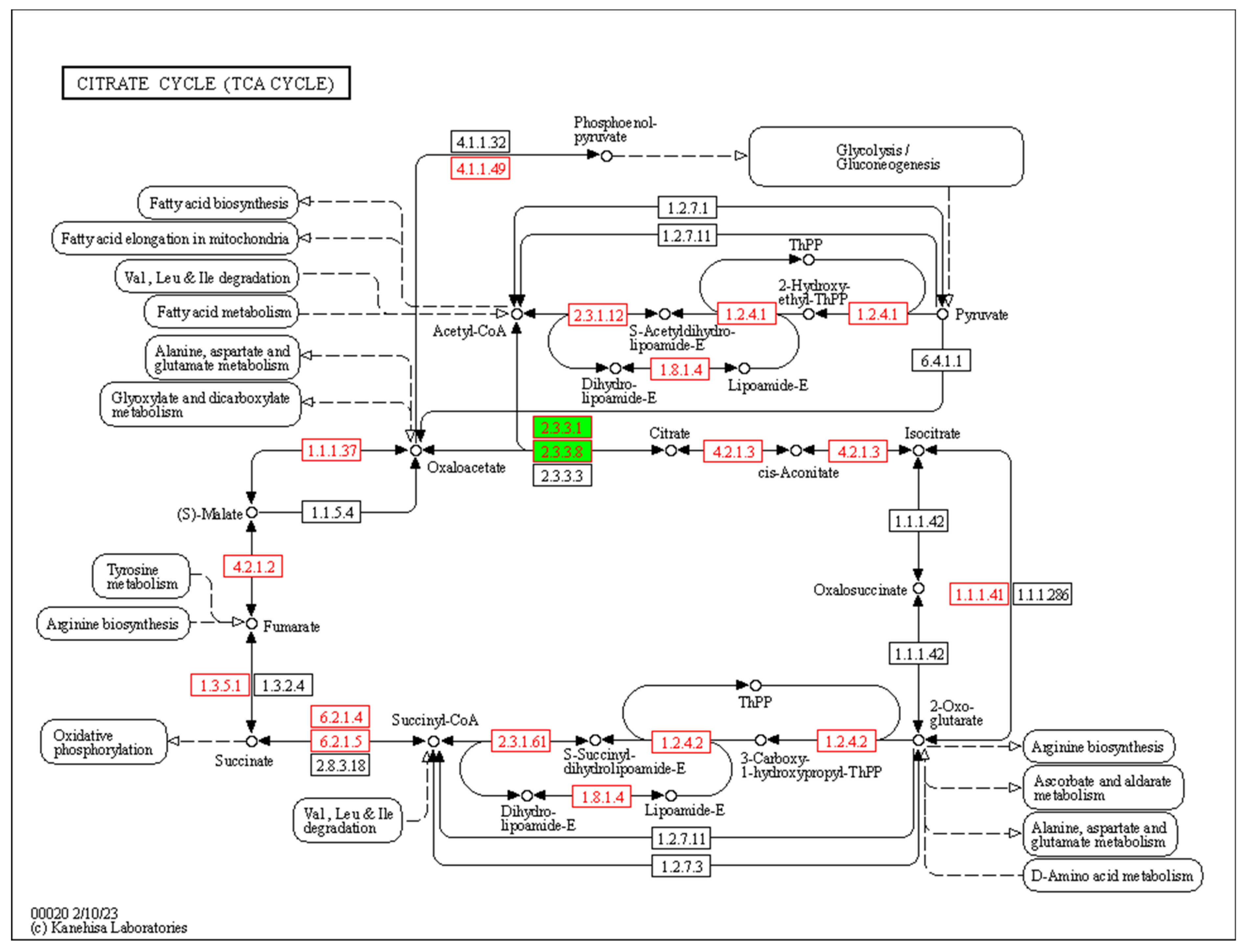
2.5. Analysis and Enrichment of Differential Expression Genes
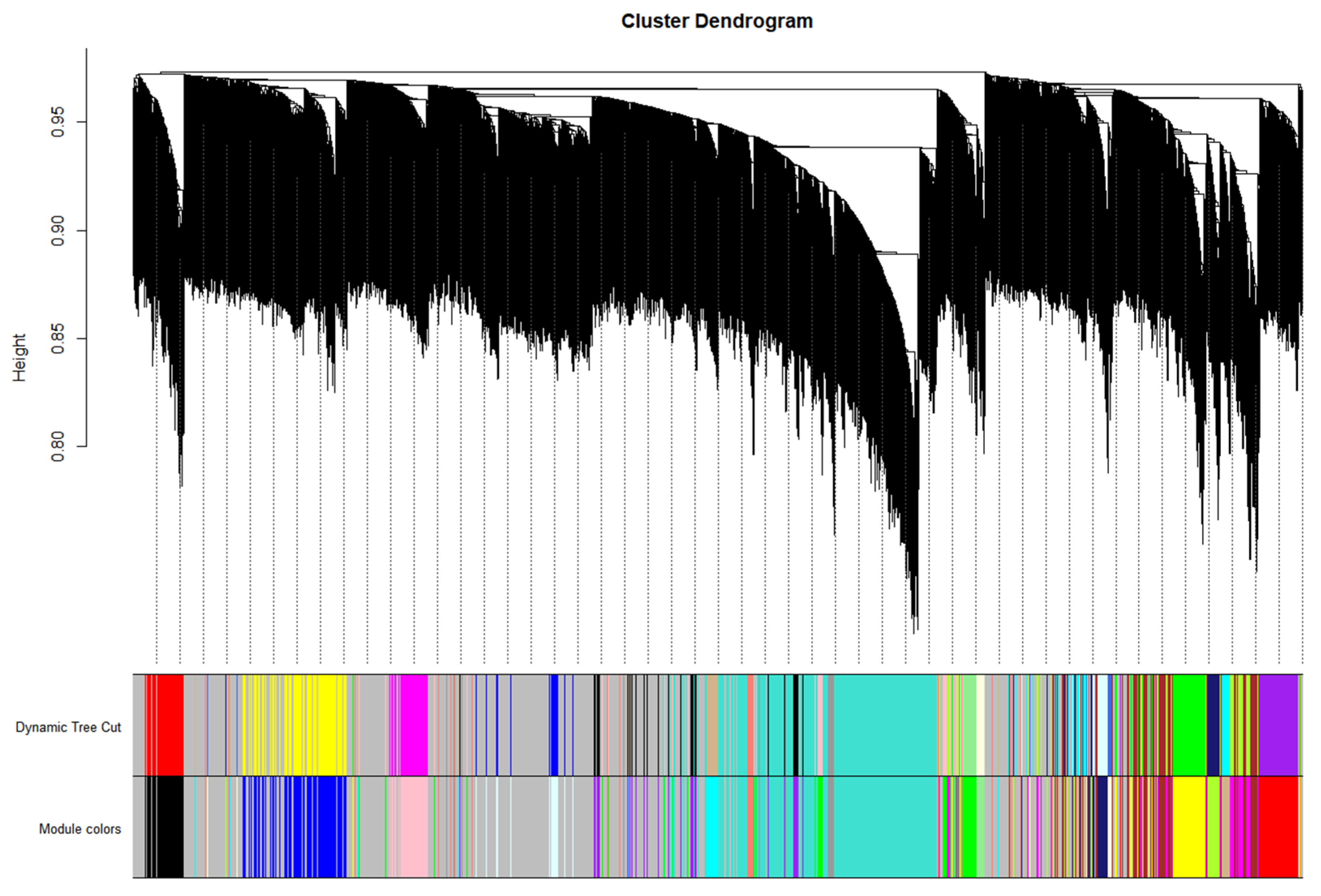
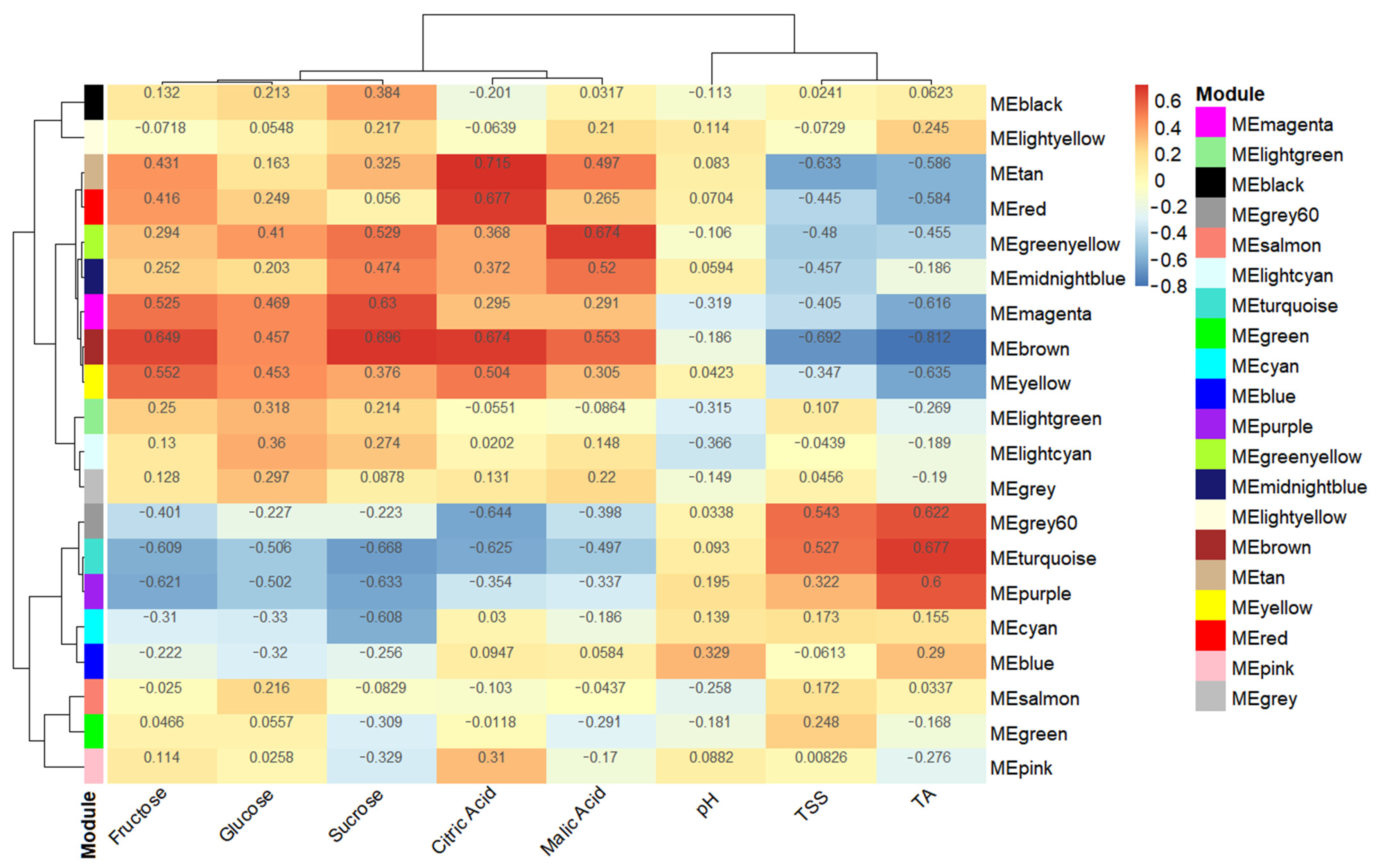
2.6. WGCNA for Identifying Hub Genes and Weight Modules
2.7. Validation of Intramodular Candidates through RT-qPCR Analysis
2.8. Construction of the VIGS Vector
2.9. Infection in Citrus Fruit with VIGS Vector
2.10. Validation of Quality Traits and VIGS Infection
3. Results
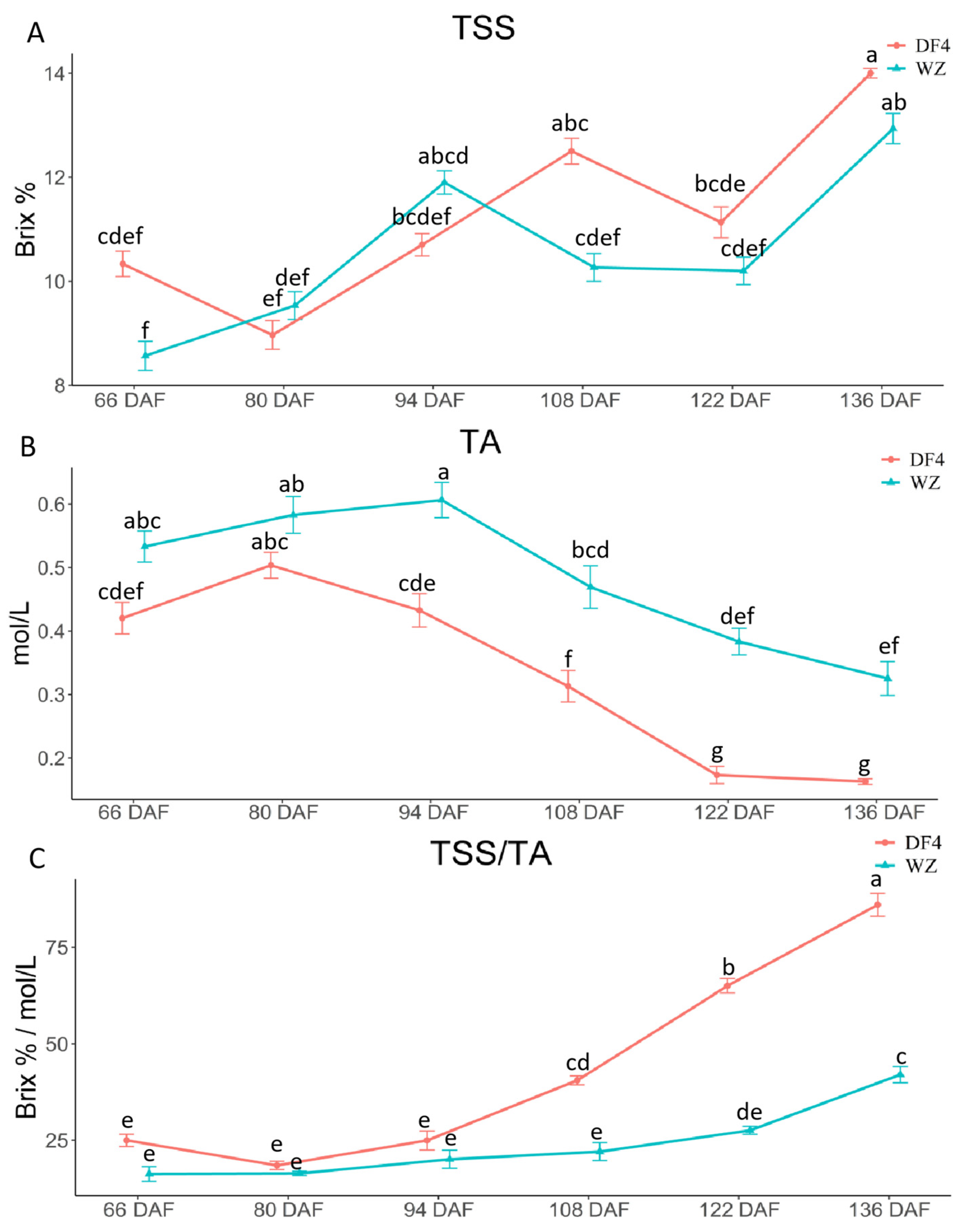
3.1. Variations among TSS, TA, and TSS/TA during Citrus Fruit Flesh Development
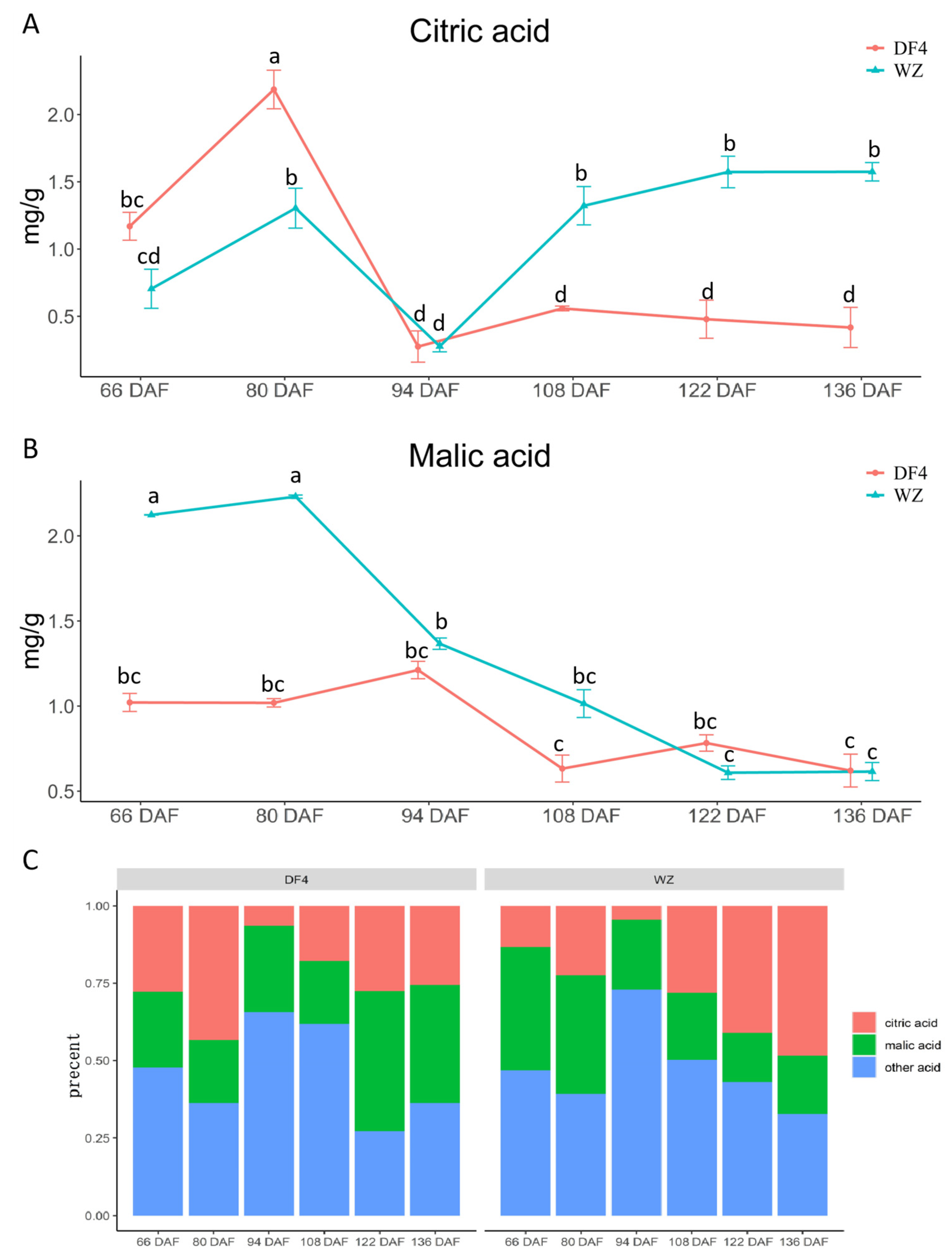
3.2. Variations of Minalic Acid and Citric Acid during Citrus Fruit Flesh Development
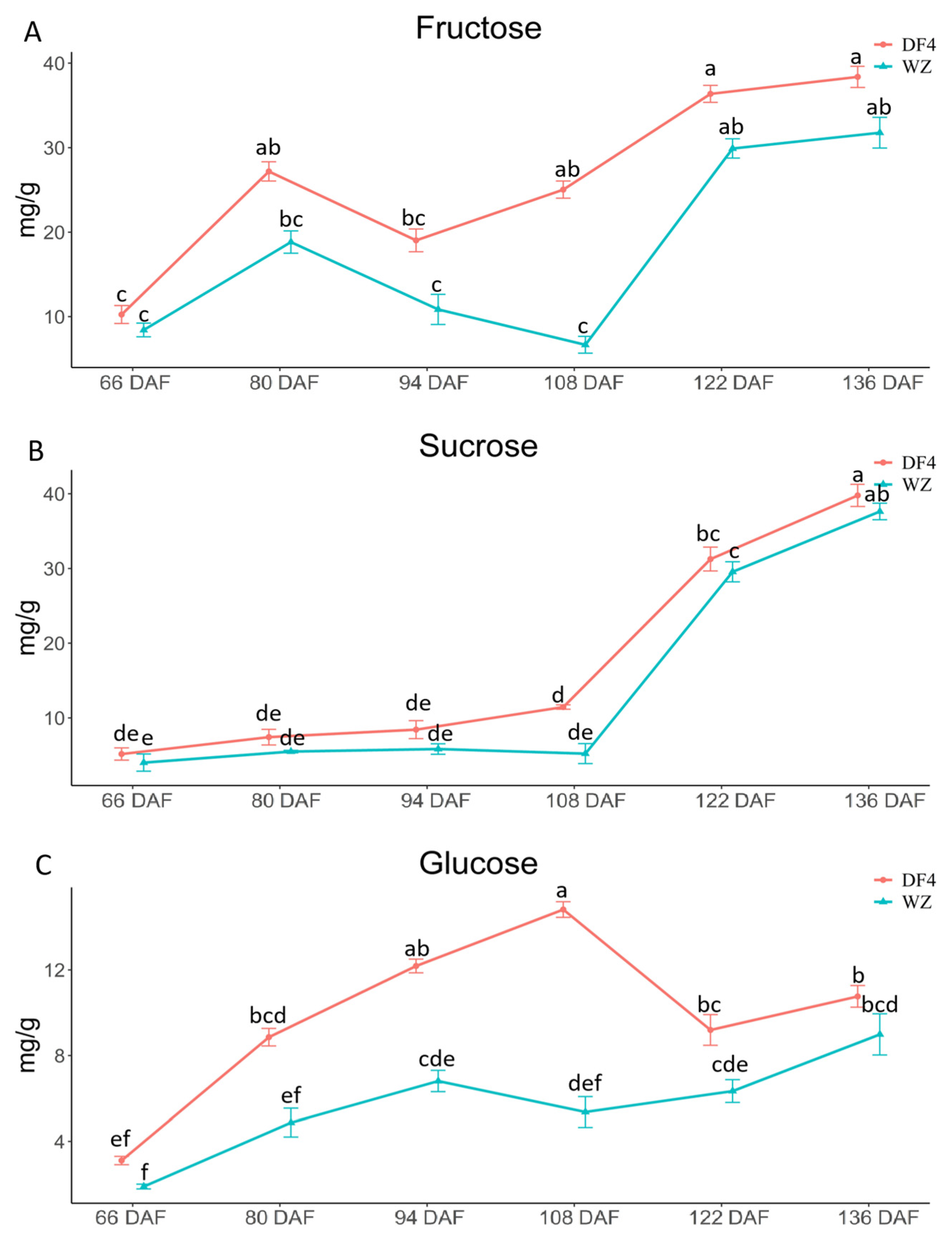
3.3. Variations in Fructose, Sucrose, and Glucose during Citrus Fruit Flesh Development
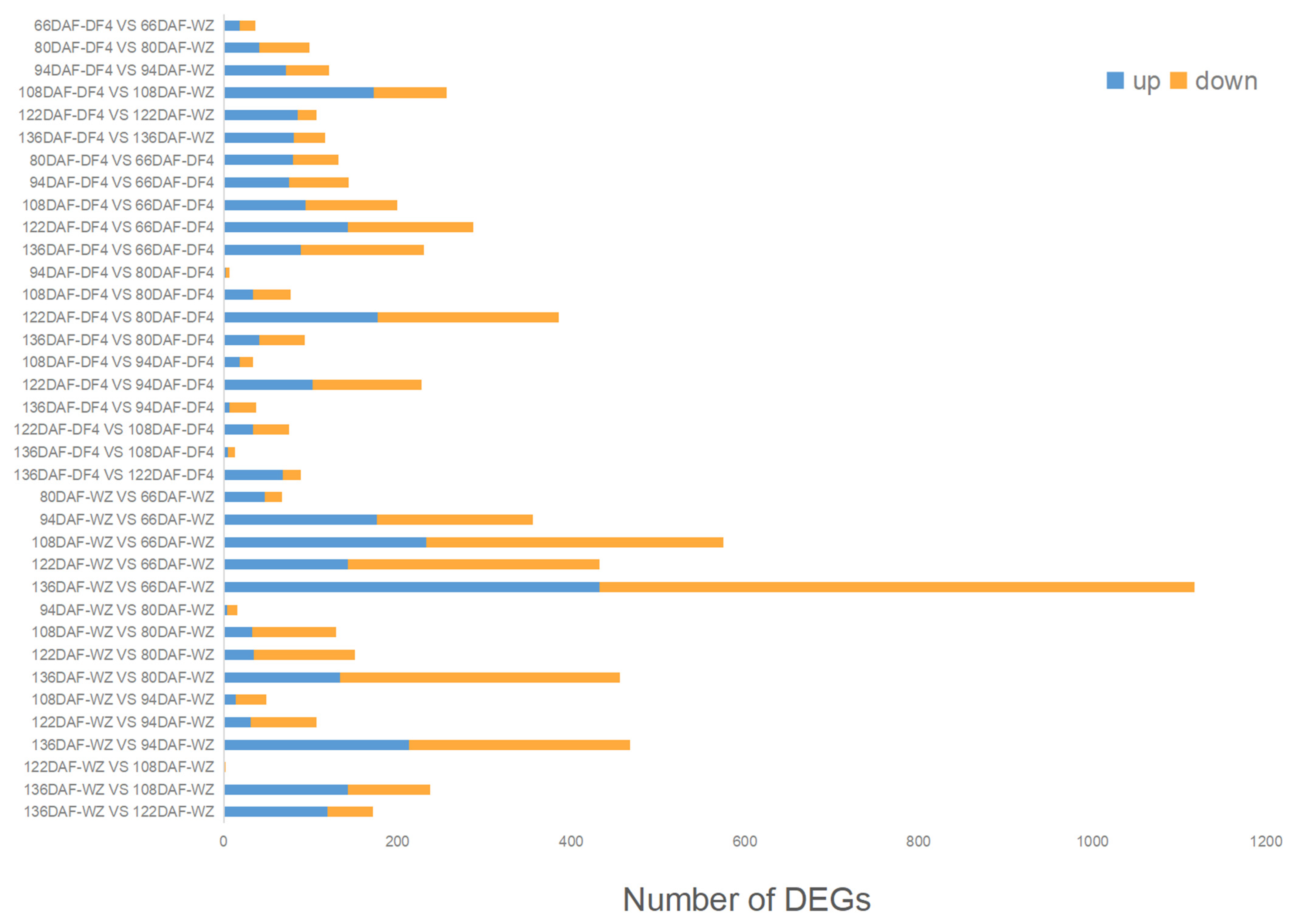
3.4. Identification and Analysis of DEGs at Different Fruit Developmental Stages
3.5. Obtaining Hub Genes during Weighted Correlation Network Analysis
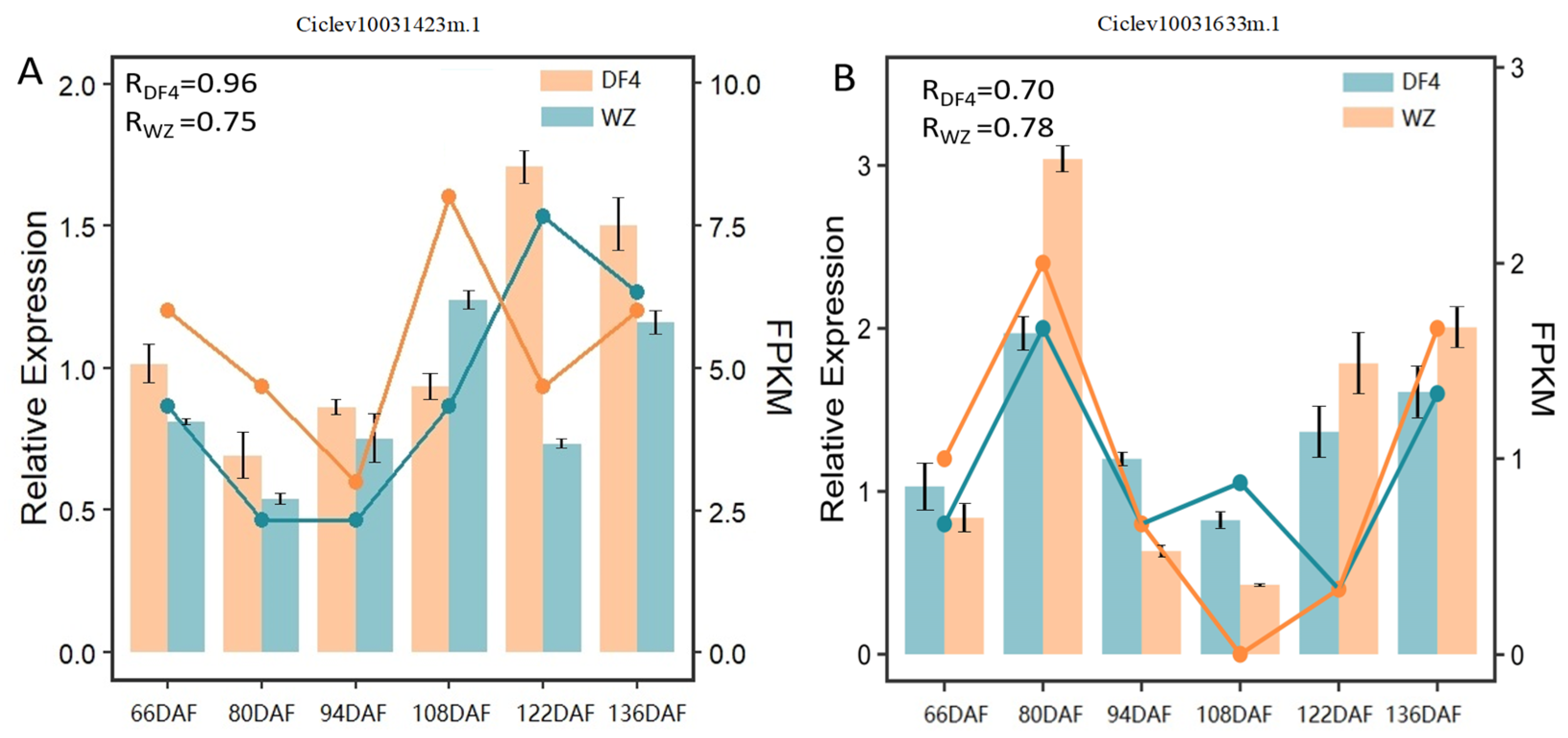
3.6. Screening Candidate Genes via RT-qPCR
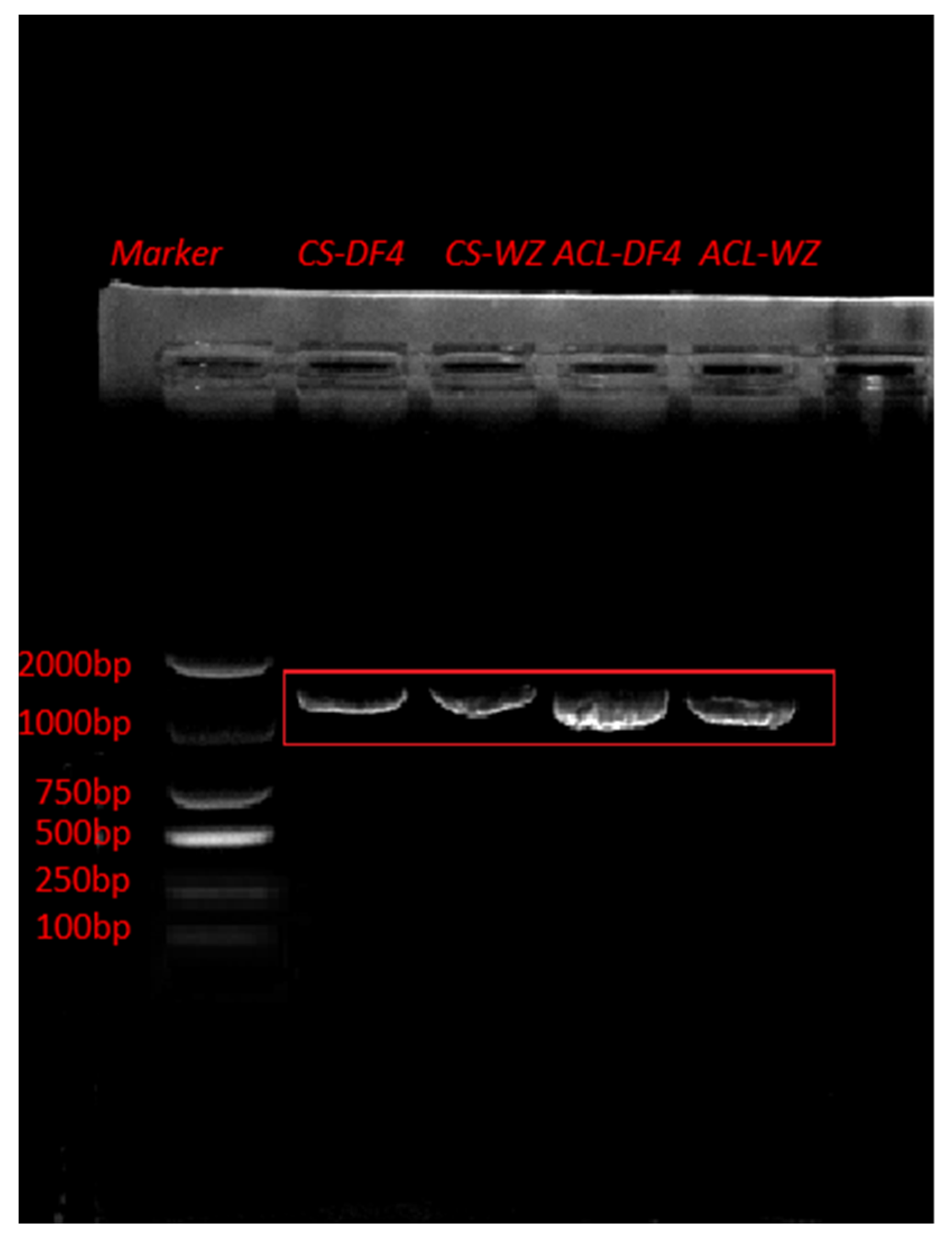
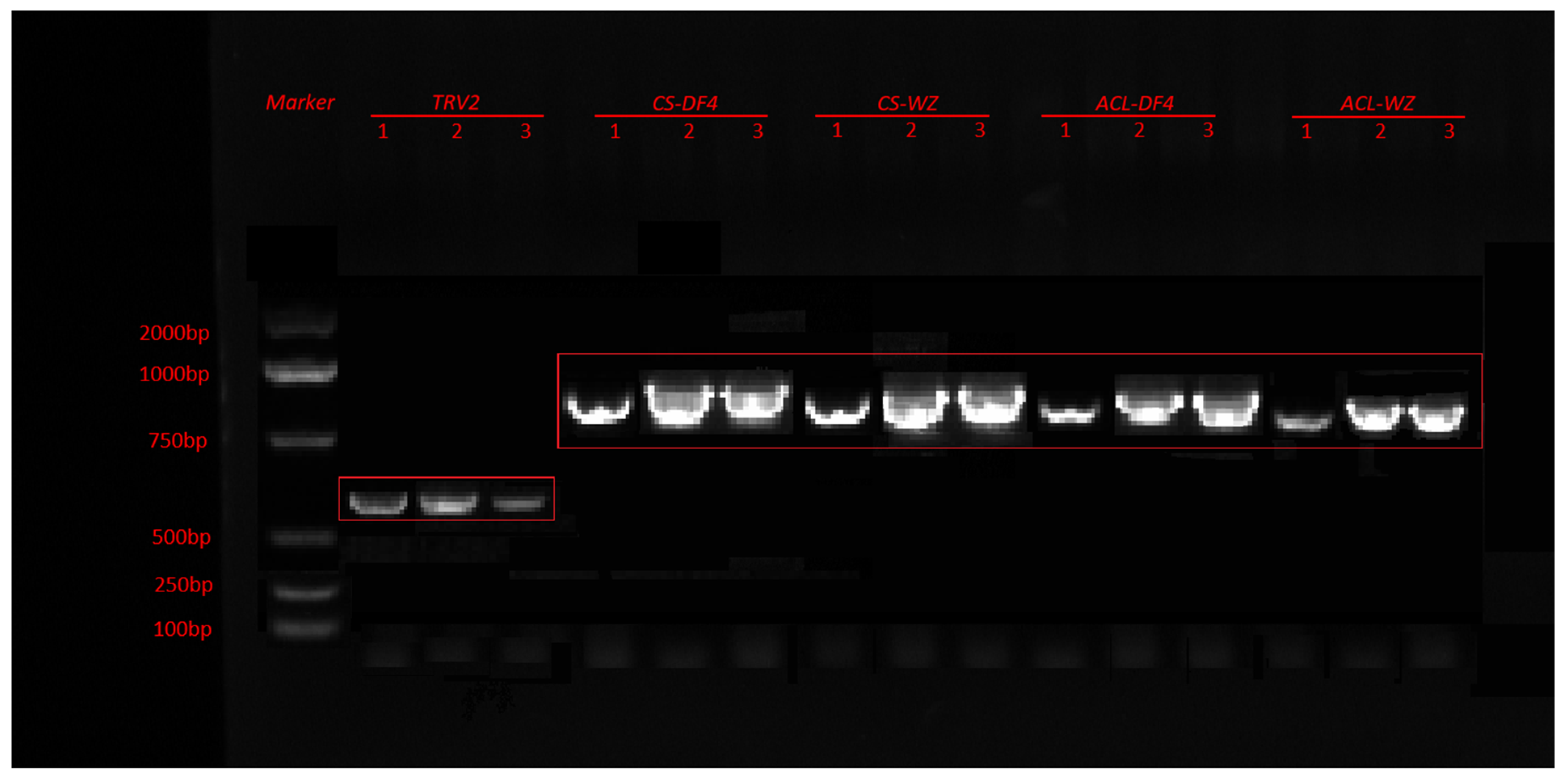
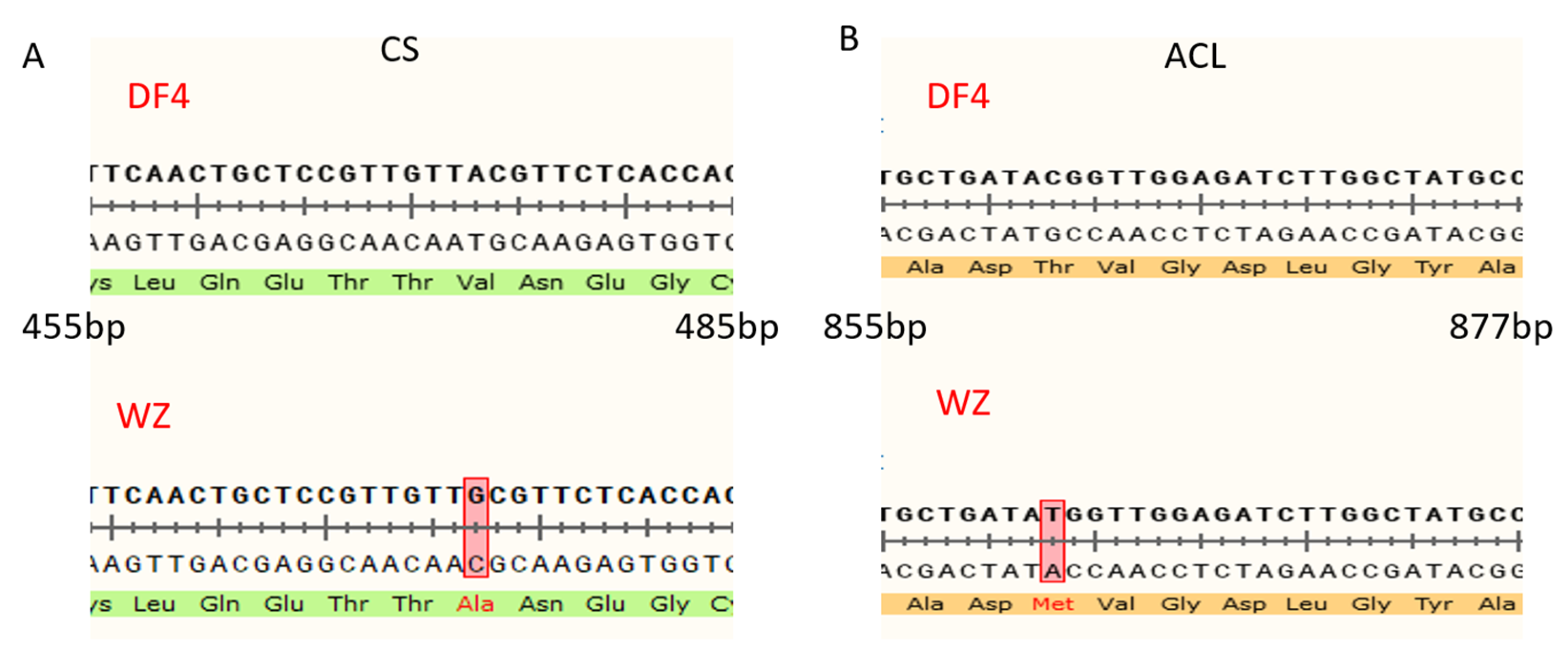
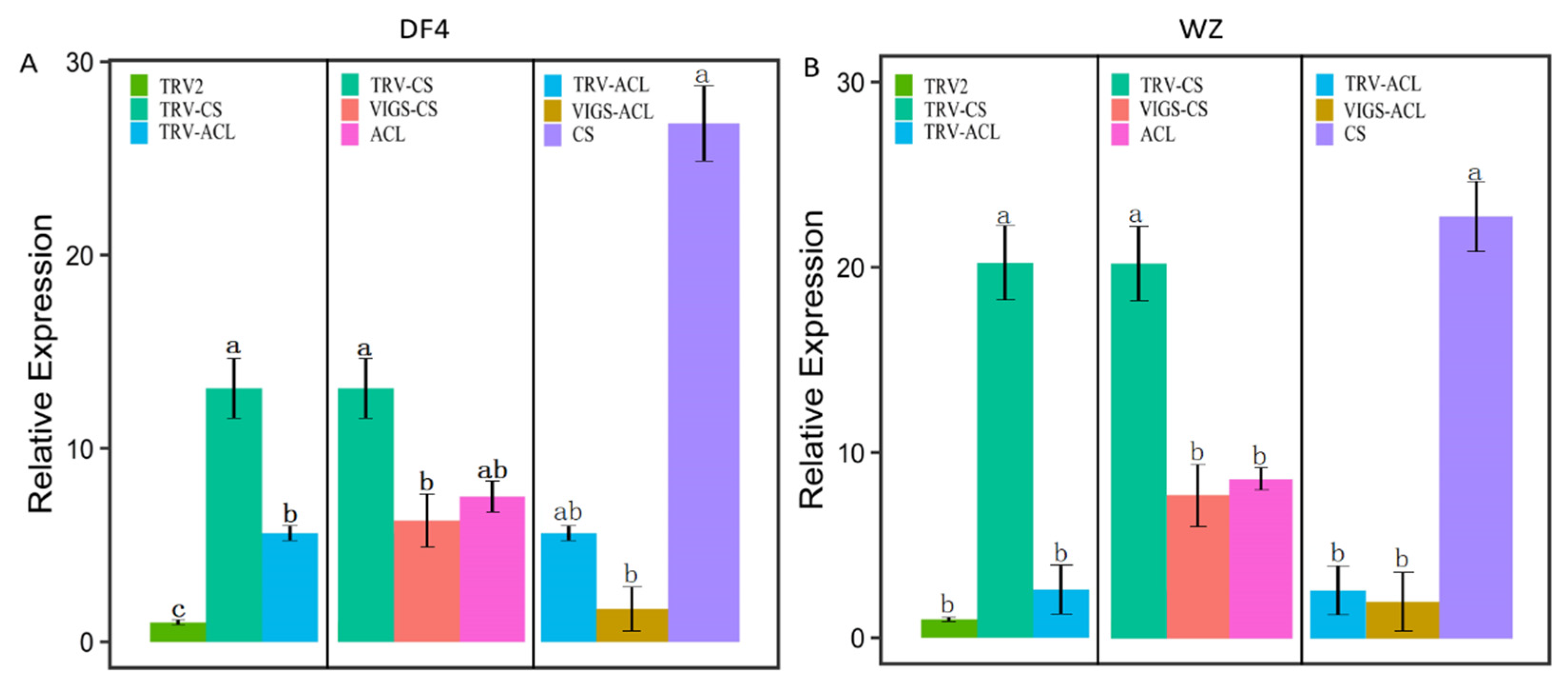
3.7. Construction of VIGS Vector and Verification of Impregnation Result
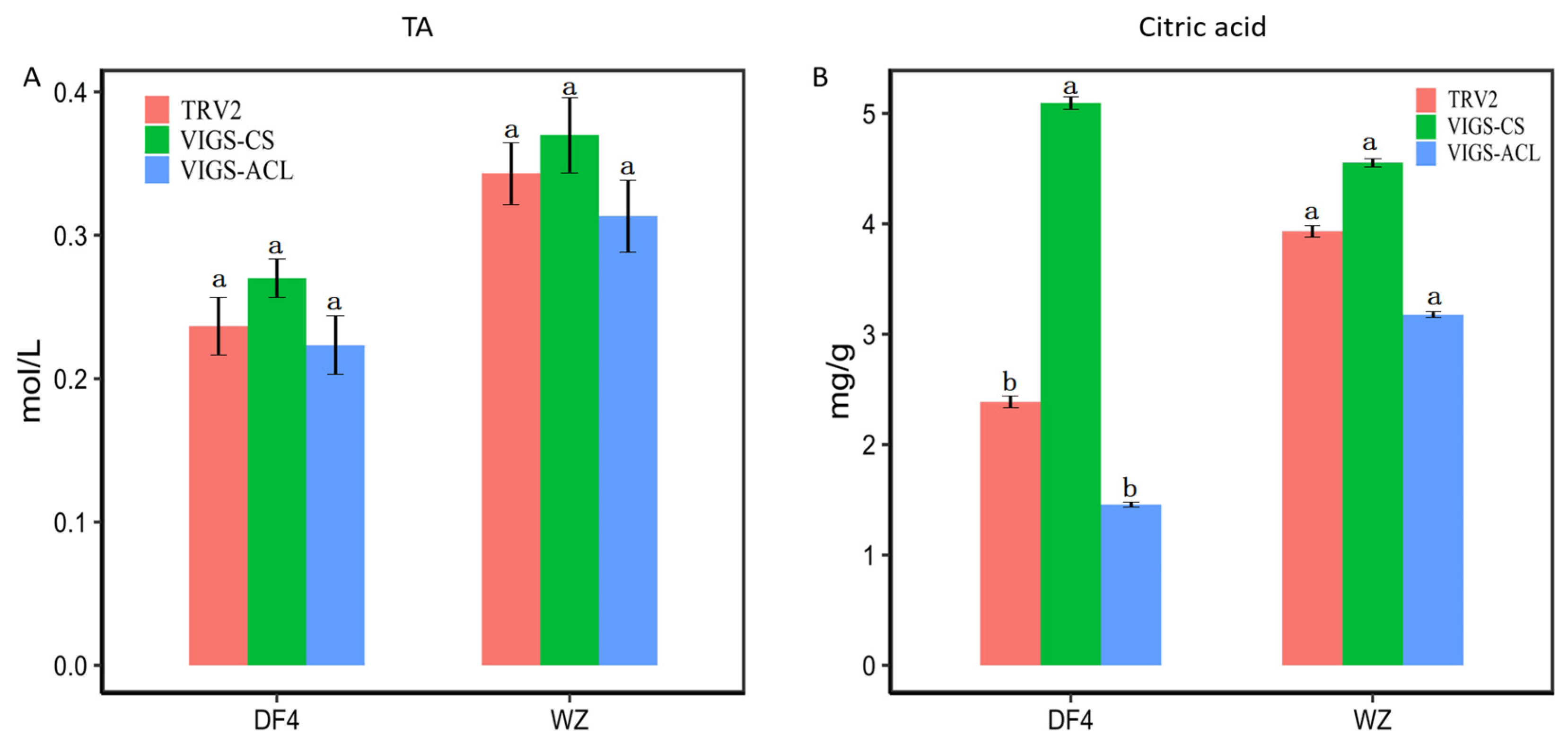
3.8. Effect of VIGS Infection on the Content of RT-qPCR Citric Acid and TA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, Q.; Chen, L.-L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.-B.; Hao, B.-H.; Lyon, M.P.; et al. The draft genome of sweet orange (Citrus sinensis). Nat. Genet. 2013, 45, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Kelebek, H.; Selli, S. Determination of volatile, phenolic, organic acid and sugar components in a Turkishcv Dortyol (Citrus sinensis L. Osbeck) orange juice. Sci. Food Agric. 2011, 91, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, L.; Yaniv, Y.; Porat, R.; Carmi, N. Mandarin fruit quality: A review. J. Sci. Food Agric. 2018, 98, 18–26. [Google Scholar] [CrossRef]
- Wang, T.; Lin, M.; Feng, X.; Wang, P.; Cao, X.; Zhang, W. Established methods and comparison of 10 organic acids based on reversed phase chromatography and hydrophilic interaction chromatography. CyTA J. Food 2022, 20, 206–217. [Google Scholar] [CrossRef]
- Yamaki, Y.T. Organic acids in the juice of citrus fruits. J. Jpn. Soc. Hortic. Sci. 1989, 58, 587–594. [Google Scholar] [CrossRef]
- Lado, J.; Gambetta, G.; Zacarias, L. Key determinants of citrus fruit quality: Metabolites and main changes during maturation. Sci. Hortic. 2018, 233, 238–248. [Google Scholar] [CrossRef]
- Emmanouilidou, M.G.; Kyriacou, M.C. Rootstock-modulated yield performance, fruit maturation and phytochemical quality of ‘Lane Late’ and ‘Delta’ sweet orange. Sci. Hortic. 2017, 225, 112–121. [Google Scholar] [CrossRef]
- Hussain, S.B.; Shi, C.-Y.; Guo, L.-X.; Kamran, H.M.; Sadka, A.; Liu, Y.-Z. Recent Advances in the Regulation of Citric Acid Metabolism in Citrus Fruit. Crit. Rev. Plant Sci. 2017, 36, 241–256. [Google Scholar] [CrossRef]
- Veldhuizen, M.G.; Siddique, A.; Rosenthal, S.; Marks, L.E. Interactions of Lemon, Sucrose and Citric Acid in Enhancing Citrus, Sweet and Sour Flavors. Chem. Senses 2018, 43, 17–26. [Google Scholar] [CrossRef]
- Oliver, D.J.; Nikolau, B.J.; Wurtele, E.S. Acetyl-CoA-Life at the metabolic nexus. Plant Sci. 2009, 176, 597–601. [Google Scholar] [CrossRef]
- Terrier, N.; Deguilloux, C.; Sauvage, F.X.; Martinoia, E.; Romieu, C. Proton pumps and anion transport in Vitis vinifera: The inorganic pyrophosphatase plays a predominant role in the energization of the tonoplast. Plant Physiol. Biochem. 1998, 36, 367–377. [Google Scholar] [CrossRef]
- Lee, W.S.; Rudd, J.J.; Kanyuka, K. Virus induced gene silencing (VIGS) for functional analysis of wheat genes involved in Zymoseptoria tritici susceptibility and resistance. Fungal. Genet. Biol. 2015, 79, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, R.; Shafiq, M.; Mansoor, S.; Briddon, R.W.; Scheffler, B.E.; Scheffler, J.; Amin, I. Virus-Induced Gene Silencing in Cultivated Cotton (Gossypium spp.) Using Tobacco Rattle Virus. Mol. Biotechnol. 2016, 58, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Orzaez, D.; Medina, A.; Torre, S.; Fernández-Moreno, J.P.; Rambla, J.L.; Fernández-Del-Carmen, A.; Butelli, E.; Martin, C.; Granell, A. A visual reporter system for virus-induced gene silencing in tomato fruit based on anthocyanin accumulation. Plant Physiol. 2009, 150, 1122–1134. [Google Scholar] [CrossRef] [PubMed]
- Ramegowda, V.; Senthil-Kumar, M.; Udayakumar, M.; Mysore, K.S. A high-throughput virus-induced gene silencing protocol identifies genes involved in multi- stress tolerance. BMC Plant Biol. 2013, 13, 193–210. [Google Scholar] [CrossRef]
- Renhui, D.; Ziniu, D.; Rencai, W.; Suming, D. Analysis of storage conditions of extra precocious Wenzhou mandarin Dafen 4. Hunan Agric. Sci. 2015, 4, 87–89, 92. [Google Scholar]
- Deyu, L.; Jianzhong, Z.; Hong, C.; Wengjing, L.; Zuyong, X. Demonstration and Popularization of Wenzhou mandarin tail as Interstock High grafting Yi Red Orange. Zhejiang Citrus 2011, 28, 18–19. [Google Scholar]
- Blanke, M. Non-invasive Assessment of Firmness and NIR Sugar (TSS) Measurement in Apple, Pear and Kiwi Fruit. Erwerbs-Obstbau 2013, 55, 19–24. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Utturkar, S.; Dassanayake, A.; Nagaraju, S.; Brown, S.D. Bacterial Differential Expression Analysis Methods. In Metabolic Pathway Engineering; Himmel, M., Bomble, Y., Eds.; Methods in Molecular Biology; Humana: New York, NY, USA, 2020; Volume 96, pp. 89–112. [Google Scholar]
- Yu, G.; He, Q. ReactomePA: An R/Bioconductor package for reactome pathway analysis and visualization. Mol. BioSyst. 2016, 12, 477–479. [Google Scholar] [CrossRef]
- Umer, M.J.; Bin Safdar, L.; Gebremeskel, H.; Zhao, S.; Yuan, P.; Zhu, H.; Kaseb, M.O.; Anees, M.; Lu, X.; He, N.; et al. Identification of key gene networks controlling organic acid and sugar metabolism during watermelon fruit development by integrating metabolic phenotypes and gene expression profiles. Hortic. Res. 2020, 7, 193. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Jawad, U.M.; Gao, L.; Gebremeskel, H.; Safdar, L.B.; Yuan, P.; Zhao, S.; Xuqiang, L.; Nan, H.; Hongju, Z.; Liu, W. Expression pattern of sugars and organic acids regulatory genes during watermelon fruit development. Sci. Hortic. 2020, 265, 109102. [Google Scholar]
- Wang, F.; Xu, Y.; Liu, X.; Shen, W.; Zhu, S.; Zhao, X. Use of TRV-mediated VIGS for functional genomics research in citrus. Plant Cell Tiss. Organ Cult. 2019, 139, 609–613. [Google Scholar] [CrossRef]
- Fantini, E.; Giuliano, G. Virus-Induced Gene Silencing as a Tool to Study Tomato Fruit Biochemistry. Plant Signal. Transduct. 2016, 1363, 65–78. [Google Scholar]
- Avi, S.; Lyudmila, S.; Itzhak, K.; Eduardo, B. Primary Metabolism in Citrus Fruit as Affected by Its Unique Structure. Front. Plant Sci. 2019, 10, 1167. [Google Scholar]
- Hussain, S.B.; Guo, L.X.; Shi, C.Y.; Khan, M.A.; Bai, Y.-X.; Du, W.; Liu, Y.-Z. Assessment of sugar and sugar accumulation-related gene expression profiles reveal new insight into the formation of low sugar accumulation trait in a sweet orange (Citrus sinensis) bud mutant. Mol. Biol. Rep. 2020, 47, 2781–2791. [Google Scholar] [CrossRef]
- Pangbom, R.M. Relative taste of selected sugars and organic acids. Food Sci. 1963, 28, 726–733. [Google Scholar] [CrossRef]
- Wang, L.; He, F.; Huang, Y.; He, J.; Yang, S.; Zeng, J.; Deng, C.; Jiang, X.; Fang, Y.; Wen, S.; et al. Genome of wild mandarin and domestication history of mandarin. Mol. Plant 2018, 11, 1024–1037. [Google Scholar] [CrossRef]
- Yehudit, A.; Candace, H.H.; Sarah, J.; Melody, W.; Deborah, P.D. A membrane-associated form of sucrose synthase and its potential role in synthesis of cellulose and callose in plants. Proc. Natl. Acad. Sci. USA 1995, 92, 9353–9357. [Google Scholar]
- Komatsu, A.; Takanokura, Y.; Moriguchi, T.; Omura, M.; Akihama, T. Differential expression of three sucrose-phosphate synthase isoforms during sucrose accumulation in citrus fruits (Citrus unshiu Marc.). Plant Sci. 1999, 140, 169–178. [Google Scholar] [CrossRef]
- Albertini, V.; Carcouet, E.; Pailly, O.; Gambotti, C.; Luro, F.; Berti, L. Changes in organic acids and sugars during early stages of development of acidic and acidless citrus fruit. Agric. Food Chem. 2006, 54, 8335–8339. [Google Scholar] [CrossRef]
- Shi, L.; Tu, B.P. Acetyl-CoA and the regulation of metabolism: Mechanisms and consequences. Curr. Opin. Cell Biol. 2015, 33, 125–131. [Google Scholar] [CrossRef]
- Loka, D.A.; Oosterhuis, D.M.; Baxevanos, D.; Noulas, C.; Hu, W. Single and combined effects of heat and water stress and recovery on cotton (Gossypium hirsutum L.) leaf physiology and sucrose metabolism. Plant Physiol. Biochem. 2020, 148, 166–179. [Google Scholar] [CrossRef]
- Ruan, Y.L. Sucrose Metabolism: Gateway to Diverse Carbon Use and Sugar Signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef]
- Hiratsuka, S.; Suzuki, M.; Nishimura, H.; Nada, K. Fruit photosynthesis in Satsuma mandarin. Plant Sci. 2015, 241, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Cao, X.; Li, F.; Li, J.; Xiong, J.; Long, G.; Cao, S.; Xie, S. Comparative transcriptome analysis reveals a global insight into molecular processes regulating citrate accumulation in sweet orange(Citrus sinensis). Physiol. Plant. 2016, 158, 463–482. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yin, X.; Xie, X.; Allan, A.C.; Ge, H.; Shen, S.; Chen, K.-S. The Citrus transcription factor, CitERF13,regulates citric acid accumulation via a protein-protein interaction with the vacuolar proton pump, CitVHA-c4. Sci. Rep. 2016, 6, 20151. [Google Scholar] [CrossRef]
- Lin, Q.; Li, S.; Dong, W.; Feng, C.; Yin, X.; Xu, C.; Sun, C.; Chen, K. Involvement of CitCHX and CitDIC in developmental-related and postharvest-hot-air driven citrate degradation in citrus fruits. PLoS ONE 2015, 10, 94–110. [Google Scholar] [CrossRef]
- Shi, C.; Song, R.; Hu, X.; Liu, X.; Jin, L.; Liu, Y. CitrusPH5-like H+-ATPase genes: Identification and transcript analysis to investigate their possible relationship with citrate accumulation in fruits. Front. Plant Sci. 2015, 6, 135. [Google Scholar] [CrossRef]
- Liu, X.; Hu, X.; Jin, L.; Shi, C.; Liu, Y.; Peng, S. Identification and transcript analysis of two glutamate decarboxylase genes, CsGAD1 and CsGAD2,reveal the strong relationship between CsGAD1 and citrate utilization in citrus fruit. Mol. Biol. Rep. 2014, 41, 6253–6262. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Xie, X.; Lin, Q.; Chen, J.; Grierson, D.; Yin, X.; Sun, C.; Chen, K. Differential expression of organic acid degradation-related genes during fruit development of Navel Oranges (Citrus sinensis) in two habitats. Plant Mol. Biol. Rep. 2013, 31, 1131–1140. [Google Scholar] [CrossRef]
- Chyper, M.; Zaidi, N.; Smans, K. ATP-citrate lyase: A mini-review. Biochem. Biophys. Res. Commun. 2012, 422, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Katz, E.; Fon, M.; Lee, Y.; Phinney, B.S.; Sadka, A.; Blumwald, E. The citrus fruit proteome: Insights into citrus fruit metabolism. Planta 2007, 226, 989–1005. [Google Scholar] [CrossRef]
- Rata, J.; Czak, R. Structure, function and regulation of the plant vacuolar H+-translocating ATPase. Biochim. Biophys. Acta Biomembr. 2000, 1465, 17–36. [Google Scholar]
- Usman, M.; Fatima, B. Mandarin (Citrus reticulata Blanco) Breeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer: Cham, Germany, 2018. [Google Scholar]
- Cuenca, J.; Garcia-Lor, A.; Navarro, L.; Aleza, P. Citrus Genetics and Breeding. In Advances in Plant Breeding Strategies: Fruits; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer: Cham, Germany, 2018. [Google Scholar]
- Khan, I.A.; Kender, W.J. Citrus breeding: Introduction and objectives. In Citrus Genetics, Breeding and Biotechnology; CABI Books; CABI International: Wallingford, UK, 2007. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, T.; Niu, J.; Sun, Z.; Chen, J.; Wang, Y.; Chen, J.; Luan, M. Transcriptome Analysis and VIGS Identification of Key Genes Regulating Citric Acid Metabolism in Citrus. Curr. Issues Mol. Biol. 2023, 45, 4647-4664. https://doi.org/10.3390/cimb45060295
Chen T, Niu J, Sun Z, Chen J, Wang Y, Chen J, Luan M. Transcriptome Analysis and VIGS Identification of Key Genes Regulating Citric Acid Metabolism in Citrus. Current Issues in Molecular Biology. 2023; 45(6):4647-4664. https://doi.org/10.3390/cimb45060295
Chicago/Turabian StyleChen, Tianxin, Juan Niu, Zhimin Sun, Jing Chen, Yue Wang, Jianhua Chen, and Mingbao Luan. 2023. "Transcriptome Analysis and VIGS Identification of Key Genes Regulating Citric Acid Metabolism in Citrus" Current Issues in Molecular Biology 45, no. 6: 4647-4664. https://doi.org/10.3390/cimb45060295
APA StyleChen, T., Niu, J., Sun, Z., Chen, J., Wang, Y., Chen, J., & Luan, M. (2023). Transcriptome Analysis and VIGS Identification of Key Genes Regulating Citric Acid Metabolism in Citrus. Current Issues in Molecular Biology, 45(6), 4647-4664. https://doi.org/10.3390/cimb45060295