
Inflammation Factors and Genistein Supplementation in Cancer—Preliminary Research

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Animals
2.2. Tumor Formation
2.3. Histopathology
2.4. Determination of 5-, 12-, and 15-Hydroxyeicosatetraenoic Acids (5-, 12-, and 15-HETE), 12-Hydroxyeicosapentaenoic Acid (12-HEPE), and Hydroxyoctadecadienoic Acids (HODE) in Rat Serum
2.5. Determination of the Contents of Interleukin-1, Interleukin-6, and Metalloproteinase-9 in the Serum of Rats
2.6. Determination of Gene Expression Activity of Interleukin-1, Interleukin-6, and Metalloproteinase-9 in Rat Blood
2.7. Statistical Analysis of the Obtained Research Results
3. Results
3.1. Effects of Nano-, Micro-, and Macrogenistein on the Development and Progression of DMBA-Induced Tumorigenesis in Rats
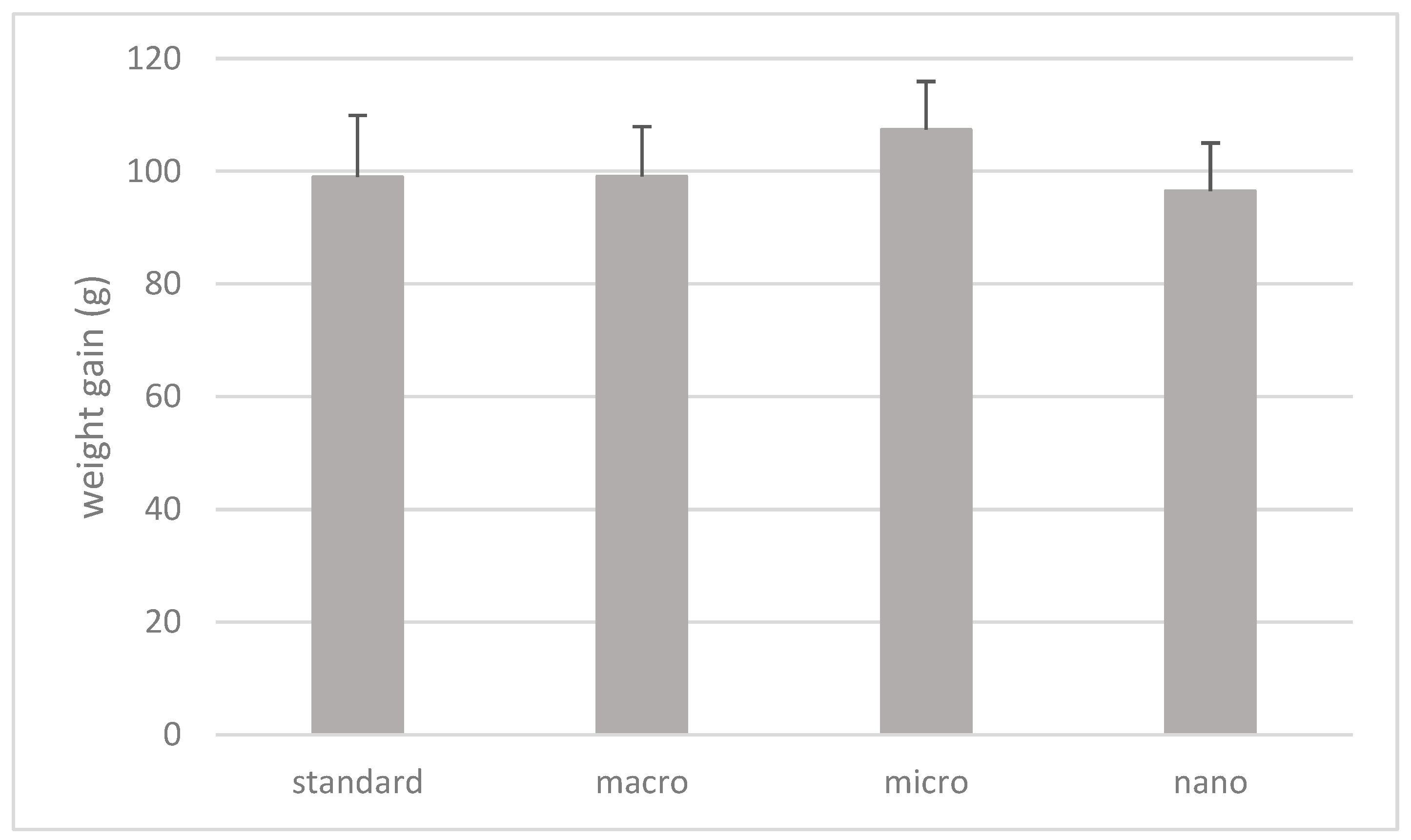
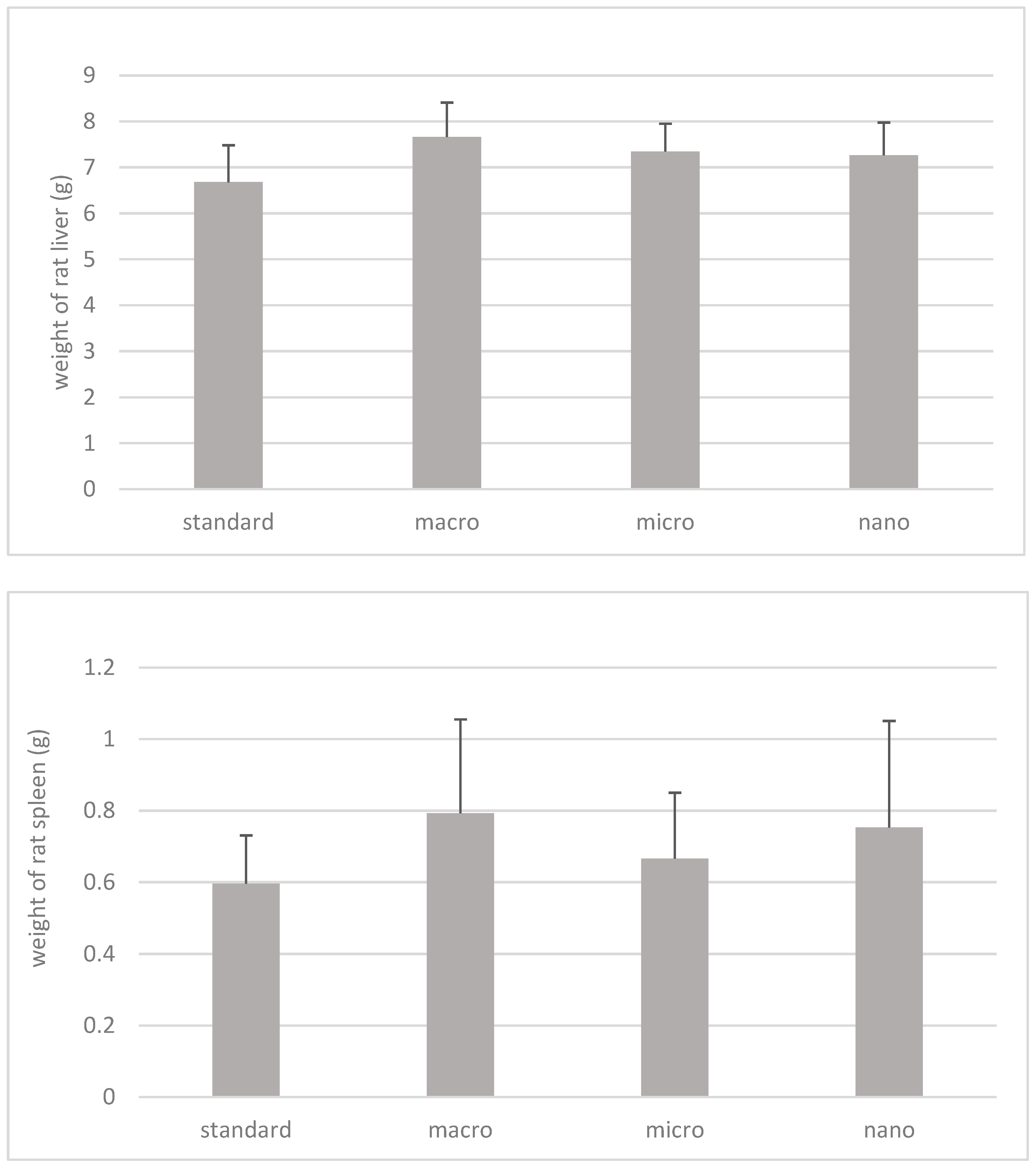
3.2. The Effect of Nano-, Micro-, and Macrogenistein Supplementation on the Body Weight of the Test Animals and the Weight of Their Organs
3.3. Effects of Nano-, Micro-, and Macrogenistein on the Content of Selected Fatty Acid Metabolites in the Serum of Rats Treated with DMBA
3.4. Effects of Nano-, Micro-, and Macrogenistein on the Contents of Interleukin-1, Interleukin-6, and Metalloproteinase-9 in the Serum of Rats Treated with DMBA
3.5. Effects of Nano-, Micro-, and Macrogenistein on Gene Expression of Interleukin-1, Interleukin-6, and Metalloproteinase-9 in the Blood Serum of Rats Treated with DMBA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharifi-Rad, J.; Quispe, C.; Imran, M.; Rauf, A.; Nadeem, M.; Gondal, T.A.; Ahmad, B.; Atif, M.; Mubarak, M.S.; Sytar, O.; et al. Genistein: An integrative overview of its mode of action, pharmacological properties, and health benefits. Oxid. Med. Cell. Longev. 2021, 30, 3268136. [Google Scholar] [CrossRef] [PubMed]
- Kołodziejczyk, J.; Olas, B.; Wachowicz, B. Właściwości lecznicze izoflawonów łagodzeniu objawów towarzyszących menopauzie i nie tylko. Kosmos 2012, 61, 331–339. [Google Scholar]
- Bhat, S.S.; Prasad, S.K.; Shivamallu, C.; Prasad, K.S.; Syed, A.; Reddy, P.; Cull, C.A.; Amachawadi, R.G. Genistein: A potent anti-breast cancer agent. Curr. Issues Mol. Biol. 2021, 43, 1502–1517. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Chatterton, R.T.; Michel, N.; Bryk, M.; Lee, O.; Ivancic, D.; Heinz, R.; Zalles, C.M.; Helenowski, I.B.; Jovanovic, B.D.; et al. Soy isoflavone supplementation for breast cancer risk reduction: A randomized phase II trial. Cancer Prev. Res. 2012, 5, 309–319. [Google Scholar] [CrossRef]
- Shike, M.; Doane, A.S.; Russo, L.; Cabal, R.; Reis-Filho, J.S.; Gerald, W.; Cody, H.; Khanin, R.; Bromberg, J.; Norton, L. The effects of soy supplementation on gene expression in breast cancer: A randomized placebo-controlled study. J. Natl. Cancer Inst. 2014, 106, dju189. [Google Scholar] [CrossRef]
- Lamartiniere, C.A.; Zhang, J.X.; Cotroneo, M.S. Genistein studies in rats: Potential for breast cancer prevention and reproductive and developmental toxicity. Am. J. Clin. Nutr. 1998, 68 (Suppl. S6), 1400S–1405S. [Google Scholar] [CrossRef]
- Ju, Y.H.; Allred, K.F.; Allred, C.D.; Helferich, W.G. Genistein stimulates growth of human breast cancer cells in a novel, postmenopausal animal model, with low plasma estradiol concentrations. Carcinogenesis 2006, 27, 1292–1299. [Google Scholar] [CrossRef]
- Liu, R.; Yua, X.; Chena, X.; Zhong, H.; Liang, C.; Xu, X.; Xu, W.; Cheng, Y.; Wang, W.; Yu, L.; et al. Individual factors define the overall effects of dietary genistein exposure on breast cancer patients. Nutr. Res. 2019, 67, 1–16. [Google Scholar] [CrossRef]
- Prescha, A.; Biernat, J. Wpływ fitoestrogenów pokarmowych na organizm człowieka. Cz. II. Przeciwdziałanie skutkom menopauzy oraz działanie przeciwnowotworowe. Bromat. Chem. Toksykol. 2008, 4, 941–948. [Google Scholar]
- Anastasiadis, S.P.; Chrissopoulou, K.; Stratakis, E.; Kavatzikidou, P.; Kaklamani, G.; Ranella, A. How the physicochemical properties of manufactured nanomaterials affect their performance in dispersion and their applications in biomedicine. Nanomaterials 2022, 12, 552. [Google Scholar] [CrossRef]
- Banys, K.; Giebultowicz, J.; Sobczak, M.; Wyrebiak, R.; Bielecki, W.; Wrzesien, R.; Bobrowska-Korczak, B. Effect of genistein supplementation on the progression of neoplasms and the level of the modified nucleosides in rats with mammary cancer. In Vivo 2021, 35, 2059–2072. [Google Scholar] [CrossRef]
- Takimoto, C.H.; Glover, K.; Huang, X.; Hayes, S.A.; Gallot, L.; Quinn, M.; Jovanovic, B.D.; Shapiro, A.; Hernandez, L.; Goetz, A.; et al. Phase I pharmacokinetic and pharmacodynamic analysis of unconjugated soy isoflavones administered to individuals with cancer. Cancer Epidem. Biomark. Prev. 2003, 12, 1213–1221. [Google Scholar]
- Frohberg, P.; Drutkowski, G.; Wobst, I. Monitoring eicosanoid biosynthesis via lipoxygenase and cyclooxygenase pathways in human whole blood by single HPLC run. J. Pharm. Biomed. Anal. 2006, 41, 1317–1324. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Walaszczyk, A.; Gabryś, D. Markery molekularne stosowane w diagnostyce raka piersi—Obecna praktyka kliniczna i perspektywy rozwoju. Nowotwory 2018, 3, 306–314. [Google Scholar]
- Cavalcanti, A.; Shirinzadeh, B.; Freitas, R.A.; Hogg, T. Nanorobot architecture for medical target identification. Nanotech 2008, 19, 015103. [Google Scholar] [CrossRef]
- Adair, J.H.; Parette, M.P.; Altinoglu, E.I.; Kester, M. Nanoparticulate alternatives for drug delivery. ACS Nano 2010, 4, 4967–4970. [Google Scholar] [CrossRef]
- Opolski, A.; Grynkiewicz, G.; Wietrzyk, J.; Radzikowski, C. Genisteina—Izoflawonoid soi o zróżnicowanym mechanizmie działania—Implikacje kliniczne w lecznictwie i prewencji chorób nowotworowych. Post. Hig. Med. Dośw. 2004, 58, 128–139. [Google Scholar]
- Mc Michael-Phillips, D.F.; Harding, C.; Morton, M.; Roberts, S.A.; Howell, A.; Potten, C.S.; Bundred, N.J. Effects of soy-protein supplementation on epithelial proliferation in the histologically normal human breast. Am. J. Clin. Nutr. 1998, 68, 1431–1435. [Google Scholar] [CrossRef]
- McCarty, M.F. Isoflavones made simple—genistein’s agonist activity for the beta-type estrogen receptor mediates their health benefits. Med. Hypotheses 2006, 66, 1093–1114. [Google Scholar] [CrossRef]
- Pons, D.G.; Nadal-Serrano, M.; Torrens-Mas, M.; Oliver, J.; Roca, P. The Phytoestrogen Genistein Affects Breast Cancer Cells Treatment Depending on the ERα/ERβ Ratio. J. Cell Biochem. 2016, 117, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Hilakivi-Clarke, L.; Andrade, J.E.; Helferich, W. Is Soy Consumption Good or Bad for the Breast? J. Nutr. 2010, 140, 2326–2334. [Google Scholar] [CrossRef]
- Francuz, T.; Czajka-Francuz, P.; Cisoń-Juek, S.; Wojnar, J. Rola zapalenia w patogenezie raka jelita grubego. Post. Hig. Med. Dosw. 2016, 70, 360–366. [Google Scholar] [CrossRef]
- Greene, E.R.; Huang, S.; Serhan, C.N.; Panigrahy, D. Regulation of inflammation in cancer by eicosanoids. Prostaglandins Other Lipid Mediat. 2011, 96, 27–36. [Google Scholar] [CrossRef]
- Calder, P.C. Eicosanoids. Essays Biochem. 2020, 64, 423–441. [Google Scholar] [PubMed]
- Banaszczak, M.; Stachowska, E. Patogeneza NAFLD w świetle wyników najnowszych badań. Post. Biochem. 2017, 63, 190–197. [Google Scholar]
- Klawitter, J.; Zafar, I.; Klawitter, J.; Pennington, A.T.; Klepacki, J.; Gitomer, B.Y.; Schrier, R.W.; Christians, U.; Edelstrin, C.L. Effects of lovastatin treatment on the metabolic distributions in the Han: SPRD rat model of polycystic kidney disease. BMC Nephrol. 2013, 14, 165. [Google Scholar] [CrossRef] [PubMed]
- Vang, K.; Ziboh, V. 15-Lipoxygenase metabolites of γ-linolenic acid/eicosapentaenoic acid suppress growth and arachidonic acid metabolism in human prostatic adenocarcinoma cells: Possible implications of dietary fatty acids. Prostaglandins Leukot. Essent. Fat. Acids 2005, 72, 363–372. [Google Scholar] [CrossRef]
- Tavakoli-Yaraki, M.; Karami-Tehrani, F. Apoptosis Induced by 13-S-hydroxyoctadecadienoic acid in the breast cancer cell lines, MCF-7 and MDA-MB-231. Iran. J. Basic Med. Sci. 2013, 16, 653–659. [Google Scholar]
- Kelavkar, U.; Glasgow, W.; Eling, T.E. The effect of 15-lipoxygenase-1 expression on cancer cells. Curr. Urol. Rep. 2002, 3, 207–214. [Google Scholar] [CrossRef]
- Jiang, W.G.; Watkins, G.; Douglas-Jones, A.; Mansel, R.E. Reduction of isoforms of 15-lipoxygenase (15-LOX)-1 and 15-LOX-2 in human breast cancer. Prostaglandins Leukot. Essent. Fat. Acids 2006, 74, 235–245. [Google Scholar] [CrossRef]
- Vaezi, M.A.; Safizadeh, B.; Eghtedari, A.R.; Ghorbanhosseini, S.S.; Rastegar, M.; Salimi, V.; Tavakoli-Yaraki, M. 15-Lipoxygenase and its metabolites in the pathogenesis of breast cancer: A double-edged sword. Lipids Health Dis. 2021, 20, 169. [Google Scholar] [CrossRef]
- Larsson, S.C.; Kumlin, M.; Ingelman-Sundberg, M.; Wolk, A. Dietary long-chain n-3 fatty acids for the prevention of cancer: A review of potential mechanisms. Am. J. Clin. Nutr. 2004, 79, 935–945. [Google Scholar] [CrossRef]
- Mahesha, H.G.; Singh, S.A.; Rao, A.G. Inhibition of lipoxygenase by soy isoflavones: Evidence of isoflavones as redox inhibitors. Arch. Biochem. Biophys. 2007, 15, 176–185. [Google Scholar] [CrossRef]
- Chechlińska, M. Rola cytokin w procesach nowotworzenia. J. Oncol. 2003, 53, 648–659. [Google Scholar]
- Łukaszewicz, M.; Mroczko, B.; Szmitkowski, M. Znaczenie kliniczne interlrukiny 6 (IL-6) jako czynnika rokowniczego w chorobie nowotworowej. Pol. Arch. Med. Wewnętrznej 2007, 117, 247–251. [Google Scholar]
- Metrowska, P.; Metrowski, S.; Smarz-Widelska, I.; Grywalska, E. Biological role, mechanism of action and the importance of interleukins in kidney diseases. Int. J. Mol. Sci. 2022, 23, 647. [Google Scholar]
- Wu, T.C.; Xu, K.; Martinek, J.; Banchereau, R.; George, J.; Turner, J.; Kim, K.I.; Zurawski, S.; Wang, X.; Blankenship, D.; et al. IL1 receptor antagonist controls transcriptional signature of inflammation in patients with metastatic breast cancer. Cancer Res. 2018, 1578, 5243–5258. [Google Scholar] [CrossRef]
- Dinarello, C.A. An interleukin-1 signature in breast cancer treated with interleukin-1 receptor blockade: Implications for treating cytokine release syndrome of checkpoint inhibitors. Cancer Res. 2018, 78, 5200–5202. [Google Scholar] [CrossRef]
- Kozłowski, L.; Zakrzewska, I.; Tokajuk, P. Concentration of interleukin-6 (IL-6), interleukin-8 (IL-8) and interleukin-10 (IL-10) in blood serum of breast cancer patients. Rocz. Akad. Med. W Bialymst. 2003, 48, 82–84. [Google Scholar]
- Gelfo, V.; Romaniello, D.; Mazzeschi, M.; Sgarzi, M.; Grilli, G.; Morselli, A.; Manzan, B.; Rihawi, K.; Lauriola, M. Roles of IL-1 in Cancer: From tumor progression to resistance to targeted therapies. Int. J. Mol. Sci. 2020, 21, 6009. [Google Scholar] [CrossRef]
- Shibayama, O.; Yoshiuchi, K.; Inagaki, M.; Matsuoka, Y.; Yoshikawa, E.; Sugawara, Y.; Akechi, T.; Wada, N.; Imoto, S.; Murakami, K.; et al. Association between adjuvant regional radiotherapy and cognitive function in breast cancer patients treated with conservation therapy. Cancer Med. 2014, 3, 702–709. [Google Scholar] [CrossRef]
- Tang, S.C.; Hsiao, Y.P.; Ko, J.L. Genistein protects against ultraviolet B- induced wrinkling and photoinflammation in in vitro and in vivo models. Genes Nutr. 2022, 17, 4. [Google Scholar] [CrossRef]
- Lee, S.R.; Kwon, S.W.; Lee, Y.H.; Kaya, P.; Kim, J.M.; Ahn, C.; Jung, E.M.; Lee, G.S.; An, B.S.; Jeung, E.B.; et al. Dietary intake of genistein suppresses hepatocellular carcinoma through AMPK-mediated apoptosis and anti-inflammation. BMC Cancer 2019, 19, 6. [Google Scholar] [CrossRef]
- Pierzynowska, K.; Podlacha, M.; Brokowska, J.; Gaffke, L.; Mantej, J.; Cyske, Z.; rintz, E.; Osiadły, M.; Bartkowski, M.; Puchalski, M.; et al. Molekularne mechanizmy działania genisteiny w świetle terapii chorób genetycznych i immunologicznych. Post. Biochem. 2018, 64, 262–275. [Google Scholar]
- Kołaczkowska, E. Metaloproteinaza 9 (MMP-9) jako szczególny przedstawiciel metaloproteinaz macierzy zewnątrzkomórkowej: Rola w napływie i apoptozie neutrofili w trakcie reakcji zapalnej. Post. Biol. Kom 2010, 37, 471–499. [Google Scholar]
- Zaręba, I.; Donejko, M.; Rysiak, E. Znaczenie i przydatność diagnostyczna mttaloproteinaz w raku piersi. J. Oncol. 2014, 64, 491–495. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | Assay ID |
|---|---|---|
| Matrix metallopeptidase-9 | MMP-9 | Rn00579162_m1 |
| Interleukin-1 beta | IL-1b | Rn00580432_m1 |
| Interleukin-6 | IL-6 | Rn01410330_m1 |
| Glyceraldehyde-3-phosphate dehydrogenase | GAPDH | Rn01775763_g1 |
| Actin beta | ACTB | Rn00667869_m1 |
| Supplementation | Rat’s Number | Tumor Weight (g) (Mean ± SD) (Week 20) | Number of Tumors per Rat (Week 20) | Incidence of Tumors per Rat in Time (Weeks of Rats Life) |
|---|---|---|---|---|
| Standard | 1 | 0.90 ± 0.78 (0.1–2.7) | 9 | 17 |
| 2 | 1.84 ± 3.35 (0.1–7.8) | 5 | 16 | |
| 3 | 0.43 ± 0.30 (0.1–0.8) | 4 | 19 | |
| 4 | 0.45 ± 0.40 (0.1–1.1) | 6 | 18 | |
| 5 | 1.02 ± 1.03 (0.1–2.6) | 6 | 16 | |
| 6 | 0.60 ± 0.00 (0.6) | 2 | 19 | |
| 7 | 1.36 ± 0.92 (0.3–1.96) | 3 | 17 | |
| 8 | 0.61 ± 0.20 (0.47–0.75) | 2 | 17 | |
| mean ± SD | 0.93 ± 1.34 (0.10–7.80) a | |||
| Macrogenistein | 1 | 2.35 ± 1.96 (0.9–5.14) | 4 | 17 |
| 2 | 0.52 ± 0.47 (0.14–1.38) | 6 | 17 | |
| 3 | 0.70 ± 0.52 (0.33–1.29) | 3 | 19 | |
| 4 | 1.83 ± 2.48 (0.13–6.39) | 6 | 18 | |
| 5 | 1.74 ± 1.79 (0.24–4.07) | 4 | 17 | |
| 6 | 0.34 | 1 | 19 | |
| 7 | 1.58 ± 1.14 (0.29–2.42) | 3 | 17 | |
| 8 | 1.13 ± 1.28 (0.25–3.03) | 4 | 17 | |
| mean ± SD | 1.27 ± 1.52 (0.14–6.39) | |||
| Microgenistein | 1 | 0.89 ± 0.40 (0.6–1.17) | 2 | 20 |
| 2 | - | - | - | |
| 3 | 6.11 | 1 | 18 | |
| 4 | 1.38 ± 0.69 (0.89–1.86) | 2 | 20 | |
| 5 | 3.31 | 1 | 17 | |
| 6 | 2.77 ± 1.81 (0.69–3.89) | 3 | 18 | |
| 7 | 1.45 ± 1.40 (0.19–2.96) | 3 | 18 | |
| 8 | 0.66 ± 0.78 (0.11–1.21) | 2 | 19 | |
| mean ± SD | 1.99 ± 1.75 (0.11–6.11) a | |||
| Nanogenistein | 1 | 1.01 ± 1.02 (0.1–2.43) | 4 | 19 |
| 2 | 4.58 ± 2.12 (3.08–6.08) | 2 | 17 | |
| 3 | 0.28 ± 0.20 (0.09–0.48) | 3 | 19 | |
| 4 | 0.20 ± 0.07 (0.1–0.26) | 4 | 20 | |
| 5 | 1.92 ± 3.74 (0.11–8.61) | 5 | 14 | |
| 6 | 1.57 ± 1.94 (0.06–4.41) | 4 | 18 | |
| 7 | 5.03 ± 6.32 (0.56–9.50) | 2 | 18 | |
| 8 | 0.30 ± 0.16 (0.18–0.41) | 2 | 20 | |
| mean ± SD | 1.59 ± 2.64 (0.06–9.50) |
| Supplementation | Tumor Grade | The Mean Number of Mitoses in the Field of View Area * |
|---|---|---|
| Standard | Adenocarcinoma grade 2 | 1.79 ± 1.25 a,b,c |
| Macrogenistein | Adenocarcinoma grade 2 | 4.46 ± 2.38 a,d |
| Microgenistein | Adenocarcinoma grade 3 | 7.33 ± 1.57 b,d |
| Nanogenistein | Adenocarcinoma grade 3 | 5.82 ± 1.57 c |
| Eicosanoids | Group | p-Value | |||
|---|---|---|---|---|---|
| Standard | Macro | Micro | Nano | ||
| 12-HEPE | 458.19 ± 208.09 b | 69.45 ± 36.42 a | 61.07 ± 12.68 a | 60.21 ± 14.18 a | 0.0013 |
| HODE | 226.82 ± 33.67 b | 110.60 ± 48.13 a | 130.52 ± 41.90 a | 116.13 ± 32.58 a | 0.0001 |
| 15-HETE | 43.10 ± 7.58 b | 29.06 ± 6.49 a | 36.82 ± 14.09 ab | 36.18 ± 8.68 ab | n.s. |
| 12-HETE | 6446 ± 1522 b | 2414 ± 917 a | 1918 ± 219 a | 1897 ± 319 a | 0.0001 |
| 5-HETE | 10.94 ± 2.81 | 6.84 ± 3.64 | 9.29 ± 6.25 | 15.73 ± 14.17 | n.s. |
| Eicosanoids | Group | p-Value | |||
|---|---|---|---|---|---|
| Standard | Macro | Micro | Nano | ||
| IL-6 | 88.47 ± 40.39 | 109.97 ± 57.64 | 99.36 ± 43.12 | 92.01 ± 16.50 | n.s. |
| IL-1 | 103.64 ± 69.86 | 86.05 ± 35.07 | 104.25 ± 32.82 | 132.43 ± 65.78 | n.s. |
| MMP-9 | 0.64 ± 0.35 | 1.67 ± 1.10 | 1.80 ± 1.05 | 2.03 ± 2.68 | n.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banyś, K.; Jelińska, M.; Wrzosek, M.; Skrajnowska, D.; Wrzesień, R.; Bielecki, W.; Bobrowska-Korczak, B. Inflammation Factors and Genistein Supplementation in Cancer—Preliminary Research. Curr. Issues Mol. Biol. 2024, 46, 2166-2180. https://doi.org/10.3390/cimb46030140
Banyś K, Jelińska M, Wrzosek M, Skrajnowska D, Wrzesień R, Bielecki W, Bobrowska-Korczak B. Inflammation Factors and Genistein Supplementation in Cancer—Preliminary Research. Current Issues in Molecular Biology. 2024; 46(3):2166-2180. https://doi.org/10.3390/cimb46030140
Chicago/Turabian StyleBanyś, Karolina, Małgorzata Jelińska, Małgorzata Wrzosek, Dorota Skrajnowska, Robert Wrzesień, Wojciech Bielecki, and Barbara Bobrowska-Korczak. 2024. "Inflammation Factors and Genistein Supplementation in Cancer—Preliminary Research" Current Issues in Molecular Biology 46, no. 3: 2166-2180. https://doi.org/10.3390/cimb46030140
APA StyleBanyś, K., Jelińska, M., Wrzosek, M., Skrajnowska, D., Wrzesień, R., Bielecki, W., & Bobrowska-Korczak, B. (2024). Inflammation Factors and Genistein Supplementation in Cancer—Preliminary Research. Current Issues in Molecular Biology, 46(3), 2166-2180. https://doi.org/10.3390/cimb46030140