_Kim.png)
The Promising Effect of Ascorbic Acid and Paracetamol as Anti-Biofilm and Anti-Virulence Agents against Resistant Escherichia coli
 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tested Isolates
2.2. Antibiotic Susceptibility Testing
2.3. Minimal Inhibitory Concentration (MIC) Assessment of Ascorbic Acid and Paracetamol
2.4. Assessment of the Ascorbic Acid and Paracetamol Effects on Bacterial Growth
2.5. Virulence Factor Inhibition Assay
2.5.1. Biofilm Inhibition Assay
2.5.2. Proteases Inhibition Assay
2.5.3. Swimming Motility Inhibition Assay
2.5.4. Serum Resistance Inhibition Assay
2.5.5. The Quantification of Gene Expression Levels of Virulence Factors Encoding Genes
2.5.6. In Vivo Mice Survival Test
2.6. Statistical Analysis
3. Results
3.1. Identification, Antibiotic Susceptibility, and Resistance Profile of the Isolates
3.2. MIC Values and the Effect of the Sub-MICs of Ascorbic Acid and Paracetamol on Bacterial Growth
3.3. The Influence of Ascorbic Acid and Paracetamol on E. coli Virulence Factors
3.3.1. Biofilm Inhibition Assay
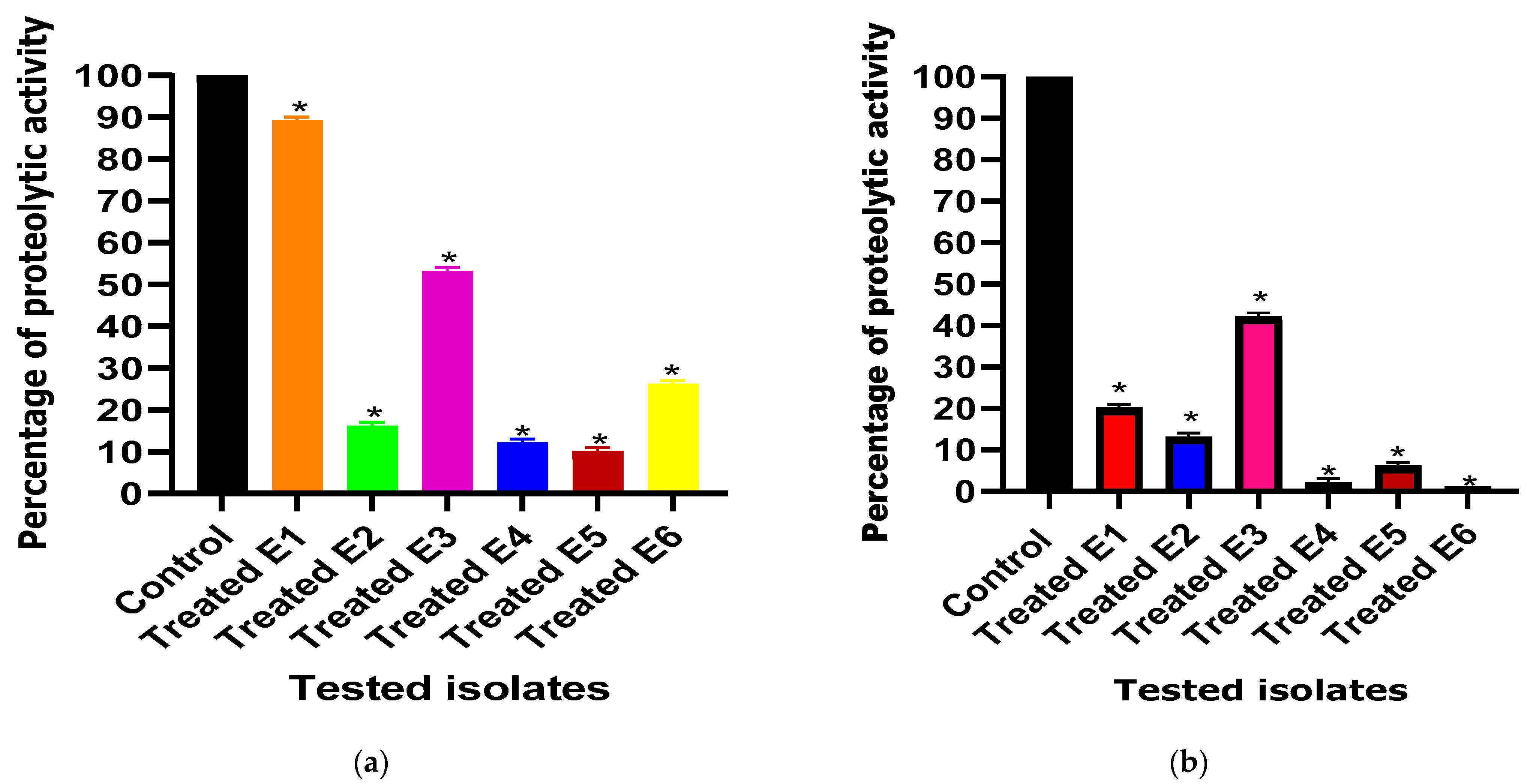
3.3.2. Total Protease Inhibition Assessment
3.3.3. Swimming Motility Inhibition Assay
3.3.4. Serum Resistance Inhibition Assay
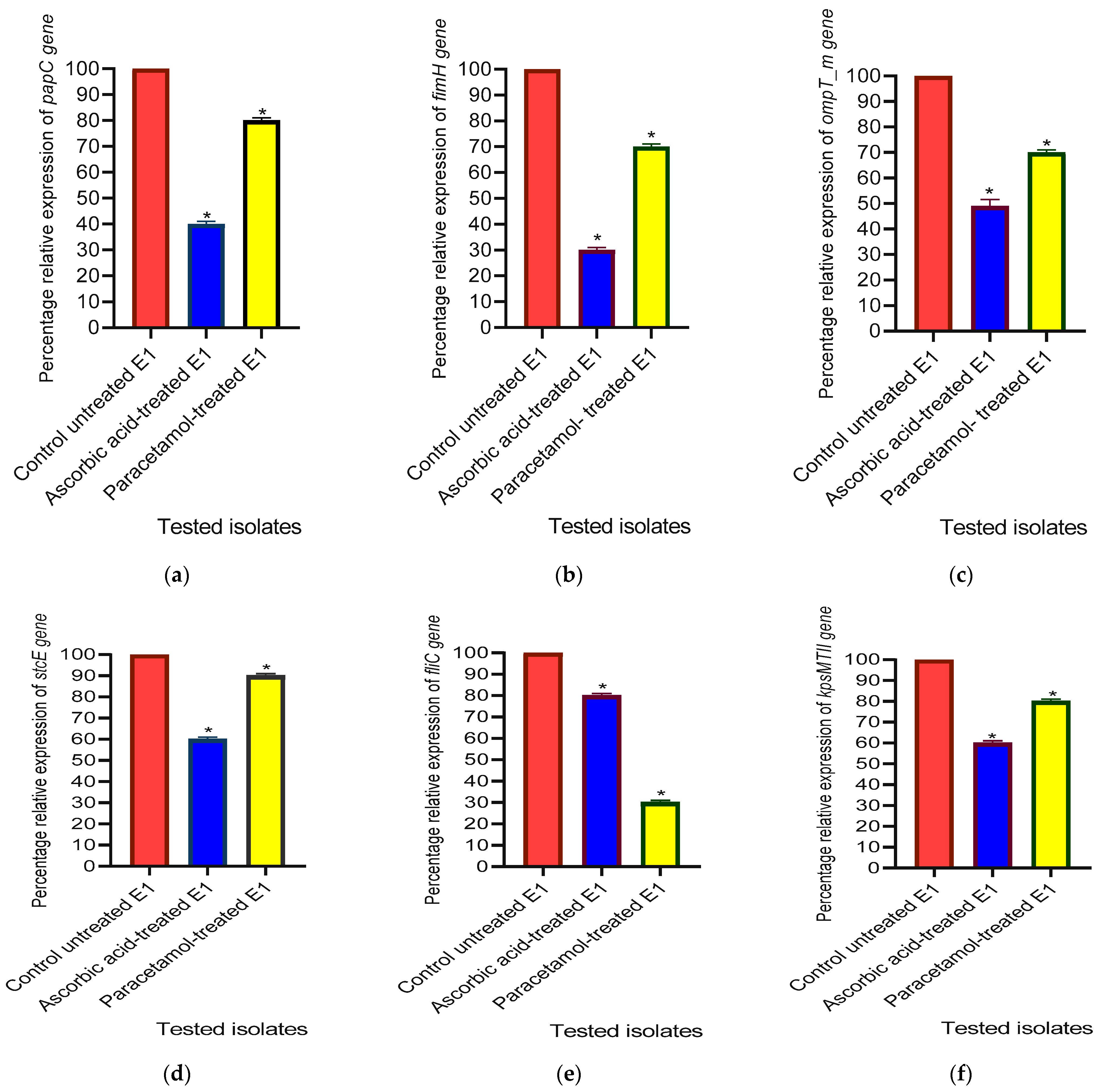
3.3.5. The Relative Gene Expression Levels of Virulence Factor-Encoding Genes in E. coli
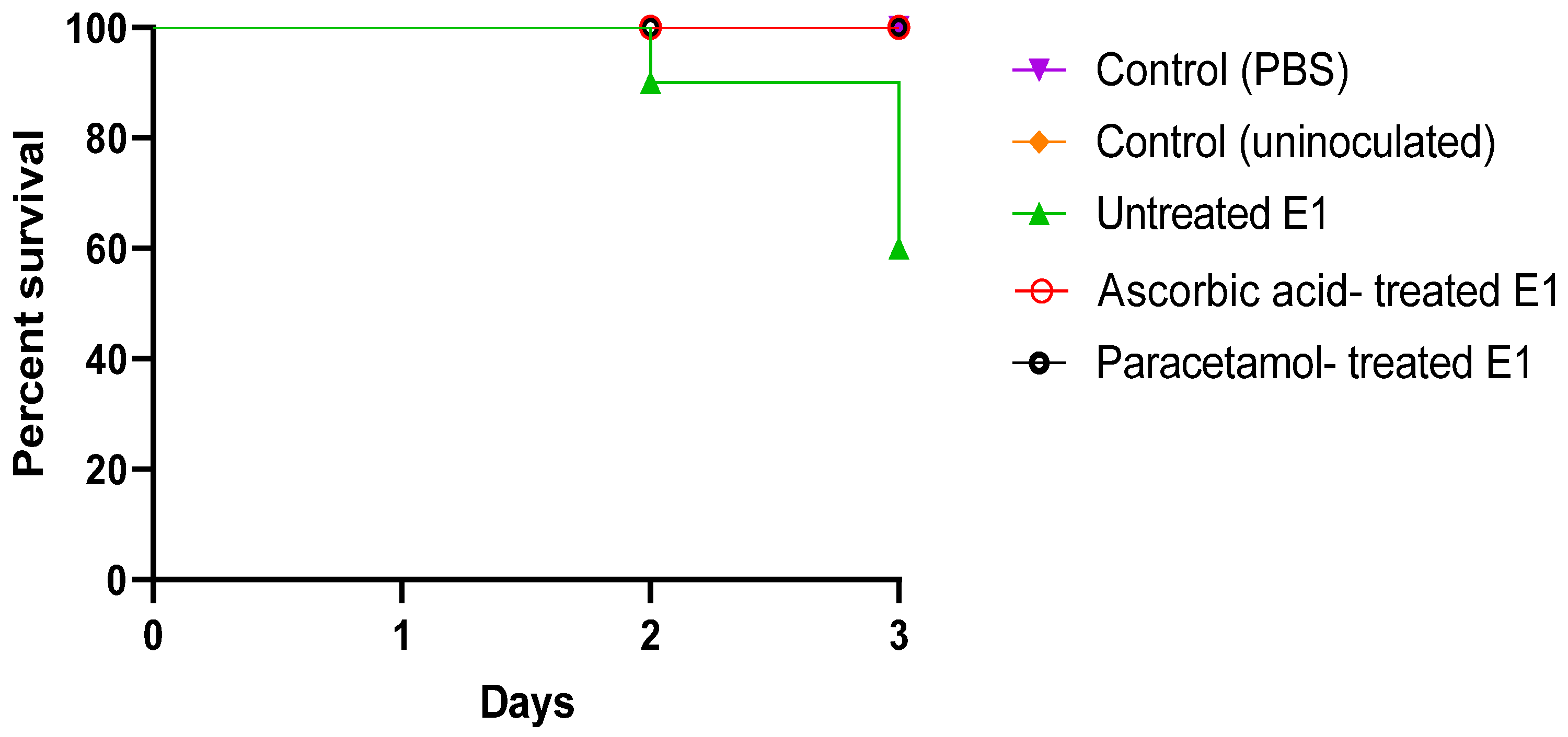
3.3.6. In Vivo Mice Survival Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pakbin, B.; Brück, W.M.; Rossen, J.W.A. Virulence Factors of Enteric Pathogenic Escherichia coli: A Review. Int. J. Mol. Sci. 2021, 22, 9922. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhu, M.; Fu, X.; Cai, J.; Chen, S.; Lin, Y.; Jiang, N.; Chen, S.; Lin, Z. Escherichia coli Causing Neonatal Meningitis During 2001–2020: A Study in Eastern China. Int. J. Gen. Med. 2021, 14, 3007–3016. [Google Scholar] [CrossRef] [PubMed]
- Terlizzi, M.E.; Gribaudo, G.; Maffei, M.E. UroPathogenic Escherichia coli (UPEC) Infections: Virulence Factors, Bladder Responses, Antibiotic, and Non-Antibiotic Antimicrobial Strategies. Front. Microbiol. 2017, 8, 1566. [Google Scholar] [CrossRef] [PubMed]
- Czajkowski, K.; Broś-Konopielko, M.; Teliga-Czajkowska, J. Urinary Tract Infection in Women. Prz. Menopauzalny 2021, 20, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, J.; Rout, P.; Sedhai, Y.R. Hemolytic Uremic Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Foschi, D.; Yakushkina, A.; Cammarata, F.; Lamperti, G.; Colombo, F.; Rimoldi, S.; Antinori, S.; Sampietro, G.M. Surgical Site Infections Caused by Multi-Drug Resistant Organisms: A Case–Control Study in General Surgery. Updates Surg. 2022, 74, 1763–1771. [Google Scholar] [CrossRef] [PubMed]
- Eltabey, S.M.; Ibrahim, A.H.; Zaky, M.M.; Saleh, M.M. Antimicrobial Susceptibility and Resistance Profile of Escherichia coli Isolates from Patients at Suez Canal University Specialized Hospital. Alfarama J. Basic. Appl. Sci. 2024, 5, 62–74. [Google Scholar] [CrossRef]
- de Kraker, M.E.A.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year Due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef] [PubMed]
- Rozwadowski, M.; Gawel, D. Molecular Factors and Mechanisms Driving Multidrug Resistance in Uropathogenic Escherichia coli—An Update. Genes 2022, 13, 1397. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; de Carvalho, L.P.S. Metabolomic Profiling Reveals Bacterial Metabolic Adaptation Strategies and New Metabolites. Curr. Opin. Chem. Biol. 2023, 74, 102287. [Google Scholar] [CrossRef]
- El-baz, R.; Said, H.S.; Abdelmegeed, E.S.; Barwa, R. Characterization of Virulence Determinants and Phylogenetic Background of Multiple and Extensively Drug Resistant Escherichia coli Isolated from Different Clinical Sources in Egypt. Appl. Microbiol. Biotechnol. 2022, 106, 1279–1298. [Google Scholar] [CrossRef]
- Grys, T.E.; Walters, L.L.; Welch, R.A. Characterization of the StcE Protease Activity of Escherichia coli O157:H7. J. Bacteriol. 2006, 188, 4646–4653. [Google Scholar] [CrossRef]
- Hidalgo, G.; Chan, M.; Tufenkji, N. Inhibition of Escherichia coli CFT073 fliC Expression and Motility by Cranberry Materials. Appl. Environ. Microbiol. 2011, 77, 6852–6857. [Google Scholar] [CrossRef]
- Mahmoud, A.T.; Ibrahem, R.A.; Salim, M.T.; Gabr, A.; Halby, H.M. Prevalence of Some Virulence Factors and Genotyping of Hospital-Acquired Uropathogenic Escherichia coli Isolates Recovered from Cancer Patients. J. Glob. Antimicrob. Resist. 2020, 23, 211–216. [Google Scholar] [CrossRef]
- Poole, N.M.; Green, S.I.; Rajan, A.; Vela, L.E.; Zeng, X.-L.; Estes, M.K.; Maresso, A.W. Role for FimH in Extraintestinal Pathogenic Escherichia coli Invasion and Translocation through the Intestinal Epithelium. Infect. Immun. 2017, 85, e00581-71. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte-Rutten, L.; Kramer, R.A.; Kroon, J.; Dekker, N.; Egmond, M.R.; Gros, P. Crystal Structure of the Outer Membrane Protease OmpT from Escherichia coli Suggests a Novel Catalytic Site. EMBO J. 2001, 20, 5033–5039. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.M.; Yousef, N.; Shafik, S.M.; Abbas, H.A. Attenuating the Virulence of the Resistant Superbug Staphylococcus Aureus Bacteria Isolated from Neonatal Sepsis by Ascorbic Acid, Dexamethasone, and Sodium Bicarbonate. BMC Microbiol. 2022, 22, 268. [Google Scholar] [CrossRef] [PubMed]
- Eltabey, S.M.; Ibrahim, A.H.; Zaky, M.M.; Saleh, M.M. Attenuation of Virulence Factors and Antibiotic Resistance in Escherichia coli Bacteria by Using Some Available Drugs; Faculty of Science, Port Said University: Port Said, Egypt, 2024. [Google Scholar]
- Elfaky, M.A.; Abdel-Hamid, M.I.; Khalifa, E.; Alshareef, W.A.; Mosbah, R.A.; Elazab, S.T.; Ghoneim, M.M.; Al-Sanea, M.M.; Bendary, M.M. Innovative Next-Generation Therapies in Combating Multi-Drug-Resistant and Multi-Virulent Escherichia coli Isolates: Insights from In Vitro, In Vivo, and Molecular Docking Studies. Appl. Microbiol. Biotechnol. 2022, 106, 1691–1703. [Google Scholar] [CrossRef] [PubMed]
- Totsika, M. Benefits and Challenges of Antivirulence Antimicrobials at the Dawn of the Post-Antibiotic Era. Drug Deliv. Lett. 2016, 6, 30–37. [Google Scholar] [CrossRef]
- Eltabey, S.M.; Ibrahim, A.H.; Zaky, M.M.; Saleh, M.M. Mitigation of Virulence in Resistant Escherichia coli by Diclofenac Sodium, Phenotypic and Genotypic Study. Delta Univ. Sci. J. 2024, 7, 150–162. [Google Scholar] [CrossRef]
- Mosallam, F.M.; Abbas, H.A.; Shaker, G.H.; Gomaa, S.E. Alleviating the Virulence of Pseudomonas Aeruginosa and Staphylococcus Aureus by Ascorbic Acid Nanoemulsion. Res. Microbiol. 2023, 174, 104084. [Google Scholar] [CrossRef]
- Rifai, A.; El-Aziz, A.; Kenawy, H. Possible Antivirulent Activity of Some Agents against Clinical Isolates of Pseudomonas Aeruginosa. Egypt J. Med. Microbiol. 2021, 30, 1–8. [Google Scholar] [CrossRef]
- Sihotang, T.S.U.; Widodo, A.D.W.; Arfijanto, M.V. Comparison of Doses of Paracetamol or Ibuprofen to Inhibit the Formation of Biofilms Pseudomonas Aeruginosa Bacteria. Int. J. Health Sci. 2022, 6, 361–367. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement M100-S25; Clinical and Laboratory Standards Institute: Malvern, PA, USA, 2015; ISBN 1-56238-990-4. [Google Scholar]
- Stepanović, S.; Vuković, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Cirković, I.; Ruzicka, F. Quantification of Biofilm in Microtiter Plates: Overview of Testing Conditions and Practical Recommendations for Assessment of Biofilm Production by Staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- El-Mowafy, S.A.; Abd El Galil, K.H.; El-Messery, S.M.; Shaaban, M.I. Aspirin Is an Efficient Inhibitor of Quorum Sensing, Virulence and Toxins in Pseudomonas Aeruginosa. Microb. Pathog. 2014, 74, 25–32. [Google Scholar] [CrossRef] [PubMed]
- AL-Ghonaiem, M.I.; Ibrahim, A.S.S.; Al-Salamah, A.A. Swimming Motility in Agrobacterium Tumefaciens Is Controlled by Quorum Sensing and Inhibited by Garlic Bulb Extract. Res. J. Microbiol. 2010, 4, 345–354. [Google Scholar] [CrossRef]
- Vaish, R.; Pradeep, M.; Setty, C.R.; Kandi, V. Evaluation of Virulence Factors and Antibiotic Sensitivity Pattern of Escherichia coli Isolated from Extraintestinal Infections. Cureus 2016, 8, e604. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Magray, M.S.U.D.; Kumar, A.; Rawat, A.K.; Srivastava, S. Identification of Escherichia coli through Analysis of 16S rRNA and 16S-23S rRNA Internal Transcribed Spacer Region Sequences. Bioinformation 2011, 6, 370–371. [Google Scholar] [CrossRef] [PubMed]
- Schiebel, J. Genotypic and Phenotypic Characteristics Associated with Biofilm Formation by Human Clinical Escherichia coli Isolates of Different Pathotypes. Appl. Environ. Microbiol. 2017, 83, e01660-17. [Google Scholar] [CrossRef]
- Desloges, I.; Taylor, J.A.; Leclerc, J.-M.; Brannon, J.R.; Portt, A.; Spencer, J.D.; Dewar, K.; Marczynski, G.T.; Manges, A.; Gruenheid, S.; et al. Identification and Characterization of OmpT-like Proteases in Uropathogenic Escherichia coli Clinical Isolates. Microbiologyopen 2019, 8, e915. [Google Scholar] [CrossRef]
- Kim, H.-S.; Lee, S.-H.; Byun, Y.; Park, H.-D. 6-Gingerol Reduces Pseudomonas Aeruginosa Biofilm Formation and Virulence via Quorum Sensing Inhibition. Sci. Rep. 2015, 5, 8656. [Google Scholar] [CrossRef] [PubMed]
- Avdić, M.; Ibrišimović, M.; Mehmedinović, N.; Hukić, M. A Novel Spectrophotometric Assay for the Determination of Biofilm Forming Capacity of Causative Agents of Urinary Tract Infections. IJERT 2017, 6, 1225–1230. [Google Scholar]
- Sharma, G.; Sharma, S.; Sharma, P.; Chandola, D.; Dang, S.; Gupta, S.; Gabrani, R. Escherichia coli Biofilm: Development and Therapeutic Strategies. J. Appl. Microbiol. 2016, 121, 309–319. [Google Scholar] [CrossRef] [PubMed]
- ElBaradei, A.; Yakout, M.A. Stenotrophomonas Maltophilia: Genotypic Characterization of Virulence Genes and The Effect of Ascorbic Acid on Biofilm Formation. Curr. Microbiol. 2022, 79, 180. [Google Scholar] [CrossRef]
- Hamed, S.; Emara, M. Antibacterial and Antivirulence Activities of Acetate, Zinc Oxide Nanoparticles, and Vitamin C Against E. Coli O157:H7 and P. Aeruginosa. Curr. Microbiol. 2023, 80, 57. [Google Scholar] [CrossRef]
- El-Mowafy, S.A.; Shaaban, M.I.; Abd El Galil, K.H. Sodium Ascorbate as a Quorum Sensing Inhibitor of Pseudomonas Aeruginosa. J. Appl. Microbiol. 2014, 117, 1388–1399. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Dong, N.; Chen, K.; Yang, X.; Zeng, P.; Hou, C.; Chi Chan, E.W.; Yao, X.; Chen, S. Bactericidal, Anti-Biofilm, and Anti-Virulence Activity of Vitamin C against Carbapenem-Resistant Hypervirulent Klebsiella Pneumoniae. iScience 2022, 25, 103894. [Google Scholar] [CrossRef]
- Seleem, N.M.; Atallah, H.; Abd El Latif, H.K.; Shaldam, M.A.; El-Ganiny, A.M. Could the Analgesic Drugs, Paracetamol and Indomethacin, Function as Quorum Sensing Inhibitors? Microb. Pathog. 2021, 158, 105097. [Google Scholar] [CrossRef]
- Abidi, S.H.; Ahmed, K.; Kazmi, S.U. The Antibiofilm Activity of Acetylsalicylic Acid, Mefenamic Acid, Acetaminophen against Biofilms Formed by P. Aeruginosa and S. Epidermidis. J. Pak. Med. Assoc. 2019, 69, 1493–1495. [Google Scholar] [CrossRef] [PubMed]
- Almulla, A.; Al-Khafaji, Z. In Vitro Applications of In Silico Designed Antibiofilm Agents for Sta Phylococcus Epidermidis. Am. J. PharmTech Res. 2015, 5, 271–282. [Google Scholar]
- Ismail, M.M.; Hassan, M.; Moawad, S.S.; Okba, M.M.; Ashour, R.M.; Fayek, N.M.; Saber, F.R. Exploring the Antivirulence Activity of Pulverulentone A, a Phloroglucinol-Derivative from Callistemon Citrinus Leaf Extract, against Multi-Drug Resistant Pseudomonas Aeruginosa. Antibiotics 2021, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Abdelraheem, W.M.; Refaie, M.M.M.; Yousef, R.K.M.; Abd El Fatah, A.S.; Mousa, Y.M.; Rashwan, R. Assessment of Antibacterial and Anti-Biofilm Effects of Vitamin C Against Pseudomonas Aeruginosa Clinical Isolates. Front. Microbiol. 2022, 13, 847449. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, İ.; Eraç, Y.; Kirmizibayrak, P.; Ermertcan, Ş. Nonsteroidal Antiinflammatory Drugs Alter Antibiotic Susceptibility and Expression Ofvirulence-Related Genes and Protein A of Staphylococcus Aureus. Turk. J. Med. Sci. 2021, 51, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Aldawsari, M.F.; Khafagy, E.-S.; Saqr, A.A.; Alalaiwe, A.; Abbas, H.A.; Shaldam, M.A.; Hegazy, W.A.H.; Goda, R.M. Tackling Virulence of Pseudomonas Aeruginosa by the Natural Furanone Sotolon. Antibiotics 2021, 10, 871. [Google Scholar] [CrossRef]
- Ahsan, M.R.; Sultan, M.Z.; Baki, M.A.; Rahman, M.A.; Hossain, M.A.; Hossain, M.A.; Amran, M.S. The Study of in Vitro and in Vivo Effects of Concurrent Administration of Paracetamol and Zinc on the Antibacterial Activity of Ciprofloxacin. Dhaka Univ. J. Pharm. Sci. 2011, 10, 137–142. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers | References |
|---|---|---|
| 16SrRNA | F */AGT TTG ATC MTG GCT CAG | [31] |
| R */GGA CTA CHA GGG TAT CTA AT | ||
| papC | F/TGA TAT CAC GCA GTC AGT AGC | [32] |
| R/CCG GCC ATA TTC ACA TAAC | ||
| fimH | F/TGC AGA ACG GAT AAG CCG TGG | [32] |
| R/GCA GTC ACC TGC CCT CCG GTA | ||
| ompT_m | F/TTT GAT GCC CCA GAT ATC TAT CGG | [33] |
| R/GGC TTT CCT GAT ATC CGG CCA TG | ||
| stcE | F/AAG GGC CCC TCT GAG GTG TCT GTTAAA CCC GTG G | [12] |
| R/AAA AA TGG CCA CGA AGT GGCCGC ACC GTC TCA GG | ||
| fliC | F/ACA GCC TCT CGC TGA TCA CTC AAA | [13] |
| R/GCG CTG TTA ATA CGC AAG CCA GAA | ||
| kpsMTII | F/GCG CAT TTG CTG ATA CTG TTG | [11] |
| R/CAT CCA GAC GAT AAG CAT GAGCA |
| Antibiotics | LEV 1 5 µg | MRP 1 10 µg | CTX 1 30 µg | SXT 1 25 µg | AK 1 30 µg | PRL 1 100 µg | DO 1 30 µg | AMC 1 30 µg | |
|---|---|---|---|---|---|---|---|---|---|
| Isolates | |||||||||
| E1 * | R 2 | S 2 | R | R | S | R | R | R | |
| E2 | R | S | R | R | S | R | S | R | |
| E3 | S | S | R | R | S | R | S | R | |
| E4 | S | S | R | R | S | R | R | R | |
| E5 | R | S | R | R | S | R | S | R | |
| E6 | R | S | R | R | S | R | S | R | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltabey, S.M.; Ibrahim, A.H.; Zaky, M.M.; Ibrahim, A.E.; Alrashdi, Y.B.A.; El Deeb, S.; Saleh, M.M. The Promising Effect of Ascorbic Acid and Paracetamol as Anti-Biofilm and Anti-Virulence Agents against Resistant Escherichia coli. Curr. Issues Mol. Biol. 2024, 46, 6805-6819. https://doi.org/10.3390/cimb46070406
Eltabey SM, Ibrahim AH, Zaky MM, Ibrahim AE, Alrashdi YBA, El Deeb S, Saleh MM. The Promising Effect of Ascorbic Acid and Paracetamol as Anti-Biofilm and Anti-Virulence Agents against Resistant Escherichia coli. Current Issues in Molecular Biology. 2024; 46(7):6805-6819. https://doi.org/10.3390/cimb46070406
Chicago/Turabian StyleEltabey, Sara M., Ali H. Ibrahim, Mahmoud M. Zaky, Adel Ehab Ibrahim, Yahya Bin Abdullah Alrashdi, Sami El Deeb, and Moustafa M. Saleh. 2024. "The Promising Effect of Ascorbic Acid and Paracetamol as Anti-Biofilm and Anti-Virulence Agents against Resistant Escherichia coli" Current Issues in Molecular Biology 46, no. 7: 6805-6819. https://doi.org/10.3390/cimb46070406