
Analysis of Intestinal Bacterial Microbiota in Individuals with and without Chronic Low Back Pain

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Stages
2.3. Statistical Analysis
2.4. Ethical Aspects
3. Results
3.1. Sample Description
3.2. Microbial Composition Analysis
3.2.1. Variables SpRIC and CD
- Species richness (SpRIC), with PG showing a higher number of taxa compared to CG (p = 0.030);
- Relative abundance of the Clostridium difficile species (a pathogenic bacterium related to inflammatory conditions of the large intestine), which is higher in PG (p = 0.011).
3.2.2. Intestinal Bacterial Microbiota Sequencing of the PG and CG Samples
3.2.3. Significant Difference between CG and PG
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2021 Low Back Pain Collaborators. Global, regional, and national burden of low back pain, 1990–2020, its attributable risk factors, and projections to 2050: A systematic analysis of the Global Burden of Disease Study 2021. Lancet Rheumatol. 2023, 5, e316–e329. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Borenstein, D.G.; Balagué, F. Low Back Pain in Adolescent and Geriatric Populations. Rheum. Dis. Clin. N. Am. 2021, 47, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Chenot, J.F.; Greitemann, B.; Kladny, B.; Petzke, F.; Pfingsten, M.; Schorr, S.G. Non-Specific Low Back Pain. Dtsch. Aerzteblatt Int. 2017, 114, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheng, H.; Wang, T.; Zhang, K.; Zhang, Y.; Kang, X. Oxidative stress in intervertebral disc degeneration: Molecular mechanisms, pathogenesis and treatment. Cell Prolif. 2023, 56, e13448. [Google Scholar] [CrossRef]
- Francisco, V.; Pino, J.; González-Gay, M.Á.; Lago, F.; Karppinen, J.; Tervonen, O.; Mobasheri, A.; Gualillo, O. A new immunometabolic perspective of intervertebral disc degeneration. Nat. Rev. Rheumatol. 2022, 18, 47–60. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef]
- Rizzetto, L.; Fava, F.; Tuohy, K.M.; Selmi, C. Connecting the immune system, systemic chronic inflammation and the gut microbiome: The role of sex. J. Autoimmun. 2018, 92, 12–34. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell Infect. Microbiol. 2021, 11, 625913. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Cai, Y.; Wang, W.; Deng, J.; Zhao, L.; Han, Z.; Wan, L. The Effect of Gut Microbiota on the Progression of Intervertebral Disc Degeneration. Orthop. Surg. 2023, 15, 858–867. [Google Scholar] [CrossRef]
- Li, W.; Lai, K.; Chopra, N.; Zheng, Z.; Das, A.; Diwan, A.D. Gut-disc axis: A cause of intervertebral disc degeneration and low back pain? Eur. Spine J. 2022, 31, 917–925. [Google Scholar] [CrossRef]
- Rajasekaran, S.; Soundararajan, D.C.R.; Tangavel, C.; Muthurajan, R.; Sri Vijay Anand, K.S.; Matchado, M.S.; Nayagam, S.M.; Shetty, A.P.; Kanna, R.M.; Dharmalingam, K. Human intervertebral discs harbour a unique microbiome and dysbiosis determines health and disease. Eur. Spine J. 2020, 29, 1621–1640. [Google Scholar] [CrossRef] [PubMed]
- Aroniadis, O.C.; Grinspan, A.M. The Gut Microbiome: A Primer for the Clinician. Am. J. Gastroenterol. 2024, 119, S2–S6. [Google Scholar] [CrossRef] [PubMed]
- Blake, M.R.; Raker, J.M.; Whelan, K. Validity and reliability of the Bristol Stool Form Scale in healthy adults and patients with diarrhoea-predominant irritable bowel syndrome. Aliment. Pharmacol. Ther. 2016, 44, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Chiarello, M.; McCauley, M.; Villéger, S.; Jackson, C.R. Ranking the biases: The choice of OTUs vs. ASVs in 16S rRNA amplicon data analysis has stronger effects on diversity measures than rarefaction and OTU identity threshold. PLoS ONE 2022, 17, e0264443. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Vaiserman, A.; Romanenko, M.; Piven, L.; Moseiko, V.; Lushchak, O.; Kryzhanovska, N.; Guryanov, V.; Koliada, A. Differences in the gut Firmicutes to Bacteroidetes ratio across age groups in healthy Ukrainian population. BMC Microbiol. 2020, 20, 221. [Google Scholar] [CrossRef] [PubMed]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska-Pietruszka, Z.; Figlerowicz, M.; Mazur-Melewska, K. The History of the Intestinal Microbiota and the Gut-Brain Axis. Pathogens 2022, 11, 1540. [Google Scholar] [CrossRef] [PubMed]
- Trastoy, B.; Naegeli, A.; Anso, I.; Sjögren, J.; Guerin, M.E. Structural basis of mammalian mucin processing by the human gut O-glycopeptidase OgpA from Akkermansia muciniphila. Nat. Commun. 2020, 11, 4844. [Google Scholar] [CrossRef] [PubMed]
- Martín, R.; Rios-Covian, D.; Huillet, E.; Auger, S.; Khazaal, S.; Bermúdez-Humarán, L.G.; Sokol, H.; Chatel, J.M.; Langella, P. Faecalibacterium: A bacterial genus with promising human health applications. FEMS Microbiol. Rev. 2023, 47, fuad039. [Google Scholar] [CrossRef]
- Feng, Y.; Duan, Y.; Xu, Z.; Lyu, N.; Liu, F.; Liang, S.; Zhu, B. An examination of data from the American Gut Project reveals that the dominance of the genus Bifidobacterium is associated with the diversity and robustness of the gut microbiota. Microbiologyopen 2019, 8, e939. [Google Scholar] [CrossRef]
- Valguarnera, E.; Wardenburg, J.B. Good Gone Bad: One Toxin Away From Disease for Bacteroides fragilis. J. Mol. Biol. 2020, 432, 765–785. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Cho, Y.; Seo, M.R.; Bae, M.H.; Kim, B.; Rho, M.; Pai, H. Quantitative characterization of Clostridioides difficile population in the gut microbiome of patients with C. difficile infection and their association with clinical factors. Sci. Rep. 2020, 10, 17608. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Shin, J.; Guevarra, R.; Lee, J.H.; Kim, D.W.; Seol, K.H.; Lee, J.H.; Kim, H.B.; Isaacson, R. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; Kong, X.B.; Liu, Z.; Jing, B.Y.; Gao, X. Automatic peak selection by a Benjamini-Hochberg-based algorithm. PLoS ONE 2013, 8, e53112. [Google Scholar] [CrossRef] [PubMed]
- Bathini, P.; Foucras, S.; Dupanloup, I.; Imeri, H.; Perna, A.; Berruex, J.L.; Doucey, M.A.; Annoni, J.M.; Auber Alberi, L. Classifying dementia progression using microbial profiling of saliva. Alzheimers Dement. 2020, 12, e12000. [Google Scholar] [CrossRef] [PubMed]
- Plugge, C.M.; Balk, M.; Zoetendal, E.G.; Stams, A.J.M. Gelria glutamica gen. nov., sp. nov., a thermophilic, obligately syntrophic, glutamate-degrading anaerobe. Int. J. Syst. Evol. Microbiol. 2002, 52 Pt 2, 401–407. [Google Scholar] [CrossRef]
- Wang, Z.; Peters, B.A.; Bryant, M.; Hanna, D.B.; Schwartz, T.; Wang, T.; Sollecito, C.C.; Usyk, M.; Grassi, E.; Wiek, F.; et al. Gut microbiota, circulating inflammatory markers and metabolites, and carotid artery atherosclerosis in HIV infection. Microbiome 2023, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Ordinola-Zapata, R.; Costalonga, M.; Dietz, M.; Lima, B.P.; Staley, C. The root canal microbiome diversity and function. A whole-metagenome shotgun analysis. Int. Endod. J. 2023, 57, 872–884. [Google Scholar] [CrossRef]
- Hawkes, C.G.; Hinson, A.N.; Vashishta, A.; Read, C.B.; Carlyon, J.A.; Lamont, R.J.; Uriarte, S.M.; Miller, D.P. Selenomonas sputigena Interactions with Gingival Epithelial Cells That Promote Inflammation. Infect. Immun. 2023, 91, e0031922. [Google Scholar] [CrossRef]
- Sasaki, M.; Shimoyama, Y.; Kodama, Y.; Ishikawa, T. Abiotrophia defectiva DnaK Promotes Fibronectin-Mediated Adherence to HUVECs and Induces a Proinflammatory Response. Int. J. Mol. Sci. 2021, 22, 8528. [Google Scholar] [CrossRef]
- Diao, J.; Yuan, C.; Tong, P.; Ma, Z.; Sun, X.; Zheng, S. Potential Roles of the Free Salivary Microbiome Dysbiosis in Periodontal Diseases. Front. Cell Infect. Microbiol. 2021, 11, 711282. [Google Scholar] [CrossRef]
- Fortes, T.O.; Alviano, D.S.; Tupinambá, G.; Padrón, T.S.; Antoniolli, A.R.; Alviano, C.S.; Seldin, L. Production of an antimicrobial substance against Cryptococcus neoformans by Paenibacillus brasilensis Sa3 isolated from the rhizosphere of Kalanchoe brasiliensis. Microbiol. Res. 2008, 163, 200–207. [Google Scholar] [CrossRef]
- Mosadegh, S.; Abtahi, H.; Amani, J.; Karizi, S.Z.; Salmanian, A.H. Protective immunization against Enterohemorrhagic Escherichia coli and Shigella dysenteriae Type 1 by chitosan nanoparticle loaded with recombinant chimeric antigens comprising EIT and STX1B-IpaD. Microb. Pathog. 2023, 184, 106344. [Google Scholar] [CrossRef]
- Ricotta, C.; Pavoine, S. A new parametric measure of functional dissimilarity: Bridging the gap between the Bray-Curtis dissimilarity and the Euclidean distance. Ecol. Model. 2022, 466, 109880. [Google Scholar] [CrossRef]
- Jarvik, J.G.; Hollingworth, W.; Heagerty, P.J.; Haynor, D.R.; Boyko, E.J.; Deyo, R.A. Three-year incidence of low back pain in an initially asymptomatic cohort: Clinical and imaging risk factors. Spine 2005, 30, 1541–1548, discussion 1549. [Google Scholar] [CrossRef]
- Brinjikji, W.; Diehn, F.E.; Jarvik, J.G.; Carr, C.M.; Kallmes, D.F.; Murad, M.H.; Luetmer, P.H. MRI Findings of Disc Degeneration are More Prevalent in Adults with Low Back Pain than in Asymptomatic Controls: A Systematic Review and Meta-Analysis. AJNR Am. J. Neuroradiol. 2015, 36, 2394–2399. [Google Scholar] [CrossRef]
- Ahmed, M.; Modic, M.T. Neck and low back pain: Neuroimaging. Neurol. Clin. 2007, 25, 439–471. [Google Scholar] [CrossRef]
- Brinjikji, W.; Luetmer, P.H.; Comstock, B.; Bresnahan, B.W.; Chen, L.E.; Deyo, R.A.; Halabi, S.; Turner, J.A.; Avins, A.L.; James, K.; et al. Systematic literature review of imaging features of spinal degeneration in asymptomatic populations. AJNR Am. J. Neuroradiol. 2015, 36, 811–816. [Google Scholar] [CrossRef]
- Carragee, E.; Alamin, T.; Cheng, I.; Franklin, T.; Haak, E.v.D.; Hurwitz, E. Are first-time episodes of serious LBP associated with new MRI findings? Spine J. 2006, 6, 624–635. [Google Scholar] [CrossRef]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.I.; McDonald, D.; et al. Best practices for analysing microbiomes. Nat. Rev. Microbiol. 2018, 16, 410–422. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | Reference |
|---|---|
| Phylum | |
| Abundance of Firmicutes and Bacteroidetes in relation to other phyla (PhylABD) | 85–95% [15] |
| Firmicutes and Bacteroidetes Ratio (FiBaRAT) | 0.7–1.0 [16] |
| Gender | |
| Gender diversity (GenDIV) | Greater than 7.0 [17] |
| Species | |
| Species richness (SpRIC) | Greater than 540 [18] |
| Protective | |
| Akkermansia muciniphila (AM) | 1–5% [19] |
| Faecalibacterium prausnitzii (FP) | 5–12% [20] |
| Bifidobacterium spp. (BS) | 1–6% [21] |
| Pathogenic | |
| Bacteroides fragilis (BF) | Less than 0.5% [22] |
| Clostridium difficilis (CD) | 0 [23] |
| Variables | CG | PG | p | ||
|---|---|---|---|---|---|
| Gender: M/F: n, (%) | 11/22 (33.3/66.6) | 16/24 (40/60) | Chi-squared = 0.1181, df = 1 p = 0.7311 | ||
| Age: mean (SD) | 39.79 ± 7.73 | 38.58 ± 8.1 | Mann–Whitney = 720.5, p = 0.5055 | ||
| * Ethnicity: W/B/other, n, (%) | 19, 9, 5 (57.5/27.2/15.1) | 27, 11, 2 (67.5/27.5/5.0) | Chi-squared = 6.2703, df = 2 p = 0.0435 | ||
| * Average income (SD) | R$ 10,636.70 ± 14,660.44 | USD 1970.96 ± 2716.55 | R$ 6161.00 ± 6868.45 | USD 1141.62 ± 1272.71 | Mann–Whitney = 637.5 p = 0.2105 |
| * BMI: mean (SD) | 23.6 ± 2.5 | 25.2 ± 2.9 | Mann–Whitney = 457 p = 0.0248 | ||
| Hours of sleep at night | <6 h: 9 6–7.5 h: 22 >7: 1 | <6 h: 18 6–7.5 h: 14 >7: 5 | Chi-squared = 7.1195, df = 2 p = 0.0284 | ||
| *N Meals/day: mean (SD) | 3.54 ± 0.9 | 3.5 ± 1.0 | Chi-squared = 7.5132, df = 4 p = 0.1111 | ||
| *N Amount of fiber: R/Po: n, (%) | 21, 12 (63.6/36.4) | 13, 27 (32.5/67.5) | Chi-squared = 5.0875, df = 1 p = 0.0241 | ||
| *N Liq at mealtimes: Y/N: n, (%) | 12, 21 (36.7/63.6) | 19, 21 (47.5/52.5) | Chi-squared = 0.19136, df = 1 p = 0.6618 | ||
| *N Larger meal: M/A/E: n, (%) | 5, 23, 5 (15.1/69.7/15.1) | 6, 27, 7 (15.0/67.5/17.5) | Chi-squared = 0.16572, df = 2 p = 0.9205 | ||
| * Route of birth: V/C/nd: n, (%) | 22, 9, 2 (66.6/27.3/6.0) | 24, 12, 4 (60.0/30.0/10.0) | Chi-squared = 0.5157, df = 2 p = 0.7720 | ||
| ** Bristol stool scale: mode | 3 | 3 | *** | ||
| ** Stool frequency: means, DP | 1 (0.7) | 1 (1.0) | Chi-squared = 0.5157, df = 2 p = 0.0308 | ||
| ** D/C oscillation: Y/N: n, (%) | 4, 29 (12.1/87.9) | 27, 13 (67.5/32.5) | Chi-squared = 20.487, df = 1 p = 6.003 × 10−6 | ||
| Variables | CG | PG | p < 0.05 |
|---|---|---|---|
| SpRIC n (dp) | 869.79 (182.520) | 962.93 (196.400) | 0.030 |
| CD m-ppm (dp) | 0.001 (0.002) | 0.003 (0.003) | 0.011 |
| Diversity Index | CG m (dp) | PG m (dp) | Adjusted p |
|---|---|---|---|
| ACE | 1192.25 (219.06) | 1330.47 (252.54) | 0.0263 |
| Observed | 844.33 (165.66) | 951.28 (191.93) | 0.0272 |
| Chao1 | 1188.68 (224.93) | 1334.59 (253.37) | 0.0263 |
| Fisher | 105.31 (24.87) | 129.58 (29.48) | 0.0045 |
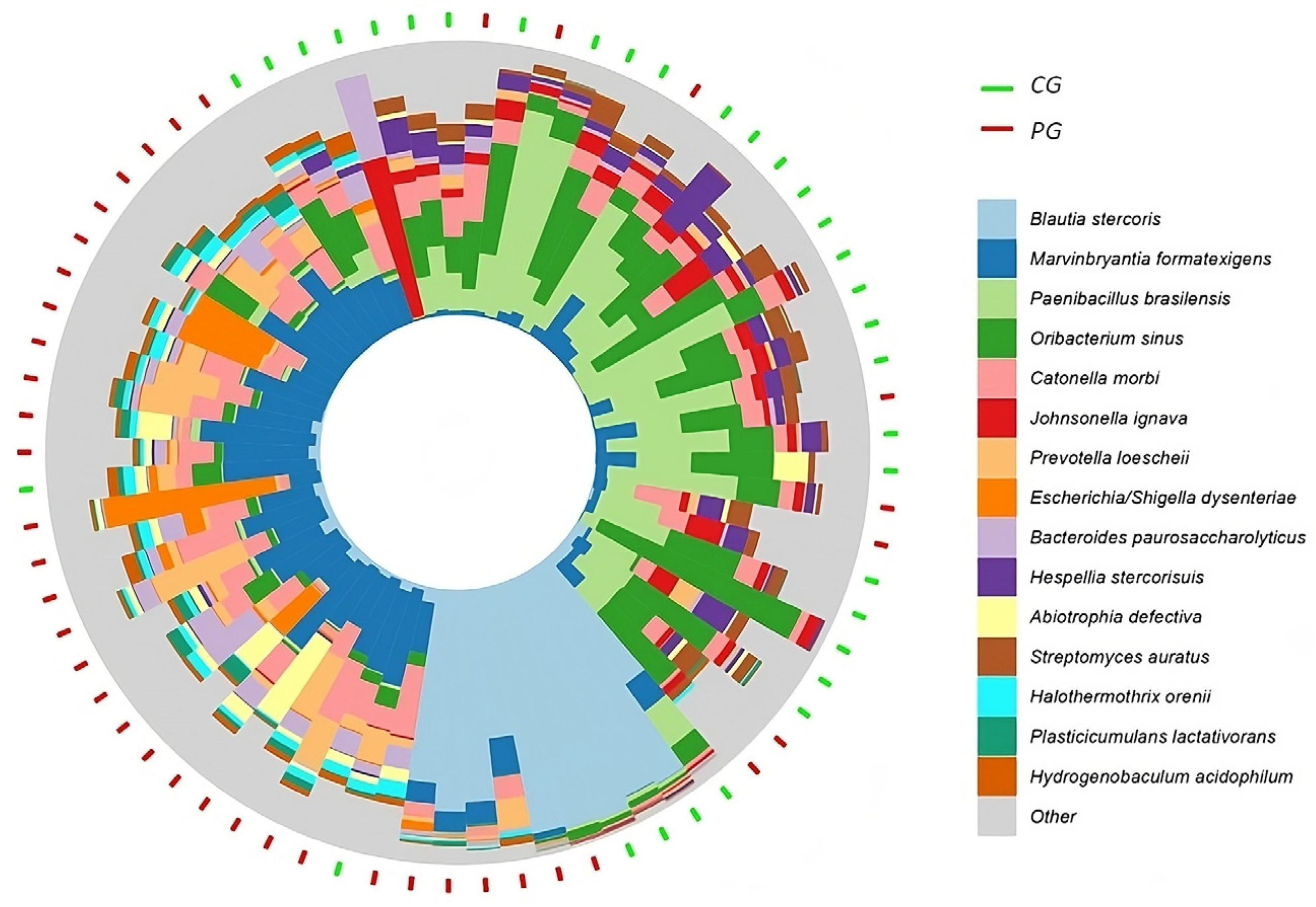
| Taxonomy (Phylum and Species) | CG | PG | Adjusted p |
|---|---|---|---|
| Phylum Firmicutes | ar-Ab (sd) | ar-Ab (sd) | |
| Blautia stercoris | 4.97 (19.43) | 7.11 (20.22) | 0.0034 |
| Paenibacillus brasilensis | 1.46 (2.02) | 0.68 (1.65) | 0.0062 |
| Oribacterium sinus | 1.39 (1.20) | 0.49 (0.56) | 0.0130 |
| Johnsonella ignava | 0.93 (3.77) | 0.08 (0.11) | 0.0012 |
| Marvinbryantia formatexigens | 0.80 (1.71) | 2.91 (2.18) | 0.0006 |
| Catonella morbi | 0.54 (0.46) | 1.31 (0.85) | 0.0056 |
| Hespellia stercorisuis | 0.36 (0.35) | 0.10 (0.10) | 0.0051 |
| Thermodesulfobium narugense | 0.13 (0.11) | 0.07 (0.14) | 0.0370 |
| Selenomonas flueggei | 0.12 (0.10) | 0.06 (0.12) | 0.0250 |
| Abiotrophia defectiva | 0.08 (0.16) | 0.57 (1.18) | 0.0023 |
| Halothermothrix orenii | 0.07 (0.18) | 0.28 (0.23) | 0.0029 |
| Filifactor villosus | 0.06 (0.06) | 0.02 (0.04) | 0.0160 |
| Butyrivibrio proteoclasticus | 0.06 (0.11) | 0.01 (0.02) | 0.0360 |
| Oribacterium asaccharolyticum | 0.06 (0.06) | 0.02 (0.04) | 0.0330 |
| Facklamia ignava | 0.05 (0.08) | 0.23 (0.23) | 0.0007 |
| Sporomusa ovata | 0.04 (0.14) | 0.09 (0.09) | 0.0160 |
| Pseudoramibacter alactolyticus | 0.03 (0.07) | 0.09 (0.10) | 0.0250 |
| Marininema halotolerans | 0.02 (0.06) | 0.16 (0.18) | 0.0021 |
| Clostridium tepidiprofundi | 0.01 (0.02) | 0 (0.02) | 0.0260 |
| Acetobacterium fimetarium | 0.01 (0.04) | 0.07 (0.09) | 0.0330 |
| Mahella australiensis | 0.01 (0.02) | 0.04 (0.06) | 0.0250 |
| Clostridium aminophilum | 0.01 (0.02) | 0.03 (0.05) | 0.0098 |
| Selenomonas sputigena | 0.01 (0.02) | 0.07 (0.14) | 0.0051 |
| Gelria glutamica | 0.01 (0.03) | 0.12 (0.11) | 0.0012 |
| Sporanaerobacter acetigenes | 0.01 (0.03) | 0.22 (0.31) | 0.0058 |
| Shimazuella kribbensis | 0.01 (0.04) | 0.10 (0.12) | 0.0110 |
| Carboxydocella manganica | 0 (0.01) | 0.05 (0.06) | 0.0055 |
| Phylum Bacteroidetes | ar-Ab (sd) | ar-Ab (sd) | |
| Bacteroides paurosaccharolyticus | 0.49 (1.97) | 0.51 (0.52) | 0.0120 |
| Prevotella loescheii | 0.20 (0.46) | 0.85 (1.38) | 0.0360 |
| Bacteroides heparinolyticus | 0.13 (0.15) | 0.05 (0.14) | 0.0340 |
| Sediminitomix flava | 0.07 (0.16) | 0.15 (0.15) | 0.0140 |
| Bacteroides barnesiae | 0.06 (0.12) | 0.16 (0.14) | 0.0058 |
| Prevotella marshii | 0.05 (0.08) | 0.01 (0.04) | 0.0160 |
| Perexilibacter aurantiacus | 0.02 (0.05) | 0.07 (0.07) | 0.0017 |
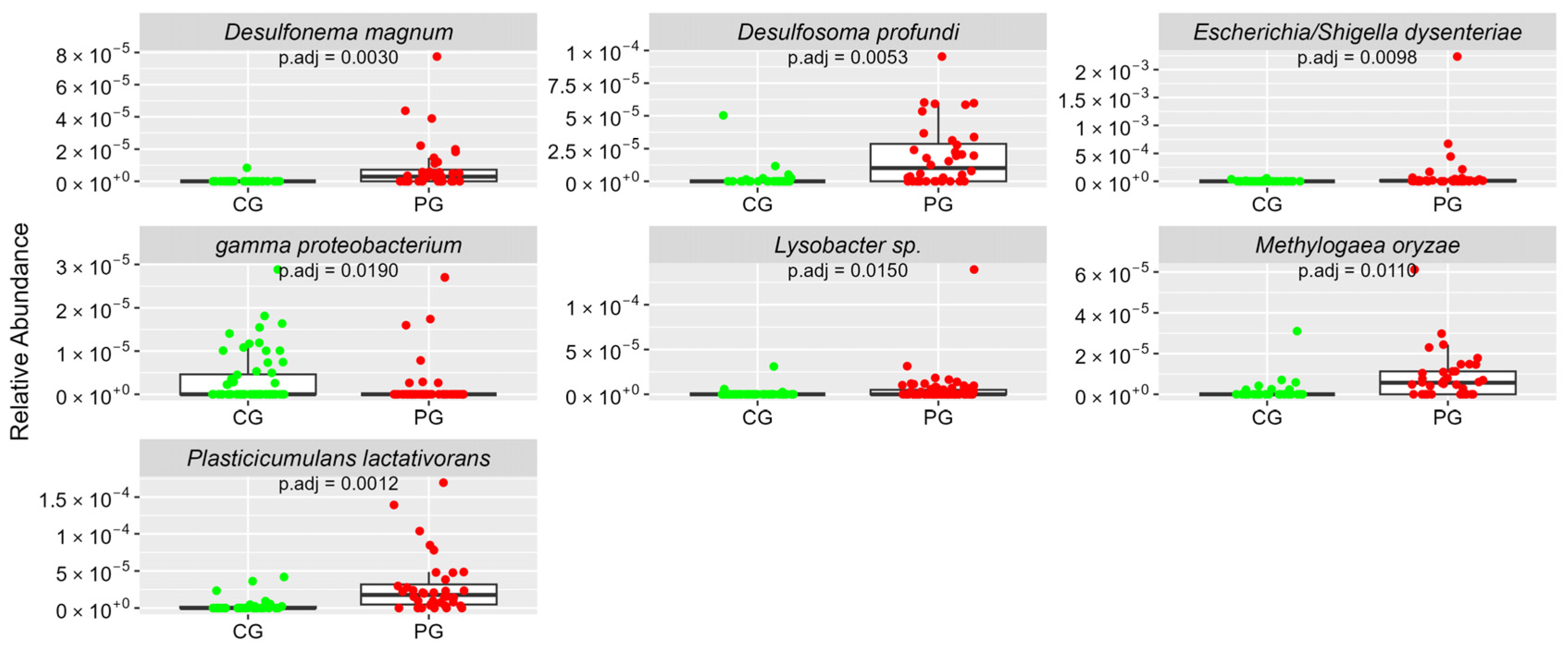
| Phylum Proteobacteria | ar-Ab (sd) | ar-Ab (sd) | |
| Escherichia/Shigella dysenteriae | 0.04 (0.12) | 1.14 (3.88) | 0.0098 |
| Plasticicumulans lactativorans | 0.04 (0.11) | 0.3 (0.39) | 0.0012 |
| Gamma proteobacterium | 0.03 (0.06) | 0.01 (0.04) | 0.0190 |
| Desulfosoma profundi | 0.02 (0.09) | 0.19 (0.24) | 0.0053 |
| Methylogaea oryzae | 0.02 (0.06) | 0.09 (0.12) | 0.0110 |
| Lysobacter sp. | 0.01 (0.04) | 0.05 (0.17) | 0.0150 |
| Desulfonema magnum | 0 (0.02) | 0.09 (0.16) | 0.0030 |
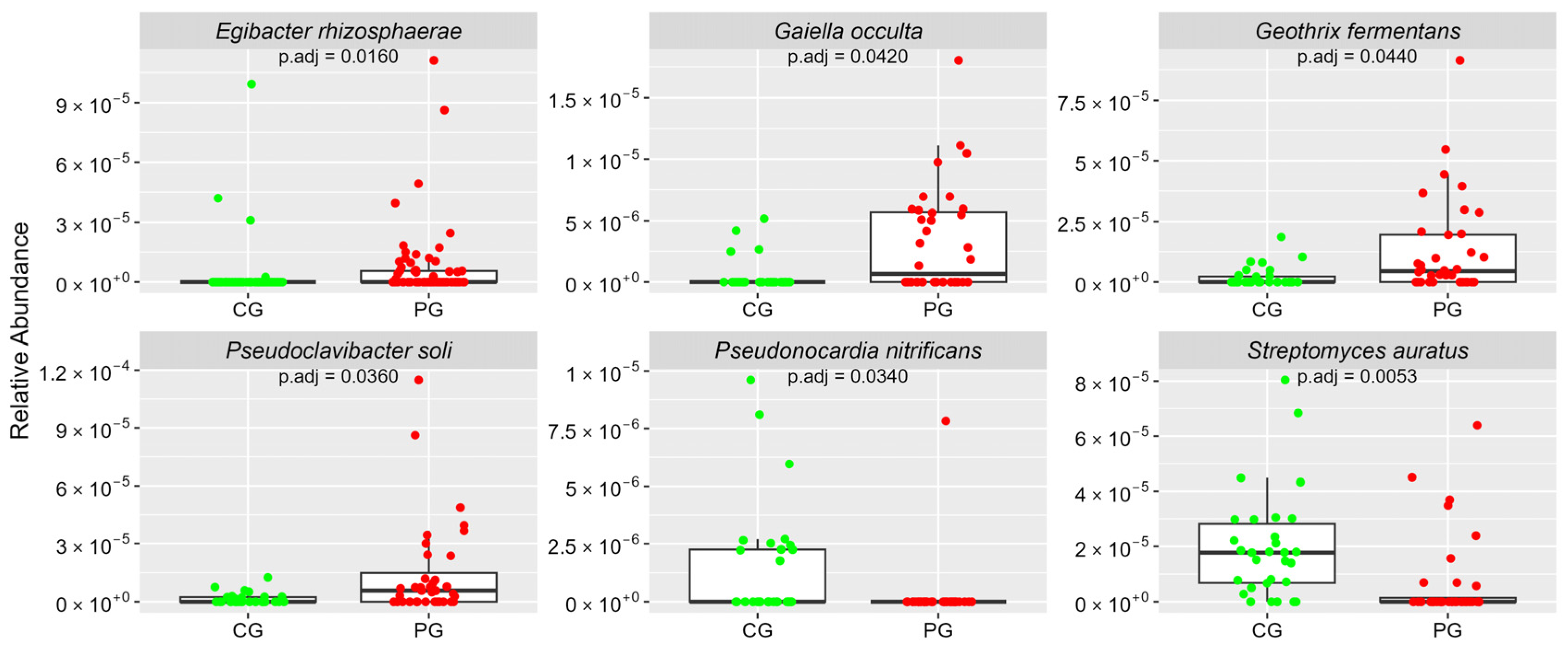
| Phylum Actinobacteria | ar-Ab (sd) | ar-Ab (sd) | |
| Geothrix fermentans | 0.02 (0.04) | 0.13 (0.20) | 0.0440 |
| Streptomyces auratus | 0.2 (0.19) | 0.07 (0.15) | 0.0053 |
| Egibacter rhizosphaerae | 0.03 (0.14) | 0.07 (0.18) | 0.0160 |
| Pseudoclavibacter soli | 0.02 (0.03) | 0.15 (0.25) | 0.0360 |
| Pseudonocardia nitrificans | 0.01 (0.02) | 0 (0.01) | 0.0340 |
| Gaiella occulta | 0 (0.01) | 0.03 (0.04) | 0.0420 |
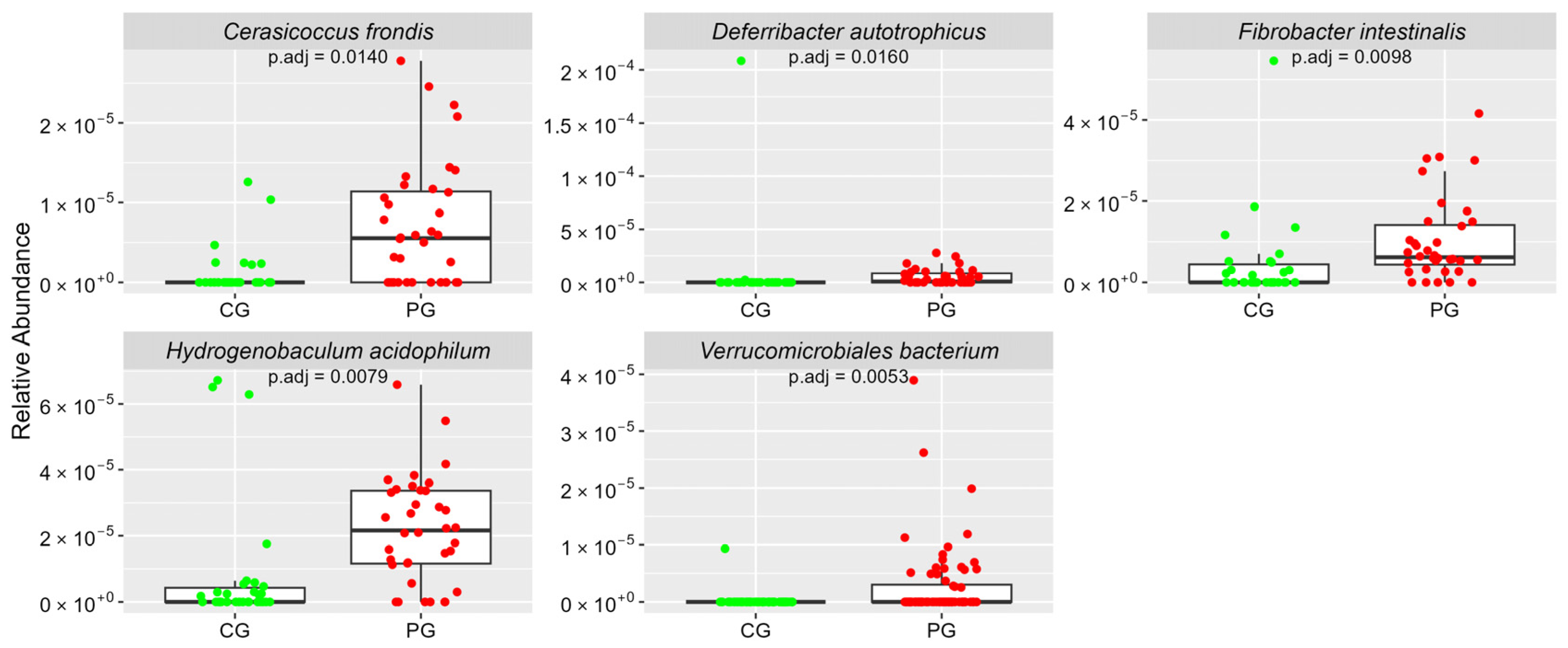
| Others | ar-Ab (sd) | ar-Ab (sd) | |
| Hydrogenobaculum acidophilum | 0.08 (0.20) | 0.22 (0.16) | 0.0079 |
| Deferribacter autotrophicus | 0.07 (0.38) | 0.05 (0.07) | 0.0160 |
| Fibrobacter intestinalis | 0.04 (0.11) | 0.10 (0.10) | 0.0098 |
| Cerasicoccus frondis | 0.01 (0.03) | 0.07 (0.08) | 0.0140 |
| Verrucomicrobiales bacterium | 0 (0.01) | 0.03 (0.06) | 0.0053 |
| Bacterial Species | CG ar-Ab | PG ar-Ab | p | Association with Pathology or Health |
|---|---|---|---|---|
| Filifactor villosus * | 0.06 (0.06) | 0.02 (0.04) | 0.0160 | Inversely associated with the severity of Alzheimer’s disease [26] |
| Gelria glutamica ** | 0.01 (0.03) | 0.12 (0.11) | 0.0012 | Glutamate-degrading bacteria [27] |
| Johnsonella ignava * | 0.93 (3.77) | 0.08 (0.11) | 0.0012 | Inversely associated with carotid plaques and inflammatory activation [28] |
| Pseudoramibacteralactolyticus ** | 0.03 (0.07) | 0.09 (0.10) | 0.0250 | Associated with endodontic infections [29] |
| Selenomonas sputigena ** | 0.01 (0.02) | 0.07 (0.14) | 0.0051 | Associated with periodontitis [30] |
| Abiotrophia defectiva ** | 0.08 (0.16) | 0.57 (1.18) | 0.0023 | Associated with the pathogenesis of endocarditis [31] |
| Catonella morbi ** | 0.54 (0.46) | 1.31 (0.85) | 0.0056 | Associated with periodontal disease [32] |
| Paenibacillus brasilensis * | 1.46 (2.02) | 0.68 (1.65) | 0.0062 | Produces antimicrobial substance against the pathological species Cryptococcus neoformans [33] |
| Escherichia/Shigella dysenteriae ** | 0.04 (0.12) | 1.14 (3.88) | 0.0098 | Causes inflammation and ulceration of the mucosa of the large intestine [34] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tieppo, A.M.; Tieppo, J.S.; Rivetti, L.A. Analysis of Intestinal Bacterial Microbiota in Individuals with and without Chronic Low Back Pain. Curr. Issues Mol. Biol. 2024, 46, 7339-7352. https://doi.org/10.3390/cimb46070435
Tieppo AM, Tieppo JS, Rivetti LA. Analysis of Intestinal Bacterial Microbiota in Individuals with and without Chronic Low Back Pain. Current Issues in Molecular Biology. 2024; 46(7):7339-7352. https://doi.org/10.3390/cimb46070435
Chicago/Turabian StyleTieppo, Antonio Martins, Júlia Silva Tieppo, and Luiz Antonio Rivetti. 2024. "Analysis of Intestinal Bacterial Microbiota in Individuals with and without Chronic Low Back Pain" Current Issues in Molecular Biology 46, no. 7: 7339-7352. https://doi.org/10.3390/cimb46070435