
TAAR8 Mediates Increased Migrasome Formation by Cadaverine in RPE Cells



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Image-Based Fecal Metabolite Library Screening
2.3. Migrasome Counting and Live Cell Imaging
2.4. Western Blot Analysis
2.5. Statistical Analyses
3. Results and Discussion
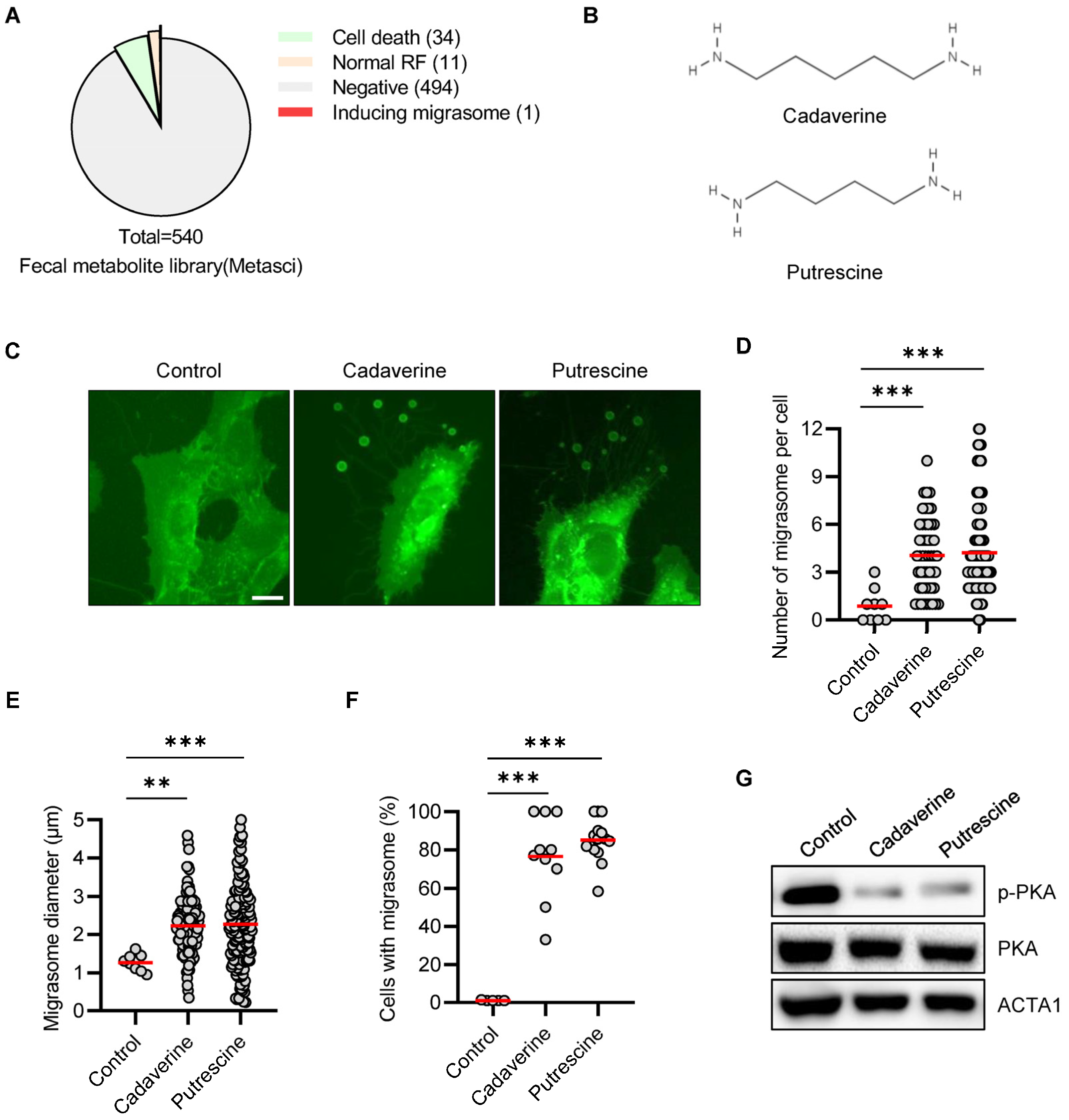
3.1. Cadaverine and Putrescine Promotes Migrasome in RPE/GFP-TSPAN4 Cells
3.2. Cadaverine and Putrescine Inhibit PKA Phosphorylation in RPE Cells
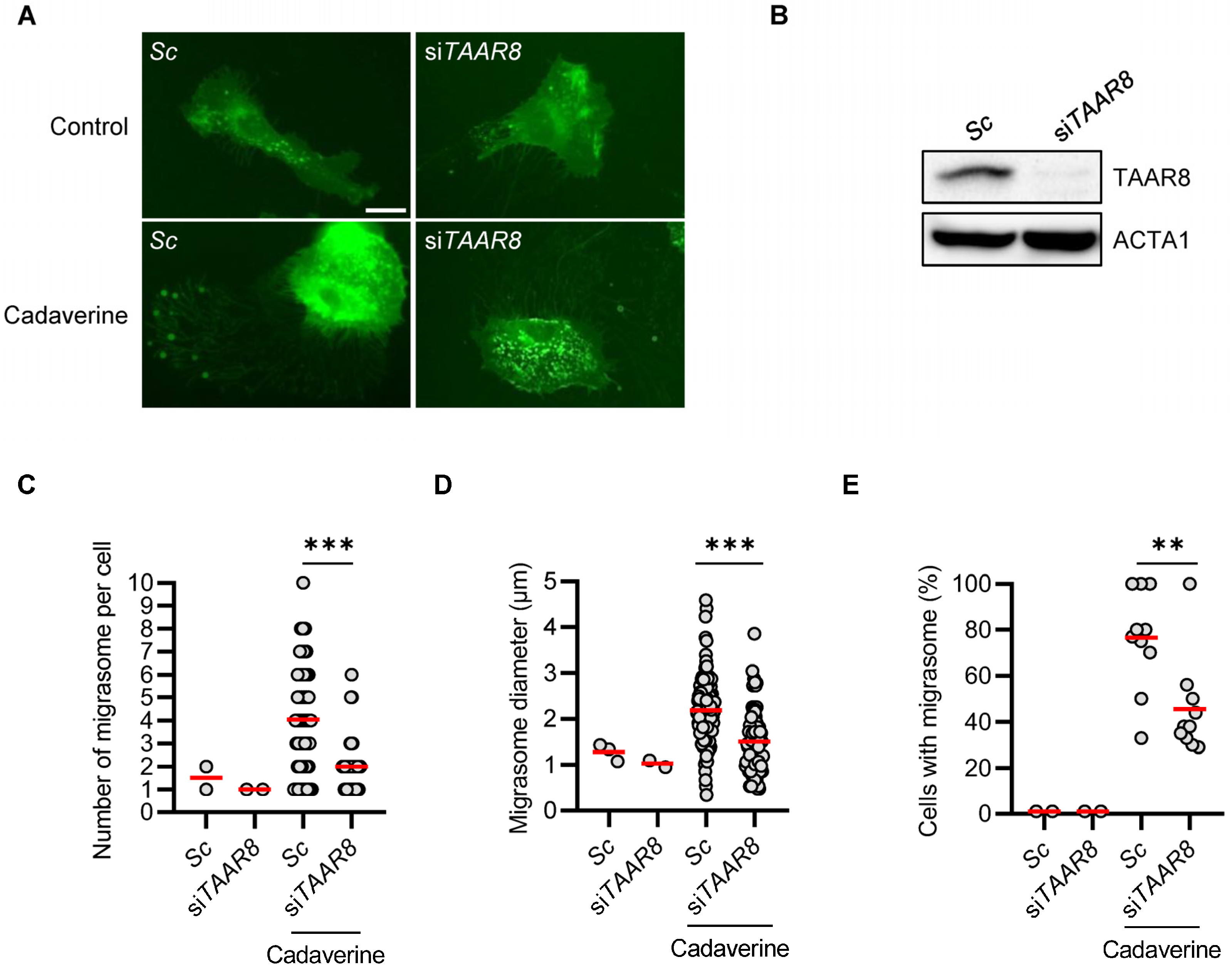
3.3. TAAR8 Knock-Down Inhibits Cadaverine-Inducing Migrasome Formation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, L.; Li, Y.; Peng, J.; Wu, D.; Zhao, X.; Cui, Y.; Chen, L.; Yan, X.; Du, Y.; Yu, L. Discovery of the migrasome, an organelle mediating release of cytoplasmic contents during cell migration. Cell Res. 2015, 25, 24–38. [Google Scholar] [CrossRef] [PubMed]
- da Rocha-Azevedo, B.; Schmid, S.L. Migrasomes: A new organelle of migrating cells. Cell Res. 2015, 25, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Xu, Y.; Ding, T.; Zu, Y.; Yang, C.; Yu, L. Pairing of integrins with ECM proteins determines migrasome formation. Cell Res. 2017, 27, 1397–1400. [Google Scholar] [CrossRef]
- Huang, Y.; Yu, L. Tetraspanin-enriched microdomains: The building blocks of migrasomes. Cell Insight 2022, 1, 100003. [Google Scholar] [CrossRef]
- Huang, Y.; Zucker, B.; Zhang, S.; Elias, S.; Zhu, Y.; Chen, H.; Ding, T.; Li, Y.; Sun, Y.; Lou, J.; et al. Migrasome formation is mediated by assembly of micron-scale tetraspanin macrodomains. Nat. Cell Biol. 2019, 21, 991–1002. [Google Scholar] [CrossRef]
- Jiang, D.; Jiang, Z.; Lu, D.; Wang, X.; Liang, H.; Zhang, J.; Meng, Y.; Li, Y.; Wu, D.; Huang, Y.; et al. Migrasomes provide regional cues for organ morphogenesis during zebrafish gastrulation. Nat. Cell Biol. 2019, 21, 966–977. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, L.; Meng, Y.; Li, B.; Yang, Y.; Gao, F. Migrasome: A new functional extracellular vesicle. Cell Death Discov. 2023, 9, 381. [Google Scholar] [CrossRef]
- Dharan, R.; Huang, Y.; Cheppali, S.K.; Goren, S.; Shendrik, P.; Wang, W.; Qiao, J.; Kozlov, M.M.; Yu, L.; Sorkin, R. Tetraspanin 4 stabilizes membrane swellings and facilitates their maturation into migrasomes. Nat. Commun. 2023, 14, 1037. [Google Scholar] [CrossRef]
- Tan, X.; He, S.; Wang, F.; Li, L.; Wang, W. Migrasome, a novel organelle, differs from exosomes. Biochem. Biophys. Rep. 2023, 35, 101500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, J.; Ding, Y.; Zhang, J.; Xu, Y.; Xu, J.; Zheng, S.; Yang, H. Migrasome and Tetraspanins in Vascular Homeostasis: Concept, Present, and Future. Front. Cell Dev. Biol. 2020, 8, 438. [Google Scholar] [CrossRef]
- Runge, K.E.; Evans, J.E.; He, Z.Y.; Gupta, S.; McDonald, K.L.; Stahlberg, H.; Primakoff, P.; Myles, D.G. Oocyte CD9 is enriched on the microvillar membrane and required for normal microvillar shape and distribution. Dev. Biol. 2007, 304, 317–325. [Google Scholar] [CrossRef]
- Jiao, H.; Jiang, D.; Hu, X.; Du, W.; Ji, L.; Yang, Y.; Li, X.; Sho, T.; Wang, X.; Li, Y.; et al. Mitocytosis, a migrasome-mediated mitochondrial quality-control process. Cell 2021, 184, 2896–2910.e13. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Liu, X.; Ye, J.; Ma, Y.; Mao, J.; Feng, D.; Wang, X. Migrasomes, a new mode of intercellular communication. Cell Commun. Signal 2023, 21, 105. [Google Scholar] [CrossRef] [PubMed]
- Kovács, T.; Mikó, E.; Vida, A.; Sebő, É.; Toth, J.; Csonka, T.; Boratkó, A.; Ujlaki, G.; Lente, G.; Kovács, P.; et al. Cadaverine, a metabolite of the microbiome, reduces breast cancer aggressiveness through trace amino acid receptors. Sci. Rep. 2019, 9, 1300. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, C.; Gomez-Tamayo, J.C.; Nebel, J.C.; Pardo, L.; Gonzalez, A. Identifying human diamine sensors for death related putrescine and cadaverine molecules. PLoS Comput. Biol. 2018, 14, e1005945. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, L.; Ebeling, M.; Kratochwil, N.A.; Bunzow, J.R.; Grandy, D.K.; Hoener, M.C. Trace amine-associated receptors form structurally and functionally distinct subfamilies of novel G protein-coupled receptors. Genomics 2005, 85, 372–385. [Google Scholar] [CrossRef] [PubMed]
- Muhlhaus, J.; Dinter, J.; Nurnberg, D.; Rehders, M.; Depke, M.; Golchert, J.; Homuth, G.; Yi, C.X.; Morin, S.; Kohrle, J.; et al. Analysis of human TAAR8 and murine Taar8b mediated signaling pathways and expression profile. Int. J. Mol. Sci. 2014, 15, 20638–20655. [Google Scholar] [CrossRef] [PubMed]
- Howe, A.K. Regulation of actin-based cell migration by cAMP/PKA. Biochim. Biophys. Acta 2004, 1692, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Galvanetto, N.; Nie, J.; Yang, Y.; Torre, V. Rac1 Promotes Cell Motility by Controlling Cell Mechanics in Human Glioblastoma. Cancers 2020, 12, 1667. [Google Scholar] [CrossRef] [PubMed]
- Romarowski, A.; Battistone, M.A.; La Spina, F.A.; Puga Molina Ldel, C.; Luque, G.M.; Vitale, A.M.; Cuasnicu, P.S.; Visconti, P.E.; Krapf, D.; Buffone, M.G. PKA-dependent phosphorylation of LIMK1 and Cofilin is essential for mouse sperm acrosomal exocytosis. Dev. Biol. 2015, 405, 237–249. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.B.; Bae, J.-E.; Park, N.Y.; Kim, Y.H.; Kim, S.H.; Hyung, H.; Yeom, E.; Choi, D.K.; Jeong, K.; Cho, D.-H. TAAR8 Mediates Increased Migrasome Formation by Cadaverine in RPE Cells. Curr. Issues Mol. Biol. 2024, 46, 8658-8664. https://doi.org/10.3390/cimb46080510
Kim JB, Bae J-E, Park NY, Kim YH, Kim SH, Hyung H, Yeom E, Choi DK, Jeong K, Cho D-H. TAAR8 Mediates Increased Migrasome Formation by Cadaverine in RPE Cells. Current Issues in Molecular Biology. 2024; 46(8):8658-8664. https://doi.org/10.3390/cimb46080510
Chicago/Turabian StyleKim, Joon Bum, Ji-Eun Bae, Na Yeon Park, Yong Hwan Kim, Seong Hyun Kim, Hyejin Hyung, Eunbyul Yeom, Dong Kyu Choi, Kwiwan Jeong, and Dong-Hyung Cho. 2024. "TAAR8 Mediates Increased Migrasome Formation by Cadaverine in RPE Cells" Current Issues in Molecular Biology 46, no. 8: 8658-8664. https://doi.org/10.3390/cimb46080510
APA StyleKim, J. B., Bae, J.-E., Park, N. Y., Kim, Y. H., Kim, S. H., Hyung, H., Yeom, E., Choi, D. K., Jeong, K., & Cho, D.-H. (2024). TAAR8 Mediates Increased Migrasome Formation by Cadaverine in RPE Cells. Current Issues in Molecular Biology, 46(8), 8658-8664. https://doi.org/10.3390/cimb46080510