
Efficacy of SGPP2 Modulation-Mediated Materials in Ameliorating Facial Wrinkles and Pore Sagging


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Preparation
2.2. Transfection of Keratinocytes with siRNA for RNAi Experiments
2.3. RNA Extraction and RT-qPCR
2.4. Conditioned-Media Experiment
2.5. Reconstructed Three-Dimensional (3D) Skin Experiment
2.6. Human Clinical Trial
2.7. Statistical Analysis
3. Results and Discussion
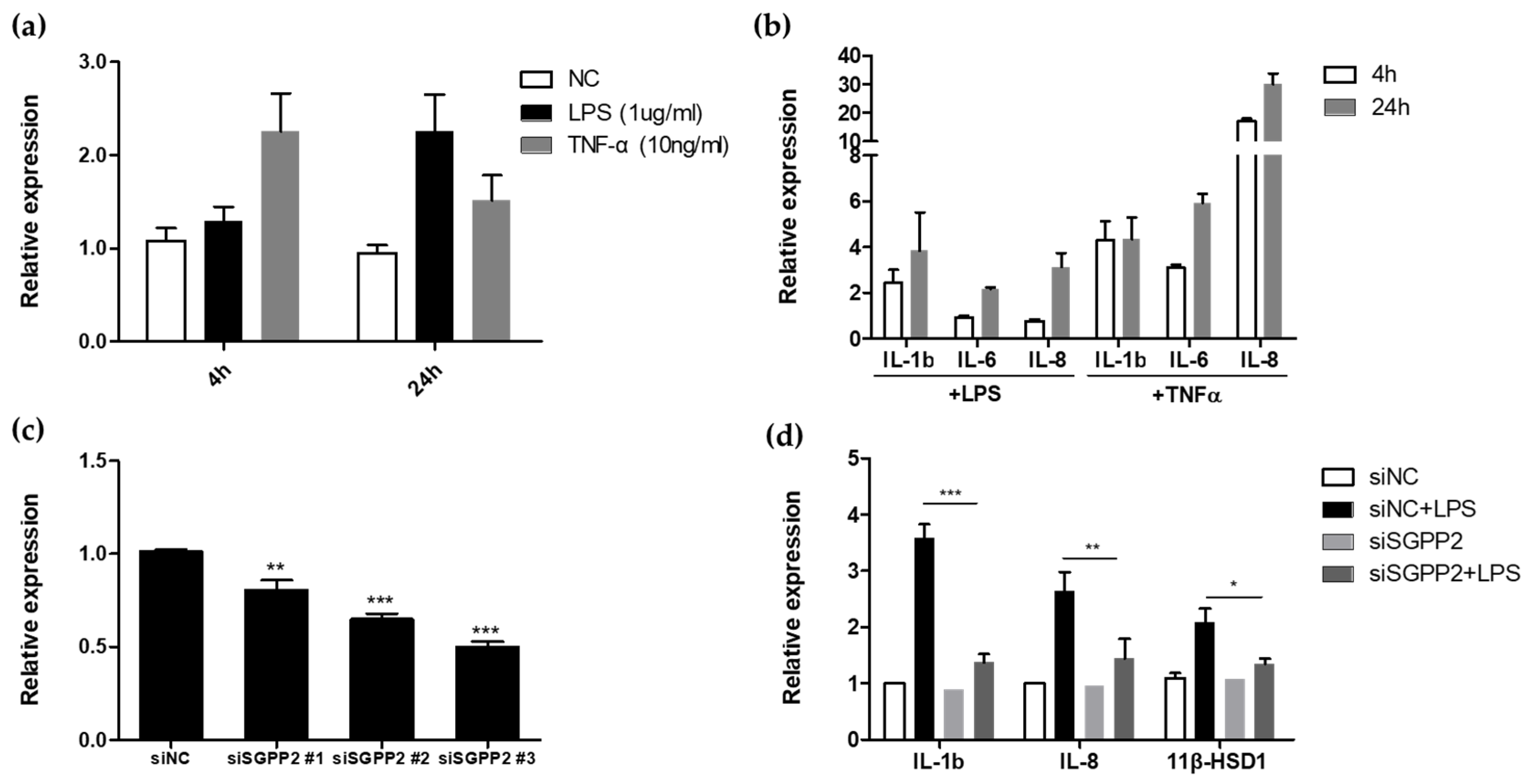
3.1. Functional Study of Wrinkle-Related Gene SGPP2 in HaCaT Cells
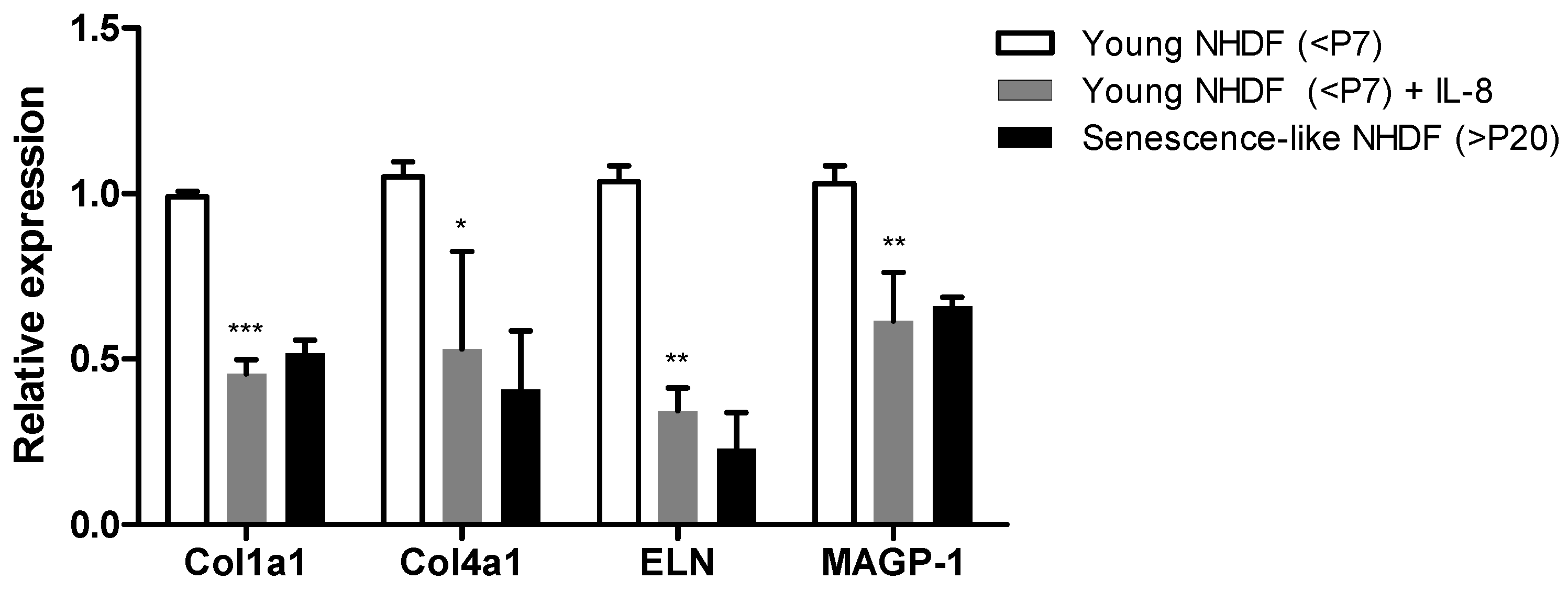
3.2. Inflammation Exacerbates Skin Fibroblast-Mediated Wrinkle Formation
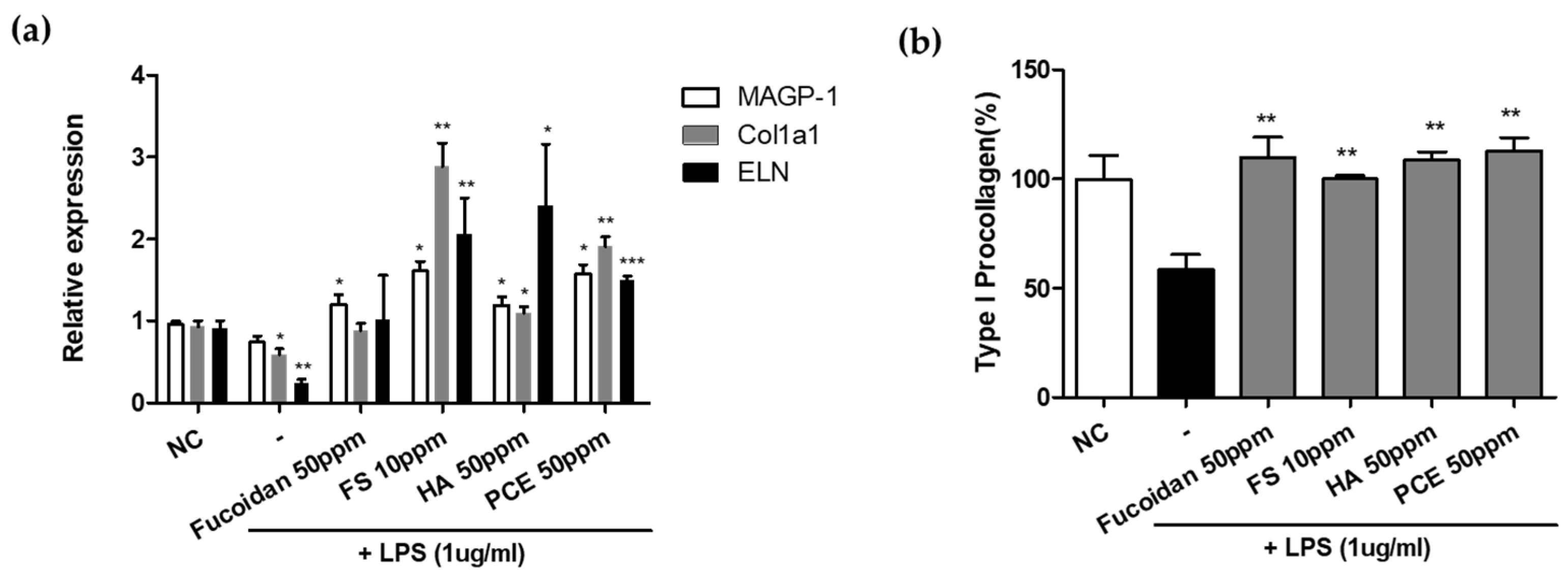
3.3. Screening Active Materials That Regulate SGPP2 Expression
3.4. Antiaging Effect of SGPP2 Expression-Regulating Materials In Vitro
3.4.1. Recovery of Inflammation-Induced Decrease in ECM-Related Gene in NHDFs
3.4.2. Effect of Enhanced Type I Procollagen Synthesis
3.5. Dermal Collagen Enhances the Efficacy of the Selected Materials within 3D Skin Model
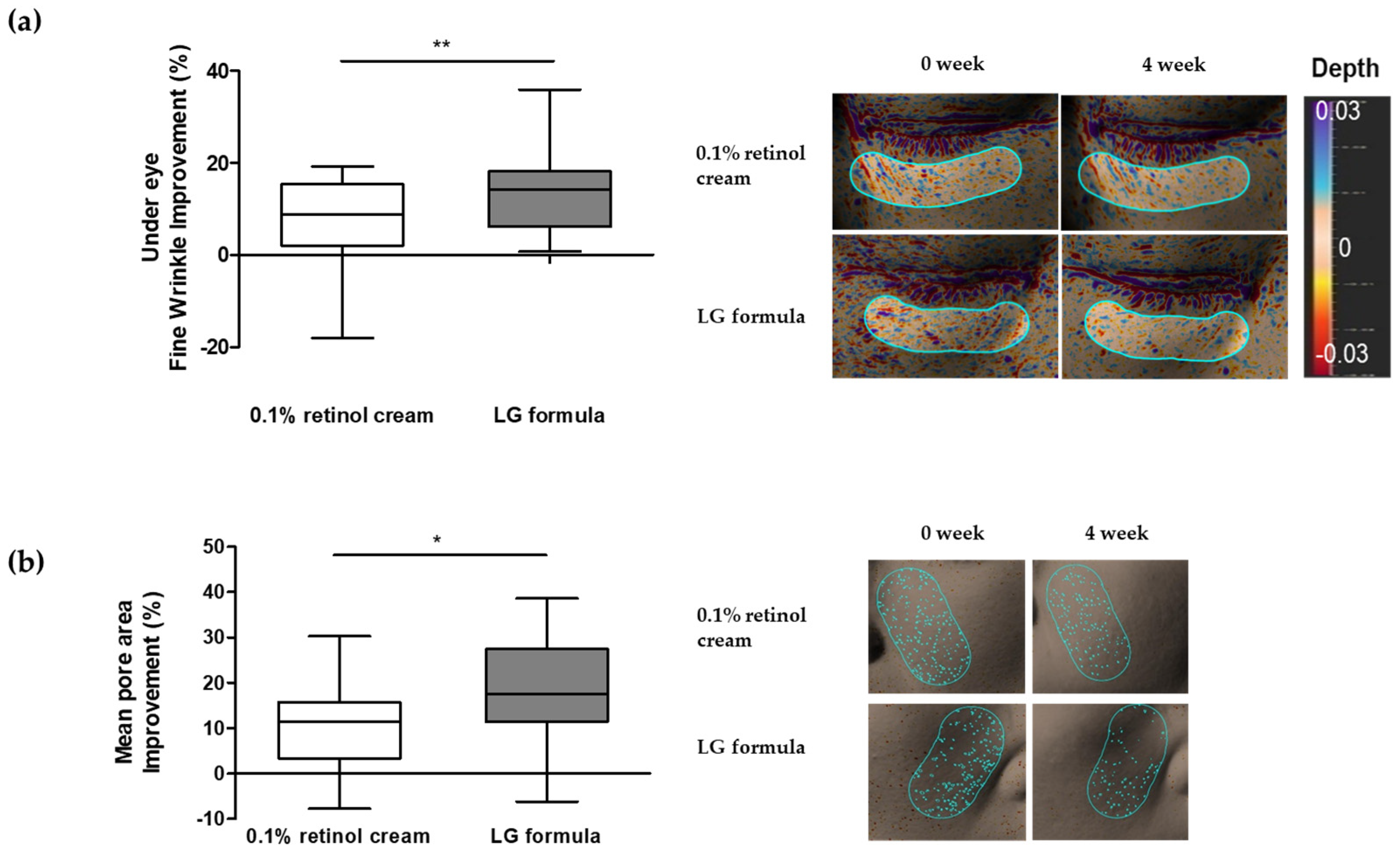
3.6. Improvement in Skin Wrinkles and Pores Using LG Formula-Containing Materials That Modulated SGPP2 Expression
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, A.S.; Dreesen, O. Biomarkers of Cellular Senescence and Skin Aging. Front. Genet. 2018, 9, 247. [Google Scholar] [CrossRef]
- Fuller, B. Role of PGE-2 and Other Inflammatory Mediators in Skin Aging and Their Inhibition by Topical Natural Anti-Inflammatories. Cosmetics 2019, 6, 6. [Google Scholar] [CrossRef]
- Salminen, A.; Kauppinen, A.; Kaarniranta, K. Emerging role of NF-kappaB signaling in the induction of senescence-associated secretory phenotype (SASP). Cell Signal. 2012, 24, 835–845. [Google Scholar] [CrossRef]
- Budamagunta, V.; Manohar-Sindhu, S.; Yang, Y.; He, Y.; Traktuev, D.O.; Foster, T.C.; Zhou, D. Senescence-associated hyper-activation to inflammatory stimuli in vitro. Aging 2021, 13, 19088–19107. [Google Scholar] [CrossRef] [PubMed]
- Bielak-Zmijewska, A.; Grabowska, W.; Ciolko, A.; Bojko, A.; Mosieniak, G.; Bijoch, L.; Sikora, E. The Role of Curcumin in the Modulation of Ageing. Int. J. Mol. Sci. 2019, 20, 1239. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.H. Prostaglandin E2 Induces Skin Aging via E-Prostanoid 1 in Normal Human Dermal Fibroblasts. Int. J. Mol. Sci. 2019, 20, 5555. [Google Scholar] [CrossRef]
- Baechle, J.J.; Chen, N.; Makhijani, P.; Winer, S.; Furman, D.; Winer, D.A. Chronic inflammation and the hallmarks of aging. Mol. Metab. 2023, 74, 101755. [Google Scholar] [CrossRef]
- Chin, T.; Lee, X.E.; Ng, P.Y.; Lee, Y.; Dreesen, O. The role of cellular senescence in skin aging and age-related skin pathologies. Front. Physiol. 2023, 14, 1297637. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, H.; Man, M.Q.; Hu, L. Aging in the dermis: Fibroblast senescence and its significance. Aging Cell 2024, 23, e14054. [Google Scholar] [CrossRef]
- Jang, S.I.; Kim, E.J.; Lee, H.K. A method of evaluating facial pores using optical 2D images and analysis of age-dependent changes in facial pores in Koreans. Skin Res. Technol. 2018, 24, 304–308. [Google Scholar] [CrossRef]
- Jung, H.J.; Ahn, J.Y.; Lee, J.I.; Bae, J.Y.; Kim, H.L.; Suh, H.Y.; Youn, J.I.; Park, M.Y. Analysis of the number of enlarged pores according to site, age, and sex. Skin Res. Technol. 2018, 24, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, B.; Miyamoto, K.; Purwar, A.; Chye, R.; Matsubara, A. New image analysis tool for facial pore characterization and assessment. Skin Res. Technol. 2019, 25, 631–638. [Google Scholar] [CrossRef]
- Lee, S.; Cherel, M.; Gougeon, S.; Jeong, E.; Lim, J.M.; Park, S.G. Identifying patterns behind the changes in skin pores using 3-dimensional measurements and K-means clustering. Skin Res. Technol. 2022, 28, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Lanoue, J.; Goldenberg, G. Enlarged facial pores: An update on treatments. Cutis 2016, 98, 33–36. [Google Scholar]
- Messaraa, C.; Metois, A.; Walsh, M.; Flynn, J.; Doyle, L.; Robertson, N.; Mansfield, A.; O’Connor, C.; Mavon, A. Antera 3D capabilities for pore measurements. Skin Res. Technol. 2018, 24, 606–613. [Google Scholar] [CrossRef]
- Nkengne, A.; Pellacani, G.; Ciardo, S.; De Carvalho, N.; Vie, K. Visible characteristics and structural modifications relating to enlarged facial pores. Skin Res. Technol. 2021, 27, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Chen, F.; Feng, X.; Shang, J.; Luo, X.; Chen, Y. Potential role of inflammaging mediated by the complement system in enlarged facial pores. J. Cosmet. Dermatol. 2024, 23, 27–32. [Google Scholar] [CrossRef]
- Inui, S.; Mori, A.; Ito, M.; Hyodo, S.; Itami, S. Reduction of conspicuous facial pores by topical fullerene: Possible role in the suppression of PGE2 production in the skin. J. Nanobiotechnol. 2014, 12, 6. [Google Scholar] [CrossRef]
- Lee, S.G.; Shin, J.G.; Kim, Y.; Leem, S.; Park, S.G.; Won, H.H.; Kang, N.G. Identification of Genetic Loci Associated with Facial Wrinkles in a Large Korean Population. J. Investig. Dermatol. 2022, 142, 2824–2827. [Google Scholar] [CrossRef]
- Cha, M.-Y.; Choi, J.-E.; Lee, D.-S.; Lee, S.-R.; Lee, S.-I.; Park, J.-H.; Shin, J.-H.; Suh, I.S.; Kim, B.H.; Hong, K.-W. Novel Genetic Associations for Skin Aging Phenotypes and Validation of Previously Reported Skin GWAS Results. Appl. Sci. 2022, 12, 11422. [Google Scholar] [CrossRef]
- Lee, S.; Ye, S.; Kim, M.; Lee, H.; Jun, S.H.; Kang, N.G. Fine Wrinkle Improvement through Bioactive Materials That Modulate EDAR and BNC2 Gene Expression. Biomolecules 2024, 14, 279. [Google Scholar] [CrossRef]
- Lee, H.; Ye, S.; Kim, J.; Jun, S.H.; Kang, N.G. Improvement in Facial Wrinkles Using Materials Enhancing PPARGC1B Expression Related to Mitochondrial Function. Curr. Issues Mol. Biol. 2024, 46, 5037–5051. [Google Scholar] [CrossRef]
- Kleuser, B.; Baumer, W. Sphingosine 1-Phosphate as Essential Signaling Molecule in Inflammatory Skin Diseases. Int. J. Mol. Sci. 2023, 24, 1456. [Google Scholar] [CrossRef]
- Noujarede, J.; Carrie, L.; Garcia, V.; Grimont, M.; Eberhardt, A.; Mucher, E.; Genais, M.; Schreuder, A.; Carpentier, S.; Segui, B.; et al. Sphingolipid paracrine signaling impairs keratinocyte adhesion to promote melanoma invasion. Cell Rep. 2023, 42, 113586. [Google Scholar] [CrossRef]
- Allende, M.L.; Sipe, L.M.; Tuymetova, G.; Wilson-Henjum, K.L.; Chen, W.; Proia, R.L. Sphingosine-1-phosphate phosphatase 1 regulates keratinocyte differentiation and epidermal homeostasis. J. Biol. Chem. 2013, 288, 18381–18391. [Google Scholar] [CrossRef]
- Baumer, W.; Rossbach, K.; Mischke, R.; Reines, I.; Langbein-Detsch, I.; Luth, A.; Kleuser, B. Decreased concentration and enhanced metabolism of sphingosine-1-phosphate in lesional skin of dogs with atopic dermatitis: Disturbed sphingosine-1-phosphate homeostasis in atopic dermatitis. J. Investig. Dermatol. 2011, 131, 266–268. [Google Scholar] [CrossRef]
- Mechtcheriakova, D.; Wlachos, A.; Sobanov, J.; Kopp, T.; Reuschel, R.; Bornancin, F.; Cai, R.; Zemann, B.; Urtz, N.; Stingl, G.; et al. Sphingosine 1-phosphate phosphatase 2 is induced during inflammatory responses. Cell Signal. 2007, 19, 748–760. [Google Scholar] [CrossRef]
- Jeong, E.T.; Jin, M.H.; Kim, M.S.; Chang, Y.H.; Park, S.G. Inhibition of melanogenesis by piceid isolated from Polygonum cuspidatum. Arch. Pharm. Res. 2010, 33, 1331–1338. [Google Scholar] [CrossRef]
- Seok, J.K.; Boo, Y.C. p-Coumaric Acid Attenuates UVB-Induced Release of Stratifin from Keratinocytes and Indirectly Regulates Matrix Metalloproteinase 1 Release from Fibroblasts. Korean J. Physiol. Pharmacol. 2015, 19, 241–247. [Google Scholar] [CrossRef]
- Oh, S.R.; Park, S.K.; Lee, P.; Kim, Y.M. The ginsenoside Rg2 downregulates MMP-1 expression in keratinocyte (HaCaT)-conditioned medium-treated human fibroblasts (Hs68). Appl. Biol. Chem. 2023, 66, 85. [Google Scholar] [CrossRef]
- Cheong, R.; Hoffmann, A.; Levchenko, A. Understanding NF-kappaB signaling via mathematical modeling. Mol. Syst. Biol. 2008, 4, 192. [Google Scholar] [CrossRef]
- Choe, S.J.; Kim, D.; Kim, E.J.; Ahn, J.S.; Choi, E.J.; Son, E.D.; Lee, T.R.; Choi, E.H. Psychological Stress Deteriorates Skin Barrier Function by Activating 11beta-Hydroxysteroid Dehydrogenase 1 and the HPA Axis. Sci. Rep. 2018, 8, 6334. [Google Scholar] [CrossRef]
- Ishii-Yonemoto, T.; Masuzaki, H.; Yasue, S.; Okada, S.; Kozuka, C.; Tanaka, T.; Noguchi, M.; Tomita, T.; Fujikura, J.; Yuji Yamamoto, Y.; et al. Glucocorticoid reamplification within cells intensifies NF-κB and MAPK signaling and reinforces inflammation in activated preadipocytes. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E930–E940. [Google Scholar]
- Itoi, S.; Terao, M.; Murota, H.; Katayama, I. 11beta-Hydroxysteroid dehydrogenase 1 contributes to the pro-inflammatory response of keratinocytes. Biochem. Biophys. Res. Commun. 2013, 440, 265–270. [Google Scholar] [CrossRef]
- Cannarozzo, G.; Fazia, G.; Bennardo, L.; Tamburi, F.; Amoruso, G.F.; Del Duca, E.; Nistico, S.P. A New 675 nm Laser Device in the Treatment of Facial Aging: A Prospective Observational Study. Photobiomodul Photomed. Laser Surg. 2021, 39, 118–122. [Google Scholar] [CrossRef]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased collagen production in chronologically aged skin: Roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef]
- Lee, D.H.; Oh, J.H.; Chung, J.H. Glycosaminoglycan and proteoglycan in skin aging. J. Dermatol. Sci. 2016, 83, 174–181. [Google Scholar] [CrossRef]
- Weinmullner, R.; Zbiral, B.; Becirovic, A.; Stelzer, E.M.; Nagelreiter, F.; Schosserer, M.; Lammermann, I.; Liendl, L.; Lang, M.; Terlecki-Zaniewicz, L.; et al. Organotypic human skin culture models constructed with senescent fibroblasts show hallmarks of skin aging. npj Aging Mech. Dis. 2020, 6, 4. [Google Scholar] [CrossRef]
- Lago, J.C.; Puzzi, M.B. The effect of aging in primary human dermal fibroblasts. PLoS ONE 2019, 14, e0219165. [Google Scholar] [CrossRef]
- Lammermann, I.; Terlecki-Zaniewicz, L.; Weinmullner, R.; Schosserer, M.; Dellago, H.; de Matos Branco, A.D.; Autheried, D.; Sevcnikar, B.; Kleissl, L.; Berlin, I.; et al. Blocking negative effects of senescence in human skin fibroblasts with a plant extract. npj Aging Mech. Dis. 2018, 4, 4. [Google Scholar] [CrossRef]
- Kamiya, Y.; Odama, M.; Mizuguti, A.; Murakami, S.; Ito, T. Puerarin blocks the aging phenotype in human dermal fibroblasts. PLoS ONE 2021, 16, e0249367. [Google Scholar] [CrossRef]
- Yang, J.; Dungrawala, H.; Hua, H.; Manukyan, A.; Abraham, L.; Lane, W.; Mead, H.; Wright, J.; Schneider, B.L. Cell size and growth rate are major determinants of replicative lifespan. Cell Cycle 2011, 10, 144–155. [Google Scholar] [CrossRef]
- Zheng, Q.; Chen, S.; Chen, Y.; Lyga, J.; Wyborski, R.; Santhanam, U. Investigation of age-related decline of microfibril-associated glycoprotein-1 in human skin through immunohistochemistry study. Clin. Cosmet. Investig. Dermatol. 2013, 6, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Lv, Y.; Sun, Y.; Wang, Y.; Zhao, Z.; Shi, C.; Chen, X.; Wang, L.; Zhang, M.; Wei, B.; et al. The anti-inflammatory activity of specific-sized hyaluronic acid oligosaccharides. Carbohydr. Polym. 2022, 276, 118699. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Dias, M.; Madusanka, D.M.D.; Han, E.J.; Kim, M.J.; Heo, S.J.; Lee, K.; Cheong, S.H.; Ahn, G. Low molecular weight fucoidan fraction ameliorates inflammation and deterioration of skin barrier in fine-dust stimulated keratinocytes. Int. J. Biol. Macromol. 2021, 168, 620–630. [Google Scholar] [CrossRef]
- Shanura Fernando, I.P.; Asanka Sanjeewa, K.K.; Samarakoon, K.W.; Kim, H.-S.; Gunasekara, U.K.D.S.S.; Park, Y.-J.; Abeytunga, D.T.U.; Lee, W.W.; Jeon, Y.-J. The potential of fucoidans from Chnoospora minima and Sargassum polycystum in cosmetics: Antioxidant, anti-inflammatory, skin-whitening, and antiwrinkle activities. J. Appl. Phycol. 2018, 30, 3223–3232. [Google Scholar] [CrossRef]
- Zorina, A.; Zorin, V.; Isaev, A.; Kudlay, D.; Vasileva, M.; Kopnin, P. Dermal Fibroblasts as the Main Target for Skin Anti-Age Correction Using a Combination of Regenerative Medicine Methods. Curr. Issues Mol. Biol. 2023, 45, 3829–3847. [Google Scholar] [CrossRef] [PubMed]
- Langhans, S.A. Three-Dimensional in Vitro Cell Culture Models in Drug Discovery and Drug Repositioning. Front. Pharmacol. 2018, 9, 6. [Google Scholar] [CrossRef]
- Zhang, Z.; Michniak-Kohn, B.B. Tissue engineered human skin equivalents. Pharmaceutics 2012, 4, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Messaraa, C.; Metois, A.; Walsh, M.; Hurley, S.; Doyle, L.; Mansfield, A.; O’Connor, C.; Mavon, A. Wrinkle and roughness measurement by the Antera 3D and its application for evaluation of cosmetic products. Skin Res. Technol. 2018, 24, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Zhang, S.; Wang, Y.; Ni, J.; Zhao, T.; Xiao, G. Anti-skin aging effects and bioavailability of collagen tripeptide and elastin peptide formulations in young and middle-aged women. J. Dermatol. Sci. Cosmet. Technol. 2024, 1, 100019. [Google Scholar]
- Kong, R.; Cui, Y.; Fisher, G.J.; Wang, X.; Chen, Y.; Schneider, L.M.; Majmudar, G. A comparative study of the effects of retinol and retinoic acid on histological, molecular, and clinical properties of human skin. J. Cosmet. Dermatol. 2016, 15, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Tucker-Samaras, S.; Zedayko, T.; Cole, C.; Miller, D.; Wallo, W.; Leyden, J.J. A stabilized 0.1% retinol facial moisturizer improves the appearance of photodamaged skin in an eight-week, double-blind, vehicle-controlled study. J. Drugs Dermatol. 2009, 8, 932–936. [Google Scholar] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Ye, S.; Jun, S.-H.; Kang, N.-G. Efficacy of SGPP2 Modulation-Mediated Materials in Ameliorating Facial Wrinkles and Pore Sagging. Curr. Issues Mol. Biol. 2024, 46, 9122-9135. https://doi.org/10.3390/cimb46080539
Kim J, Ye S, Jun S-H, Kang N-G. Efficacy of SGPP2 Modulation-Mediated Materials in Ameliorating Facial Wrinkles and Pore Sagging. Current Issues in Molecular Biology. 2024; 46(8):9122-9135. https://doi.org/10.3390/cimb46080539
Chicago/Turabian StyleKim, Juhyun, Sanghyun Ye, Seung-Hyun Jun, and Nae-Gyu Kang. 2024. "Efficacy of SGPP2 Modulation-Mediated Materials in Ameliorating Facial Wrinkles and Pore Sagging" Current Issues in Molecular Biology 46, no. 8: 9122-9135. https://doi.org/10.3390/cimb46080539
APA StyleKim, J., Ye, S., Jun, S.-H., & Kang, N.-G. (2024). Efficacy of SGPP2 Modulation-Mediated Materials in Ameliorating Facial Wrinkles and Pore Sagging. Current Issues in Molecular Biology, 46(8), 9122-9135. https://doi.org/10.3390/cimb46080539