
Intranasal Insulin Eases Autism in Rats via GDF-15 and Anti-Inflammatory Pathways
 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Procedures
2.3. Behavioral Tests
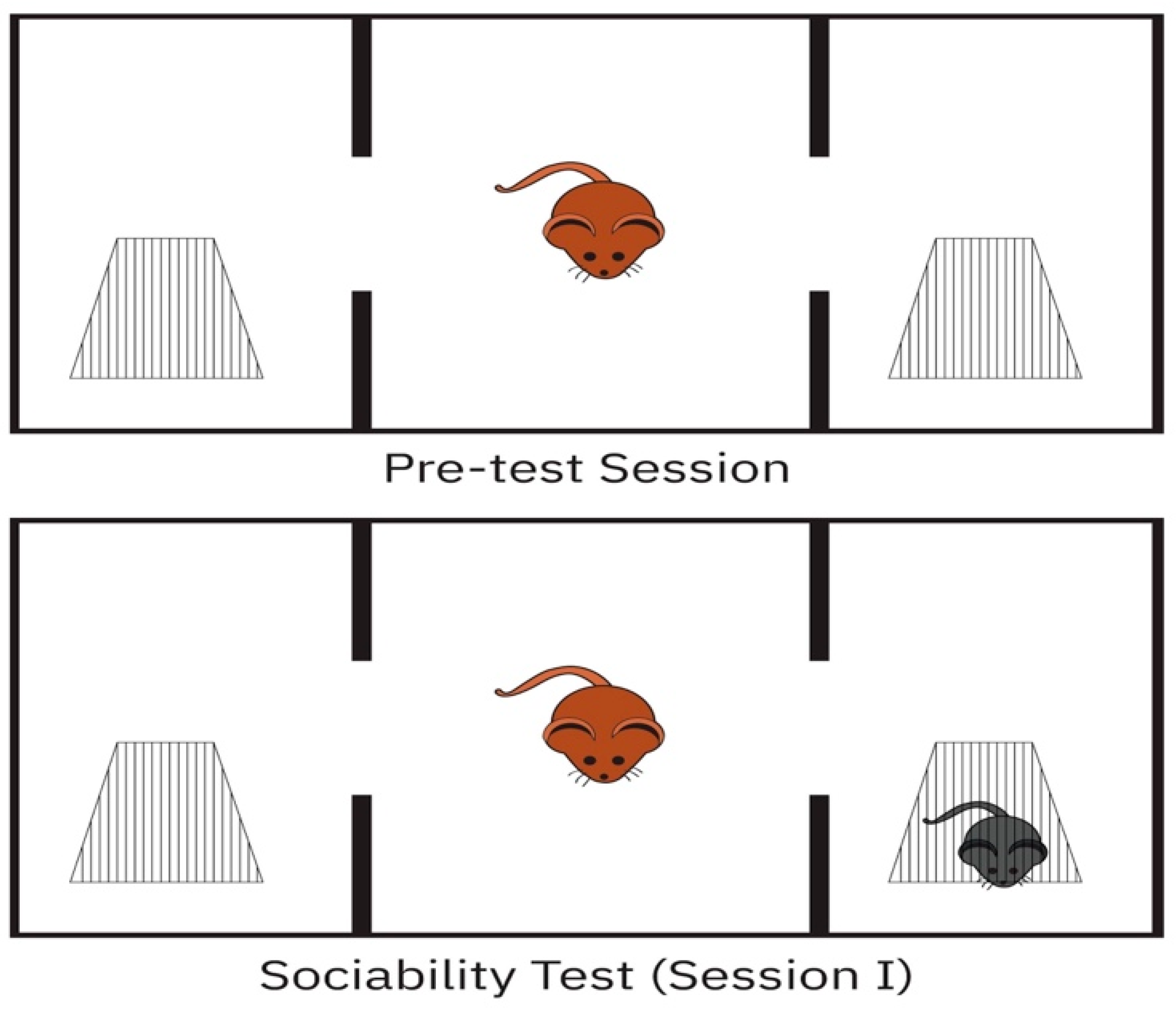
2.3.1. Three-Chamber Sociability Test
2.3.2. Open-Field
2.3.3. Passive Avoidance Learning (PAL)
2.4. Hippocampus and Cerebellum Histopathology
2.5. Brain Biochemical Analysis
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Measurement of Brain Lipid Peroxidation (MDA)
2.8. Measurement of Brain Protein Levels
2.9. Statistical Analysis
3. Results
3.1. Biochemical Investigations of Brain Tissue
3.2. Behavioural Tests
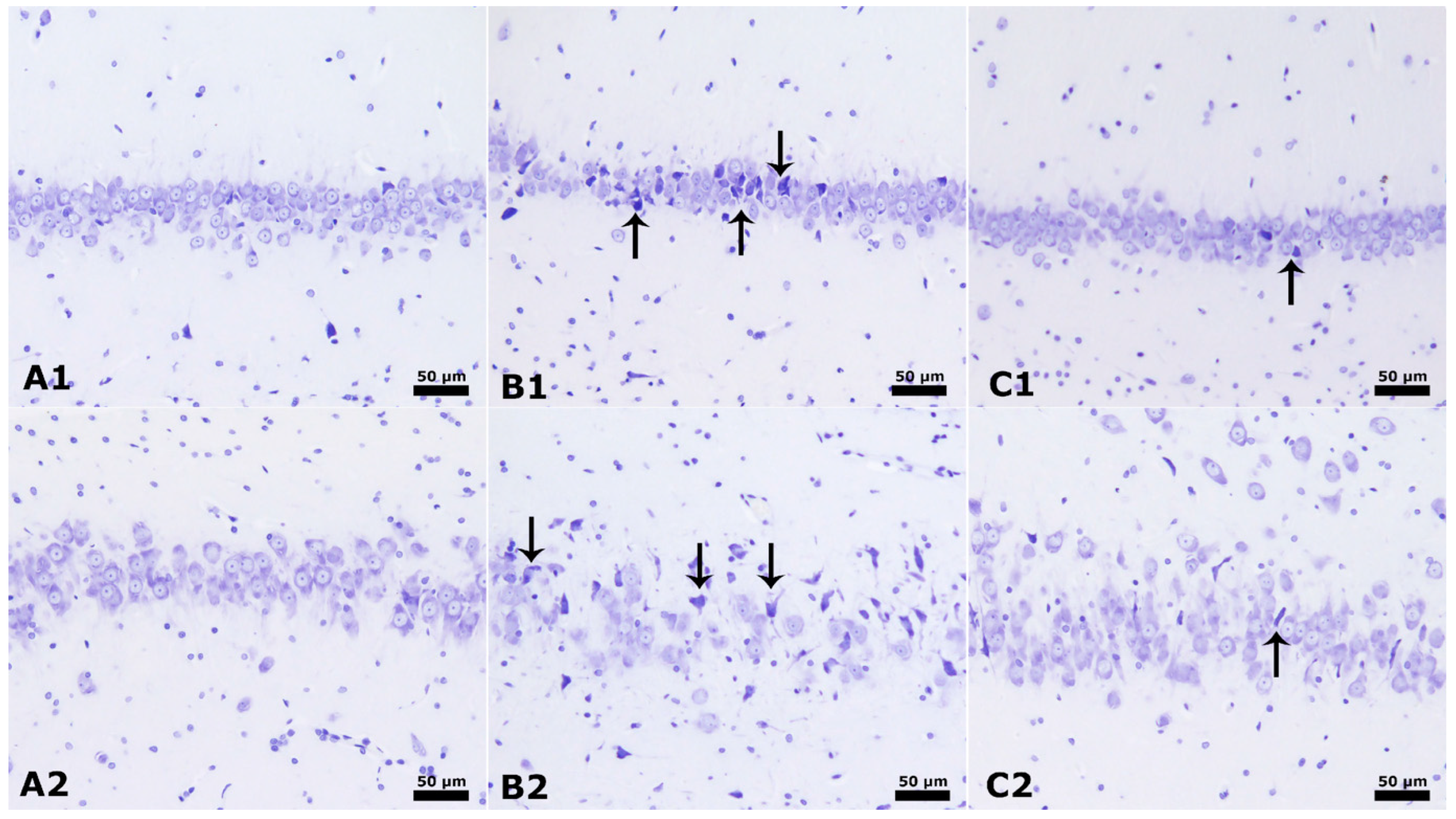
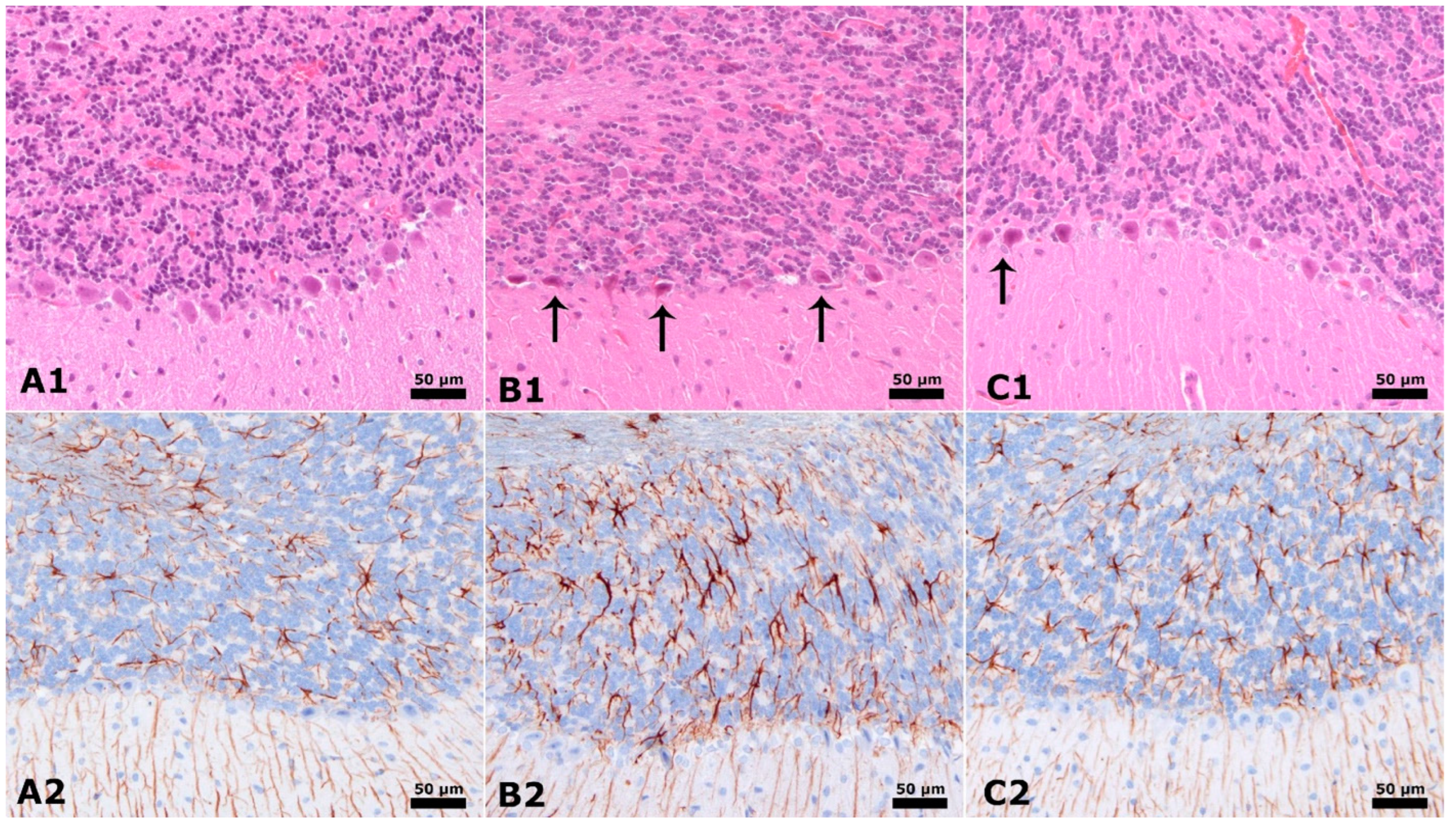
3.3. Neuronal Counts and GLIAL Fibrillary Acidic Proteins
3.4. Hippocampus and Cerebellum Histopathology Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Genovese, A.; Butler, M.G. The autism spectrum: Behavioural, psychiatric and genetic associations. Genes 2023, 14, 677. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; King, B.H. Autism spectrum disorder: A review. JAMA 2023, 329, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Shenouda, J.; Barrett, E.; Davidow, A.L.; Sidwell, K.; Lescott, C.; Halperin, W.; Silenzio, V.; Zahorodny, W. Prevalence and disparities in the detection of autism without intellectual disability. Pediatrics 2023, 151, e2022056594. [Google Scholar] [CrossRef]
- Byres, L.; Morris, E.; Austin, J. Exploring Autistic adults’ perspectives on genetic testing for autism. Genet. Med. 2023, 25, 100021. [Google Scholar] [CrossRef]
- Yasuda, Y.; Matsumoto, J.; Miura, K.; Hasegawa, N.; Hashimoto, R. Genetics of autism spectrum disorders and future direction. J. Hum. Genet. 2023, 68, 193–197. [Google Scholar] [CrossRef]
- Joon, P.; Kumar, A.; Parle, M. The Resemblance between the Propionic Acid Rodent Model of Autism and Autism Spectrum Disorder. J. Psych. Neuroche Res. 2023, 1, 8. [Google Scholar]
- Bagcioglu, E.; Solmaz, V.; Erbas, O.; Özkul, B.; Çakar, B.; Uyanikgil, Y.; Söğüt, İ. Modafinil improves autism-like behavior in rats by reducing neuroinflammation. J. Neuroimmune Pharmacol. 2023, 18, 9–23. [Google Scholar] [CrossRef]
- Lagod, P.P.; Abdelli, L.S.; Naser, S.A. An In Vivo Model of Propionic Acid-Rich Diet-Induced Gliosis and Neuro-Inflammation in Mice (FVB/N-Tg (GFAPGFP) 14Mes/J): A Potential Link to Autism Spectrum Disorder. Int. J. Mol. Sci. 2024, 25, 8093. [Google Scholar] [CrossRef]
- Meeking, M.M.; MacFabe, D.F.; Mepham, J.R.; Foley, K.A.; Tichenoff, L.J.; Boon, F.H.; Kavaliers, M.; Ossenkopp, K.P. Propionic acid induced behavioural effects of relevance to autism spectrum disorder evaluated in the hole board test with rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 97, 109794. [Google Scholar] [CrossRef]
- Sharma, A.R.; Batra, G.; Saini, L.; Sharma, S.; Mishra, A.; Singla, R.; Singh, A.; Singh, R.S.; Jain, A.; Bansal, S.; et al. Valproic acid and propionic acid modulated mechanical pathways associated with autism spectrum disorder at prenatal and neonatal exposure. CNS Neurol. Disord.-Drug Targets 2022, 21, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Aishworiya, R.; Valica, T.; Hagerman, R.; Restrepo, B. An update on psychopharmacological treatment of autism spectrum disorder. Neurotherapeutics 2023, 19, 248–262. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.F.; Brubaker, P.L. The discovery of insulin revisited: Lessons for the modern era. J. Clin. Investig. 2021, 131, e142239. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Hossain, K.S.; Das, S.; Kundu, S.; Adegoke, E.O.; Rahman, M.A.; Hannan, M.A.; Uddin, M.J.; Pang, M.G. Role of insulin in health and disease: An update. Int. J. Mol. Sci. 2021, 22, 6403. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Cui, J.; He, Q.; Chen, Z.; Arvan, P.; Liu, M. Proinsulin misfolding and endoplasmic reticulum stress during the development and progression of diabetes. Mol. Asp. Med. 2015, 42, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.E.; Newgard, C.B. Mechanisms controlling pancreatic islet cell function in insulin secretion. Nat. Rev. Mol. Cell Biol. 2021, 22, 142–158. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Mechanisms of insulin action and insulin resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef]
- Hallschmid, M. Intranasal insulin. J. Neuroendocrinol. 2021, 33, e12934. [Google Scholar] [CrossRef]
- Zhao, L.; Teter, B.; Morihara, T.; Lim, G.P.; Ambegaokar, S.S.; Ubeda, O.J.; Frautschy, S.A.; Cole, G.M. Insulin-degrading enzyme as a downstream target of insulin receptor signaling cascade: Implications for Alzheimer’s disease intervention. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 11120–11126. [Google Scholar] [CrossRef]
- Banks, W.A.; Owen, J.B.; Erickson, M.A. Insulin in the brain: There and back again. Pharmacol. Ther. 2012, 136, 82–93. [Google Scholar] [CrossRef]
- Hallschmid, M. Intranasal insulin for Alzheimer’s disease. CNS Drugs 2021, 35, 21–37. [Google Scholar] [CrossRef]
- Edgerton, D.S.; Kraft, G.; Smith, M.; Farmer, B.; Williams, P.E.; Coate, K.C.; Printz, R.L.; O’Brien, R.M.; Cherrington, A.D. Insulin’s direct hepatic effect explains the inhibition of glucose production caused by insulin secretion. JCI Insight 2017, 2, e91863. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Zabolotny, J.M.; Huang, H.; Lee, H.; Kim, Y.B. Insulin in the nervous system and the mind: Functions in metabolism, memory, and mood. Mol. Metab. 2016, 5, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Zwanenburg, R.J.; Bocca, G.; Ruiter, S.A.; Dillingh, J.H.; Flapper, B.C.; van den Heuvel, E.R.; van Ravenswaaij-Arts, C.M. Is there an effect of intranasal insulin on development and behaviour in Phelan-McDermid syndrome? A randomized, double-blind, placebo-controlled trial. Eur. J. Hum. Genet. EJHG 2016, 24, 1696–1701. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Yuan, L.J.; Yang, Y.; Zhang, M.; Chen, W.F. IGF-1 inhibits MPTP/MPP+-induced autophagy on dopaminergic neurons through the IGF-1R/PI3K-Akt-mTOR pathway and GPER. Am. J. Physiol.-Endocrinol. Metab. 2020, 319, E734–E743. [Google Scholar] [CrossRef] [PubMed]
- Bou Khalil, R. Is insulin growth factor-1 the future for treating autism spectrum disorder and/or schizophrenia? Med. Hypotheses 2017, 99, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Vahdatpour, C.; Dyer, A.H.; Tropea, D. Insulin-Like Growth Factor 1 and Related Compounds in the Treatment of Childhood-Onset Neurodevelopmental Disorders. Front. Neurosci. 2016, 10, 450. [Google Scholar] [CrossRef]
- Riikonen, R. Treatment of autistic spectrum disorder with insulin-like growth factors. Eur. J. Paediatr. Neurol. EJPN Off. J. Eur. Paediatr. Neurol. Soc. 2016, 20, 816–823. [Google Scholar] [CrossRef]
- Fine, J.M.; Stroebel, B.M.; Faltesek, K.A.; Terai, K.; Haase, L.; Knutzen, K.E.; Kosyakovsky, J.; Bowe, T.J.; Fuller, A.K.; Frey, W.H.; et al. Intranasal delivery of low-dose insulin ameliorates motor dysfunction and dopaminergic cell death in a 6-OHDA rat model of Parkinson’s Disease. Neurosci. Lett. 2020, 714, 134567. [Google Scholar] [CrossRef]
- Iravanpour, F.; Dargahi, L.; Rezaei, M.; Haghani, M.; Heidari, R.; Valian, N.; Ahmadiani, A. Intranasal insulin improves mitochondrial function and attenuates motor deficits in a rat 6-OHDA model of Parkinson’s disease. CNS Neurosci. Ther. 2021, 27, 308–319. [Google Scholar] [CrossRef]
- Doğan, M.; Albayrak, Y.; Erbaş, O. Torasemide Improves the Propionic Acid-Induced Autism in Rats: A Histopathological and Imaging Study. Alpha Psychiatry 2023, 24, 22–31. [Google Scholar] [CrossRef]
- Erdogan, M.A.; Bozkurt, M.F.; Erbas, O. Effects of prenatal testosterone exposure on the development of autism-like behaviours in offspring of Wistar rats. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 2023, 83, 201–215. [Google Scholar] [CrossRef]
- Karakaş, L.; Solmaz, V.; Bağcıoğlu, E.; Ozkul, B.; Söğüt, İ.; Uyanıkgil, Y.; Erbaş, O. Lowering propionic acid levels by regulating gut microbiota with ursodeoxycholic acid appears to regress autism symptoms: An animal study. J. Health Sci. Med./JHSM 2023, 6, 791–799. [Google Scholar] [CrossRef]
- Bozkurt, M.F.; Bhaya, M.N.; Sever, İ.H.; Özkul, B.; Erbaş, O. Medroxyprogesterone acetate improves propionic acid-induced autism rat model and magnetic resonance spectroscopic correlation. Bangladesh J. Pharmacol. 2022, 17, 56–65. [Google Scholar] [CrossRef]
- Jiang, L.H.; Zhang, Y.N.; Wu, X.W.; Song, F.F.; Guo, D.Y. Effect of insulin on the cognizing function and expression of hippocampal Aβ1–40 of rat with Alzheimer disease. Chin. Med. J. 2008, 121, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Furubayashi, T.; Arai, M.; Inoue, D.; Kimura, S.; Kiriyama, A.; Kusamori, K.; Katsumi, H.; Yutani, R.; Sakane, T.; et al. Delivery of Oxytocin to the Brain for the Treatment of Autism Spectrum Disorder by Nasal Application. Mol. Pharm. 2018, 15, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Yatawara, C.; Einfeld, S.; Hickie, I.; Davenport, T.; Guastella, A. The effect of oxytocin nasal spray on social interaction deficits observed in young autistic children: A randomized clinical crossover trial. Mol. Psychiatry 2015, 21, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Li, X.; Zhong, Y. Inflammatory cytokines: Potential biomarkers of immunologic dysfunction in autism spectrum disorders. Mediat. Inflamm. 2015, 2015, 531518. [Google Scholar] [CrossRef]
- Vanhala, R.; Turpeinen, U.; Riikonen, R. Low levels of insulin-like growth factor-I in cerebrospinal fluid in children with autism. Dev. Med. Child Neurol. 2001, 43, 614–616. [Google Scholar]
- Manco, M.; Guerrera, S.; Ravà, L.; Ciofi Degli Atti, M.; Di Vara, S.; Valeri, G.; Vicari, S. Cross-sectional investigation of insulin resistance in youths with autism spectrum disorder. Any role for reduced brain glucose metabolism? Transl. Psychiatry 2021, 11, 229. [Google Scholar] [CrossRef]
- Altun, H.; Şahin, N.; Kurutaş, E.B.; Karaaslan, U.; Sevgen, F.H.; Fındıklı, E. Assessment of malondialdehyde levels, superoxide dismutase, and catalase activity in children with autism spectrum disorders. Psychiatry Clin. Psychopharmacol. 2018, 28, 408–415. [Google Scholar] [CrossRef]
- Meguid, N.A.; Dardir, A.A.; Abdel-Raouf, E.R.; Hashish, A. Evaluation of Oxidative Stress in Autism: Defective Antioxidant Enzymes and Increased Lipid Peroxidation. Biol. Trace Elem. Res. 2011, 143, 58–65. [Google Scholar] [CrossRef]
- Ciafrè, S.; Carito, V.; Ferraguti, G.; Greco, A.; Ralli, M.; Aloe, G.I.A.N.; Chaldakov, G.; Messina, M.P.; Attilia, M.L.; Ceccarelli, R.; et al. Nerve growth factor in brain diseases. Biomed. Rev. 2018, 29, 1–16. [Google Scholar] [CrossRef]
- Havrankova, J.; Schmechel, D.; Roth, J.; Brownstein, M. Identification of insulin in rat brain. Proc. Natl. Acad. Sci. USA 1978, 75, 5737–5741. [Google Scholar] [CrossRef] [PubMed]
- Mielke, J.G.; Nicolitch, K.; Avellaneda, V.; Earlam, K.; Ahuja, T.; Mealing, G.; Messier, C. Longitudinal study of the effects of a high-fat diet on glucose regulation, hippocampal function, and cerebral insulin sensitivity in C57BL/6 mice. Behav. Brain Res. 2006, 175, 374–382. [Google Scholar] [CrossRef]
- Wan, Q.; Xiong, Z.G.; Man, H.Y.; Ackerley, C.A.; Braunton, J.; Lu, W.Y.; Becker, L.E.; MacDonald, J.F.; Wang, Y.T. Recruitment of functional GABAA receptors to postsynaptic domains by insulin. Nature 1997, 388, 686–690. [Google Scholar] [CrossRef] [PubMed]
- Beattie, J.; Allan, G.J.; Lochrie, J.D.; Flint, D.J. Insulin-like growth factor-binding protein-5 (IGFBP-5): A critical member of the IGF axis. Biochem. J. 2006, 395, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Plum, L.; Schubert, M.; Brüning, J.C. The role of insulin receptor signaling in the brain. Trends Endocrinol. Metab. 2005, 16, 59–65. [Google Scholar] [CrossRef]
- Wischhusen, J.; Melero, I.; Fridman, W.H. Growth/differentiation factor-15 (GDF-15): From biomarker to novel targetable immune checkpoint. Front. Immunol. 2020, 11, 951. [Google Scholar] [CrossRef]
- Than, U.T.T.; Nguyen, L.T.; Nguyen, P.H.; Nguyen, X.H.; Trinh, D.P.; Hoang, D.H.; Nguyen, P.A.T.; Dang, V.D. Inflammatory mediators drive neuroinflammation in autism spectrum disorder and cerebral palsy. Sci. Rep. 2023, 13, 22587. [Google Scholar] [CrossRef]
- Iglesias, P.; Silvestre, R.A.; Díez, J.J. Growth differentiation factor 15 (GDF-15) in endocrinology. Endocrine 2023, 81, 419–431. [Google Scholar] [CrossRef]
- Marques, S.S.; Liebaug, A.; Maurer, S.; Rothenbacher, D.; Brenner, R.E.; Riegger-Koch, J. p53-induced GDF-15 expression promotes a pro-regenerative response in human chondrocytes upon cartilage injury. bioRxiv 2024. [Google Scholar] [CrossRef]
- Abdelhak, A.; Foschi, M.; Abu-Rumeileh, S.; Yue, J.K.; D’Anna, L.; Huss, A.; Oeckl, P.; Ludolph, A.C.; Kuhle, J.; Petzold, A.; et al. Blood GFAP as an emerging biomarker in brain and spinal cord disorders. Nat. Rev. Neurol. 2022, 18, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lee, S.; Won, J.; Jin, Y.; Hong, Y.; Hur, T.Y.; Kim, J.H.; Lee, S.R.; Hong, Y. Pathophysiological and neurobehavioral characteristics of a propionic acid-mediated autism-like rat model. PLoS ONE 2018, 13, e0192925. [Google Scholar]
- Abdelli, L.S.; Samsam, A.; Naser, S.A. Propionic Acid Induces Gliosis and Neuro-inflammation through Modulation of PTEN/AKT Pathway in Autism Spectrum Disorder. Sci. Rep. 2019, 9, 8824. [Google Scholar] [CrossRef]
- Stern, M. Insulin Signaling and Autism. Front. Endocrinol. 2011, 2, 54. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 Normal Control | Group 2 PPA + Saline | Group 3 PPA + Insulin | |
|---|---|---|---|
| Brain MDA level (nmol/gr protein) | 50.1 ± 0.9 | 164.5 ± 9.5 ** | 97.1 ± 8.2 ## |
| Brain TNF-alfa level (pg/mg protein) | 12.5 ± 1.1 | 196.1 ± 12.3 ** | 91.8 ± 7.7 ## |
| Brain IL-2 level (pg/mg protein) | 2.1 ± 0.2 | 215.2 ± 14.1 ** | 85.1 ± 9.3 ## |
| Brain IL-17 level (pg/mg protein) | 195.3 ± 20.1 | 524.5 ± 17.4 * | 412.7 ± 10.6 # |
| Brain GDF-15 level (pg/mg protein) | 13.08 ± 0.6 | 19.3 ± 0.8 ** | 24.05 ± 1.1 # |
| Brain NGF level (pg/mg protein) | 78.3 ± 2.4 | 39.4 ± 3.1 ** | 66.3 ± 5.4 # |
| Group 1 Normal Control | Group 2 PPA + Saline | Group 3 PPA + Insulin | |
|---|---|---|---|
| Sociability test: The percentage (%) of time spent with a stranger rat | 65.9 ± 3.5 | 29.4 ± 2.2 * | 63.8 ± 3.5 ## |
| Open Field Test: Amount of ambulation | 17.6 ± 2.5 | 8.3 ± 1.9 * | 15.4 ± 2.1 # |
| Passive avoidance learning (PAL) Latency (Sec.) | 266.4 ± 19.5 | 85.4 ± 20.5 * | 214.5 ± 16.2 ## |
| Group 1 Normal Control | Group 2 PPA + Saline | Group 3 PPA + Insulin | |
|---|---|---|---|
| Neuronal Count CA1 | 81.2 ± 3.9 | 65.3 ± 4.8 ** | 77.9 ± 1.8 ## |
| Neuronal Count CA3 | 44.3 ± 3.7 | 34.5 ± 3.1 ** | 39.8 ± 6.3 # |
| GFAP immunostaining index (CA1) | 35.8 ± 1.4 | 46.9 ± 4.06 * | 41.7 ± 0.8 # |
| GFAP immunostaining index (CA3) | 33.3 ± 0.9 | 42.1 ± 2.3 * | 35.7 ± 1.5 # |
| Purkinje Count Cerebellum | 21.7 ± 1.3 | 15.2 ± 0.8 * | 18.6 ± 1.2 # |
| GFAP immunostaining index (Cerebellum) | 21.4 ± 2.3 | 27.1 ± 2.9 * | 22.5 ± 1.8 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arda, D.B.; Tunç, K.C.; Bozkurt, M.F.; Bora, E.S.; Çiğel, A.; Erbaş, O. Intranasal Insulin Eases Autism in Rats via GDF-15 and Anti-Inflammatory Pathways. Curr. Issues Mol. Biol. 2024, 46, 10530-10544. https://doi.org/10.3390/cimb46090624
Arda DB, Tunç KC, Bozkurt MF, Bora ES, Çiğel A, Erbaş O. Intranasal Insulin Eases Autism in Rats via GDF-15 and Anti-Inflammatory Pathways. Current Issues in Molecular Biology. 2024; 46(9):10530-10544. https://doi.org/10.3390/cimb46090624
Chicago/Turabian StyleArda, Duygu Burcu, Kerem Can Tunç, Mehmet Fatih Bozkurt, Ejder Saylav Bora, Ayşe Çiğel, and Oytun Erbaş. 2024. "Intranasal Insulin Eases Autism in Rats via GDF-15 and Anti-Inflammatory Pathways" Current Issues in Molecular Biology 46, no. 9: 10530-10544. https://doi.org/10.3390/cimb46090624
APA StyleArda, D. B., Tunç, K. C., Bozkurt, M. F., Bora, E. S., Çiğel, A., & Erbaş, O. (2024). Intranasal Insulin Eases Autism in Rats via GDF-15 and Anti-Inflammatory Pathways. Current Issues in Molecular Biology, 46(9), 10530-10544. https://doi.org/10.3390/cimb46090624