
Biosynthesis and Extraction of Chlorophyll, Carotenoids, Anthocyanins, and Betalaine In Vivo and In Vitro

Abstract
1. Introduction
2. Extraction of Plant Pigments
2.1. Liquid–Liquid Extraction or Solid–Liquid Extraction
2.2. Supercritical Fluid [Extraction], SFE
2.3. Correct to Ultrasound-Assisted Extraction, UAE
2.4. Microwave-Assisted Extraction, MAE
3. Biosynthesis of Plant Pigments
3.1. Biosynthesis Pathways of Plant Pigments
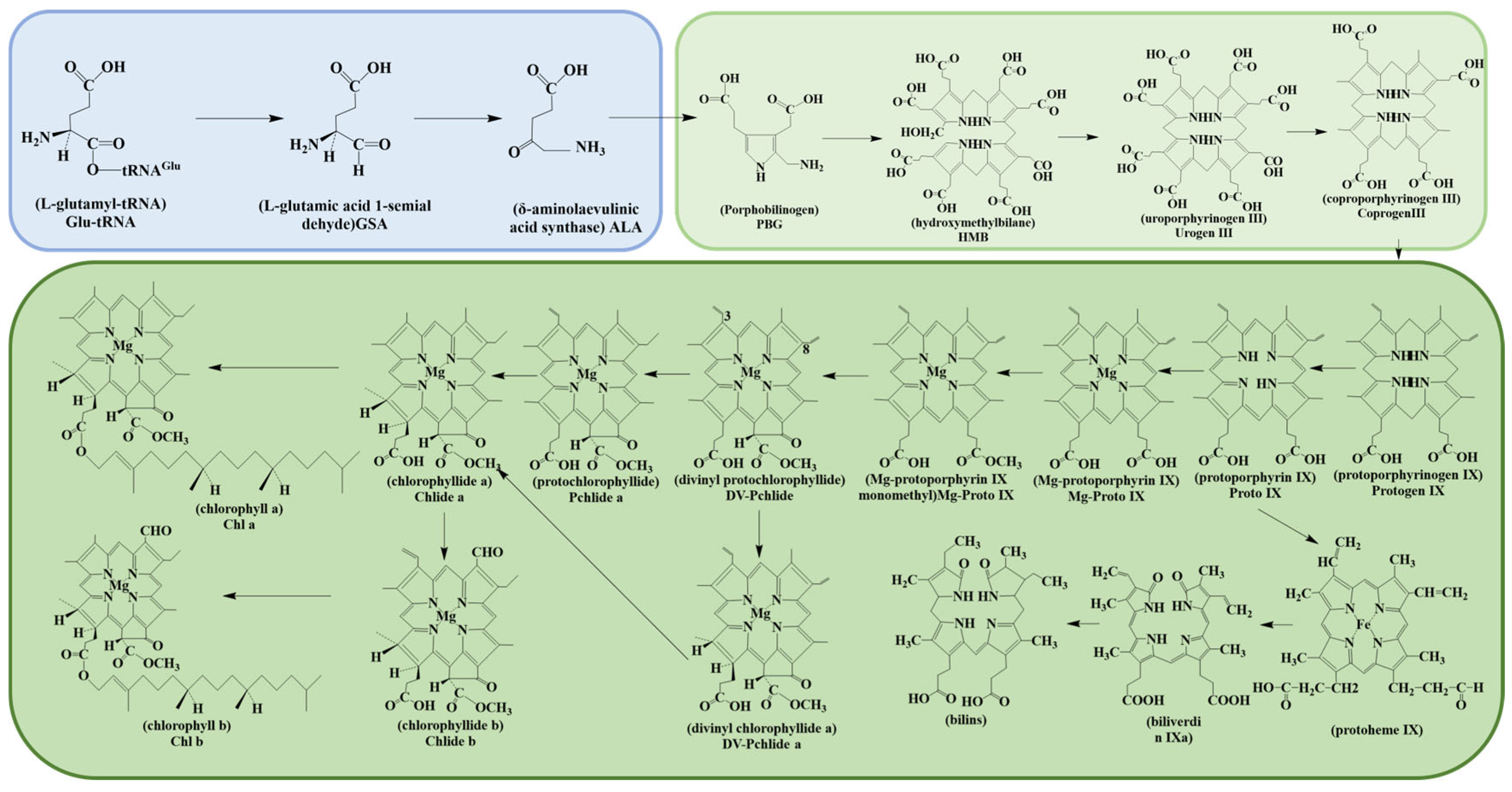
3.1.1. Chlorophyll
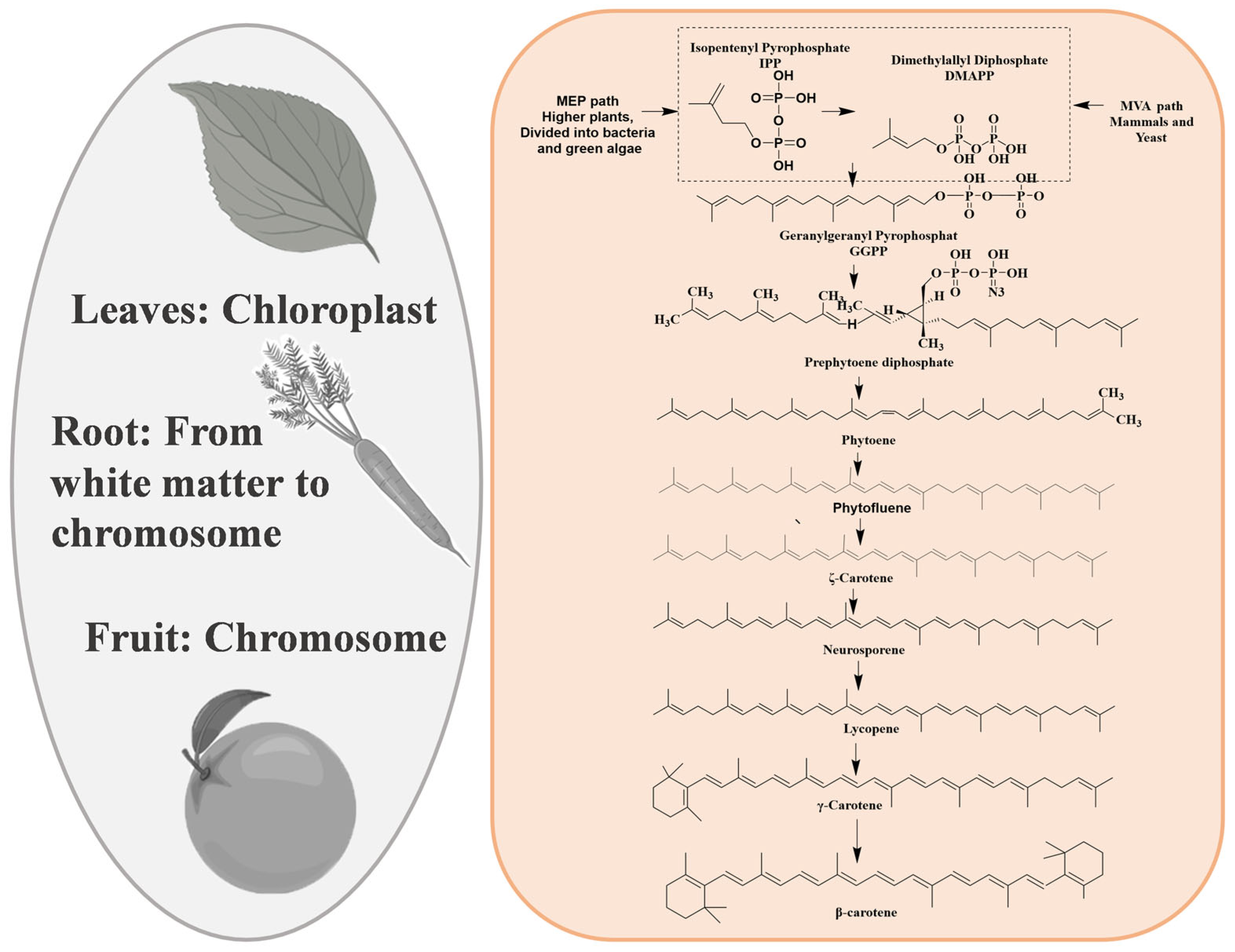
3.1.2. Carotenoids
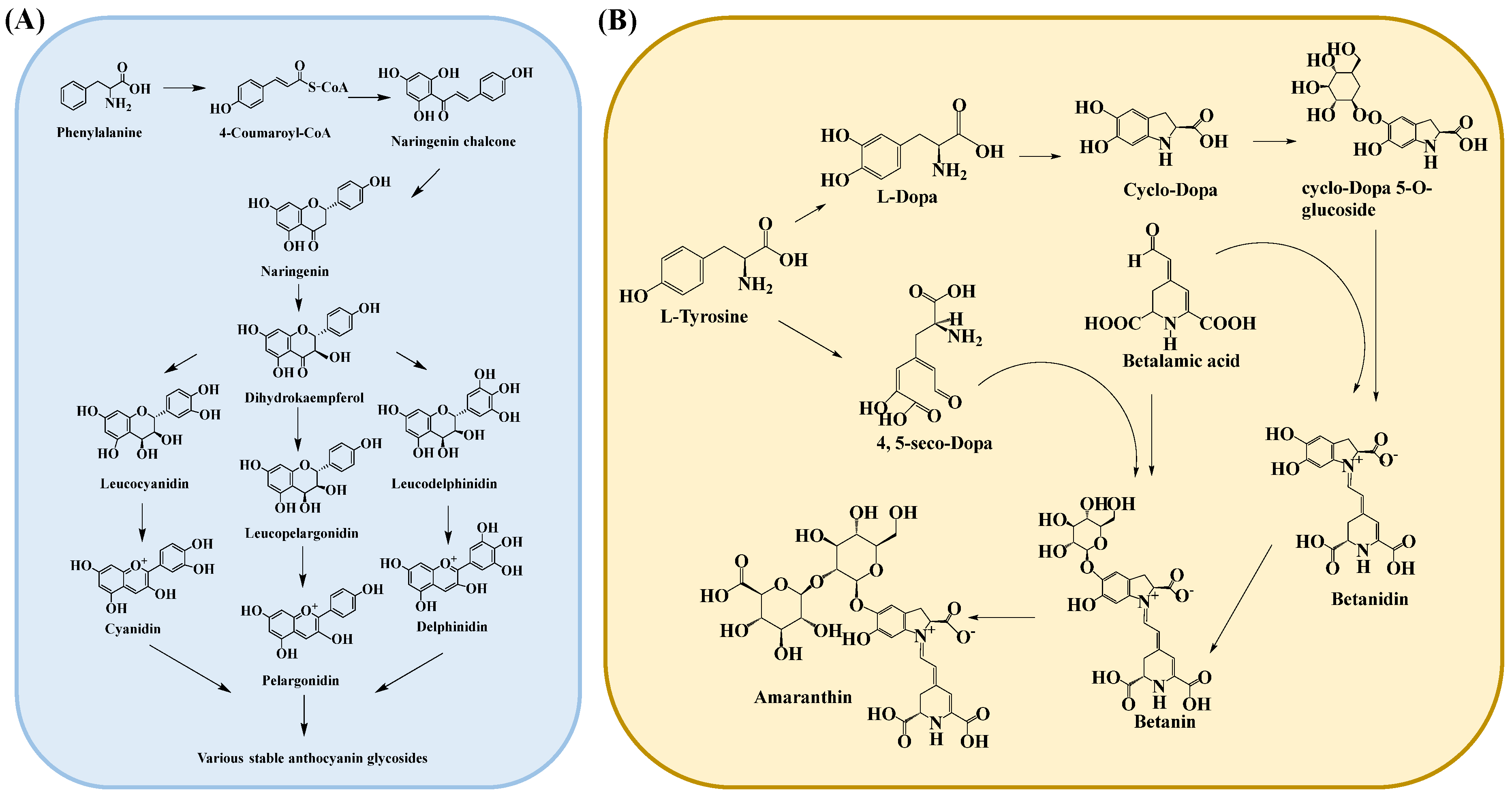
3.1.3. Anthocyanins
3.1.4. Betalaine
3.2. Heterologous Synthesis of Plant Pigments
3.2.1. Chlorophyll Synthesis In Vitro
3.2.2. Carotenoids Synthesis In Vitro
3.2.3. Anthocyanins Synthesis In Vitro
3.2.4. Betalaine Synthesis In Vitro
4. Improvement in Production of Plant Pigments
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Chen, C.X. Overview of Plant Pigments. In Pigments in Fruits and Vegetables: Genomics and Dietetics; Chen, C., Ed.; Springer: New York, NY, USA, 2015; pp. 1–7. [Google Scholar]
- Kamble, P.N.; Giri, S.P.; Mane, R.S.; Tiwana, A. Estimation of chlorophyll content in young and adult leaves of some selected plants. Univers. J. Environ. Res. Technol. 2015, 5, 306–310. [Google Scholar]
- Ali, K.A.; Noraldeen, S.S.; Yaseen, A.A. An Evaluation Study for Chlorophyll Estimation Techniques. Sarhad J. Agric. 2021, 37, 1458–1465. [Google Scholar] [CrossRef]
- Fernandes, A.S.; do Nascimento, T.C.; Jacob-Lopes, E.; De Rosso, V.V.; Zepka, L.Q. Carotenoids: A brief overview on its structure, biosynthesis, synthesis, and applications. Prog. Carotenoid Res. 2018, 1, 1–17. [Google Scholar]
- Zakar, T.; Laczko-Dobos, H.; Toth, T.N.; Gombos, Z. Carotenoids Assist in Cyanobacterial Photosystem II Assembly and Function. Front. Plant Sci. 2016, 7, 295. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in Human Nutrition and Health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Karak, P. Biological Activities of Flavonoids: An Overview. Int. J. Pharm. Sci. Res. 2019, 10, 1567–1574. [Google Scholar]
- Saati, E.A. Antioxidant Power of Rose Anthocyanin Pigment. J. Eng. Appl. Sci. 2016, 11, 1201–1204. [Google Scholar]
- Sokolova, D.V.; Shvachko, N.A.; Mikhailova, A.S.; Popov, V.S.; Solovyeva, A.E.; Khlestkina, E.K. Characterization of Betalain Content and Antioxidant Activity Variation Dynamics in Table Beets (Beta vulgaris L.) with Differently Colored Roots. Agronomy 2024, 14, 999. [Google Scholar] [CrossRef]
- Sakuta, M. Diversity in plant red pigments: Anthocyanins and betacyanins. Plant Biotechnol. Rep. 2014, 8, 37–48. [Google Scholar] [CrossRef]
- Nabi, B.G.; Mukhtar, K.; Ahmed, W.; Manzoor, M.F.; Ranjha, M.M.A.N.; Kieliszek, M.; Bhat, Z.F.; Aadil, R.M. Natural pigments: Anthocyanins, carotenoids, chlorophylls, and betalains as colorants in food products. Food Biosci. 2023, 52. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant Flavonoids: Classification, Distribution, Biosynthesis, and Antioxidant Activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ahammed, G.J. Plant Stress Response and Adaptation via Anthocyanins: A Review. Plant Stress. 2023, 10. [Google Scholar] [CrossRef]
- Lu, X.; Li, W.; Wang, Q.; Wang, J.; Qin, S. Progress on the Extraction, Separation, Biological Activity, and Delivery of Natural Plant Pigments. Molecules 2023, 28, 5364. [Google Scholar] [CrossRef] [PubMed]
- Azmin, S.N.H.M.; Sulaiman, N.S.; Nor, M.S.M.; Abdullah, P.S.; Kari, Z.A.; Pati, S. A Review on Recent Advances on Natural Plant Pigments in Foods: Functions, Extraction, Importance and Challenges. Appl. Biochem. Biotechnol. 2022, 194, 4655–4672. [Google Scholar] [CrossRef] [PubMed]
- Lyu, X.; Lyu, Y.; Yu, H.; Chen, W.; Ye, L.; Yang, R. Biotechnological Advances for Improving Natural Pigment Production: A State-of-the-Art Review. Bioresour. Bioprocess. 2022, 9, 8. [Google Scholar] [CrossRef]
- Rodríguez-Mena, A.; Ochoa-Martínez, L.A.; González-Herrera, S.M.; Rutiaga-Quiñones, O.M.; González-Laredo, R.F.; Olmedilla-Alonso, B. Natural Pigments of Plant Origin: Classification, Extraction and Application in Foods. Food Chem. 2023, 398, 133908. [Google Scholar] [CrossRef]
- Kumar, G.; Upadhyay, S.; Yadav, D.K.; Malakar, S.; Dhurve, P.; Suri, S. Application of Ultrasound Technology for Extraction of Color Pigments from Plant Sources and their Potential Bio-Functional Properties: A review. J. Food Process. Eng. 2023, 46. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; Rajauria, G.; Tiwari, B. Conventional Extraction Techniques: Solvent Extraction. In Sustainable Seaweed Technologies; Elsevier: Amsterdam, The Netherlands, 2020; pp. 171–189. [Google Scholar]
- Ngamwonglumlert, L.; Devahastin, S.; Chiewchan, N. Molecular Structure, Stability and Cytotoxicity of Natural Green Colorants Produced from Centella asiatica L. Leaves Treated by Steaming and Metal Complexations. Food Chem. 2017, 232, 387–394. [Google Scholar] [CrossRef]
- López-Cruz, R.; Sandoval-Contreras, T.; Iñiguez-Moreno, M. Plant Pigments: Classification, Extraction, and Challenge of Their Application in the Food Industry. Food Bioprocess. Technol. 2023, 16, 2725–2741. [Google Scholar] [CrossRef]
- Ngamwonglumlert, L.; Devahastin, S.; Chiewchan, N. Natural Colorants: Pigment Stability and Extraction Yield En-hancement Via Utilization of Appropriate Pretreatment and Extraction Methods. Crit. Rev. Food Sci. Nutr. 2017, 57, 3243–3259. [Google Scholar] [CrossRef]
- Linares, G.; Rojas, M.L. Ultrasound-Assisted Extraction of Natural Pigments from Food Processing by-Products: A Review. Front. Nutr. 2022, 9, 891462. [Google Scholar] [CrossRef] [PubMed]
- Novais, C.; Molina, A.K.; Abreu, R.M.V.; Santo-Buelga, C.; Ferreira, I.C.F.R.; Pereira, C.; Barros, L. Natural Food Colorants and Preservatives: A Review, a Demand, and a Challenge. J. Agric. Food Chem. 2022, 70, 2789–2805. [Google Scholar] [CrossRef] [PubMed]
- Didion, Y.P.; Tjalsma, T.G.; Su, Z.; Malankowska, M.; Pinelo, M. What Is Next? The Greener Future of Solid Liquid Extraction of Biobased Compounds: Novel Techniques and Solvents Overpower Traditional Ones. Sep. Purif. Technol. 2023, 320. [Google Scholar] [CrossRef]
- Joseph, J.A.; Denisova, N.A.; Arendash, G.; Gordon, M.; Diamond, D.; Shukitt-Hale, B.; Morgan, D. Blueberry Supplementation Enhances Signaling and Prevents Behavioral Deficits in an Alzheimer Disease Model. Nutr. Neurosci. 2003, 3, 153–162. [Google Scholar] [CrossRef]
- Butnariu, M. Methods of Analysis (Extraction, Separation, Identification and Quantification) of Carotenoids from Natural Products. J. Ecosyst. Ecography 2016, 6, 193. [Google Scholar] [CrossRef]
- Liberal, Â.; Molina, A.K.; Pereira, C.; Dias, M.I.; Ferreira, I.C.; Barros, L. Solid-Liquid Extraction of Polyphenols. In Technologies to Recover Polyphenols from AgroFood By-Products and Wastes; Academic Press: Cambridge, MA, USA, 2022; pp. 73–112. [Google Scholar]
- Tzanova, M.T.; Yaneva, Z.; Ivanova, D.; Toneva, M.; Grozeva, N.; Memdueva, N. Green Solvents for Extraction of Natural Food Colorants from Plants: Selectivity and Stability Issues. Foods 2024, 13, 605. [Google Scholar] [CrossRef]
- Ashenafi, E.L.; Nyman, M.C.; Shelley, J.T.; Mattson, N.S. Spectral Properties and Stability of Selected Carotenoid and Chlorophyll Compounds in Different Solvent Systems. Food Chem. Advances. 2023, 2, 100178. [Google Scholar] [CrossRef]
- Morón-Ortiz, A.; Mapelli-Brahm, P.; Meléndez-Martínez, A.J. Sustainable Green Extraction of Carotenoid Pigments: Innovative Technologies and Bio-Based Solvents. Antixidants. 2024, 13, 239. [Google Scholar] [CrossRef]
- Yadav, S.; Tiwari, K.S.; Gupta, C.; Tiwari, M.K.; Khan, A.; Sonkar, S.P. A Brief Review on Natural Dyes, Pigments: Recent Advances and Future Perspectives. Results Chem. 2023, 5, 100733. [Google Scholar] [CrossRef]
- Tzima, S.; Georgiopoulou, I.; Louli, V.; Magoulas, K. Recent Advances in Supercritical CO2 Extraction of Pigments, Lipids and Bioactive Compounds from Microalgae. Molecules. 2023, 28, 1410. [Google Scholar] [CrossRef]
- Elmaaty, T.A.; Sayed-Ahmed, K.; Magdi, M.; Elsisi, H. An eco-friendly method of extracting alizarin from Rubia tinctorum roots under supercritical carbon dioxide and its application to wool dyeing. Sci. Rep. 2023, 13, 30. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.S.L.; Magalhães, R.D.; Fraga, S.; de Souza, P.T.; de Lima, J.P.; Meirelles, A.J.D.A.; Sampaio, K.A. Extraction of Bioactive Compounds from Butia capitata Fruits Using Supercritical Carbon Dioxide and Pressurized Fluids. J. Supercrit. Fluids 2023, 199, 105959. [Google Scholar] [CrossRef]
- Peng, L.-Q.; Cao, J.; Du, L.-J.; Zhang, Q.-D.; Xu, J.-J.; Chen, Y.-B.; Shi, Y.-T.; Li, R.-R. Rapid Ultrasonic and Microwave-Assisted Micellar Extraction Of Zingiberone, Shogaol and Gingerols from Gingers Using Biosurfactants. J. Chromatogr. 2017, 1515, 37–44. [Google Scholar] [CrossRef]
- Wang, H.T.; Shi, L.; Yang, X.Y.; Hong, R.; Li, L. Ultrasonic-Assisted Extraction of Natural Yellow Pigment from Physalis pubescens L. and Its Antioxidant Activities. J. Chem. 2018, 1, 7861639. [Google Scholar] [CrossRef]
- Sharma, M.; Bhat, R. Extraction of Carotenoids from Pumpkin Peel and Pulp: Comparison between Innovative Green Extraction Technologies (Ultrasonic and Microwave-Assisted Extractions Using Corn Oil). Foods 2021, 10, 787. [Google Scholar] [CrossRef]
- Chesnokova, N.Y.; Levochkina, L.V.; Prikhod’ko, Y.V.; A Kuznetsova, A.; A Chebukin, P. Influence of Ultrasound and Extraction Conditions on the Intensity of Extraction of Anthocyanin Pigment from Berry Raw Materials. IOP Conf. Ser. Earth Environ. Sci. 2021, 848, 012211. [Google Scholar] [CrossRef]
- Surmanidze, N.; Vanidze, M.; Djafaridze, I.; Davitadze, R.; Qarcivadze, I.; Khakhutaishvili, M.; Kalandia, A. Optimization of the Method of Ultrasonic Extraction of Lycopene with a Green Extract from the Fruit of Elaeagnus Umbellata, Common in Western Georgia. Food Sci. Nutr. 2024, 12, 3593–3601. [Google Scholar] [CrossRef]
- Ordoñez-Santos, L.E.; Martínez-Girón, J.; Rodríguez-Rodríguez, D.X. Extraction of Total Carote-noids from Peach Palm Fruit (Bactris gasipaes) Peel by Means of Ultrasound Application and Vegetable Oil. Dyna 2019, 86, 91–96. [Google Scholar]
- Čulina, P.; Balbino, S.; Jokić, S.; Dragović-Uzelac, V.; Pedisić, S. Efficiency of Supercritical CO2 and Ultrasound-Assisted Extraction Techniques for Isolation of Bioactive Molecules from Sea Buckthorn (Elaeagnus rhamnoides (L.) A. Nelson) Berry Oils and Cakes. Processes 2024, 12, 698. [Google Scholar] [CrossRef]
- Molina, A.K.; Corrêa, R.C.G.; Prieto, M.A.; Pereira, C.; Barros, L. Bioactive Natural Pigments’ Extraction, Isolation, and Stability in Food Applications. Molecules 2023, 28, 1200. [Google Scholar] [CrossRef]
- Pramudika, S.; Paramita, V. Efficiency of Betacyanin and Betaxanthin Extraction from Red Beets (Beta vulgaris L.) using Microwave-Assisted Extraction (Mae). J. Vocat. Stud. Appl. Res. 2024, 5, 68–72. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave Assisted Extraction of Phenolic Compounds from Four Economic Brown Macroalgae Species and Evaluation of Their Antioxidant Activities and Inhibitory Effects on A-Amylase, A-Glucosidase, Pancreatic Lipase and Tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef]
- Kiruthika, M.; Shivaswamy, M.S. Extraction of Provitamin and Non-Provitamin Carotenoid Using Conventional and Modern Extraction Methods—A Review. Food Humanit. 2024, 2, 100241. [Google Scholar] [CrossRef]
- Silva, J.P.P.; Bolanho, B.C.; Stevanato, N.; Massa, T.B.; da Silva, C. Ultrasound-Assisted Extraction of Red Beet Pigments (Beta vulgaris L.): Influence of Operational Parameters and Kinetic Modeling. J. Food Process. Preserv. 2020, 44, e14762. [Google Scholar] [CrossRef]
- López-Bascón, M.A.; De Castro, M.D.L. Soxhlet Extraction. In Liquid-Phase Extraction; Elsevier: Amsterdam, The Netherlands, 2020; pp. 327–354. [Google Scholar]
- Mathews, A.; Arbal, A.V.; Kaarunya, A.; Jha, P.K.; Le-Bail, A.; Rawson, A. Conventional Vs Modern Extraction Techniques in the Food Industry. In Extraction Processes in the Food Industry; Elsevier: Amsterdam, The Netherlands, 2024; pp. 97–146. [Google Scholar]
- Wani, F.A.; Rashid, R.; Jabeen, A.; Brochier, B.; Yadav, S.; Aijaz, T.; Makroo, H.A.; Dar, B.N. Valorisation of Food Wastes to Produce Natural Pigments Using Non-Thermal Novel Extraction Methods: A Review. Int. J. Food Sci. Technol. 2021, 56, 4823–4833. [Google Scholar] [CrossRef]
- Goti, D.; Dasgupta, S. A Comprehensive Review of Conventional and Non-Conventional Solvent Extraction Techniques. J. Pharmacogn. Phytochem. 2023, 12, 202–211. [Google Scholar] [CrossRef]
- Bhadange, Y.A.; Carpenter, J.; Saharan, V.K. A Comprehensive Review on Advanced Extraction Techniques for Retrieving Bioactive Components from Natural Sources. ACS Omega 2024, 9, 31274–31297. [Google Scholar] [CrossRef]
- Da Silva, D.V.T.; Baião, D.D.S.; Magalhães, A.; Almeida, N.F.; Conte, C.A.; Paschoalin, V.M.F. Combining Conventional Organic Solvent Extraction, Ultrasound-Assisted Extraction, and Chromatographic Techniques to Obtain Pure Betanin from Beetroot for Clinical Purposes. Antioxidants 2023, 12, 1823. [Google Scholar] [CrossRef]
- Sooch, B.S.; Sandhu, N.; Mann, M.K.; Ray, R.C. Valorization of Beetroot Waste for Extraction of Natural Dye for Textile and Food Applications. In Roots, Tubers, and Bulb Crop Wastes: Management by Biorefinery Approaches; Springer: Singapore, 2024; pp. 237–260. [Google Scholar]
- Lozano Pérez, A.S.; Lozada Castro, J.J.; Guerrero Fajardo, C.A. Application of Microwave Energy to Biomass: A Comprehensive Review of Microwave-Assisted Technologies, Optimization Parameters, and the Strengths and Weaknesses. J. Manuf. Mater. Process. 2024, 8, 121. [Google Scholar] [CrossRef]
- Wang, P.; Grimm, B. Organization of chlorophyll biosynthesis and insertion of chlorophyll into the chlorophyll-binding proteins in chloroplasts. Photosynth. Res. 2015, 126, 189–202. [Google Scholar] [CrossRef]
- Wang, H.; Li, Z.; Yuan, L.; Zhou, H.; Hou, X.; Liu, T. Cold Acclimation can Specifically Inhibit Chlorophyll Biosynthesis in Young Leaves of Pakchoi. BMC Plant Biol. 2021, 21, 172. [Google Scholar] [CrossRef] [PubMed]
- Qiu, N.; Jiang, D.; Wang, X.; Wang, B.; Zhou, F. Advances in the Members and Biosynthesis of Chlorophyll Family. Photosynthetica 2019, 57, 974–984. [Google Scholar] [CrossRef]
- Fang, Y.; Zhao, S.; Zhang, F.L.; Zhao, A.G.; Zhang, W.X.; Zhang, M.; Liu, L. The Arabidopsis Glutamyl-Trna Reductase (Glutr) Forms a Ternary Complex with Flu and Glutr-Binding Protein. Sci. Rep. 2016, 6, 19756. [Google Scholar] [CrossRef]
- Wu, Y.; Liao, W.; Dawuda, M.M.; Hu, L.; Yu, J. 5-Aminolevulinic Acid (Ala) Biosynthetic and Metabolic Pathways and Its Role in Higher Plants: A Review. Plant Growth Regul. 2019, 87, 357–374. [Google Scholar] [CrossRef]
- Tan, S.; Cao, J.; Xia, X.; Li, Z. Advances in 5-Aminolevulinic Acid Priming to Enhance Plant Tolerance to Abiotic Stress. Int. J. Mol. Sci. 2022, 23, 702. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.; Wang, L. Study on the Mechanism of Exogenous 5-Aminolevulinic Acid (Ala) in Regulating the Photo-synthetic Efficiency of Pear Leaves. Forests 2024, 15, 769. [Google Scholar] [CrossRef]
- Qaisar, U.; Afzal, M.; Tayyeb, A. Commercial Applications of Plant Pigments. Int. J. Biotech. Trends Technol. 2019, 9, 18–22. [Google Scholar] [CrossRef]
- Yuniati, Y.; Handarini, K.; Mahfud, M. Pigment Extraction Method for Anthocyanin Natural Resources in Indonesia: A Review. ASEAN J. Chem. Eng. 2024, 24, 66–78. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Zuluaga, M.Y.A.; Monterisi, S.; Rouphael, Y.; Colla, G.; Lucini, L.; Cesco, S.; Pii, Y. Different vegetal protein hydrolysates distinctively alleviate salinity stress in vegetable crops: A case study on tomato and lettuce. Front. Plant Sci. 2023, 14, 1077140. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, B.; Mu, B.; Zheng, X.; Zhao, F.; Lan, W.; Fu, A.; Luan, S. A Thylakoid Membrane Protein Functions Synergistically with GUN5 in Chlorophyll Biosynthesis. Plant Commun. 2020, 1, 100094. [Google Scholar] [CrossRef] [PubMed]
- Garnett, J.M.; Wellner, N.; Mayes, A.G.; Downey, G.; Kemsley, E.K. Using Induced Chlorophyll Production to Monitor the Physiological State of Stored Potatoes (Solanum tuberosum L.). Postharvest Biol. Technol. 2018, 145, 222–229. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid Metabolism in Plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Shao, Z.; Zhang, M.; Wang, Q. Regulation of carotenoid metabolism in tomato. Mol. Plant 2015, 8, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef]
- Zia-Ul-Haq, M. Past, Present and Future of Carotenoids Research. In Carotenoids: Structure and Function in the Human Body; Springer: Cham, Switzerland, 2021; pp. 827–854. [Google Scholar]
- Swapnil, P.; Meena, M.; Singh, S.K.; Dhuldhaj, U.P.; Harish; Marwal, A. Vital Roles of Carotenoids in Plants and Humans to Deteriorate Stress with Its Structure, Biosynthesis, Metabolic Engineering and Functional Aspects. Curr. Plant Biol. 2021, 26, 100203. [Google Scholar] [CrossRef]
- Sathasivam, R.; Radhakrishnan, R.; Kim, J.K.; Park, S.U. An Update on Biosynthesis and Regulation of Carotenoids in Plants. S. Afr. J. Bot. 2021, 140, 290–302. [Google Scholar] [CrossRef]
- Quian-Ulloa, R.; Stange, C. Carotenoid Biosynthesis and Plastid Development in Plants: The Role of Light. Int. J. Mol. Sci. 2021, 22, 1184. [Google Scholar] [CrossRef]
- Sun, T.; Rao, S.; Zhou, X.; Li, L. Plant carotenoids: Recent advances and future perspectives. Mol. Hortic. 2022, 2, 3. [Google Scholar] [CrossRef]
- Zhou, X.; Rao, S.; Wrightstone, E.; Sun, T.; Lui, A.C.W.; Welsch, R.; Li, L. Phytoene Synthase: The Key Rate-Limiting Enzyme of Carotenoid Biosynthesis in Plants. Front. Plant Sci. 2022, 13, 884720. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.E.; Hunter, C.N. Engineering Chlorophyll, Bacteriochlorophyll, and Carotenoid Biosynthetic Pathways in Escherichia coli. ACS Synth. Biol. 2023, 12, 2236–2244. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N.; Nakagawa, M.; Kobayashi, K.; Yamano, S.; Izawa, Y.; Nakamura, K.; Harashima, K. Elucidation of the Erwinia uredovora Carotenoid Biosynthetic Pathway by Functional Analysis of Gene Products Expressed in Escherichia coli. J. Bacteriol. 1990, 172, 6704–6712. [Google Scholar] [CrossRef] [PubMed]
- Liang, M.-H.; Zhu, J.; Jiang, J.-G. Carotenoids Biosynthesis and Cleavage Related Genes from Bacteria to Plants. Crit. Rev. Food Sci. Nutr. 2018, 58, 2314–2333. [Google Scholar] [CrossRef]
- Cazzonelli, C.I. Carotenoids in nature: Insights from plants and beyond. Funct. Plant Biol. 2011, 38, 833–847. [Google Scholar] [CrossRef]
- Yan, H.; Pei, X.; Zhang, H.; Li, X.; Zhang, X.; Zhao, M.; Chiang, V.L.; Sederoff, R.R.; Zhao, X. MYB-Mediated Regulation of Anthocyanin Biosynthesis. Int. J. Mol. Sci. 2021, 22, 3103. [Google Scholar] [CrossRef]
- Majumder, J.; Subrahmanyeswari, T.; Gantait, S. Natural Biosynthesis, Pharmacological Applications, and Sustainable Biotechnological Production of Ornamental Plant-Derived Anthocyanin: Beyond Colorants and Aesthetics. 3 Biotech. 2024, 14, 175. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, X.; Gao, X.; Wu, W.; Zhou, B. Light induced regulation pathway of anthocyanin biosynthesis in plants. Int. J. Mol. Sci. 2021, 22, 11116. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid Biosynthetic Pathways in Plants: Versatile Targets for Metabolic Engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef]
- Menconi, J.; Perata, P.; Gonzali, S. In Pursuit of Purple: Anthocyanin Biosynthesis in Fruits of the Tomato Clade. Trends Plant Sci. 2024, 29, 589–604. [Google Scholar] [CrossRef]
- Bean, A.; Sunnadeniya, R.; Akhavan, N.; Campbell, A.; Brown, M.; Lloyd, A. Gain-of-Function Mutations in Beet DODA2 Identify Key Residues for Betalain Pigment Evolution. New Phytol. 2018, 219, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, Y.; Long, T.; Wang, S.; Yang, J. Regulation Mechanism of Plant Pigments Biosynthesis: Anthocyanins, Carotenoids, and Betalains. Metabolites 2022, 12, 871. [Google Scholar] [CrossRef] [PubMed]
- DeLoache, W.C.; Russ, Z.N.; Narcross, L.; Gonzales, A.M.; Martin, V.J.J.; Dueber, J.E. An Enzyme-Coupled Biosensor Enables (S)-Reticuline Production in Yeast from Glucose. Nat. Chem. Biol. 2015, 11, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Sunnadeniya, R.; Bean, A.; Brown, M.; Akhavan, N.; Hatlestad, G.; Gonzalez, A.; Symonds, V.V.; Lloyd, A. Tyrosine Hy-droxylation in Betalain Pigment Biosynthesis Is Performed by Cytochrome P450 Enzymes in Beets (Beta vulgaris L.). PLoS ONE 2016, 11, e0149417. [Google Scholar] [CrossRef] [PubMed]
- Timoneda, A.; Feng, T.; Sheehan, H.; Walker-Hale, N.; Pucker, B.; Lopez-Nieves, S.; Guo, R.; Brockington, S. The Evolution of Betalain Biosynthesis in Caryophyllales. New Phytol. 2019, 224, 71–85. [Google Scholar] [CrossRef]
- Ipekoglu, E.M.; Göçmen, K.; Öz, M.T.; Gürgan, M.; Yücel, M. Cloning and Heterologous Expression of Chlorophyll a Syn-thase in Rhodobacter Sphaeroides. J. Basic. Microbiol. 2017, 57, 238–244. [Google Scholar] [CrossRef]
- Liu, X.Y.; Cao, J.J.; Cheng, X.; Zhu, W.F.; Sun, Y.; Wan, X.C.; Liu, L.L. Csrve1 Promotes Seasonal Greening of Albino Camellia Sinensis Cv. Huangkui by Activating Chlorophyll Biosynthesis. Tree Physiol. 2023, 43, 1432–1443. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, W.; Zhan, X.; Zhang, M.; Xiao, Y.; Hou, X.; Gao, M.; Xiao, B.; Gao, Y. CsCHLI Plays an Important Role in Chlorophyll Biosynthesis of Tea Plant (Camellia sinensis). Beverage Plant Res. 2024, 4, e004. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, S.; Shao, X.; Park, J.-B.; Jeong, S.-H.; Park, H.-J.; Kwak, W.-J.; Wei, G.; Kim, S.-W. Challenges and Tackles in Metabolic Engineering for Microbial Production of Carotenoids. Microb. Cell Factories 2019, 18, 55. [Google Scholar] [CrossRef]
- Naz, T.; Ullah, S.; Nazir, Y.; Li, S.; Iqbal, B.; Liu, Q.; Mohamed, H.; Song, Y. Industrially Important Fungal Carotenoids: Advancements in Biotechnological Production and Extraction. J. Fungi 2023, 9, 578. [Google Scholar] [CrossRef]
- Hattan, J.I.; Furubayashi, M.; Maoka, T.; Takemura, M.; Misawa, N. Reconstruction of the Native Biosynthetic System of Carotenoids in E. coli—Biosynthesis of a Series of Carotenoids Specific to Paprika Fruit. ACS Synth. Biol. 2023, 12, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Liu, N.; Greisen, P.; Li, J.; Qiao, K.; Huang, S.; Stephanopoulos, G. Removal of Lycopene Substrate Inhibition Enables High Carotenoid Productivity in Yarrowia lipolytica. Nat. Commun. 2022, 13, 572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-X.; Xu, L.-W.; Xu, Y.-S.; Li, J.; Ma, W.; Sun, X.-M.; Huang, H. Integration of Genetic Engineering and Multi-Factor Fermentation Optimization for Co-Production of Carotenoid and DHA in Schizochytrium sp. Bioresour. Technol. 2024, 394, 130250. [Google Scholar] [CrossRef] [PubMed]
- Sereti, F.; Alexandri, M.; Papadaki, A.; Papapostolou, H.; Kopsahelis, N. Carotenoids Production by Rhodosporidium Palu-digenum Yeasts: Characterization of Chemical Composition, Antioxidant and Antimicrobial Properties. J. Biotechnol. 2024, 386, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Zhang, Y.; Li, W.; Li, Z.; Zhang, D.; Mo, Q.; Cao, M.; Yuan, J. Multidimensional Optimization of Saccharomyces cerevisiae for Carotenoid Overproduction. BioDesign Res. 2024, 6, 0026. [Google Scholar] [CrossRef]
- Postma, E.D.; Hassing, E.-J.; Mangkusaputra, V.; Geelhoed, J.; de la Torre, P.; van den Broek, M.; Mooiman, C.; Pabst, M.; Daran, J.-M.; Daran-Lapujade, P. Modular, synthetic chromosomes as new tools for large scale engineering of metabolism. Metab. Eng. 2022, 72, 1–13. [Google Scholar] [CrossRef]
- Ji, X.; Li, Z.; Zhang, M.; Lang, S.; Song, X. ChMYB1-ChbHLH42-ChTTG1 Module Regulates Abscisic Acid-Induced Anthocyanin Biosynthesis in Cerasus humilis. Hortic. Plant J. 2024, 10, 51–65. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Y.; Deng, Y.; Yao, Q.; Xiong, A. Synthesis of Betanin by Expression of the Core Betalain Biosynthetic Pathway in Carrot. Hortic. Plant J. 2024, 10, 732–742. [Google Scholar] [CrossRef]
- Thomsen, P.T.; Meramo, S.; Ninivaggi, L.; Pasutto, E.; Babaei, M.; Avila-Neto, P.M.; Pastor, M.C.; Sabri, P.; Rago, D.; Parekh, T.U.; et al. Beet red food colourant can be produced more sustainably with engineered Yarrowia lipolytica. Nat. Microbiol. 2023, 8, 2290–2303. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, X.; Li, J.; Meng, Y.; Zhao, G.-R. Improvement of Betanin Biosynthesis in Saccharomyces cerevisiae by Metabolic Engineering. Synth. Syst. Biotechnol. 2023, 8, 54–60. [Google Scholar] [CrossRef]
- Chen, K.; Yu, Y.; Zhang, Z.; Hu, B.; Liu, X.; James, A.A.; Tan, A. Engineering a Complex, Multiple Enzyme-Mediated Synthesis of Natural Plant Pigments in the Silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2023, 120, e2306322120. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Huang, X.; Chen, J.L.; Qin, Y.Y.; Gao, X.L. Recent Research and Prospects of Non-Thermal Physical Technologies in Green and High-Efficient Extraction of Natural Pigments: A Review. Innov. Food Sci. Emerg. Technol. 2024, 92, 103593. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Method | Control Parameter | Advantages | Limitations | Reference |
|---|---|---|---|---|
| Soxhlet Extraction | Eextraction solvent, extraction solvent reflux times (extraction time), ratio of solid to liquid, and extraction temperature | the instrument is low low-cost, simple, and can be used for multiple extractions;, the continuous flow of fresh solvents to the sample is beneficial to the extraction of the sample;, and the final extract does not need to be filtered | the process time is long, glassware is easy to damage, the open system is prone to solvent leakage, high boiling point temperature causes the thermal degradation of compounds, and cosolvent increases the cost | [28,48,49] |
| Supercritical Fluid [Extraction], SFE | Solvent, temperature, pressure, the addition of modifiers, solid to solvent ratio, and extraction time | high selectivity, it is suitable for highly sensitive applications, heat sensitive compounds are not easy to degrade, CO2 as a solvent is non-toxic and easy to obtain, and has a wide range of applications | the equipment is expensive, the operation is complex, CO2 is not suitable for some polar compounds, the parameter conditions are difficult, and the processing scale is small | [50,51,52] |
| Ultrasound-Assisted Extraction, UAE | Frequency, power intensity, amplitude, extraction time, temperature, type of ultrasonic, and treatment device (probe and bath) | shorten the extraction time, process automation, and reduce the consumption of organic solvents | requires professional equipment, only for laboratory scale (limited application), high energy consumption, extraction equipment conditions and ultrasonic conditions, and complicated operation procedures | [25,52,53] |
| Microwave-assisted Extraction, MAE) | Ssolvent feed ratio, solvent composition, moisture content, characteristics of plant samples, irradiation time, effect of stirring, microwave energy density, and extraction temperature | high efficiency and productivity, selective extraction of target compounds, high reaction rate, the product and process have high quality, reduce environmental pollution, and can reduce the degradation of heat-resistant components and reduce costs | the uneven heating leads to incomplete extraction, not suitable for reaction monitoring, high cost, complicated operation procedure, and limited application | [53,54,55] |
| Four Major Plant Pigments | Existence in Plants | The Most Representative Components | Reference |
|---|---|---|---|
| Chlorophyll | Higher plants and other organisms capable of photosynthesis | Chl a, Chl b as shown in Figure 1. | |
| Carotenoids | Yellow-orange vegetables and fruits, orange fruit, dark green vegetables, and lycopene | 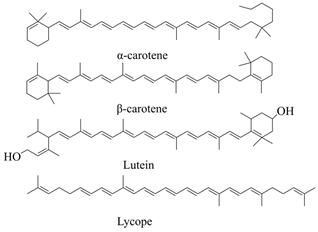 | [63] |
| Anthocyanins | Tissues of purple sweet potato, grape, blood orange, red head cabbage, blueberry, eggplant, cherry, red berry, strawberry, mulberry, hawthorn, morning glory, Elderberries, etc. | 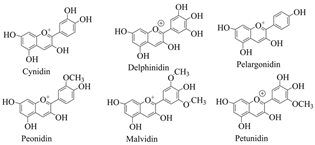 | [64,65] |
| Betalaine | Chenopodiaceae, Amaranthaceae, Cactaceae, Mirabilis, and Phytolacca plants and some higher fungi. | 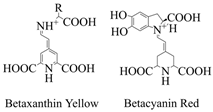 | [66] |
| Methods of Improvement | Optimization Methods | Biological Excitation Factors | Reference |
|---|---|---|---|
| Plant cell culture and tissue culture | (1) Selection and development of high-yield cell lines; (2) optimizing cell culture process; (3) including the optimization of culture conditions; (4) recovering cells by immobilization; (5) redesign the bioreactor. | (1) Exogenous compounds secreted by microorganisms and insects when attacking plants; (2) exogenous compounds formed by plant enzymes degrading microbial cells, such as fungal and bacterial lysates and polysaccharides of microbial cell walls; (3) plant cell wall fragments degraded by pathogens; (4) intracellular proteins or compounds synthesized by plants in response to pathogen attack or abiotic stress (e.g., plant hormones); (5) non-biological elicitors (such as heavy metals, ultraviolet radiation, inorganic salts, etc.). | [16] |
| Microbial culture | (1) Medium; (2) process parameters; (3) extraction conditions. | (1) Cell growth; (2) nutritional factors (carbon source, nitrogen source, and C/N ratio); (3) microbial parameters (spore age, seed age, and inoculation age); (4) environmental conditions. | |
| Heterologous biosynthesis | (1) Engineering of rate-limiting biological components/factors (promoters, protein engineering to optimize enzyme properties, cofactors); (2) metabolic network engineering (supply of precursor substances to target products, enhancement of target product formation ability, inhibition of competitive pathways); (3) the engineering of cell system (external environment and cell expression). | Functional genes and responsible pathways. | |
| Co-culture system | Modularization | The biosynthetic pathway is divided into different host strains. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Wang, H.; Xiang, X.; Fu, J.; Wang, X.; Zhou, Y.; Xing, W. Biosynthesis and Extraction of Chlorophyll, Carotenoids, Anthocyanins, and Betalaine In Vivo and In Vitro. Curr. Issues Mol. Biol. 2024, 46, 10662-10676. https://doi.org/10.3390/cimb46090633
Yu X, Wang H, Xiang X, Fu J, Wang X, Zhou Y, Xing W. Biosynthesis and Extraction of Chlorophyll, Carotenoids, Anthocyanins, and Betalaine In Vivo and In Vitro. Current Issues in Molecular Biology. 2024; 46(9):10662-10676. https://doi.org/10.3390/cimb46090633
Chicago/Turabian StyleYu, Xinxin, Hao Wang, Xingchun Xiang, Jingjing Fu, Xin Wang, Yuanhang Zhou, and Wang Xing. 2024. "Biosynthesis and Extraction of Chlorophyll, Carotenoids, Anthocyanins, and Betalaine In Vivo and In Vitro" Current Issues in Molecular Biology 46, no. 9: 10662-10676. https://doi.org/10.3390/cimb46090633
APA StyleYu, X., Wang, H., Xiang, X., Fu, J., Wang, X., Zhou, Y., & Xing, W. (2024). Biosynthesis and Extraction of Chlorophyll, Carotenoids, Anthocyanins, and Betalaine In Vivo and In Vitro. Current Issues in Molecular Biology, 46(9), 10662-10676. https://doi.org/10.3390/cimb46090633