
Assessment of Five Pesticides as Endocrine-Disrupting Chemicals: Effects on Estrogen Receptors and Aromatase

 , ,
, ,  , and
, and
Abstract
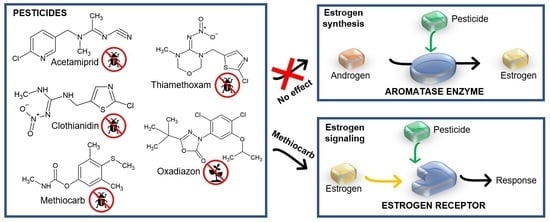
:
1. Introduction
2. Materials and Methods
2.1. Pesticides
2.2. ELISA Assay
2.3. MELN Gene Reporter Assay
2.4. Statistical Analysis
3. Results
3.1. Pesticide Effect on Estrogen Biosynthesis
3.2. Pesticide Effect on Estrogen Signaling
4. Discussion

{kind=link}
{kind=link}
{kind=link}
| Pesticide | Pesticide Type | Assay | EEF | Reference |
|---|---|---|---|---|
| Methiocarb | Carbamate insecticide | Gene reporter assay (MELN cells) | 8.00 × 10−8 | Present study |
| Imidacloprid | Neonicotinoid insecticide | Gene reporter assay (MELN cells) | 5.40 × 10−10 | [29] |
| Thiacloprid | Neonicotinoid insecticide | Gene reporter assay (MELN cells) | 3.70 × 10−9 | [29] |
| Chlorpyrifos | Organophosphate insecticide | Yeast Estrogen Screen assay | 2.90 × 10−3 | [39] |
| Dieldrin | Organochloride insecticide | ER-CALUX assay (T47D Luc cells) | 2.40 × 10−7 | [40] |
| Endosulfan | Organochloride insecticide | ER-CALUX assay (T47D Luc cells) | 1.00 × 10−6 | [40] |
| Permethrin | Pyrethroid insecticide | Yeast Estrogen Screen assay | 1.00 × 10−7—no estrogenic activity | [41] |
| Chlordane | Organochlorine insecticide | ER-CALUX assay (T47D Luc cells) | 9.60 × 10−7 | [40] |
| DDT | Organochlorine insecticide | ER-CALUX assay (T47D Luc cells) | 9.10 × 10−6 | [42] |
| Alachlor | Chloroacetanilide herbicide | Receptor binding assay | 8.00 × 10−6 | [43] |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kabir, E.; Jahan, S.A. Exposure to pesticides and the associated human health effects. Sci. Total Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Bourguignon, D. EU Policy and Legislation on Pesticides. Plant Protection Products and Biocides; Publications Office of the European Union Luxembourg: Luxembourg, 2017. [Google Scholar]
- Tudi, M.; Ruan, H.D.; Wang, L.; Lyu, J.; Sadler, R.; Connell, D.; Chu, C.; Phung, D. Agriculture Development, Pesticide Application and Its Impact on the Environment. Int. J. Environ. Res. Public Health 2021, 18, 1112. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations—FAO. FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/RP (accessed on 10 December 2021).
- Frank, S.D.; Tooker, J.F. Opinion: Neonicotinoids pose undocumented threats to food webs. Proc. Natl. Acad. Sci. USA 2020, 117, 22609–22613. [Google Scholar] [CrossRef] [PubMed]
- Handford, C.E.; Elliott, C.T.; Campbell, K. A review of the global pesticide legislation and the scale of challenge in reaching the global harmonization of food safety standards. Integr. Environ. Assess. Manag. 2015, 11, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Samsidar, A.; Siddiquee, S.; Shaarani, S.M. A review of extraction, analytical and advanced methods for determination of pesticides in environment and foodstuffs. Trends Food Sci. Technol. 2018, 71, 188–201. [Google Scholar] [CrossRef]
- Barbosa, M.O.; Moreira, N.F.F.; Ribeiro, A.R.; Pereira, M.F.R.; Silva, A.M.T. Occurrence and removal of organic micropollutants: An overview of the watch list of EU Decision 2015/495. Water Res. 2016, 94, 257–279. [Google Scholar] [CrossRef]
- Starner, K.; Goh, K.S. Detections of the Neonicotinoid Insecticide Imidacloprid in Surface Waters of Three Agricultural Regions of California, USA, 2010–2011. Bull. Environ. Contam. Toxicol. 2012, 88, 316–321. [Google Scholar] [CrossRef]
- Olisah, C.; Okoh, O.O.; Okoh, A.I. Occurrence of organochlorine pesticide residues in biological and environmental matrices in Africa: A two-decade review. Heliyon 2020, 6, e03518. [Google Scholar] [CrossRef]
- Masiá, A.; Ibañez, M.; Blasco, C.; Sancho, J.V.; Pico, Y.; Hernandez, F. Combined use of liquid chromatography triple quadrupole mass spectrometry and liquid chromatography quadrupole time-of-flight mass spectrometry in systematic screening of pesticides and other contaminants in water samples. Anal. Chim. Acta 2013, 761, 117–127. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Hyne, R.V. Detection and analysis of neonicotinoids in river waters—Development of a passive sampler for three commonly used insecticides. Chemosphere 2014, 99, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Köhler, H.-R.; Triebskorn, R.; Meierbachtol, T.; Harper, J.; Humphrey, N. Wildlife Ecotoxicology of Pesticides: Can We Track Effects to the Population Level and Beyond? Science 2013, 341, 759–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide Exposure, Safety Issues, and Risk Assessment Indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.E.; Meade, B.J. Potential Health Effects Associated with Dermal Exposure to Occupational Chemicals. Environ. Health Insights 2014, 8, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mnif, W.; Hassine, A.I.H.; Bouaziz, A.; Bartegi, A.; Thomas, O.; Roig, B. Effect of Endocrine Disruptor Pesticides: A Review. Int. J. Environ. Res. Public Health 2011, 8, 2265–2303. [Google Scholar] [CrossRef] [Green Version]
- Kabir, E.R.; Rahman, M.S.; Rahman, I. A review on endocrine disruptors and their possible impacts on human health. Environ. Toxicol. Pharmacol. 2015, 40, 241–258. [Google Scholar] [CrossRef]
- Combarnous, Y. Endocrine Disruptor Compounds (EDCs) and agriculture: The case of pesticides. Comptes Rendus Biol. 2017, 340, 406–409. [Google Scholar] [CrossRef]
- Kiyama, R.; Wada-Kiyama, Y. Estrogenic endocrine disruptors: Molecular mechanisms of action. Environ. Int. 2015, 83, 11–40. [Google Scholar] [CrossRef]
- Briz, V.; Molina-Molina, J.-M.; Sánchez-Redondo, S.; Fernández, M.F.; Grimalt, J.O.; Olea, N.; Rodríguez-Farré, E.; Suñol, C. Differential Estrogenic Effects of the Persistent Organochlorine Pesticides Dieldrin, Endosulfan, and Lindane in Primary Neuronal Cultures. Toxicol. Sci. 2011, 120, 413–427. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Yi, B.-R.; Go, R.-E.; Hwang, K.-A.; Nam, K.-H.; Choi, K.-C. Methoxychlor and triclosan stimulates ovarian cancer growth by regulating cell cycle- and apoptosis-related genes via an estrogen receptor-dependent pathway. Environ. Toxicol. Pharmacol. 2014, 37, 1264–1274. [Google Scholar] [CrossRef]
- Baravalle, R.; Ciaramella, A.; Baj, F.; Di Nardo, G.; Gilardi, G. Identification of endocrine disrupting chemicals acting on human aromatase. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2018, 1866, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Higley, E.B.; Newsted, J.L.; Zhang, X.; Giesy, J.P.; Hecker, M. Assessment of chemical effects on aromatase activity using the H295R cell line. Environ. Sci. Pollut. Res. 2010, 17, 1137–1148. [Google Scholar] [CrossRef] [PubMed]
- Andersen, H.R.; Vinggaard, A.M.; Rasmussen, T.H.; Gjermandsen, I.M.; Bonefeld-Jørgensen, E.C. Effects of Currently Used Pesticides in Assays for Estrogenicity, Androgenicity, and Aromatase Activity In Vitro. Toxicol. Appl. Pharmacol. 2002, 179, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Di Nardo, G.; Breitner, M.; Sadeghi, J.; Castrignanò, S.; Mei, G.; Di Venere, A.; Nicolai, E.; Allegra, P.; Gilardi, G. Dynamics and Flexibility of Human Aromatase Probed by FTIR and Time Resolved Fluorescence Spectroscopy. PLoS ONE 2013, 8, e82118. [Google Scholar]
- Huang, N.; Pandey, A.V.; Agrawal, V.; Reardon, W.; Lapunzina, P.D.; Mowat, D.; Jabs, E.W.; Van Vliet, G.; Sack, J.; Flück, C.E.; et al. Diversity and Function of Mutations in P450 Oxidoreductase in Patients with Antley-Bixler Syndrome and Disordered Steroidogenesis. Am. J. Hum. Genet. 2005, 76, 729–749. [Google Scholar] [CrossRef] [Green Version]
- Balaguer, P.; François, F.; Comunale, F.; Fenet, H.; Boussioux, A.-M.; Pons, M.; Nicolas, J.-C.; Casellas, C. Reporter cell lines to study the estrogenic effects of xenoestrogens. Sci. Total Environ. 1999, 233, 47–56. [Google Scholar] [CrossRef]
- Zhang, C.; Schilirò, T.; Gea, M.; Bianchi, S.; Spinello, A.; Magistrato, A.; Gilardi, G.; Di Nardo, G. Molecular Basis for Endocrine Disruption by Pesticides Targeting Aromatase and Estrogen Receptor. Int. J. Environ. Res. Public Health 2020, 17, 5664. [Google Scholar] [CrossRef]
- Laville, N.; Balaguer, P.; Brion, F.; Hinfray, N.; Casellas, C.; Porcher, J.-M.; Aït-Aïssa, S. Modulation of aromatase activity and mRNA by various selected pesticides in the human choriocarcinoma JEG-3 cell line. Toxicology 2006, 228, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Westlund, P.; Yargeau, V. Investigation of the presence and endocrine activities of pesticides found in wastewater effluent using yeast-based bioassays. Sci. Total Environ. 2017, 607–608, 744–751. [Google Scholar] [CrossRef]
- Mesnage, R.; Biserni, M.; Genkova, D.; Wesolowski, L.; Antoniou, M.N. Evaluation of neonicotinoid insecticides for oestrogenic, thyroidogenic and adipogenic activity reveals imidacloprid causes lipid accumulation. J. Appl. Toxicol. 2018, 38, 1483–1491. [Google Scholar] [CrossRef]
- Kojima, H.; Katsura, E.; Takeuchi, S.; Niiyama, K.; Kobayashi, K. Screening for estrogen and androgen receptor activities in 200 pesticides by in vitro reporter gene assays using Chinese hamster ovary cells. Environ. Health Perspect. 2004, 112, 524–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tange, S.; Fujimoto, N.; Uramaru, N.; Wong, F.F.; Sugihara, K.; Ohta, S.; Kitamura, S. In vitro metabolism of methiocarb and carbaryl in rats, and its effect on their estrogenic and antiandrogenic activities. Environ. Toxicol. Pharmacol. 2016, 41, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Jurado, A.; Walther, M.; Díaz-Cruz, M.S. Occurrence, fate and environmental risk assessment of the organic microcontaminants included in the Watch Lists set by EU Decisions 2015/495 and 2018/840 in the groundwater of Spain. Sci. Total Environ. 2019, 663, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Ewence, A.; Brescia, S.; Johnson, I.; Rumsby, P.C. An approach to the identification and regulation of endocrine disrupting pesticides. Food Chem. Toxicol. 2015, 78, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ma, J.; Meng, Y.; Wei, Y.; Xie, S.; Jiang, P.; Wang, Z.; Chen, X.; Liu, Z.; Zhong, K.; et al. Exposure to Oxadiazon-Butachlor causes cardiac toxicity in zebrafish embryos. Environ. Pollut. 2020, 265, 114775. [Google Scholar] [CrossRef]
- Pamplona-Silva, M.T.; Mazzeo, D.E.C.; Bianchi, J.; Marin-Morales, M.A. Estrogenic Compounds: Chemical Characteristics, Detection Methods, Biological and Environmental Effects. Water Air Soil Pollut. 2018, 229, 144. [Google Scholar] [CrossRef] [Green Version]
- Echeverri-Jaramillo, G.; Jaramillo-Colorado, B.; Sabater-Marco, C.; Castillo-López, M.Á. Cytotoxic and estrogenic activity of chlorpyrifos and its metabolite 3,5,6-trichloro-2-pyridinol. Study of marine yeasts as potential toxicity indicators. Ecotoxicology 2021, 30, 104–117. [Google Scholar] [CrossRef]
- Legler, J.; van den Brink, C.E.; Brouwer, A.; Murk, A.J.; Van Der Saag, P.T.; Vethaak, A.D.; Van Der Burg, B. Development of a stably transfected estrogen receptor-mediated luciferase reporter gene assay in the human T47D breast cancer cell line. Toxicol. Sci. 1999, 48, 55–66. [Google Scholar] [CrossRef]
- Tyler, C.R.; Beresford, N.; van der Woning, M.; Sumpter, J.P.; Tchorpe, K. Metabolism and environmental degradation of pyrethroid insecticides produce compounds with endocrine activities. Environ. Toxicol. Chem. 2000, 19, 801–809. [Google Scholar] [CrossRef]
- Legler, J.; Dennekamp, M.; Vethaak, A.D.; Brouwer, A.; Koeman, J.H.; van der Burg, B.; Murk, A.J. Detection of Estrogenic Activity in Sediment-Associated Compounds Using In Vitro Reporter Gene Assays. Sci. Total Environ. 2002, 293, 69–83. [Google Scholar] [CrossRef]
- Scippo, M.-L.; Argiris, C.; Muller, M.; Willemsen, P.; Martial, J.; Maghuin-Rogister, G. Recombinant human estrogen, androgen and progesterone receptors for detection of potential endocrine disruptors. Anal. Bioanal. Chem. 2004, 378, 664–669. [Google Scholar] [CrossRef] [PubMed]


| Pesticide | Relative Activity (% of C+) | ||
|---|---|---|---|
| 0.5 × 10−6 M | 1 × 10−6 M | 5 × 10−6 M | |
| Acetamiprid | 94.7 ± 11.8 | 99.7 ± 4.7 | 87.5 ± 17.6 |
| Clothianidin | 74.1 ± 24.7 | 112.9 ± 14.9 | 97.9 ± 5.2 |
| Thiamethoxam | 101.4 ± 13.2 | 84.1 ± 11.6 | 95.6 ± 17.1 |
| Methiocarb | 98.1 ± 0.2 | 96.5 ± 0.1 | 90.9 ± 5.4 |
| Oxadiazon | 87.8 ± 0.7 | 100.1 ± 13.1 | 101.2 ± 16.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gea, M.; Zhang, C.; Tota, R.; Gilardi, G.; Di Nardo, G.; Schilirò, T. Assessment of Five Pesticides as Endocrine-Disrupting Chemicals: Effects on Estrogen Receptors and Aromatase. Int. J. Environ. Res. Public Health 2022, 19, 1959. https://doi.org/10.3390/ijerph19041959
Gea M, Zhang C, Tota R, Gilardi G, Di Nardo G, Schilirò T. Assessment of Five Pesticides as Endocrine-Disrupting Chemicals: Effects on Estrogen Receptors and Aromatase. International Journal of Environmental Research and Public Health. 2022; 19(4):1959. https://doi.org/10.3390/ijerph19041959
Chicago/Turabian StyleGea, Marta, Chao Zhang, Roberta Tota, Gianfranco Gilardi, Giovanna Di Nardo, and Tiziana Schilirò. 2022. "Assessment of Five Pesticides as Endocrine-Disrupting Chemicals: Effects on Estrogen Receptors and Aromatase" International Journal of Environmental Research and Public Health 19, no. 4: 1959. https://doi.org/10.3390/ijerph19041959