Activation of the Dormant Secondary Metabolite Production by Introducing Gentamicin-Resistance in a Marine-Derived Penicillium purpurogenum G59


Abstract
:1. Introduction
2. Results
2.1. Finding Conditions for Introducing Gentamicin-Resistance in Penicillium purpurogenum G59

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tests in | Test and Control Groups a | Gentamicin (mg/mL) | Dilution Ratio b |
|---|---|---|---|
| 20% DMSO | Tests in 20% DMSO with gentamicin | 0.5, 1, 2, 5, 10, 20, 50, 100 | OSS:DMSO = 4:1 |
| Control in 20% DMSO without gentamicin | 0 | OSS:DMSO = 4:1 | |
| Controls in water with gentamicin | 0.5, 1, 2, 5, 10, 20, 50, 100 | OSS:H2O = 4:1 | |
| Control in water without gentamicin | 0 | OSS:H2O = 4:1 | |
| 50% DMSO | Tests in 50% DMSO with gentamicin | 1, 2, 5, 10, 20 | OSS:DMSO = 1:1 |
| Control in 50% DMSO without gentamicin | 0 | OSS:DMSO = 1:1 | |
| Controls in water with gentamicin | 1, 2, 5, 10, 20 | OSS:H2O = 1:1 | |
| Control in water without gentamicin | 0 | OSS:H2O = 1:1 |


2.2. Mutant Selection
| DMSO% (v/v) | Gentamicin (mg/mL) | Numbers of Mutant Strains Selected on the Days of Gentamicin-Treatment at 4 °C | ||||||
|---|---|---|---|---|---|---|---|---|
| 1st d | 3rd d | 5th d | 10th d | 15th d | 20th d | Total | ||
| 20% | 0.5 | 1 | 1 | 3 | NS | 2 | ND | 7 |
| 1 | 1 | 1 | NS | 1 | 6 | ND | 9 | |
| 2 | 1 | 1 | NS | 4 | 12 | ND | 18 | |
| 5 | 1 | 1 | 3 | 3 | 2 | ND | 10 | |
| 10 | 1 | 2 | 10 | NS | 7 | ND | 20 | |
| 20 | 1 | 1 | 3 | 27 | 1 | ND | 33 | |
| 50 | 1 | 1 | 10 | 4 | 7 | ND | 23 | |
| 100 | 1 | 1 | 4 | 5 | 5 | ND | 16 | |
| Sum | 8 | 9 | 33 | 44 | 42 | 0 | 136 | |
| 50% | 1 | NS | NC | NC | 1 | NC | NC | 1 |
| 2 | 9 | 4 | 5 | 8 | NC | 8 | 34 | |
| 5 | 4 | NC | NC | NC | NC | NC | 4 | |
| 10 | NS | NC | NC | 2 | NC | NC | 2 | |
| 20 | 4 | NC | NC | NC | NC | NC | 4 | |
| Sum | 17 | 4 | 5 | 11 | 0 | 8 | 45 | |
| Total | 25 | 13 | 38 | 55 | 42 | 8 | 181 | |
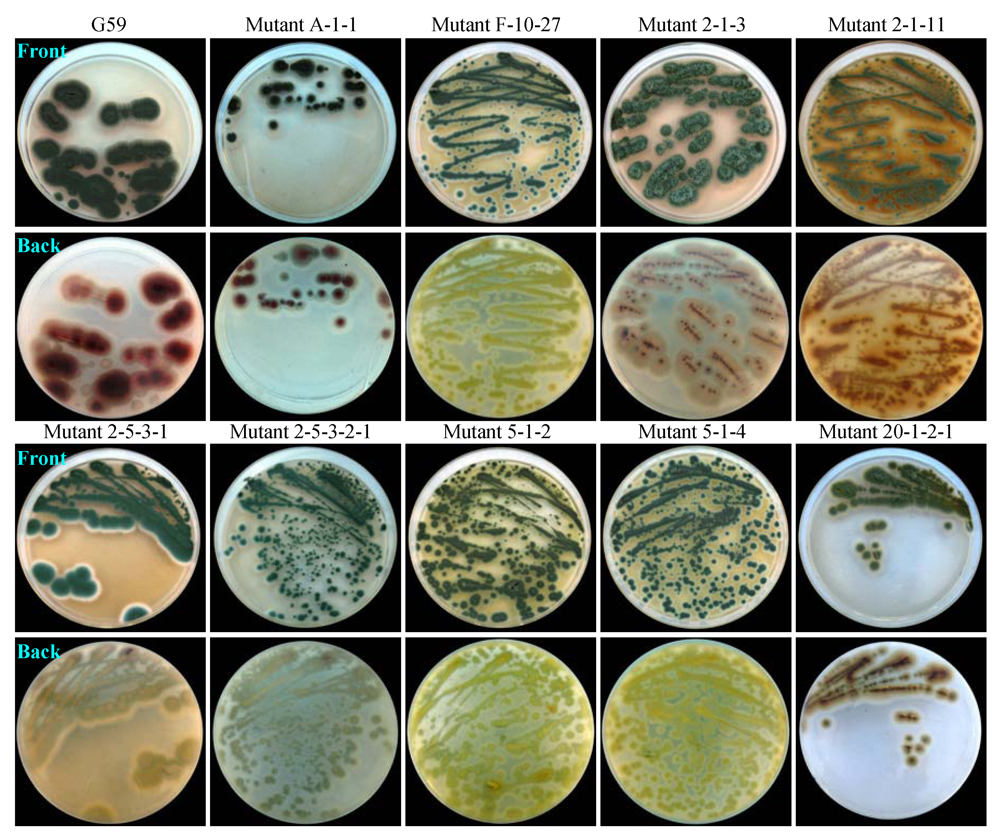
2.3. Phenotypes of G59 Mutants

2.4. Inhibitory Effect of G59 Mutant Samples on K562 Cells
| No. | Strain | Condition for Selecting Mutant Strain | IR% (Mean ± SD, n = 3) b | ||
|---|---|---|---|---|---|
| DMSO (%) | Gentamicin (mg/mL) | Treated Time (days) | |||
| 0 | G59 | — | — | — | 5.8 ± 0.5 |
| 1 | A-1-1 | 20% | 0.5 | 1 | 42.0 ± 12.7 |
| 2 | F-10-27 | 20% | 20 | 10 | 45.5 ± 10.8 |
| 3 | 2-1-3 | 50% | 2 | 1 | 43.5 ± 10.3 |
| 4 | 2-1-11 | 50% | 2 | 1 | 40.0 ± 7.3 |
| 5 | 2-5-3-1 | 50% | 2 | 5 | 80.7 ± 0.7 |
| 6 | 2-5-3-2-1 | 50% | 2 | 5 | 46.1 ± 16.1 |
| 7 | 5-1-2 | 50% | 5 | 1 | 47.2 ± 19.8 |
| 8 | 5-1-4 | 50% | 5 | 1 | 43.7 ± 2.8 |
| 9 | 20-1-2-1 | 50% | 20 | 1 | 52.2 ± 18.1 |

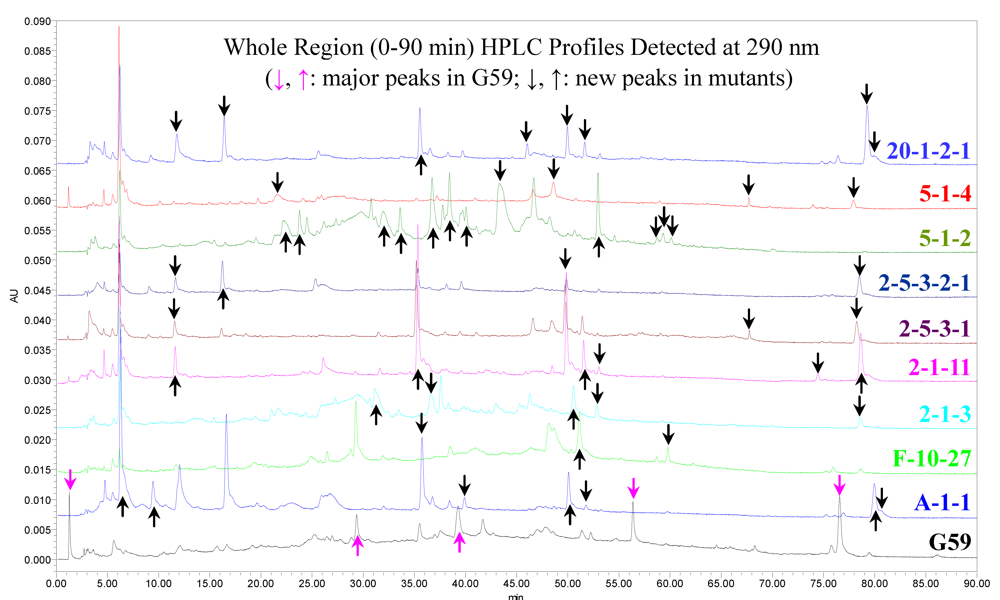
2.5. TLC and HPLC Analysis of the EtOAc Extracts of G59 and Its Nine Mutant Cultures

2.6. Producing Fermentation, Isolation and Identification of Antitumor Metabolites 1–4

2.7. Inhibitory Effect of Compounds 1–4 on the Proliferation of K562 Cells
| Compound | IR% at the Given Concentration (μg/mL) | IC50 (μg/mL) | |||||
|---|---|---|---|---|---|---|---|
| 100 μg/mL | 50 μg/mL | 25 μg/mL | 12.5 μg/mL | 6.25 μg/mL | 3.125 μg/mL | ||
| 1 | 34.6 | 28.4 | 20.6 | 14.3 | 12.6 | 7.6 | >100 |
| 2 | 60.8 | 43.1 | 37.7 | 35.9 | 31.6 | 22.7 | 58.4 |
| 3 | 31.7 | 20.8 | 17.5 | 14.1 | 8.9 | 6.7 | >100 |
| 4 | 67.1 | 44.6 | 41.6 | 25.0 | 2.3 | 1.1 | 52.6 |
3. Discussions

4. Experimental Section
4.1. The Original Strain and Human Tumor Cell Line
4.2. Preparation of Spore Suspensions for Strain G59 and Its Mutant 5-1-4
4.3. Mutant Selection by Treatment of G59 Spores with Gentamycin in Aqueous DMSO
4.4. Resistance Test for Acquired Resistance of Mutant 5-1-4 to Gentamicin
4.5. Fermentation and Sample Preparation for MTT Assay and Chemical Analysis
4.6. MTT Assay
4.7. TLC and HPLC Analysis
4.8. Time Course Experiment for Mutant 5-1-4 Fermentation
4.9. Fermentation and Extract Preparation
4.10. Isolation of Antitumor Metabolites 1–4 Produced by the Mutant 5-1-4
4.11. Physicochemical and Spectral Data for Compounds 1–4
5. Conclusions
Acknowledgments
References
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar]
- Baker, D.D.; Chu, M.; Oza, U.; Rajgarhia, V. The value of natural products to future pharmaceutical discovery. Nat. Prod. Rep. 2007, 24, 1225–1244. [Google Scholar]
- Gordaliza, M. Natural products as leads to anticancer drugs. Clin.Transl. Oncol. 2007, 9, 767–776. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar]
- Clardy, J.C.; Fischbach, M.A.; Walsh, C.T. New antibiotics from bacterial natural products. Nat. Biotechnol. 2006, 24, 1541–1550. [Google Scholar]
- Donadio, S.; Maffioli, S.; Monciardini, P.; Sosio, M.; Jabes, D. Antibiotic discovery in the twenty-first century: Current trends and further perspectives. J. Antibiot. 2010, 63, 423–430. [Google Scholar]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar]
- Ahmed, H.Y.; Shikh, H.H.E.; Ghoneimy, E.A.; Ragab, A.M.; Saad, A.A.; Hashem, A.M.; Sayed, R.M.E.; Shikh, M.E.M.E. Fungal secondary metabolites: A promising source of antineoplastic drugs. Curr.Cancer Ther. Rev. 2010, 6, 222–228. [Google Scholar]
- Suryanarayanan, T.S.; Thirunavukkarasu, N.; Govindarajulu, M.B.; Sasse, F.; Jansen, R.; Murali, T.S. Fungal endophytes and bioprospecting. Fungal Biol. Rev. 2009, 23, 9–19. [Google Scholar]
- Saleem, M.; Ali, M.S.; Hussain, S.; Jabbar, A.; Ashraf, M.; Lee, Y.S. Marine natural products of fungal origin. Nat. Prod. Rep. 2007, 24, 1142–1152. [Google Scholar]
- Gao, S.-S.; Li, X.-M.; Du, F.-Y.; Li, C.-S.; Proksch, P.; Wang, B.-G. Secondary metabolites from a marine-derived endophytic fungus Penicillium chrysogenum QEN-24S. Mar. Drugs 2011, 9, 59–70. [Google Scholar]
- Shushni, M.A.M.; Singh, R.; Mentel, R.; Lndequit, U. Balticolid: A new 12-membered macrolide with antiviral activity from an Ascomycetous fungus of marine origin. Mar. Drugs 2011, 9, 844–851. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, J.; Liu, P.; Wang, W.; Zhu, W. Three new compounds from Aspergillus terreus PT06-2 grown in a high salt medium. Mar. Drugs 2011, 9, 1368–1378. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, Z.; Sun, K.; Zhu, W. Effects of high salt stress on secondary metabolite production in the marine-derived fungus Spicaria elegans. Mar. Drugs 2011, 9, 535–542. [Google Scholar] [CrossRef]
- Schneider, P.; Misiek, M.; Hoffmeister, D. In vivo and in vitro production options for fungal secondary metabolites. Mol. Pharm. 2008, 5, 234–242. [Google Scholar] [CrossRef]
- Shwab, E.K.; Keller, N.P. Regulation of secondary metabolite production in filamentous ascomycetes. Mycol. Res. 2008, 112, 225–230. [Google Scholar]
- Yu, J.H.; Keller, N. Regulation of secondary metabolism in filamentous fungi. Annu.Rev. Phytopathol. 2005, 43, 437–458. [Google Scholar]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef]
- Misiek, M.; Hoffmeister, D. Fungal genetics, genomics, and secondary metabolites in pharmaceutical sciences. Planta Med. 2007, 73, 103–115. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar]
- Saruwatari, T.; Praseuth, A.P.; Sato, M.; Torikai, K.; Noguchi, H.; Watanabe, K. A comprehensive overview on genomically directed assembly of aromatic polyketides and macrolide lactones using fungal megasynthases. J. Antibiot. 2011, 64, 9–17. [Google Scholar]
- Fox, E.M.; Howlett, B.J. Secondary metabolism: Regulation and role in fungal biology. Curr.Opin. Microbiol. 2008, 11, 481–487. [Google Scholar]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol.Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar]
- Handelsman, J. Metagenomics: Application of genomics to uncultured microorganisms. Microbiol.Mol. Biol. Rev. 2004, 68, 669–685. [Google Scholar]
- Lewis, K.; Epstein, S.; D’Onofrio, A.; Ling, L.L. Uncultured microorganisms as a source of secondary metabolites. J. Antibiot. 2010, 63, 468–476. [Google Scholar]
- Hertweck, C. Hidden biosynthetic treasures brought to light. Nat. Chem. Biol. 2009, 5, 450–452. [Google Scholar]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites—Strategies to activate silent gene clusters. Fungal Genet.Biol. 2011, 48, 15–22. [Google Scholar]
- Brakhage, A.A.; Schuemann, J.; Bergmann, S.; Scherlach, K.; Schroeckh, V.; Hertweck, C. Activation of Fungal Silent Gene Clusters: A New Avenue to Drug Discovery. In Progress in Drug Research, Natural Compounds as Drugs Volume II; Petersen, F., Amstutz, R., Eds.; Birkhäuser Verlag AG: Basel, Switzerland, 2008; Volume 66, pp. 1–12. [Google Scholar]
- Bergmann, S.; Schümann, J.; Scherlach, K.; Lange, C.; Brakhage, A.A.; Hertweck, C. Genomics-driven discovery of PKS-NRPS hybrid metabolites from Aspergillus nidulans. Nat. Chem. Biol. 2007, 3, 213–217. [Google Scholar]
- Bentley, S.D.; Chater, K.F.; Cerdeño-Tárraga, A.-M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar]
- Ikeda, H.; Ishikawa, J.; Hnamoto, A.; Shimose, M.; Kikuchi, H.; Shiba, T.; Sakaki, Y.; Hattori, M.; Ōmura, S. Complete genome sequence and comparative analysis of the industrial microorganism Streptomyces avermitilis. Nat. Biotechnol. 2003, 21, 526–531. [Google Scholar] [CrossRef]
- Oliynyk, M.; Samborskyy, M.; Lester, J.B.; Mironenko, T.; Scott, N.; Dickens, S.; Haydock, S.F.; Leadlay, P.F. Complete genome sequence of the erythromycin-producing bacterium Saccharopolyspora erythraea NPRL2338. Nat. Biotechnol. 2007, 25, 447–453. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Ishikawa, J.; Hara, H.; Suzuki, H.; Ikenoya, M.; Ikeda, H.; Yamashita, A.; Hattori, M.; Horinouchi, S. Genome sequence of the streptomycin-producing microorganism Streptomyces griseus IFO 13350. J. Bacteriol. 2008, 190, 4050–4060. [Google Scholar]
- Galagan, J.E.; Calvo, S.E.; Borkovich, K.A.; Selker, E.U.; Read, N.D.; Jaffe, D.; FitzHugh, W.; Ma, L.J.; Smirnov, S.; Purcell, S.; et al. The genome sequence of the filamentous fungus Neurospora crassa. Nature 2003, 422, 859–868. [Google Scholar]
- Martinez, D.; Larrondo, L.F.; Purnam, N.; Gelpke, M.D.S.; Huang, K.; Chapman, J.; Helfenbein, K.G.; Ramaiya, P.; Detter, J.C.; Larimer, F.; et al. Genome sequence of the lignocellulose degrading fungus Phanerochaete chrysosporium strain RP78. Nat. Biotechnol. 2004, 22, 695–700. [Google Scholar]
- Pel, H.J.; Winde, J.H.D.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; de Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar]
- Donadio, S.; Sosio, M.; Lancini, G. Impact of the first Streptomyces genome sequence on the discovery and production of bioactive substances. Appl. Microbiol. Biotechnol. 2002, 60, 377–380. [Google Scholar] [CrossRef]
- Bode, H.B.; Müller, R. The impact of bacterial genomics on natural product research. Angew.Chem. Int. Ed. 2005, 44, 6828–6846. [Google Scholar]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar]
- Christian, O.E.; Compton, J.; Christian, K.R.; Mooberry, S.L.; Valeriote, F.A.; Crews, P. Using jasplakinolide to turn on pathways that enable the isolation of new chaetoglobosins from Phomospis asparagi. J. Nat. Prod. 2005, 68, 1592–1597. [Google Scholar] [CrossRef]
- Paranagama, P.A.; Wijeratne, E.M.K.; Gunatilaka, A.A.L. Uncovering biosynthetic potential of plant-associated fungi: Effect of culture conditions on metabolite production by Paraphaeosphaeria quadriseptata and Chaetomium chiversii. J. Nat. Prod. 2007, 70, 1939–1945. [Google Scholar] [CrossRef]
- Williams, R.B.; Henrikson, J.C.; Hoover, A.R.; Lee, A.E.; Cichewicz, R.H. Epigenetic remodeling of the fungal secondary metabolome. Org. Biomol. Chem. 2008, 6, 1895–1897. [Google Scholar]
- Henrikson, J.C.; Hoover, A.R.; Joyner, P.M.; Cichewicz, R.H. A chemical epigenetics approach for engineering the in situ biosynthesis of a cryptic natural product from Aspergillus niger. Org. Biomol. Chem. 2009, 7, 435–438. [Google Scholar]
- Fisch, K.M.; Gillaspy, A.F.; Gipson, M.; Henrikson, J.C.; Hoover, A.R.; Jackson, L.; Najar, F.Z.; Wägele, H.; Cichewicz, R.H. Chemical induction of silent biosynthetic pathway transcription in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1199–1213. [Google Scholar] [CrossRef]
- Cichewicz, R.H. Epigenome manipulation as a pathway to new natural product scaffolds and their congeners. Nat. Prod. Rep. 2010, 27, 11–22. [Google Scholar]
- Wang, X.; Filho, J.G.S.; Hoover, A.R.; King, J.B.; Ellis, T.K.; Powell, D.R.; Cichewicz, R.H. Chemical epigenetics alters the secondary metabolite composition of guttate excreted by an atlantic-forest-soil-derived Penicillium citreonigrum. J. Nat. Prod. 2010, 73, 942–948. [Google Scholar] [CrossRef]
- Ochi, K.; Okamoto, S.; Tozawa, Y.; Inaoka, T.; Hosaka, T.; Xu, J.; Kurosawa, K. Ribosome engineering and secondary metabolite production. Adv. Appl. Microbiol. 2004, 56, 155–184. [Google Scholar]
- Ochi, K. From microbial differentiation to ribosome engineering. Biosci.Biothenol. Biochem. 2007, 71, 1373–1386. [Google Scholar]
- Hosaka, T.; Ohnishi-Kameyama, M.; Muramatsu, H.; Murakami, K.; Tsurumi, Y.; Kodani, S.; Yoshida, M.; Fujie, A.; Ochi, K. Antibacterial discovery in actinomycetes strains with mutations in RNA polymerase or ribosomal protein S12. Nat. Biotechnol. 2009, 27, 462–464. [Google Scholar]
- Tian, C.K.; Cui, C.B.; Han, X.X. Isolation of fungal strains in unusual environment and screening for their antitumor activity. J. Int. Pharm. Res. 2008, 35, 401–405. [Google Scholar]
- Shao, C.L.; Wang, C.Y.; Wei, M.Y.; Gu, Y.C.; Xia, X.K.; She, Z.G.; Lin, Y.C. Structure elucidation of two new xanthone derivatives from the marine fungus Penicillium sp. (ZZF 32#) from the South China Sea. Magn. Reson. Chem. 2008, 46, 1066–1069. [Google Scholar]
- Arai, K.; Kimura, K.; Mushiroda, T.; Yamamoto, Y. Structures of Fructigenines A and B, new alkaloids isolated from Penicillium fructigenum TAKEUCHI. Chem. Pharm. Bull. 1989, 37, 2937–2939. [Google Scholar] [CrossRef]
- Shimada, A.; Kusano, M.; Takeuchi, S.; Fujioka, S.; Inokuchi, T.; Kimura, Y. Aspterric acid and 6-hydroxymellein, inhibitors of pollen development in Arabidopsis thaliana, produced by Aspergillus terreus. Z. Naturforsch. 2002, 57c, 459–464. [Google Scholar]
- Sankawa, U.; Ebizuka, Y.; Noguchi, H.; Isikawa, Y.; Kitaghawa, S.; Yamamoto, Y.; Kobayashi, T.; Irtak, Y. Biosynthesis of citrinin in Aspergillus terreus. Tetrahedron 1983, 39, 3583–3591. [Google Scholar] [CrossRef]
- Wu, C.J.; Cui, C.B.; Tian, C.K.; Li, C.W. Antitumor metabolites produced by two Penicillium purpurogenum G59 mutants. J. Int. Pharm. Res. 2010, 37, 122–126. [Google Scholar]
- Shima, J.; Hesketh, A.; Okamoto, S.; Kawamoto, S.; Ochi, K. Induction of Actinorhodin production by rpsL (encoding ribosomal protein S12) mutations that confer streptomycin resistance in Streptomyces lividans and Streptomyces coelicolor A3(2). J. Bacteriol. 1996, 178, 7276–7284. [Google Scholar]
- Hu, H.; Zhang, Q.; Ochi, K. Activation of antibiotic biosynthesis by specified mutations in the rpoB gene (endoding the RNA polymerase β subunit) of Streptomyces lividans. J. Bacteriol. 2002, 184, 3984–3991. [Google Scholar] [CrossRef]
- Lai, C.; Xu, J.; Tozawa, Y.; Okamoto-Hosoya, Y.; Yao, X.; Ochi, K. Genetic and physiological characterization of rpoB mutations that activate antibiotic production in Streptomyces lividans. Microbiology 2002, 148, 3365–3373. [Google Scholar]
- Inaoka, T.; Takahashi, K.; Yada, H.; Yoshida, M.; Ochi, K. RNA polymerase mutation activates the production of a dormant antibiotic 3,3'-neotrehalosadiamine via an autoinduction mechanism in Bacillus subtilis. J. Biol. Chem. 2004, 279, 3885–3892. [Google Scholar]
- Talà, A.; Wang, G.; Zemanova, M.; Okamoto, S.; Ochi, K.; Alifano, P. Activation of dormant bacterial genes by Nonomuraea sp. strain ATCC 39727 mutant-type RNA polymerase. J. Bacteriol. 2009, 191, 805–814. [Google Scholar]
- Shakil, S.; Khan, R.; Zarrilli, R.; Khan, A.U. Aminoglycosides versus bacteria—A description of the action, resistance mechanism, and nosocomial battleground. J. Biomed. Sci. 2008, 15, 5–14. [Google Scholar]
- Eustice, D.C.; Wilhelm, J.M. Fidelity of the eukaryotic condon-anticodon interaction: Interference by aminoglycoside antibiotics. Biochemistry 1984, 23, 1462–1467. [Google Scholar]
- Yoshizawa, S.; Fourmy, D.; Puglisi, J.D. Structural origins of gentamicin antibiotic action. EMBO J. 1998, 17, 6437–6448. [Google Scholar]
- Recht, M.I.; Douthwaite, S.; Puglist, J.D. Basis for prokaryotic specificity of action of aminoglycoside antibiotics. EMBO J. 1999, 18, 3133–3138. [Google Scholar]
- Lee, H.B.; Kim, Y.; Kim, J.C.; Choi, G.J.; Park, S.-H.; Kim, C.-J.; Jung, H.S. Activity of some aminoglycoside antibiotics against true fungi, Phytophthora and Pythium species. J. Appl. Microbiol. 2005, 99, 836–843. [Google Scholar] [CrossRef]
- Chang, J.-H.; Moon, H.-S. Synthesis and anti-inflammatory activity of fructigenine A derivatives. Biotechnol. Bioprocess Eng. 2004, 9, 59–61. [Google Scholar] [CrossRef]
- Xin, Z.H.; Zhu, W.M.; Gu, Q.Q.; Fang, Y.C.; Duan, L.; Cui, C.B. A new cytotoxic compound from Penicillium auratiogriseum, symbiotic or epiphytic fungus of sponge Mycale plumose. Chin. Chem. Lett. 2005, 16, 1227–1229. [Google Scholar]
- Yu, F.-Y.; Liao, Y.-C.; Chang, C.-H.; Liu, B.-H. Citrinin induces apoptosis in HL-60 cells via activation of the mitochondrial pathway. Toxicol. Lett. 2006, 161, 143–151. [Google Scholar]
- Chen, C.-C.; Chan, W.-H. Inhibition of citrinin-induced apoptotic biochemical signaling in human hepatoma G2 cells by resveratrol. Int. J. Mol. Sci. 2009, 10, 3338–3357. [Google Scholar]
- Chai, Y.J.; Cui, C.B.; Li, C.W.; Hua, W. Antitumor metabolites newly produced by a gentamicin-resistant mutant of Penicillium purpurogenum G59. J. Int. Pharm. Res. 2011, 38, 216–222. [Google Scholar]
- Pottz, G.E.; Rampey, J.H.; Benjamin, F. The effect of dimethyl sulfoxide (DMSO) on antibiotic sensitivity of a group of medically important microorganisms: Preliminary report. Ann. N. Y. Acad. Sci. 1967, 141, 261–272. [Google Scholar]
- Anchordoguy, T.J.; Carpenter, J.F.; Crowe, J.H.; Crowe, L.M. Temperature-dependent perturbation of phospholipid bilayers by dimethylsulfoxide. Biochim.Biophys. Acta 1992, 1104, 117–122. [Google Scholar] [CrossRef]
- Chen, G.; Wang, G.-Y.-S.; Li, X.; Wters, B.; Davies, J. Enhanced production of microbial metabolites in the presence of dimethyl sulphoxide. J. Antibiot. 2000, 53, 1145–1153. [Google Scholar]
- Yu, Z.-W.; Quinn, P.J. The modulation of membrane structure and stability by dimethyl sulphoxide (Review). Mol. Membr. Biol. 1998, 15, 59–68. [Google Scholar]
- Notman, R.; Noro, M.; O’Malley, B.; Anwar, J. Molecular basis for dimethylsulfoxide (DMSO) action on lipid membranes. J. Am. Chem. Soc. 2006, 128, 13982–13983. [Google Scholar]
Supplementary Files
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chai, Y.-J.; Cui, C.-B.; Li, C.-W.; Wu, C.-J.; Tian, C.-K.; Hua, W. Activation of the Dormant Secondary Metabolite Production by Introducing Gentamicin-Resistance in a Marine-Derived Penicillium purpurogenum G59. Mar. Drugs 2012, 10, 559-582. https://doi.org/10.3390/md10030559
Chai Y-J, Cui C-B, Li C-W, Wu C-J, Tian C-K, Hua W. Activation of the Dormant Secondary Metabolite Production by Introducing Gentamicin-Resistance in a Marine-Derived Penicillium purpurogenum G59. Marine Drugs. 2012; 10(3):559-582. https://doi.org/10.3390/md10030559
Chicago/Turabian StyleChai, Yun-Jing, Cheng-Bin Cui, Chang-Wei Li, Chang-Jing Wu, Cong-Kui Tian, and Wei Hua. 2012. "Activation of the Dormant Secondary Metabolite Production by Introducing Gentamicin-Resistance in a Marine-Derived Penicillium purpurogenum G59" Marine Drugs 10, no. 3: 559-582. https://doi.org/10.3390/md10030559
APA StyleChai, Y.-J., Cui, C.-B., Li, C.-W., Wu, C.-J., Tian, C.-K., & Hua, W. (2012). Activation of the Dormant Secondary Metabolite Production by Introducing Gentamicin-Resistance in a Marine-Derived Penicillium purpurogenum G59. Marine Drugs, 10(3), 559-582. https://doi.org/10.3390/md10030559