Cellular and Transcriptional Responses of Crassostrea gigas Hemocytes Exposed in Vitro to Brevetoxin (PbTx-2)

Abstract
:1. Introduction
2. Results
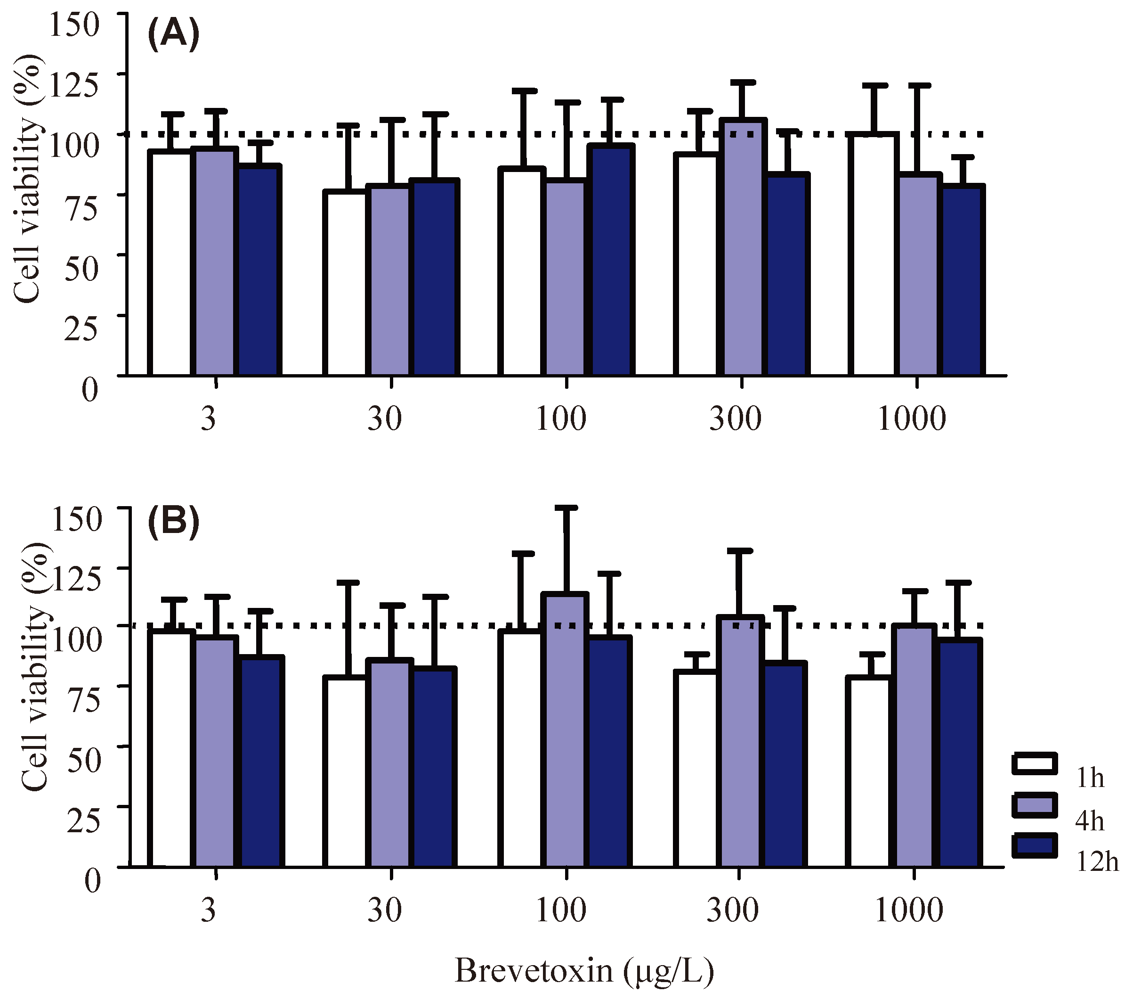
2.1. Cellular Parameters

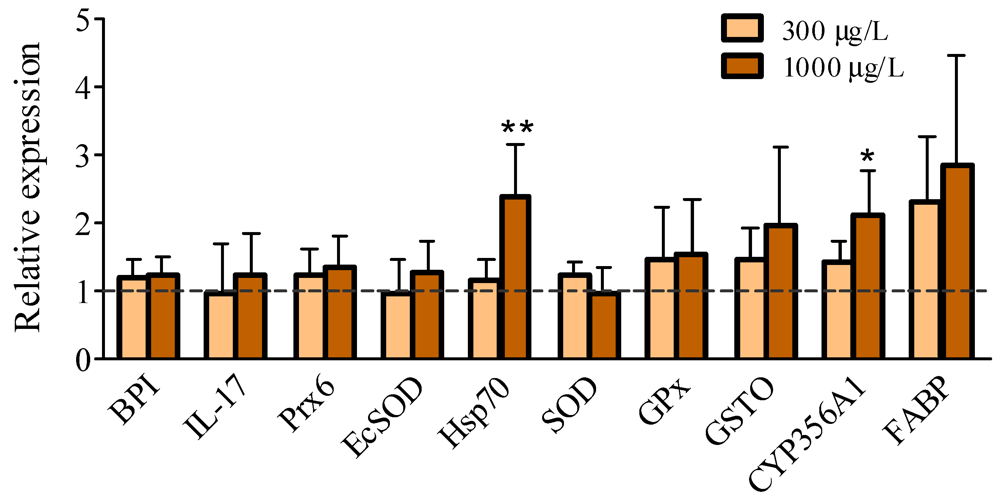
2.2. Gene Expression Levels

3. Discussion
4. Experimental Section
4.1. Animals
4.2. Hemolymph Extraction and Hemocyte Incubation with Brevetoxin (PbTx-2)
4.3. Cellular Viability Assays and Apoptosis
4.4. Extraction of Total RNA and cDNA Synthesis
4.5. Real-Time Quantitative PCR (qPCR)

{kind=link}
{kind=link}
| Gene Name | Primers | Sequence 5'-3' | GenBank |
|---|---|---|---|
| Glyceraldehyde 3-phosphate dehydrogenase | CgGAPDH-Fw | GCTGTGACACCATTGGAGAA | AJ544886.1 |
| CgGAPDH-Rv | ACCAATGACGCAACAAGCGA | ||
| Peroxiredoxin 6 | CgPrx6-Fw | GAGCCAGAGTTCAAGAAGAG | AM265552.1 |
| CgPrx6-Rv | TGCATTGTCCTTTTCGGCTG | ||
| Interleukin-17 | CgIL17-Fw | ACTGAGGCTCGATGCAAGTG | EF190193.1 |
| CgIL17-Rv | AGCCTTCTTGCTTCATGTGG | ||
| Bactericidal/permeability | CgBPI-Fw | GATAGAAATAGGAATGGACGG | HM992925.1 |
| increasing protein | CgBPI-Rv | GTTATAGATCCACGCTGCTCC | |
| Heat shock protein 70 | CgHsp70-Fw | TCATCAAGTGGATGGACCAG | AB122063.1 |
| CgHsp70-Rv | CATTCCTCCAGGCATGCCA | ||
| Extracellular superoxide dismutase | CgEcSOD-Fw | GCTGTGACACCATTGGAGAA | DQ010420.1 |
| CgEcSOD-Rv | ACCAATGACGCAACAAGCGA | ||
| Glutathione S-transferase omega | CgGSTO-Fw | TGATGAGTTCACCACCGCAA | AJ557141.1 |
| CgGSTO-Rv | TTCAAACCATGGCCACAGCA | ||
| Cytochrome P450 isoform 356A1 | CgCYP3561A-Fw | ATGAAACCCGCGAAACCAGA | EF645271.1 |
| CgCYP3561A-Rv | TAAATTCGGCTTCACGCCCT | ||
| Superoxide dismutase | CgSOD-Fw | TCAACAAAGAGCATGGCGTC | AJ496219.1 |
| CgSOD-Rv | TTTCCGGTCGTCTTACTGAG | ||
| Glutathione peroxidase | CgGPx-Fw | TCAAGATCCGAGATGTCGTC | EF692639.1 |
| CgGPx-Rv | ACTCGGTTTCCAGACATGAG | ||
| Fatty acid binding protein | CgFABP-Fw | GTTTGAGGGAAACTGGGAATGC | EU069496 |
| CgFABP-Rv | TCCGTCGGAATATGTCAGTTTAGC |
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Wang, D.Z. Neurotoxins from marine dinoflagellates: A brief review. Mar Drugs 2008, 6, 349–371. [Google Scholar]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar]
- Brand, L.E.; Campbell, L.; Bresnan, E. KARENIA: The biology and ecology of a toxic genus. Harmful Algae 2011, 14, 156–178. [Google Scholar]
- Plakas, S.M.; Jester, E.L.E.; El Said, K.R.; Granade, H.R.; Abraham, A.; Dickey, R.W.; Scott, P.S.; Flewelling, L.J.; Henry, M.; Blum, P.; et al. Monitoring of brevetoxins in the Karenia brevis bloom-exposed Eastern oyster (Crassostrea virginica). Toxicon 2008, 52, 32–38. [Google Scholar] [CrossRef]
- Plakas, S.M.; Dickey, R.W. Advances in monitoring and toxicity assessment of brevetoxins in molluscan shellfish. Toxicon 2010, 56, 137–149. [Google Scholar]
- Shumway, S.E. A review of the effects of algal blooms on shellfish and aquaculture. J. World Aquac. Soc. 1990, 21, 65–104. [Google Scholar]
- Leverone, J.R.; Shumway, S.E.; Blake, N.J. Comparative effects of the toxic dinoflagellate Karenia brevis on clearance rates in juveniles of four bivalve molluscs from Florida, USA. Toxicon 2007, 49, 634–645. [Google Scholar]
- Leverone, J.R.; Blake, N.J.; Pierce, R.H.; Shumway, S.E. Effects of the dinoflagellate Karenia brevis on larval development in three species of bivalve mollusc from Florida. Toxicon 2006, 48, 75–84. [Google Scholar] [CrossRef]
- Wang, L.P.; Yan, T.; Zhou, M.J. Impacts of HAB species Heterosigma akashiwo on early development of the scallop Argopecten irradians Lamarck. Aquaculture 2006, 255, 374–383. [Google Scholar] [CrossRef]
- Keppler, C.J.; Lewitus, A.J.; Ringwood, A.H.; Hoguet, J.; Staton, T. Sublethal cellular effects of short-term raphidophyte and brevetoxin exposures on the eastern oyster Crassostrea virginica. Mar. Ecol. Prog. Ser. 2006, 312, 141–147. [Google Scholar] [CrossRef]
- Keppler, C.J.; Hoguet, J.; Smith, K.; Ringwood, A.H.; Lewitus, A.J. Sublethal effects of the toxic alga Heterosigma akashiwo on the southeastern oyster (Crassostrea virginica). Harmful Algae 2005, 4, 275–285. [Google Scholar] [CrossRef]
- Khan, S.; Arakawa, O.; Onoue, Y. Neurotoxins in a toxic red tide of Heterosigma akashiwo (Raphidophyceae) in Kagoshima Bay, Japan. Aquac. Res. 1997, 28, 9–14. [Google Scholar] [CrossRef]
- Hégaret, H.; da Silva, P.M.; Wikfors, G.H.; Haberkorn, H.; Shumway, S.E.; Soudant, P. In vitro interactions between several species of harmful algae and haemocytes of bivalve molluscs. Cell Biol. Toxicol. 2011, 27, 249–266. [Google Scholar] [CrossRef]
- Hégaret, H.; Smolowitz, R.M.; Sunila, I.; Shumway, S.E.; Alix, J.; Dixon, M.; Wikfors, G.H. Combined effects of a parasite, QPX, and the harmful-alga, Prorocentrum minimum on northern quahogs, Mercenaria mercena. Mar. Environ. Res. 2010, 69, 337–344. [Google Scholar] [CrossRef]
- da Silva, P.M.; Hégaret, H.; Lambert, C.; Wikfors, G.H.; Le Goic, N.; Shumway, S.E.; Soudant, P. Immunological responses of the Manila clam (Ruditapes philippinarum) with varying parasite (Perkinsus olseni) burden, during a long-term exposure to the harmful alga, Karenia selliformis, and possible interactions. Toxicon 2008, 51, 563–573. [Google Scholar] [CrossRef]
- Galimany, E.; Sunila, I.; Hégaret, H.; Ramon, M.; Wikfors, G.H. Pathology and immune response of the blue mussel (Mytilus edulis L.) after an exposure to the harmful dinoflagellate Prorocentrum minimum. Harmful Algae 2008, 7, 630–638. [Google Scholar] [CrossRef]
- Galimany, E.; Sunila, I.; Hégaret, H.; Ramon, M.; Wikfors, G.H. Experimental exposure of the blue mussel (Mytilus edulis, L.) to the toxic dinoflagellate Alexandrium fundyense: Histopathology, immune responses, and recovery. Harmful Algae 2008, 7, 702–711. [Google Scholar] [CrossRef]
- Hégaret, H.; da Silva, P.M.; Wikfors, G.H.; Lambert, C.; De Bettignies, T.; Shumway, S.E.; Soudant, P. Hemocyte responses of Manila clams, Ruditapes philippinarum, with varying parasite, Perkinsus olseni, severity to toxic-algal exposures. Aquat. Toxicol. 2007, 84, 469–479. [Google Scholar] [CrossRef]
- Hégaret, H.; Shumway, S.E.; Wikfors, G.H. Effects of harmful algae on physiology and hemocyte parameters of the Northern Bay scallop, Argopecten irradians irradians. J. Shellfish Res. 2007, 26, 1316–1316. [Google Scholar]
- Hégaret, H.; Wikfors, G.H. Time-dependent changes in hemocytes of eastern oysters, Crassostrea virginica, and northern bay scallops, Argopecten irradians irradians, exposed to a cultured strain of Prorocentrum minimum. Harmful Algae 2005, 4, 187–199. [Google Scholar] [CrossRef]
- Hégaret, H.; Wikfors, G.H. Effects of natural and field-simulated blooms of the dinoflagellate Prorocentrum minimum upon hemocytes of eastern oysters, Crassostrea virginica, from two different populations. Harmful Algae 2005, 4, 201–209. [Google Scholar]
- Mello, D.F.; Proença, L.A.O.; Barracco, M.A. Comparative study of various immune parameters in three bivalve species during a natural bloom of Dinophysis acuminata in Santa Catarina Island, Brazil. Toxins 2010, 2, 1166–1178. [Google Scholar] [CrossRef]
- Dizer, H.; Fischer, B.; Harabawy, A.S.A.; Hennion, M.C.; Hansen, P.D. Toxicity of domoic acid in the marine mussel Mytilus edulis. Aquat. Toxicol. 2001, 55, 149–156. [Google Scholar] [CrossRef]
- Jones, T.O.; Whyte, J.N.C.; Ginther, N.G.; Townsend, L.D.; Iwama, G.K. Hemocyte changes in the pacific oyster, Crassostrea gigas, caused by exposure to domoic acid in the diatom Pseudonitzschia pungens f. multiseries. Toxicon 1995, 33, 347–353. [Google Scholar] [CrossRef]
- Song, L.; Wang, L.; Qiu, L.; Zhang, H. Bivalve Immunity. In Invertebrate Immunity; Soderhall, K., Ed.; Springer: New York, NY, USA, 2010; Volume 708, p. 316. [Google Scholar]
- Hedgecock, D.; Gaffney, P.; Goulletquer, P.; Guo, X.; Reece, K.; Warr, G. The case for sequencing the Pacific oyster genome. J. Shellfish Res. 2005, 24, 429–441. [Google Scholar]
- Jose, S.; Mohandas, A.; Philip, R.; Singh, I.S.B. Primary hemocyte culture of Penaeus monodon as an in vitro model for white spot syndrome virus titration, viral and immune related gene expression and cytotoxicity assays. J. Invertebr. Pathol. 2010, 105, 312–321. [Google Scholar] [CrossRef]
- Araya, M.T.; Markham, F.; Mateo, D.R.; McKenna, P.; Johnson, G.R.; Berthe, F.C.J.; Siah, A. Identification and expression of immune-related genes in hemocytes of soft-shell clams, Mya arenaria, challenged with Vibrio splendidus. Fish Shellfish Immunol. 2010, 29, 557–564. [Google Scholar] [CrossRef]
- Medeiros, I.D.; Siebert, M.N.; Toledo-Silva, G.; Rodrigues, T.B.; Marques, M.R.F.; Bainy, A.C.D. Induced gene expression in oyster Crassostrea gigas exposed to sewage. Environ. Toxicol. Pharmacol. 2008, 26, 362–365. [Google Scholar] [CrossRef]
- Trevisan, R.; Flesch, S.; Mattos, J.J.; Milani, M.R.; Bainy, A.C.D.; Dafre, A.L. Zinc exposure targets peroxiredoxin 6, thioredoxin reductase and glutathione metabolism in brown mussel Perna perna gills. Comp. Biochem. Physiol. to be submitted..
- Green, T.J.; Dixon, T.J.; Devic, E.; Adlard, R.D.; Barnes, A.C. Differential expression of genes encoding anti-oxidant enzymes in Sydney rock oysters, Saccostrea glomerata (Gould) selected for disease resistance. Fish Shellfish Immunol. 2009, 26, 799–810. [Google Scholar] [CrossRef]
- David, E.; Tanguy, A.; Moraga, D. Peroxiredoxin 6 gene: A new physiological and genetic indicator of multiple environmental stress response in Pacific oyster Crassostrea gigas. Aquat. Toxicol. 2007, 84, 389–398. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar]
- Paillard, C.; AshtonAlcox, K.A.; Ford, S.E. Changes in bacterial densities and hemocyte parameters in eastern oysters, Crassostrea virginica, affected by juvenile oyster disease. Aquat. Living Resour. 1996, 9, 145–158. [Google Scholar]
- Allam, B.; Paillard, C. Defense factors in clam extrapallial fluids. Dis. Aquat. Org. 1998, 33, 123–128. [Google Scholar]
- Ford, S.E.; Bricelj, V.M.; Lambert, C.; Paillard, C. Deleterious effects of a nonPST bioactive compound(s) from Alexandrium tamarense on bivalve hemocytes. Mar. Biol. 2008, 154, 241–253. [Google Scholar] [CrossRef]
- Franchini, A.; Milandri, A.; Poletti, R.; Ottaviani, E. Immunolocalization of yessotoxins in the mussel Mytilus galloprovincialis. Toxicon 2003, 41, 967–970. [Google Scholar] [CrossRef]
- Hégaret, H.; Wikfors, G.H.; Soudant, P.; Lambert, C.; Shumway, S.E.; Berard, J.B.; Lassus, P. Toxic dinoflagellates (Alexandrium fundyense and A-catenella) have minimal apparent effects on oyster hemocytes. Mar. Biol. 2007, 152, 441–447. [Google Scholar] [CrossRef]
- Jackson, S.A.; Uhlinger, K.R.; Clegg, J.S. Duration of induced thermal tolerance and tissue-specific expression of hsp/hsc70 in the eastern oyster, Crassostrea virginica and the pacific oyster, Crassostrea gigas. Aquaculture 2011, 317, 168–174. [Google Scholar] [CrossRef]
- Clegg, J.S.; Uhlinger, K.R.; Jackson, S.A.; Cherr, G.N.; Rifkin, E.; Friedman, C.S. Induced thermotolerance and the heat shock protein-70 family in the Pacific oyster Crassostrea gigas. Mol. Mar. Biol. Biotechnol. 1998, 7, 21–30. [Google Scholar]
- Shamseldin, A.A.; Clegg, J.S.; Friedman, C.S.; Cherr, G.N.; Pillai, M.C. Induced thermotolerance in the Pacific oyster, Crassostrea gigas. J. Shellfish Res. 1997, 16, 487–491. [Google Scholar]
- Farcy, E.; Voiseux, C.; Lebel, J.M.; Fievet, B. Transcriptional expression levels of cell stress marker genes in the Pacific oyster Crassostrea gigas exposed to acute thermal stress. Cell Stress Chaperones 2009, 14, 371–380. [Google Scholar] [CrossRef]
- Hamdoun, A.M.; Cheney, D.P.; Cherr, G.N. Phenotypic plasticity of hsp70 and hsp70 gene expression in the Pacific oyster (Crassostrea gigas): Implications for thermal limits and induction of thermal tolerance. Biol. Bull. 2003, 205, 160–169. [Google Scholar] [CrossRef]
- Clayton, M.E.; Steinmann, R.; Fent, K. Different expression patterns of heat shock proteins hsp 60 and hsp 70 in zebra mussels (Dreissena polymorpha) exposed to copper and tributyltin. Aquat. Toxicol. 2000, 47, 213–226. [Google Scholar] [CrossRef]
- Boutet, I.; Tanguy, A.; Moraga, D. Response of the Pacific oyster Crassostrea gigas to hydrocarbon contamination under experimental conditions. Gene 2004, 329, 147–157. [Google Scholar] [CrossRef]
- Song, L.; Wu, L.; Ni, D.; Chang, Y.; Xu, W.; Xing, K. The cDNA cloning and mRNA expression of heat shock protein 70 gene in the haemocytes of bay scallop (Argopecten irradians, Lamarck 1819) responding to bacteria challenge and naphthalin stress. Fish Shellfish Immunol. 2006, 21, 335–345. [Google Scholar] [CrossRef]
- Piano, A.; Valbonesi, P.; Fabbri, E. Expression of cytoprotective proteins, heat shock protein 70 and metallothioneins, in tissues of Ostrea edulis exposed to heat and heavy metals. Cell Stress Chaperones 2004, 9, 134–142. [Google Scholar] [CrossRef]
- La Porte, P. Mytilus trossulus hsp70 as a biomarker for arsenic exposure in the marine environment: Laboratory and real-world results. Biomarkers 2005, 10, 417–428. [Google Scholar] [CrossRef]
- Mićović, V.; Bulog, A.; Kučić, N.; Jakovac, H.; Radošević-Stašić, B. Metallothioneins and heat shock proteins 70 in marine mussels as sensors of environmental pollution in Northern Adriatic Sea. Environ. Toxicol. Pharmacol. 2009, 28, 439–447. [Google Scholar]
- Hamer, B.; Hamer, D.P.; Müller, W.E.G.; Batel, R. Stress-70 proteins in marine mussel Mytilus galloprovincialis as biomarkers of environmental pollution: A field study. Environ. Int. 2004, 30, 873–882. [Google Scholar] [CrossRef]
- Tirard, C.T.; Grossfeld, R.M.; Volety, A.K.; Chu, F.L.E. Heat-shock proteins of the oyster parasite perkinsus-marinus. Dis. Aquat. Org. 1995, 22, 147–151. [Google Scholar]
- Encomio, V.G.; Chu, F.L.E. Seasonal variation of heat shock protein 70 in eastern oysters (Crassostrea virginica) infected with Perkinsus marinus (Dermo). J. Shellfish Res. 2005, 24, 167–175. [Google Scholar]
- Cellura, C.; Toubiana, M.; Parrinello, N.; Roch, P. Specific expression of antimicrobial peptide and HSP70 genes in response to heat-shock and several bacterial challenges in mussels. Fish Shellfish Immunol. 2007, 22, 340–350. [Google Scholar]
- Fabbri, E.; Valbonesi, P.; Franzellitti, S. HSP expression in bivalves. Invertebr. Surviv. J. 2008, 5, 135–161. [Google Scholar]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef]
- Tedengren, M.; Olsson, B.; Reimer, O.; Brown, D.C.; Bradley, B.P. Heat pretreatment increases cadmium resistance and HSP 70 levels in Baltic Sea mussels. Aquat. Toxicol. 2000, 48, 1–12. [Google Scholar]
- Snyder, M.J. Cytochrome P450 enzymes in aquatic invertebrates: Recent advances and future directions. Aquat. Toxicol. 2000, 48, 529–547. [Google Scholar]
- de Toledo-Silva, G.; Siebert, M.N.; Medeiros, I.D.; Sincero, T.C.M.; Moraes, M.O.; Goldstone, J.V.; Bainy, A.C.D. Cloning a new cytochrome P450 isoform (CYP356A1) from oyster Crassostrea gigas. Mar. Environ. Res. 2008, 66, 15–18. [Google Scholar] [CrossRef]
- Guo, F.; An, T.; Rein, K.S. Human metabolites of brevetoxin PbTx-2: Identification and confirmation of structure. Toxicon 2010, 56, 648–651. [Google Scholar]
- Radwan, F.F.Y.; Ramsdell, J.S. Characterization of in vitro oxidative and conjugative metabolic pathways for brevetoxin (PbTx-2). Toxicol. Sci. 2006, 89, 57–65. [Google Scholar]
- Washburn, B.S.; Baden, D.G.; Gassman, N.J.; Walsh, P.J. Brevetoxin: Tissue distribution and effect on cytochrome P450 enzymes in fish. Toxicon 1994, 32, 799–805. [Google Scholar]
- Plakas, S.M.; Wang, Z.H.; El Said, K.R.; Jester, E.L.E.; Granade, H.R.; Flewelling, L.; Scott, P.; Dickey, R.W. Brevetoxin metabolism and elimination in the Eastern oyster (Crassostrea virginica) after controlled exposures to Karenia brevis. Toxicon 2004, 44, 677–685. [Google Scholar] [CrossRef]
- Plakas, S.M.; El Said, K.R.; Jester, E.L.E.; Granade, H.R.; Musser, S.M.; Dickey, R.W. Confirmation of brevetoxin metabolism in the Eastern oyster (Crassostrea virginica) by controlled exposures to pure toxins and to Karenia brevis cultures. Toxicon 2002, 40, 721–729. [Google Scholar] [CrossRef]
- Livingstone, D.R. The fate of organic xenobiotics in aquatic ecosystems: Quantitative and qualitative differences in biotransformation by invertebrates and fish. Comp. Biochem. Physiol A 1998, 120, 43–49. [Google Scholar]
- Esteves, A.; Ehrlich, R. Invertebrate intracellular fatty acid binding proteins. Comp. Biochem. Physiol. C 2006, 142, 262–274. [Google Scholar]
- Velkov, T.; Chuang, S.; Wielens, J.; Sakellaris, H.; Charman, W.N.; Porter, C.J.H.; Scanlon, M.J. The interaction of lipophilic drugs with intestinal fatty acid-binding protein. J. Biol. Chem. 2005, 280, 17769–17776. [Google Scholar]
- Gonzalez, M.; Gueguen, Y.; Destoumieux-Garzon, D.; Romestand, B.; Fievet, J.; Pugniere, M.; Roquet, F.; Escoubas, J.M.; Vandenbulcke, F.; Levy, O.; et al. Evidence of a bactericidal permeability increasing protein in an invertebrate, the Crassostrea gigas Cg-BPI. Proc. Natl. Acad. Sci. USA 2007, 104, 17759–17764. [Google Scholar]
- Roberts, S.; Gueguen, Y.; de Lorgeril, J.; Goetz, F. Rapid accumulation of an interleukin 17 homolog transcript in Crassostrea gigas hemocytes following bacterial exposure. Dev. Comp. Immunol. 2008, 32, 1099–1104. [Google Scholar] [CrossRef]
- Gonzalez, M.; Romestand, B.; Fievet, J.; Huvet, A.; Lebart, M.-C.; Gueguen, Y.; Bachère, E. Evidence in oyster of a plasma extracellular superoxide dismutase which binds LPS. Biochem. Biophys. Res. Commun. 2005, 338, 1089–1097. [Google Scholar]
- Robinson, M.W.; Hutchinson, A.T.; Dalton, J.P.; Donnelly, S. Peroxiredoxin: A central player in immune modulation. Parasite Immunol. 2010, 32, 305–313. [Google Scholar]
- Jo, P.G.; Choi, Y.K.; Choi, C.Y. Cloning and mRNA expression of antioxidant enzymes in the Pacific oyster, Crassostrea gigas in response to cadmium exposure. Comp. Biochem. Physiol. C 2008, 147, 460–469. [Google Scholar]
- Trevisan, R.; Arl, M.; Sacchet, C.L.; Engel, C.S.; Danielli, N.M.; Mello, D.F.; Brocardo, C.; Maris, A.F.; Dafre, A.L. Antioxidant deficit in gills of Pacific oyster (Crassostrea gigas) exposed to chlorodinitrobenzene increases menadione toxicity. Aquat. Toxicol. 2012, 108, 85–93. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/nuccore (accessed on 12 August 2011).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29. [Google Scholar]
- StatSoft, version 7.0, StatSoft Inc.: Tulsa, OK, USA, 2004.
- Sample Availability: Available from the authors.
Supplementary Files
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mello, D.F.; De Oliveira, E.S.; Vieira, R.C.; Simoes, E.; Trevisan, R.; Dafre, A.L.; Barracco, M.A. Cellular and Transcriptional Responses of Crassostrea gigas Hemocytes Exposed in Vitro to Brevetoxin (PbTx-2). Mar. Drugs 2012, 10, 583-597. https://doi.org/10.3390/md10030583
Mello DF, De Oliveira ES, Vieira RC, Simoes E, Trevisan R, Dafre AL, Barracco MA. Cellular and Transcriptional Responses of Crassostrea gigas Hemocytes Exposed in Vitro to Brevetoxin (PbTx-2). Marine Drugs. 2012; 10(3):583-597. https://doi.org/10.3390/md10030583
Chicago/Turabian StyleMello, Danielle F., Eliza S. De Oliveira, Renato C. Vieira, Erik Simoes, Rafael Trevisan, Alcir Luiz Dafre, and Margherita Anna Barracco. 2012. "Cellular and Transcriptional Responses of Crassostrea gigas Hemocytes Exposed in Vitro to Brevetoxin (PbTx-2)" Marine Drugs 10, no. 3: 583-597. https://doi.org/10.3390/md10030583
APA StyleMello, D. F., De Oliveira, E. S., Vieira, R. C., Simoes, E., Trevisan, R., Dafre, A. L., & Barracco, M. A. (2012). Cellular and Transcriptional Responses of Crassostrea gigas Hemocytes Exposed in Vitro to Brevetoxin (PbTx-2). Marine Drugs, 10(3), 583-597. https://doi.org/10.3390/md10030583