
Long-Term Feeding of Chitosan Ameliorates Glucose and Lipid Metabolism in a High-Fructose-Diet-Impaired Rat Model of Glucose Tolerance



Abstract
:
1. Introduction
2. Results
2.1. Effects of Chitosan on Body Weight and Tissue Weight in HF-Diet-Fed Rats


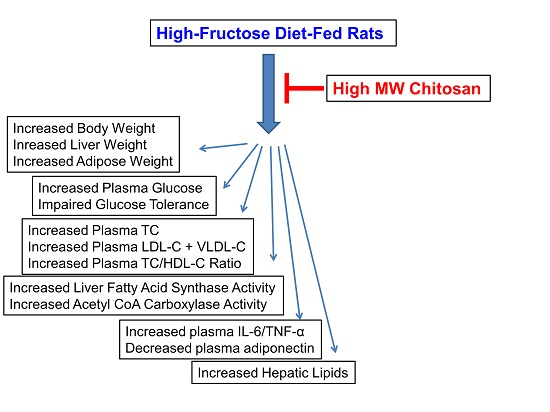{kind=link}
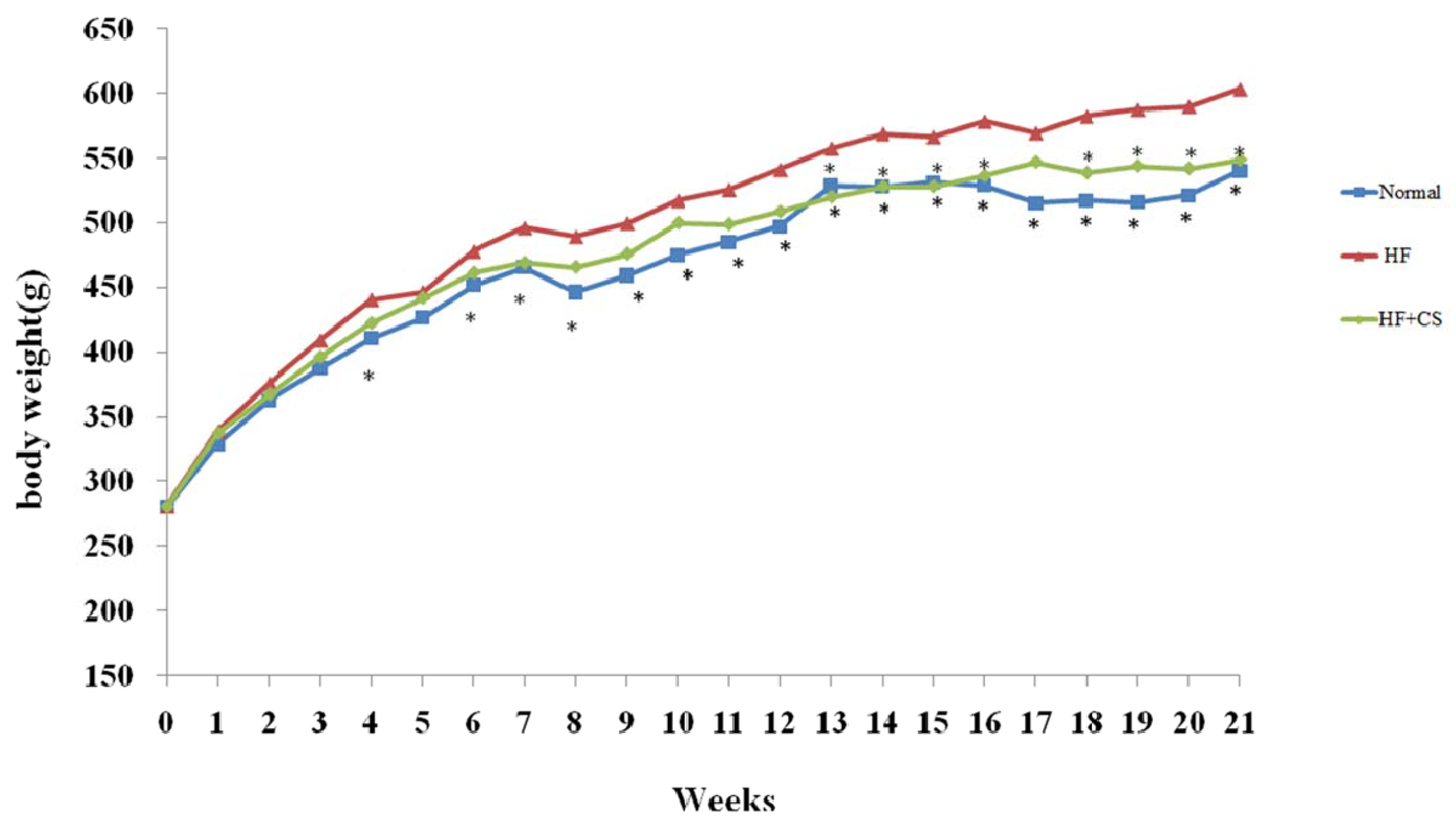{kind=link}
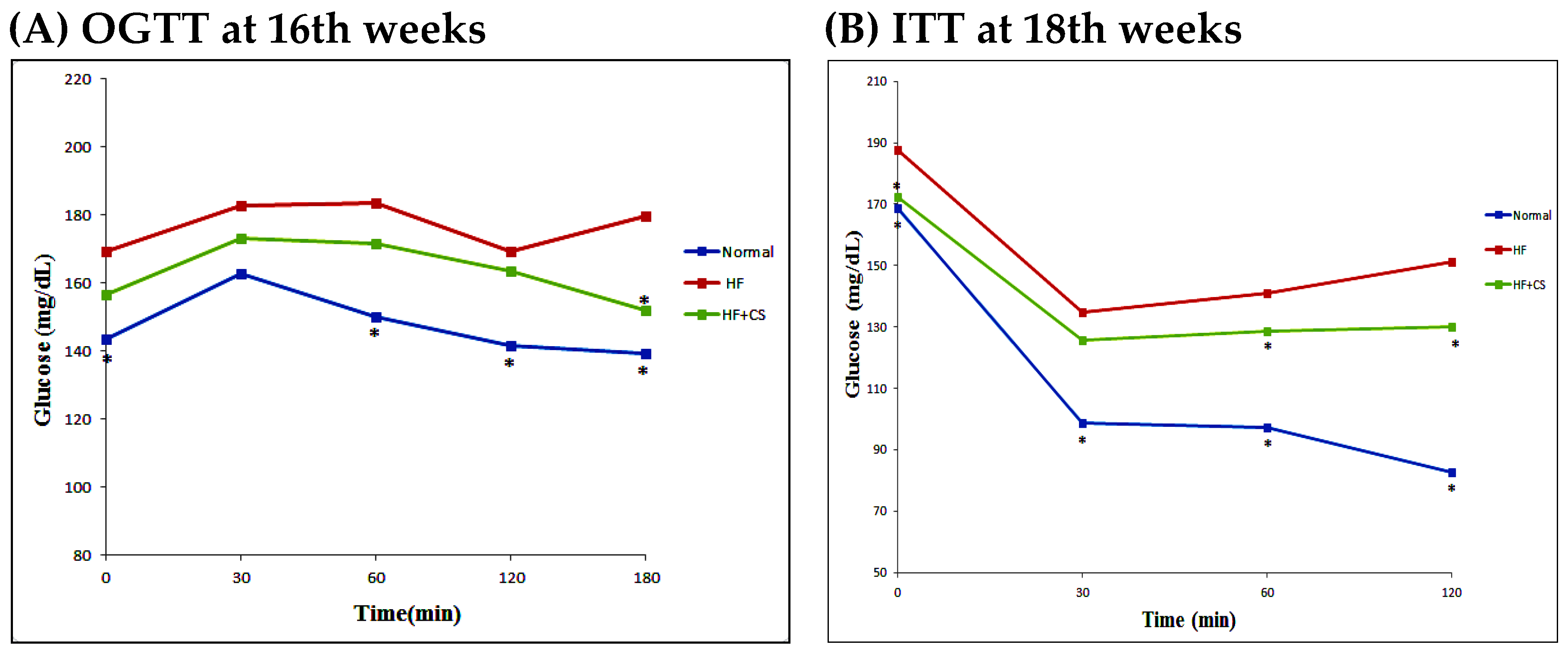{kind=link}
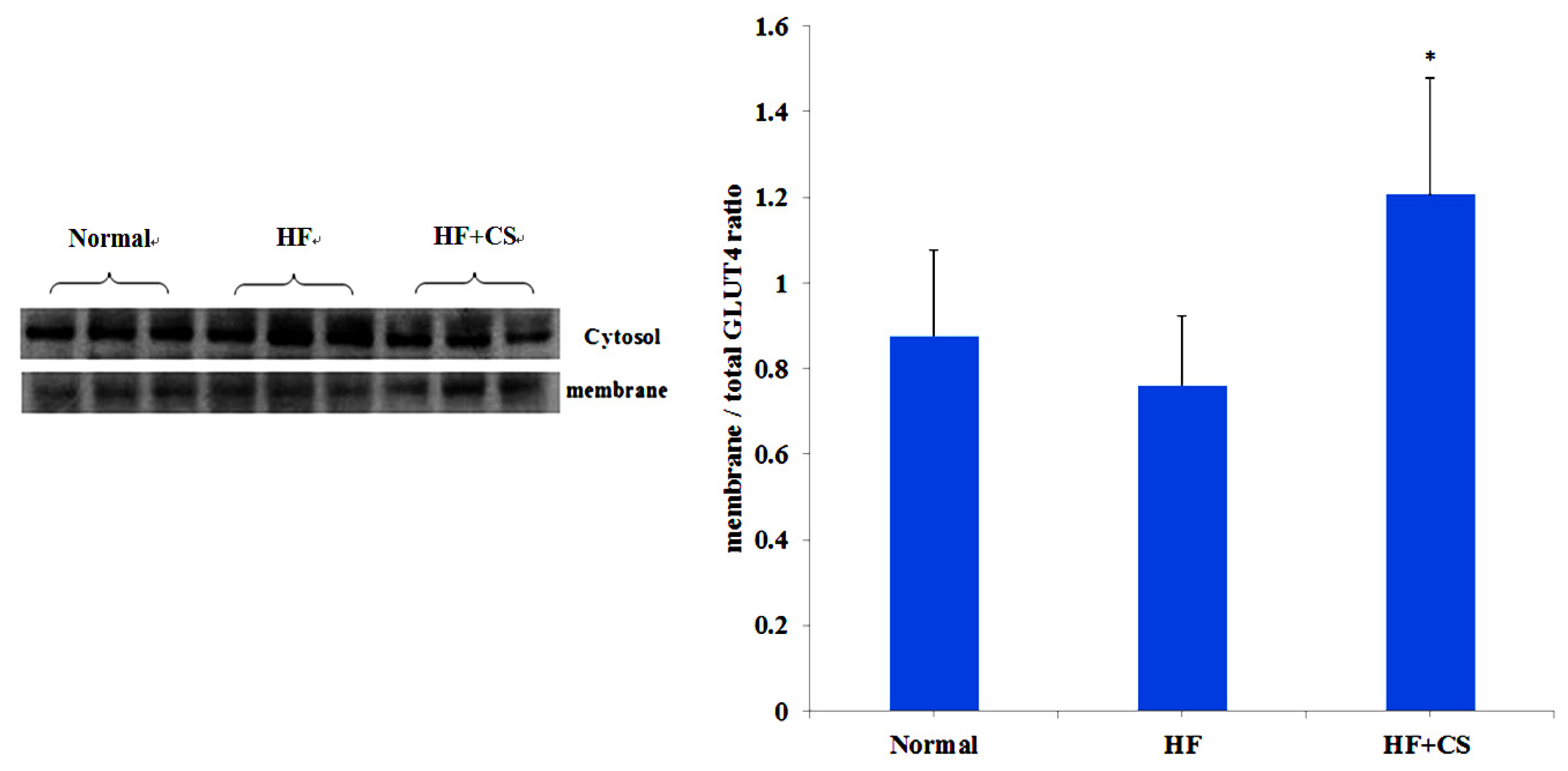{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue Weight | Groups | ||
|---|---|---|---|
| Normal | HF | HF + CS | |
| Liver weight (g) | 28.1 ± 4.1 * | 35.7 ± 6.3 | 23.7 ± 5.2 * |
| Relative liver weight (g/100 g BW) | 5.3 ± 0.6 * | 6.1 ± 0.9 | 4.4 ± 0.7 * |
| Adipose tissue weight(g) | 21.4 ± 5.1 * | 28.6 ± 8.4 | 19.2 ± 5.6 * |
| Relative adipose tissue weight (g/100 g BW) | 4.0 ± 1.0 | 4.9 ± 1.2 | 3.6 ± 1.1 * |
| Retroperitoneal adipose weight(g) | 13.9 ± 3.3 * | 18.7 ± 5.7 | 12.5 ± 4.0 * |
| Relative retroperitoneal adipose weight (g/100 g BW) | 2.6 ± 0.6 | 3.2 ± 0.8 | 2.4 ± 0.8 * |
| Epididymal adipose weight(g) | 7.5 ± 2.3 | 9.9 ± 2.9 | 7.1 ± 2.2 * |
| Relative epididymal adipose weight (g/100 g BW) | 1.4 ± 0.4 | 1.7 ± 0.5 | 1.3± 0.4 * |
2.2. Effects of Chitosan on Plasma Glucose and Lipid Metabolism in HF-Diet-Fed Rats
| Parameters | Groups | ||
|---|---|---|---|
| Normal | HF | HF + CS | |
| Glucose (mg/dL) | 187.0 ± 7.6 * | 215.9 ± 32.6 | 191.3 ± 19.1 |
| Insulin (μg/L) | 0.9 ± 0.3 | 0.8 ± 0.2 | 0.6 ± 0.2 * |
| Total cholesterol (mg/dL) | 193.4 ± 31.0 * | 288.2 ± 110.5 | 177.4 ± 34.6 * |
| HDL-C (mg/dL) | 53.6 ± 8.3 | 51.2 ± 17.4 | 63.6 ± 17.8 |
| LDL-C + VLDL-C (mg/dL) | 147.4 ± 35.6 * | 231.8 ± 102.1 | 113.8 ± 26.2 * |
| TC/HDL-C | 3.9 ± 0.9 * | 5.1 ± 1.3 | 3.0 ± 0.3 * |
| HDL-C/(LDL-C + VLDL-C) | 0.4 ± 0.2 | 0.3 ± 0.1 | 0.5 ± 0.1 * |
| Triglyceride (mg/dL) | 60.9 ± 11.4 | 65.0 ± 6.0 | 92.2 ± 26.0 * |



| Parameters | Groups | ||
|---|---|---|---|
| Normal | HF | HF + CS | |
| TNF-α (pg/mL) | 16.4 ± 1.0 * | 20.6 ± 4.4 | 15.0 ± 4.2 * |
| IL-6 (pg/mL) | 19.3 ± 1.7 * | 23.3 ± 2.9 | 20.4 ± 2.9 * |
| Adiponectin (μg/mL) | 16.5 ± 2.5 * | 8.00 ± 2.8 | 11.5 ± 3.1 * |
| Leptin (ng/mL) | 6.5 ± 2.0 | 9.8 ± 3.5 | 4.68 ± 1.5 * |
| Free fatty acids | 1.1 ±0.3 | 1.1 ±0.2 | 1.4 ±0.3 * |
| Hepatic Lipids | Groups | ||
|---|---|---|---|
| Normal | HF | HF + CS | |
| Total Cholesterol | |||
| (mg/g liver) | 83.6 ± 14.5 * | 121.0 ± 19.3 | 62.5 ± 17.0 * |
| (g/liver) | 2.4 ± 0.5 * | 4.3 ± 1.0 | 1.1 ± 0.4 * |
| Triglyceride | |||
| (mg/g liver) | 43.7 ± 6.8 * | 57.9 ± 16.0 | 30.4 ± 11. * |
| (g/liver) | 1.2 ± 0.3 * | 2.0 ± 0.6 | 0.8 ± 0.2 * |
| Fecal Weight and Lipids | Groups | ||
|---|---|---|---|
| Normal | HF | HF + CS | |
| Feces wet weight (g/day) | 2.2 ± 0.3 | 2.3 ± 0.3 | 2.4 ± 0.4 |
| Feces dry weight (g/day) | 1.9 ± 0.3 | 2.0 ± 0.3 | 1.9 ± 0.2 |
| Total Cholesterol | |||
| (mg/g feces) | 5.2 ± 2.0 | 5.4 ± 1.3 | 16.5 ± 3.0 * |
| (mg/day) | 10.2 ± 3.5 | 10.7 ± 2.3 | 30.1 ± 4.6 * |
| Triglyceride | |||
| (mg/g feces) | 1.7 ± 0.4 | 1.6 ± 0.3 | 3.4 ± 0.6 * |
| (mg/day) | 3.1 ± 0.9 | 3.2 ± 0.6 | 6.3 ± 1.4 * |
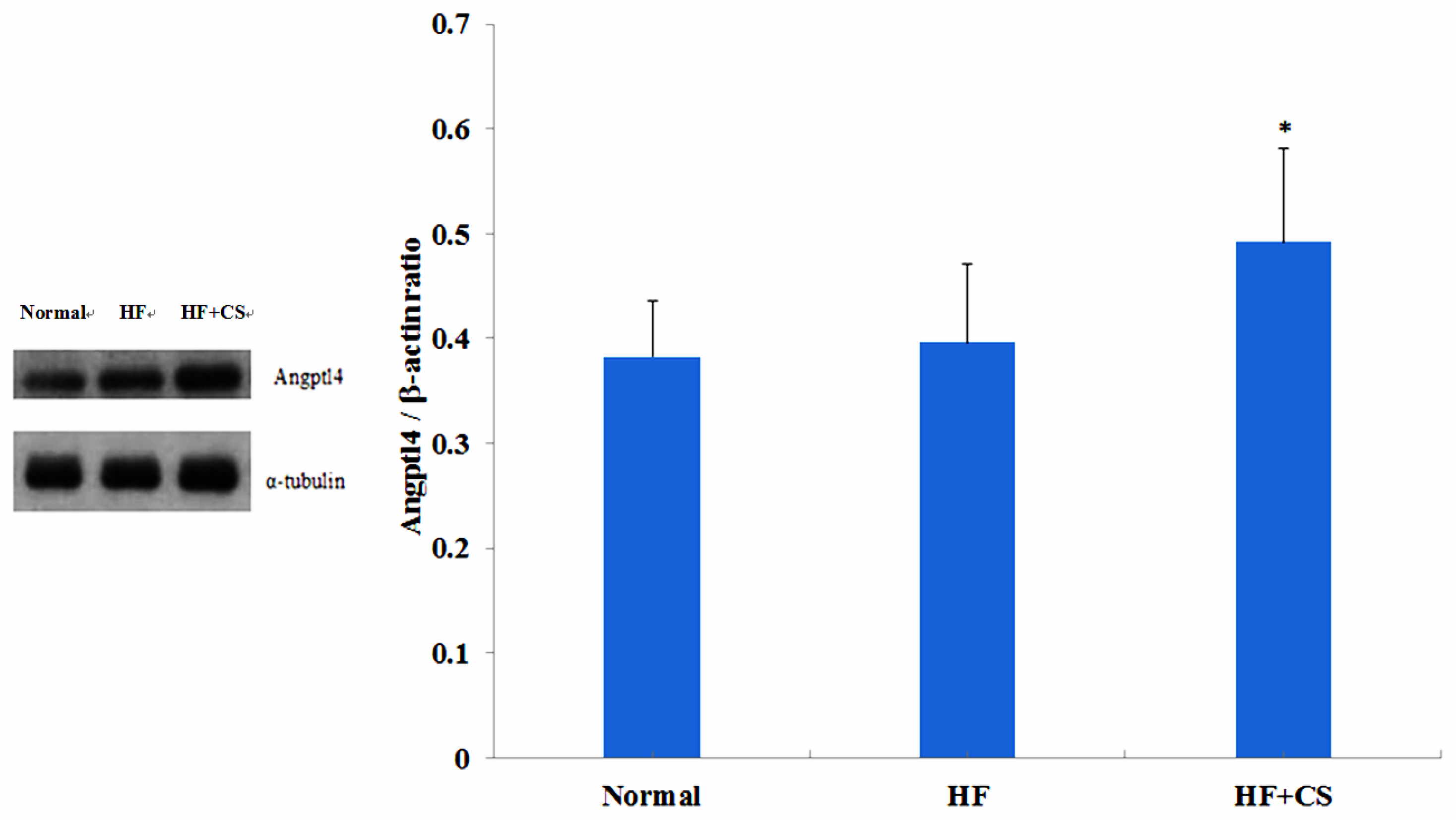
2.3. Effects of Chitosan on the ANGPTL4 Protein Expression

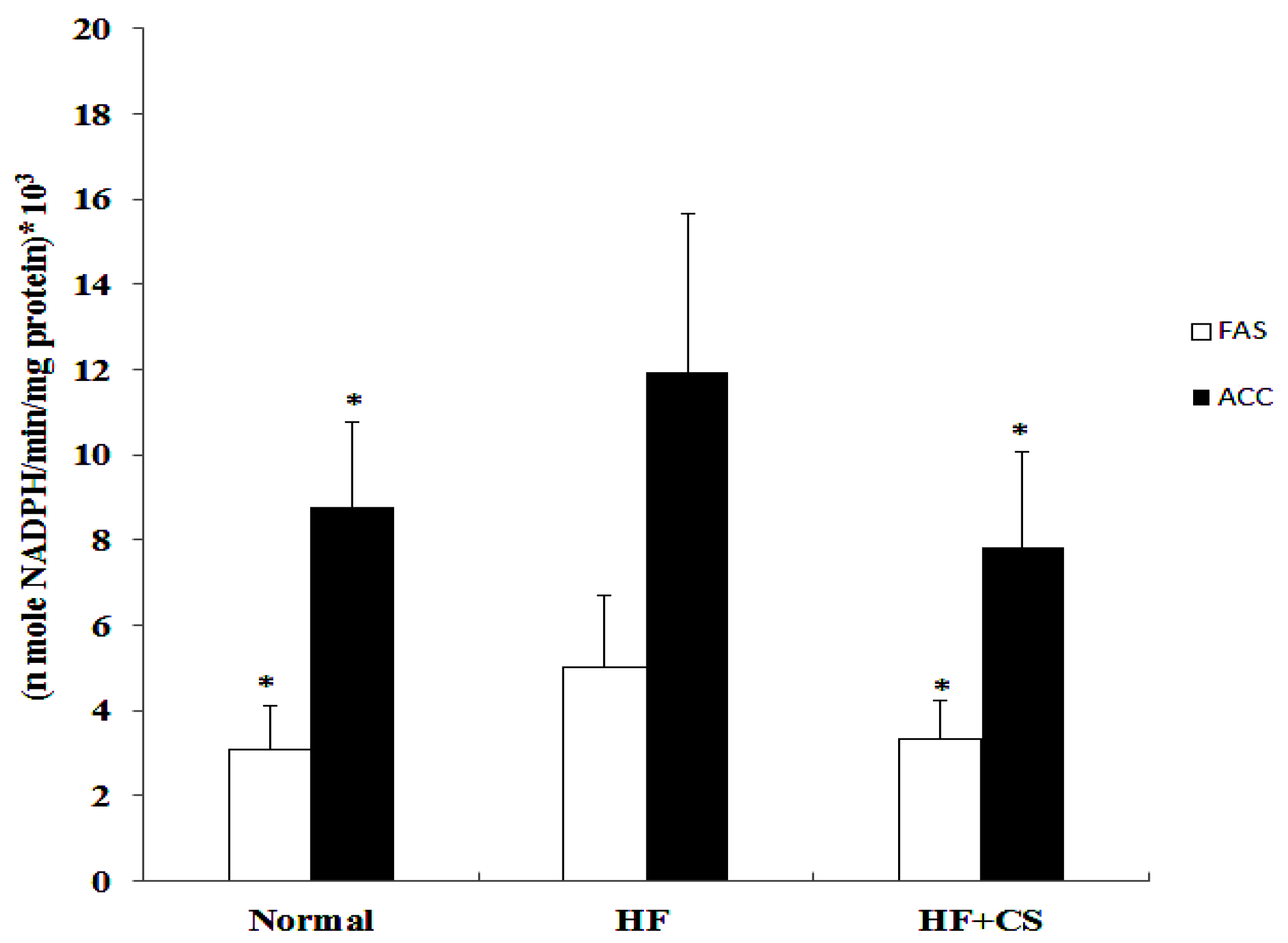
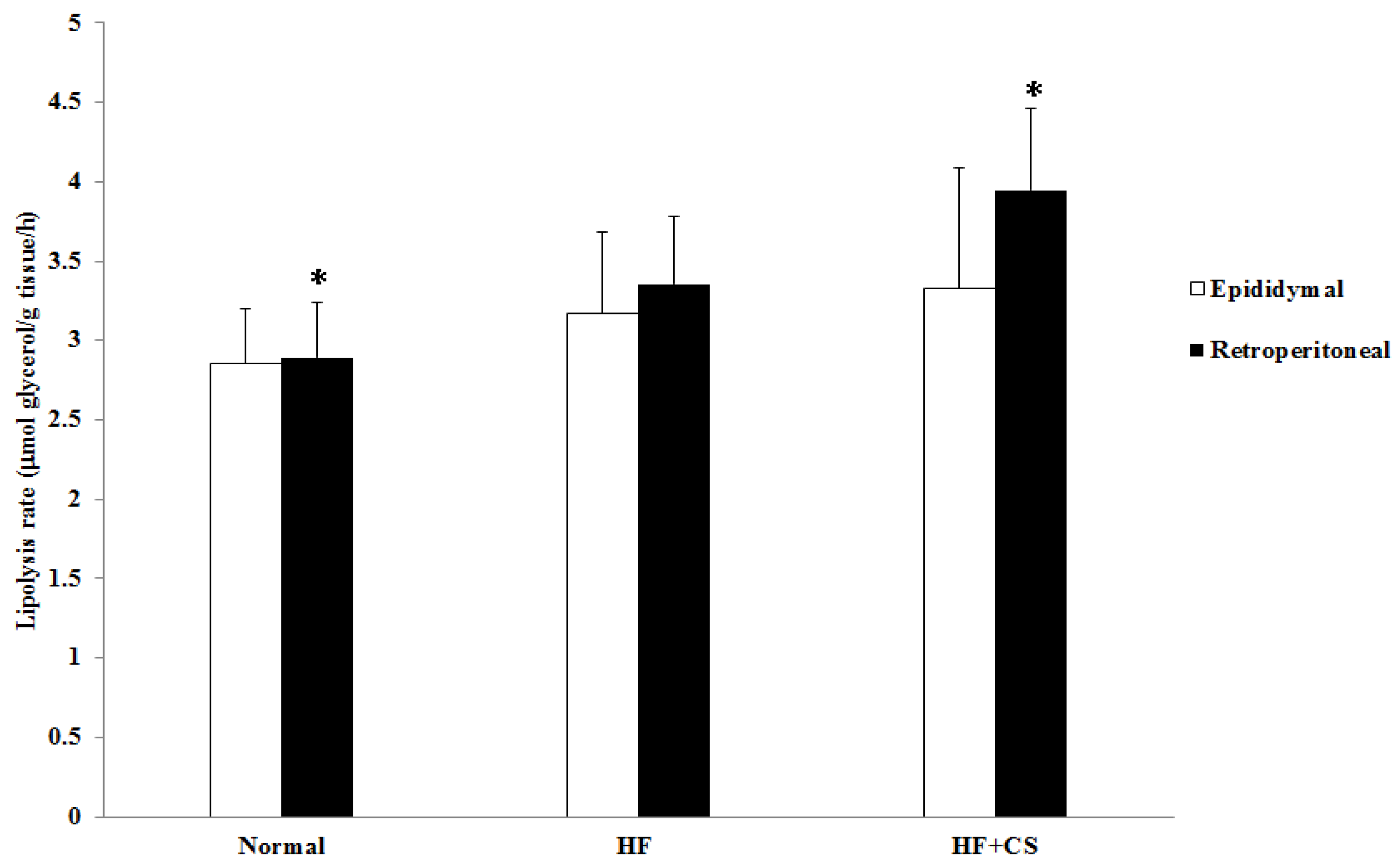
2.4. Effects of Chitosan on the Hepatic Enzyme Activities and Lipolysis

3. Discussion
4. Experimental Section
4.1. High-MW Chitosan
4.2. Animals and Diets
| Ingredient (%) | Normal * | HF | HF + C |
|---|---|---|---|
| Corn starch | 63.1 | ||
| Fructose | 63.1 | 63.1 | |
| Casein | 20 | 20 | 20 |
| Lard | 5 | 5 | 5 |
| Soybean oil | 1 | 1 | 1 |
| Vitamin mixture 1 | 1 | 1 | 1 |
| Mineral mixture 2 | 4 | 4 | 4 |
| Cholesterol | 0.5 | 0.5 | 0.5 |
| Cholic acid | 0.2 | 0.2 | 0.2 |
| Choline chloride | 0.2 | 0.2 | 0.2 |
| Cellulose | 5 | 5 | |
| Chitosan 3 | 5 |
4.3. Collection of Blood and Tissue Samples
4.4. Determination of Plasma Glucose, Insulin, Adiponectin, Leptin, Free Fatty Acids, and Tumor Necrosis Factor-α (TNF-α)
4.5. Determination of Plasma Lipid Concentration
4.6. Determination of Hepatic Lipid Metabolism Enzymes
4.7. Lipolysis Rate Measurement
4.8. Western Blot Analysis
4.9. Statistical Evaluation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bays, H.E.; Chapman, R.H.; Grandy, S. The relationship of body mass index to diabetes mellitus, hypertension and dyslipidaemia: Comparison of data from two national surveys. Int. J. Clin. Pract. 2007, 61, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Natali, A.; Ferrannini, E. Hypertension, insulin resistance, and the metabolic syndrome. Endocrinol. Metab. Clin. N. Am. 2004, 33, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.K.; Yetley, E.A. Intakes and food sources of fructose in the United States. Am. J. Clin. Nutr. 1993, 58, 737–747. [Google Scholar]
- Marriott, B.P.; Cole, N.; Lee, E. National estimates of dietary fructose intake increased from 1977 to 2004 in the United States. J. Nutr. 2009, 139, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Tappy, L.; Lê, K.A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Catena, C.; Giacchetti, G.; Novello, M.; Colussi, G.; Cavarape, A.; Sechi, L.A. Cellular mechanisms of insulin resistance in rats with fructose-induced hypertension. Am. J. Hypertens. 2003, 16, 973–978. [Google Scholar] [CrossRef]
- Bantle, J.P. Dietary fructose and metabolic syndrome and diabetes. J. Nutr. 2009, 139, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Arachchi, J.K.V.; Jeon, Y.J. Food applications of chitin and chitosan. Trends Food Sci. Technol. 1999, 10, 37–51. [Google Scholar] [CrossRef]
- Yao, H.T.; Chiang, M.T. Plasma lipoprotein cholesterol in rats fed on a diet enriched in chitosan and cholesterol. J. Nutr. Sci. Vitaminol. 2002, 48, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.T.; Chiang, M.T. Effect of chitosan on plasma lipids, hepatic lipids and fecal bile acids in hamsters. J. Food Drug Anal. 2006, 14, 183–189. [Google Scholar]
- Liu, S.H.; Chang, Y.H.; Chiang, M.T. Chitosan reduces gluconeogenesis and increases glucose uptake in skeletal muscle in streptozotocin-induced diabetic rats. J. Agric. Food Chem. 2010, 58, 5795–5800. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.L.; Yao, H.T.; Chiang, M.T. Chitosan reduces plasma adipocytokines and lipid accumulation in liver and adipose and ameliorates insulin resistance in diabetic rats. J. Med. Food 2012, 15, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Ito, M. Antidiabetic action of low molecular weight chitosan in genetically obese diabetic KK-Ay mice. Biol. Pharm. Bull. 2002, 25, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.T.; Huang, S.Y.; Chiang, M.T. A comparative study on hypoglycemic and hypocholesterolemic effects of high and low molecular weight chitosan in streptozotocin-induced diabetic rats. Food Chem. Toxicol. 2008, 46, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Mhurchu, C.N.; Poppitt, S.D.; McGill, A.T.; Leahy, F.E.; Bennet, D.A.; Lin, R.B.; Ormrod, D.; Ward, L.; Strik, C.; Rodgers, A. The effect of dietary supplement chitosan on body weight: A randomized controlled trail in 250 overweight and obese adults. Int. J. Obes. Relat. Disord. 2004, 28, 1149–1156. [Google Scholar] [CrossRef] [PubMed]
- Gallaher, D.D.; Gallaher, C.M.; Mahrt, G.J.; Carr, T.P.; Hollingshead, C.H.; Hesslink, R., Jr.; Wise, J. A glucomannan and chitosan fiber supplement decreases plasma cholesterol and increases cholesterol excretion in overweight normocholesterolemic humans. J. Am. Coll. Nutr. 2002, 21, 428–433. [Google Scholar] [CrossRef]
- Rizzo, M.; Giglio, R.V.; Nikolic, D.; Patti, A.M.; Campanella, C.; Cocchi, M.; Katsiki, N.; Montalto, G. Effects of chitosan on plasma lipids and lipoproteins: A 4-month prospective pilot study. Angiology 2014, 65, 538–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patti, A.M.; Katsiki, N.; Nikolic, D.; Al-Rasadi, K.; Rizzo, M. Nutraceuticals in lipid-lowering treatment: A narrative review on the role of chitosan. Angiology 2015, 66, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Zubiría, M.G.; Fariña, J.P.; Moreno, G.; Gagliardino, J.J.; Spinedi, E.; Giovambattista, A. Excess fructose intake-induced hypertrophic visceral adipose tissue results from unbalanced precursor cell adipogenic signals. FEBS J. 2013, 280, 5864–5874. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Chan, I.M.; Yang, T.H.; Liu, S.H.; Chiang, M.T. Supplementation of chitosan alleviates high-fat diet-enhanced lipogenesis rats via adenosine monophosphate (AMP)-activated protein kinase activation and inhibition of lipogensis-associated genes. J. Agric. Food Chem. 2015, 63, 2979–2988. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; He, S.P.; Chiang, M.T. Effects of long-term feeding of chitosan on postprandial lipid responses and lipid metabolism in a high-sucrose-diet-impaired-glucose-tolerant rat model. J. Agric. Food Chem. 2012, 60, 4306–4313. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, Y. The metabolic syndrome and adipocytokines. FEBS Lett. 2006, 580, 2917–2921. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Okuda, H.; Morimoto, C.; Tsujita, T. Effect of substracts on cyclic AMP-dependent lipolytic reaction of hormone-sensitive lipase. J. Lipid Res. 1994, 35, 1267–1273. [Google Scholar] [PubMed]
- Samuel, V.T. Fructose induced lipogenesis: From sugar to fat to insulin resistance. Trends Endocrinol. Metab. 2011, 22, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Costabile, G.; Annuzzi, G.; di Marino, L.; de Natale, C.; Giacco, R.; Bozztto, L.; Cipriano, P.; Santangelo, C.; Masella, R.; Rivellese, A.A. Fasting and post-prandial adipose tissue lipoprotein lipase and hormone-sensitive lipase in obesity and type 2 diabetes. J. Endocrinol. Investig. 2014, 34, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Gotto, A.M. Lipid management in diabetic patients: Lessons from prevention trials. Am. J. Med. 2002, 112, 19–26. [Google Scholar] [CrossRef]
- Xu, G.; Huang, X.; Qiu, L.; Wu, J.; Hu, Y. Mechanism study of chitosan on lipid metabolism in hyperlipidemic rats. Asia Pac. J. Clin. Nutr. 2007, 16, 313–317. [Google Scholar] [PubMed]
- Lichtenstein, L.; Berbee, J.F.; van Dijk, K.W.; van Dijk, S.J.; Bensadoun, A.; Kema, I.P.; Voshol, P.J.; Muller, M.; Resen, P.C.; Kersten, S. Angptl4 upregulates cholesterol synthesis in liver via inhibition of LPL- and HL-dependent hepatic cholesterol uptake. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2420–2427. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, L.; Kersten, S. Modulation of plasma TG lipolysis by angiopoietin-like protein and GPIHBP1. Biochim. Biophys. Acta 2010, 1801, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandehroo, M.; Knoch, B.; Muller, M.; Kersten, S. Peroxisome proliferator-activated receptor α target genes. PPAR Res. 2010, 61, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Staiger, H.; Haas, C.; Machann, J.; Werner, R.; Weisser, M.; Schick, F.; Machicao, F.; Stefan, N.; Fritsche, A.; Haring, H.U. Muscle-dervied angiopoietin-like protein 4 is induced by fatty acids via PPARδ and is of metabolic relevance in human. Diabetes 2008, 58, 578–589. [Google Scholar]
- Karadeniz, F.; Kim, S.K. Antidiabetic activities of chitosan and its derivatives: A mini review. In Marine Carbohydrates: Fundamentals and Applications, Part B; Kim, S., Ed.; Elsevier Inc.: Oxford, UK, 2014; pp. 15–31. [Google Scholar]
- Kerch, G. The potential of chitosan and its derivatives in prevention and treatment of age-related diseases. Mar. Drugs 2015, 13, 2158–2182. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Osaki, T.; Minami, S.; Okamoto, Y. Anticancer and anti-inflammatory properties of chitin and chitosan oligosaccharides. J. Funct. Biomater. 2015, 6, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Anraku, M.; Michihara, A.; Yasufuku, T.; Akasaki, K.; Tsuchiya, D.; Nishio, H.; Maruyama, T.; Otagiri, M.; Maezaki, Y.; Kondo, Y.; et al. The antioxidative and antilipidemic effects of different molecular weight chitosans in metabolic syndrome model rats. Biol. Pharm. Bull. 2010, 33, 1994–1998. [Google Scholar] [CrossRef] [PubMed]
- Takehisa, F.; Suzuki, Y. Effect of guar gum and cholestyramine on plasma lipoprotein cholesterol in rats. J. Jpn. Soc. Nutr. Food Sci. 1990, 43, 269–274. [Google Scholar] [CrossRef]
- Goodridge, A.G. Regulation of the activity of acetyl coenzyme A carboxylase by palmitoyl coenzyme A and citrate. J. Biol. Chem. 1972, 247, 6946–6952. [Google Scholar] [PubMed]
- Nepokroeff, C.M.; Lakshmanan, M.R.; Porter, J.W. Fatty-acid synthetase from rat liver. Methods Enzymol. 1975, 35, 37–44. [Google Scholar] [PubMed]
- Berger, J.J.; Barnard, R.J. Effect of diet on fat cell size and hormone-sensitive lipase activity. J. Appl. Physiol. 1999, 87, 227–232. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.-H.; Cai, F.-Y.; Chiang, M.-T. Long-Term Feeding of Chitosan Ameliorates Glucose and Lipid Metabolism in a High-Fructose-Diet-Impaired Rat Model of Glucose Tolerance. Mar. Drugs 2015, 13, 7302-7313. https://doi.org/10.3390/md13127067
Liu S-H, Cai F-Y, Chiang M-T. Long-Term Feeding of Chitosan Ameliorates Glucose and Lipid Metabolism in a High-Fructose-Diet-Impaired Rat Model of Glucose Tolerance. Marine Drugs. 2015; 13(12):7302-7313. https://doi.org/10.3390/md13127067
Chicago/Turabian StyleLiu, Shing-Hwa, Fang-Ying Cai, and Meng-Tsan Chiang. 2015. "Long-Term Feeding of Chitosan Ameliorates Glucose and Lipid Metabolism in a High-Fructose-Diet-Impaired Rat Model of Glucose Tolerance" Marine Drugs 13, no. 12: 7302-7313. https://doi.org/10.3390/md13127067
APA StyleLiu, S. -H., Cai, F. -Y., & Chiang, M. -T. (2015). Long-Term Feeding of Chitosan Ameliorates Glucose and Lipid Metabolism in a High-Fructose-Diet-Impaired Rat Model of Glucose Tolerance. Marine Drugs, 13(12), 7302-7313. https://doi.org/10.3390/md13127067