
Persistent Contamination of Octopuses and Mussels with Lipophilic Shellfish Toxins during Spring Dinophysis Blooms in a Subtropical Estuary

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
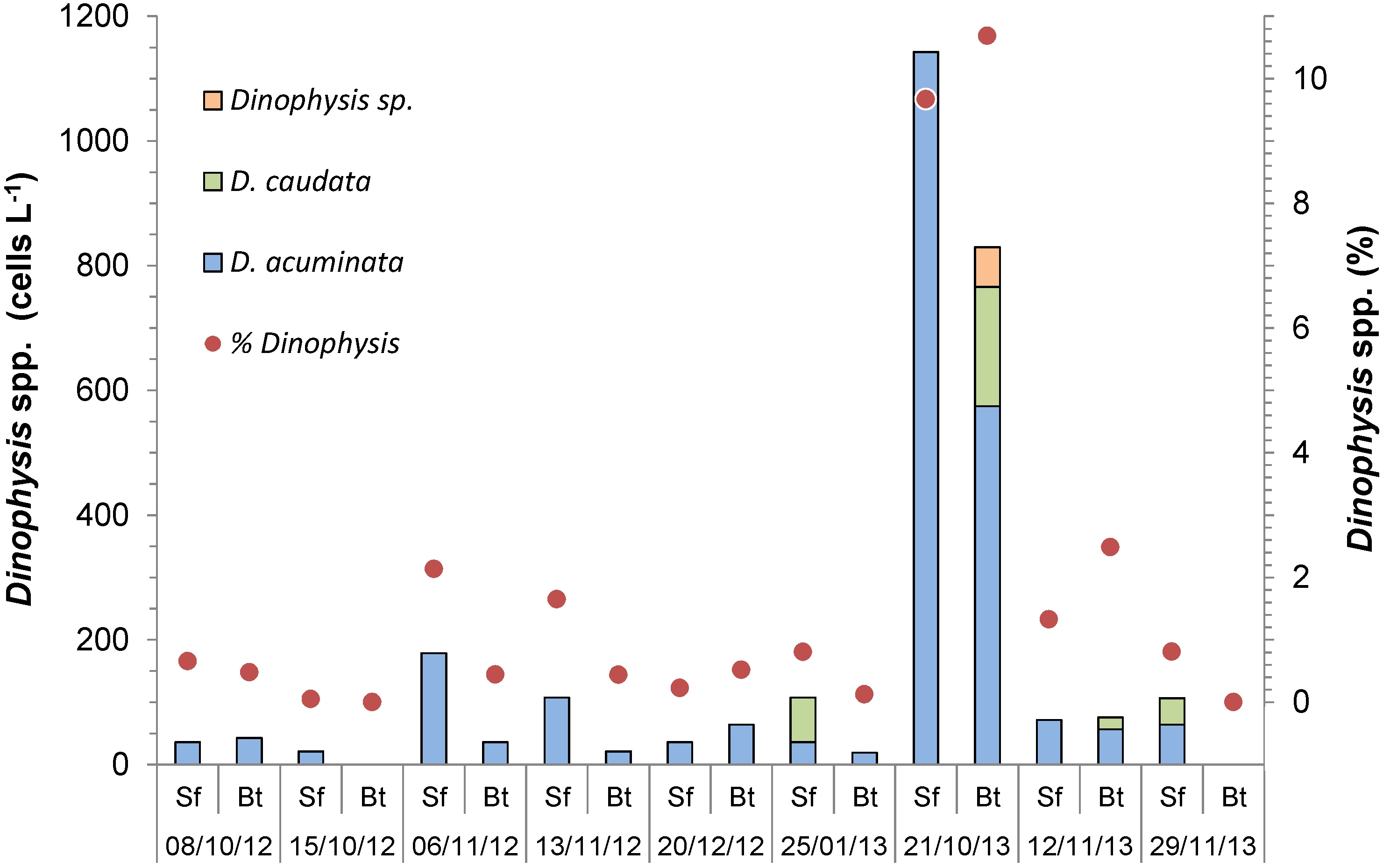
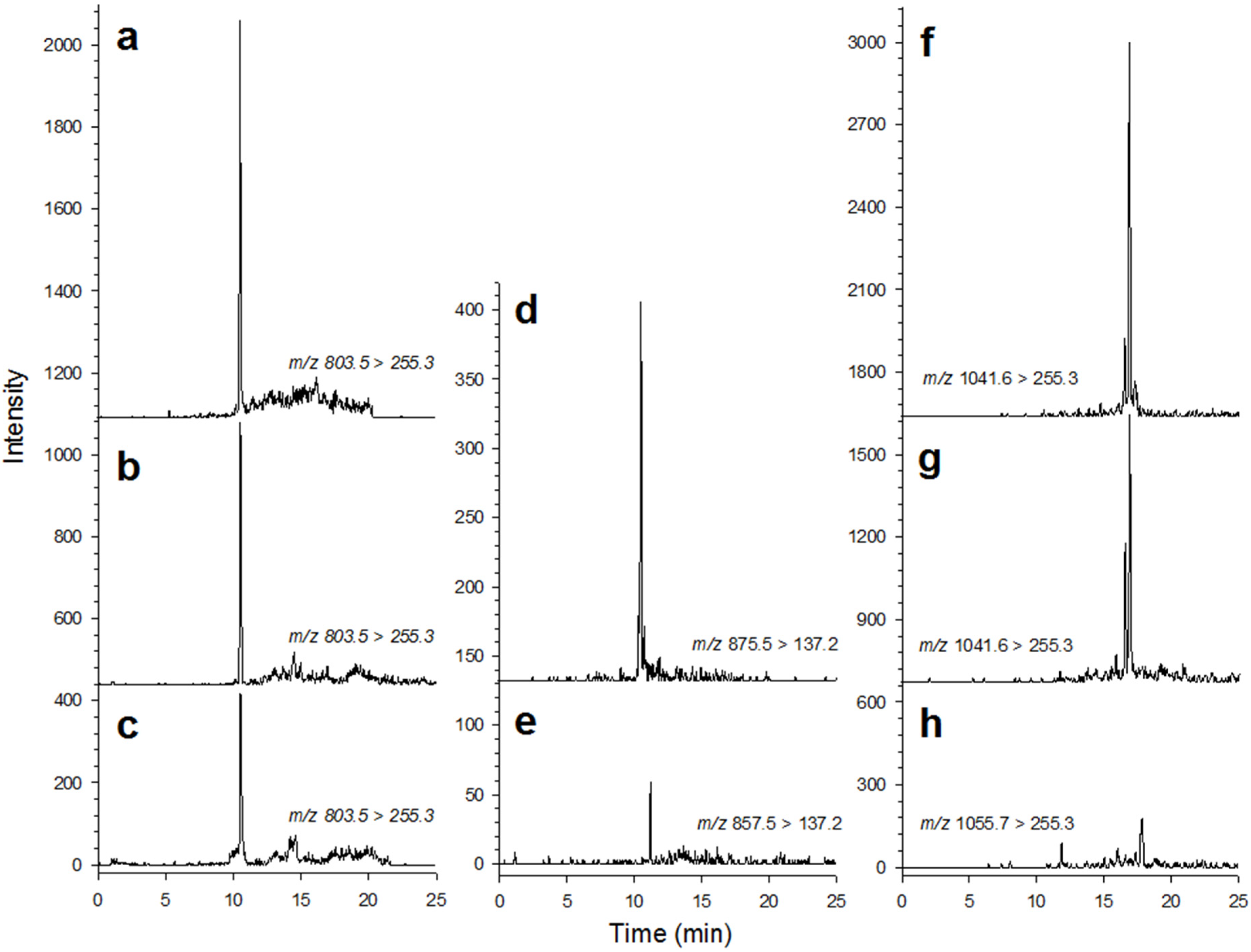
2.1. Cell Abundance and Toxin Concentration in the Plankton



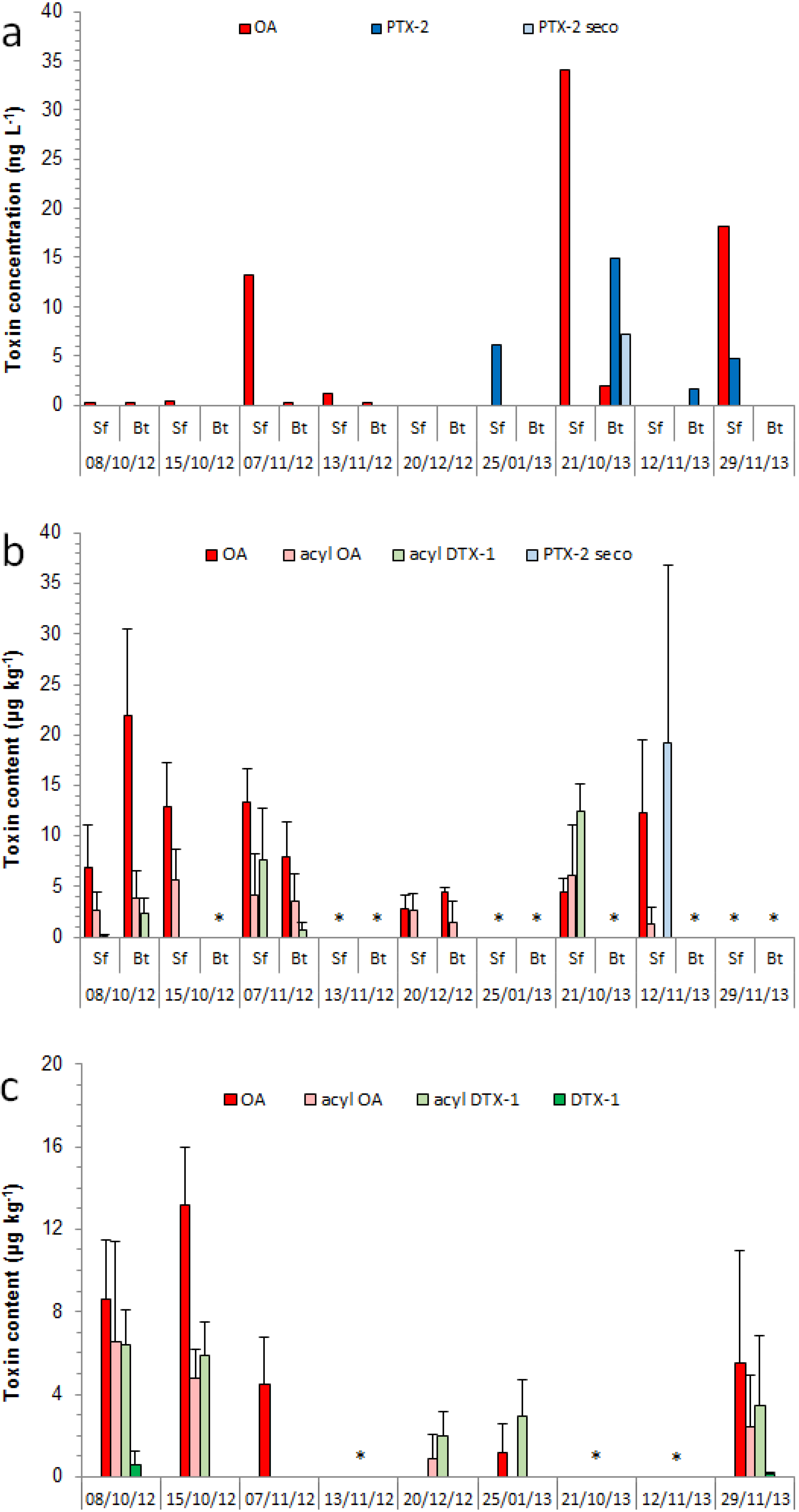
2.2. Toxin in Mussels and Octopuses

{kind=link}
{kind=link}
{kind=link}
| Tissue | OA | acyl OA | acyl DTX-1 | PTX-2 seco | |||||
|---|---|---|---|---|---|---|---|---|---|
| µg kg−1 | n | µg kg−1 | n | µg kg−1 | n | µg kg−1 | n | ||
| Hepatopancreas | avg. | 22.2 ± 17.9 | (25/26) | 8.87 ± 10.3 | (17/26) | 7.81 ± 15.2 | (10/26) | 6.34 ± 27.2 | (2/26) |
| max. | 69.7 | 43.2 | 53.3 | 137.0 | |||||
| Non-visceral tissues | avg. | 0.94 ± 1.30 | (11/26) | nd | nd | nd | |||
| max. | 4.04 | ||||||||
| OA | acyl OA | acyl DTX-1 | DTX-1 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Tissue | ng g−1 | n | ng g−1 | n | ng g−1 | n | ng g−1 | n | |
| Digestive gland | avg. | 5.49 ± 5.66 | (4/5) | 2.44 ± 3.94 | (3/5) | 3.41 ± 3.02 | (4/5) | 0.11 ± 0.44 | (1/5) |
| max. | 17.3 | 14.4 | 8.8 | 1.7 | |||||
| Stomach + intestine | avg. | 0.43 ± 1.70 | (1/5) | 0.06 ± 0.23 | (1/5) | 0.04 ± 0.16 | (1/5) | nd | |
| max. | 6.6 | 0.9 | 0.6 | ||||||
| Gills | avg. | 2.48 ± 3.94 | (3/5) | 0.02 ± 0.09 | (1/5) | nd | nd | ||
| max. | 13.2 | 0.4 | |||||||
| Kidneys | avg. | 1.12 ± 2.48 | (2/5) | 0.10 ± 0.33 | (1/5) | nd | nd | ||
| max. | 7.3 | 1.0 | |||||||
| Arm | avg. | 3.72 ± 11.3 | (2/5) | nd | nd | nd | |||
| max. | 43.7 | ||||||||
| Gonads | avg. | nd | nd | nd | nd | ||||
| max. | |||||||||
| Mantle | avg. | nd | nd | nd | nd | ||||
| max. | |||||||||
3. Experimental Section
3.1. Field Sampling
3.2. Toxin Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Reguera, B.; Riobó, P.; Rodríguez, F.; Díaz, P.A.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis Toxins: Causative Organisms, Distribution and Fate in Shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Holmes, C.F.B.; Tsukitani, Y. Okadaic acid, a new probe for studying cellular regulation. Trends Biochem. Sci. 1990, 15, 98–102. [Google Scholar] [CrossRef]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku district. J. Nippon Suisan Gakkaishi 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Identification of Dinophysis fortii as the causative organism of diarrhetic shellfish poisoning. Bull. Jpn. Soc. Sci. Fish. 1980, 46, 1405–1411. [Google Scholar] [CrossRef]
- Valdiglesias, V.; Prego-Faraldo, M.V.; Pásaro, E.; Méndez, J.; Laffon, B. Okadaic Acid: More than a Diarrheic Toxin. Mar. Drugs 2013, 11, 4328–4349. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M. Unique features of the okadaic acid activity class of tumor promoters. J. Cancer Res. Clin. Oncol. 1999, 125, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Moestrup, Ø.; Akselman, R.; Fraga, S.; Hansen, G.; Hoppenrath, M.; Iwataki, M.; Komárek, J.; Larsen, J.; Lundholm, N.; Zingone, A. (Eds.) IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. Available online: http://www.marinespecies.org/HAB (accessed on 20 March 2015).
- Miles, C.O.; Wilkins, A.L.; Munday, R.; Dines, M.H.; Hawkes, A.D.; Briggs, L.R.; Sandvik, M.; Jensen, D.J.; Cooney, J.M.; Holland, P.T.; et al. Isolation of pectenotoxin-2 from Dinophysis acuta and its conversion to pectenotoxin-2 seco acid, and preliminary assessment of their acute toxicities. Toxicon 2004, 43, 1–9. [Google Scholar] [PubMed]
- Bricelj, V.M.; Shumway, S.E. Paralytic shellfish toxins in bivalve molluscs: Occurrence, transfer kinetics and biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Reizopoulou, S.; Strogyloudi, E.; Giannakourou, A.; Pagou, K.; Hatzianestis, I.; Pyrgaki, C.; Granéli, E. Okadaic acid accumulation in macrofilter feeders subjected to natural blooms of Dinophysis acuminata. Harmful Algae 2008, 7, 228–234. [Google Scholar] [CrossRef]
- Vale, P.; Sampayo, M.A.M. First confirmation of human diarrhoeic poisonings by okadaic acid esters after ingestion of razor clams (Solen marginatus) and green crabs (Carcinus maenas) in Aveiro lagoon, Portugal and detection of okadaic acid esters in phytoplankton. Toxicon 2002, 40, 989–996. [Google Scholar] [CrossRef]
- Sipiä, V.; Kankaanpää, H.; Meriluoto, J. The first observation of okadaic acid in flounder in the Baltic Sea. Sarsia 2000, 85, 471–475. [Google Scholar]
- Mafra, L.L., Jr.; Tavares, C.P.S.; Schramm, M.A. Diarrheic toxins in field-sampled and cultivated Dinophysis spp. cells from southern Brazil. J Appl .Phycol. 2014, 26, 1727–1739. [Google Scholar] [CrossRef]
- Costa, P.R.; Rosa, R.; Sampayo, M.A.M. Tissue distribution of the amnesic shellfish toxin, domoic acid, in Octopus vulgaris from the Portuguese coast. Mar. Biol. 2004, 144, 971–976. [Google Scholar] [CrossRef]
- Costa, P.R.; Rosa, R.; Pereira, J.; Sampayo, M.A.M. Detection of domoic acid, the amnesic shellfish toxin, in the digestive gland of Eledone cirrhosa and E. moschata (Cephalopoda, Octopoda) from the Portuguese coast. Aquat. Living Resour. 2005, 18, 395–400. [Google Scholar] [CrossRef]
- Robertson, A.; Stirling, D.; Robillot, C.; Llewellyn, L.; Negri, A. First report of saxitoxin in octopi. Toxicon 2004, 44, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.R.; Botelho, M.J.; Rodrigues, S.M. Accumulation of paralytic shellfish toxins in digestive gland of Octopus vulgaris during bloom events including the dinoflagellate Gymnodinium catenatum. Mar. Pollut. Bull. 2009, 58, 1739–1765. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, A.; Costa, P.R. Distribution and selective elimination of paralytic shellfish toxins in different tissues of Octopus vulgaris. Harmful Algae 2011, 10, 732–737. [Google Scholar] [CrossRef]
- Lopes, V.M.; Baptista, M.; Repolho, T.; Rosa, R.; Costa, P.R. Uptake, transfer and elimination kinetics of paralytic shellfish toxins in common octopus (Octopus vulgaris). Aquat. Toxicol. 2014, 146, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Hackett, J.D.; Tong, M.; Kulis, D.M.; Fux, E.; Hess, P.; Bire, R.; Anderson, D.M. DSP toxin production de novo in cultures of Dinophysis acuminata (Dinophyceae) from North America. Harmful Algae 2009, 8, 873–879. [Google Scholar] [CrossRef]
- Fux, E.; Smith, J.L.; Tong, M.; Guzman, L.; Anderson, D.M. Toxin profiles of five geographical isolates of Dinophysis spp. from North and South America. Toxicon 2012, 57, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Kulis, D.M.; Fux, E.; Smith, J.L.; Hess, P.; Zhou, Q.X.; Anderson, D.M. The effects of growth phase and light intensity on toxin production by Dinophysis acuminata from the northeastern United States. Harmful Algae 2011, 10, 254–264. [Google Scholar] [CrossRef]
- Tong, M.; Smith, J.L.; Richlen, M.; Steidinger, K.A.; Kulis, D.M.; Fux, E.; Anderson, D.M. Characterization and comparison of toxin-producing isolates of Dinophysis acuminata from New England and Canada. J. Phycol. 2015, 51, 66–81. [Google Scholar] [CrossRef]
- Kamiyama, T.; Suzuki, T. Production of dinophysistoxin-1 and pectenotoxin-2 by a culture of Dinophysis acuminata (Dinophyceae). Harmful Algae 2009, 8, 312–317. [Google Scholar] [CrossRef]
- Kamiyama, T.; Nagai, S.; Suzuki, T.; Miyamura, K. Effect of temperature on production of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 by Dinophysis acuminata in culture experiments. Aquat. Microb. Ecol. 2010, 60, 193–202. [Google Scholar] [CrossRef]
- Nagai, S.; Suzuki, T.; Nishikawa, T.; Kamiyama, T. Differences in the production and excretion kinetics of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 between cultures of Dinophysis acuminata and Dinophysis fortii isolated from western Japan. J. Phycol. 2011, 47, 1326–1337. [Google Scholar] [CrossRef]
- Report of the Joint FAO/IOC/WHO ad hoc Expert Consultation on Biotoxins in Bivalve Molluscs. Available online: http://unesdoc.unesco.org/images/0013/001394/139421e.pdf (accessed on 8 April 2015).
- Proença, L.A.O.; Schramm, M.A.; Tamanaha, M.S.; Alves, T.P. Diarrhoetic shellfish poisoning (DSP) outbreak in subtropical southwest Atlantic. Harmful Algae News 2007, 33, 19–20. [Google Scholar]
- Mafra, L.L., Jr.; Ribas, T.; Alves, T.P.; Proença, L.A.O.; Schramm, M.A.; Uchida, H.; Suzuki, T. Differential okadaic acid accumulation and detoxification by oysters and mussels during natural and simulated Dinophysis blooms. Fish. Sci. 2015. [Google Scholar] [CrossRef]
- Suzuki, T.; Mackenzie, L.; Stirling, D.; Adamson, J. Pectenotoxin-2 seco acid: A toxin converted from pectenotoxin-2 by the New Zealand Greenshell mussel, Perna canaliculus. Toxicon 2001, 39, 507–514. [Google Scholar] [CrossRef]
- Nishitani, G.; Nagai, S.; Sakiyama, S.; Kamiyama, T. Successful cultivation of the toxic dinoflagellate Dinophysis caudata (Dinophyceae). Plankton Benthos Res. 2008, 3, 78–85. [Google Scholar] [CrossRef]
- Marasigan, A.N.; Sato, S.; Fukuyo, Y.; Kodama, M. Accumulation of a high level of diarrhetic shellfish toxins in the green mussel Perna viridis during a bloom of Dinophysis caudata and Dinophysis miles in Saipan Bay, Panay Island, the Philippines. Fish. Sci. 2001, 67, 994–996. [Google Scholar] [CrossRef]
- Mafra, L.L., Jr.; (Federal University of Paraná State, Curitiba, Brazil); Nagai, S.; Uchida, H.; Suzuki, T.; (National Research Institute of Fisheries Science, Yokohama, Japan). Unpublished work. 2014.
- Kim, J.H.; Lee, K.J.; Suzuki, T.; Kang, Y.S.; Kim, P.H.; Song, K.C.; Lee, T.S. Seasonal variability of lipophilic shellfish toxins in bivalves and waters, and abundance of Dinophysis spp. in Jinhae Bay, Korea. J. Shellfish Res. 2010, 29, 1061–1067. [Google Scholar] [CrossRef]
- Hatfield, C.L.; Gauglitz, E.J., Jr.; Barnett, H.J.; Lund, J.A.K.; Wekell, J.C.; Eklund, M. The fate of domoic acid in Dungeness crab (Cancer magister) as a function of processing. J. Shellfish Res. 1995, 14, 359–363. [Google Scholar]
- Choi, M.C.; Yu, P.K.N.; Hsieh, D.P.H.; Lam, P.K.S. Trophic transfer of paralytic shellfish toxins from clams (Ruditapes philippinarum) to gastropods (Nassarius festivus). Chemosphere 2006, 64, 1642–1649. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.A. Diet of dolphins, Delphinus delphis, off the Portuguese continental coast. J. Mar. Biol. Assoc. UK 1999, 79, 531–540. [Google Scholar] [CrossRef]
- Blanco, C.; Salomón, O.; Raga, J.A. Diet of the bottlenose dolphin (Tursiops truncatus) in the western Mediterranean Sea. J. Mar. Biol. Assoc. UK 2001, 81, 1053–1058. [Google Scholar] [CrossRef]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M.G. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Mangold, K. Eledone Moschata. In Cephalopod Life Cycles: Species Accounts; Boyle, P.R., Ed.; Academic Press: London, UK, 1983; Volume 1, pp. 387–400. [Google Scholar]
- Miramand, P.; Bentley, D. Concentration and distribution of heavy metals in tissues of two cephalopods, Eledone cirrhosa and Sepia officinalis, from the French coast of the English Channel. Mar. Biol. 1992, 114, 407–414. [Google Scholar] [CrossRef]
- Raimundo, J.; Costa, P.; Vale, C.; Costa, M.H.; Moura, I. DNA damage and metal accumulation in four tissues of feral Octopus vulgaris from two coastal areas in Portugal. Ecotoxicol. Environ. Saf. 2010, 73, 1543–1547. [Google Scholar] [CrossRef] [PubMed]
- Lopes, V.M.; Lopes, A.R.; Costa, P.; Rosa, R. Cephalopods as vectors of harmful algal bloom toxins in marine food webs. Mar. Drugs 2013, 11, 3381–3409. [Google Scholar] [CrossRef] [PubMed]
- Rosa, R.; Marques, A.M.; Nunes, M.L.; Bandarra, N.; Sousa Reis, C. Spatial–temporal changes in dimethyl acetal (octadecanal) levels of Octopus vulgaris (Mollusca, Cephalopoda): Relation to feeding ecology. Sci. Mar. 2004, 68, 227–236. [Google Scholar] [CrossRef]
- Kvitek, R.G.; Beitler, M.K. Relative insensitivity of butter clam neurons to saxitoxin: A pre-adaptation for sequestering paralytic shellfish poisoning toxins as a chemical defense. Mar. Ecol. Prog. Ser. 1991, 69, 47–54. [Google Scholar] [CrossRef]
- Yamassaki, G.E. Dinâmica populacional de Octopus vulgaris (Cuvier, 1797) no Complexo Estuarino de Paranaguá, Paraná, Brasil. B.Sc. Thesis, Universidade Federal do Paraná, Paraná, Brazil, 2011. [Google Scholar]
- Balech, E. Los dinoflagelados del Atlántico Sudoccidental. (Publ. Espec. st. Esp. Oceanogr.); Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 1988; p. 310. [Google Scholar]
- Steidiner, K.A.; Tangen, K. Dinoflagellates. In Identifying Marine Phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 387–584. [Google Scholar]
- Haraguchi, L.; Odebrecht, C. Dinophysiales (Dinophyceae) no extremo Sul do Brasil (inverno de 2005, verão de 2007). Biota Neotrop. 2010, 10, 101–114. [Google Scholar] [CrossRef]
- Suzuki, T.; Jin, T.; Shirota, T.; Mitsuya, T.; Okumura, Y.; Kamiyama, T. Quantification of lipophilic toxins associated with diarrhetic shellfish poisoning in Japanese bivalves by liquid chromatography-mass spectrometry and comparison with mouse bioassay. Fish. Sci. 2005, 71, 1370–1378. [Google Scholar] [CrossRef]
- Suzuki, T.; Miyazono, A.; Baba, K.; Sugawara, R.; Kamiyama, T. LC-MS/MS analysis of okadaic acid analogues and other lipophilic toxins in single-cell isolates of several Dinophysis species collected in Hokkaido, Japan. Harmful Algae 2009, 8, 233–238. [Google Scholar] [CrossRef]
- Quilliam, M.A.; Hess, P.; Dell’Aversano, C. Recent developments in the analysis of phycotoxins by liquid chromatography-mass spectrometry. In Mycotoxins and Phycotoxins in Perspective at the Turn of the Century, Proceedings of the X International IUPAC Symposium, Guaruja, Brazil, 21–25 May 2000; de Koe, W.J., Samson, R.A., van Egmond, H.P., Gilbert, J., Sabino, M., Eds.; pp. 383–391.
- Quilliam, M.A. Chemical Method for Lipophilic Shellfish Toxins. In Manual on Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., Eds.; UNESCO Publishing: Paris, France, 2003; pp. 211–245. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mafra, L.L., Jr.; Lopes, D.; Bonilauri, V.C.; Uchida, H.; Suzuki, T. Persistent Contamination of Octopuses and Mussels with Lipophilic Shellfish Toxins during Spring Dinophysis Blooms in a Subtropical Estuary. Mar. Drugs 2015, 13, 3920-3935. https://doi.org/10.3390/md13063920
Mafra LL Jr., Lopes D, Bonilauri VC, Uchida H, Suzuki T. Persistent Contamination of Octopuses and Mussels with Lipophilic Shellfish Toxins during Spring Dinophysis Blooms in a Subtropical Estuary. Marine Drugs. 2015; 13(6):3920-3935. https://doi.org/10.3390/md13063920
Chicago/Turabian StyleMafra, Luiz L., Jr., Daiana Lopes, Vanessa C. Bonilauri, Hajime Uchida, and Toshiyuki Suzuki. 2015. "Persistent Contamination of Octopuses and Mussels with Lipophilic Shellfish Toxins during Spring Dinophysis Blooms in a Subtropical Estuary" Marine Drugs 13, no. 6: 3920-3935. https://doi.org/10.3390/md13063920