Marine-Derived Penicillium Species as Producers of Cytotoxic Metabolites


Abstract
:1. Introduction
2. Alkaloids
3. Terpenes, Meroterpenes, and Steroids
4. Polyketides
5. Lipopeptides
6. Miscellaneous Compounds
7. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
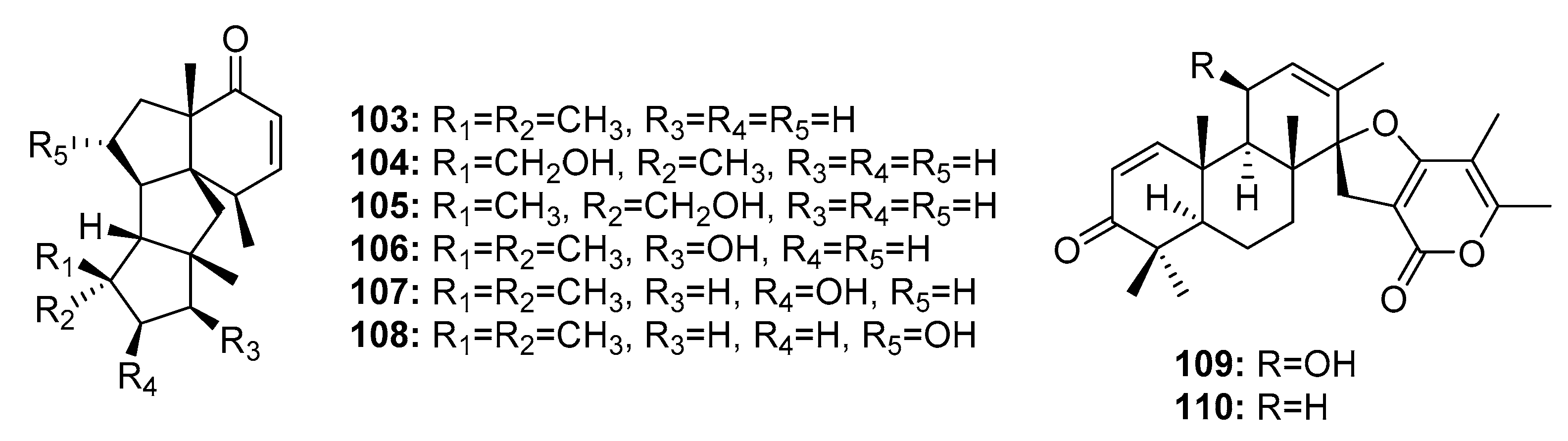{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Producing Stain | Environment Source | Type | Cell Lines/Brine Shrimp | IC50, LD50, or IR (%) | Target | Reference |
|---|---|---|---|---|---|---|---|
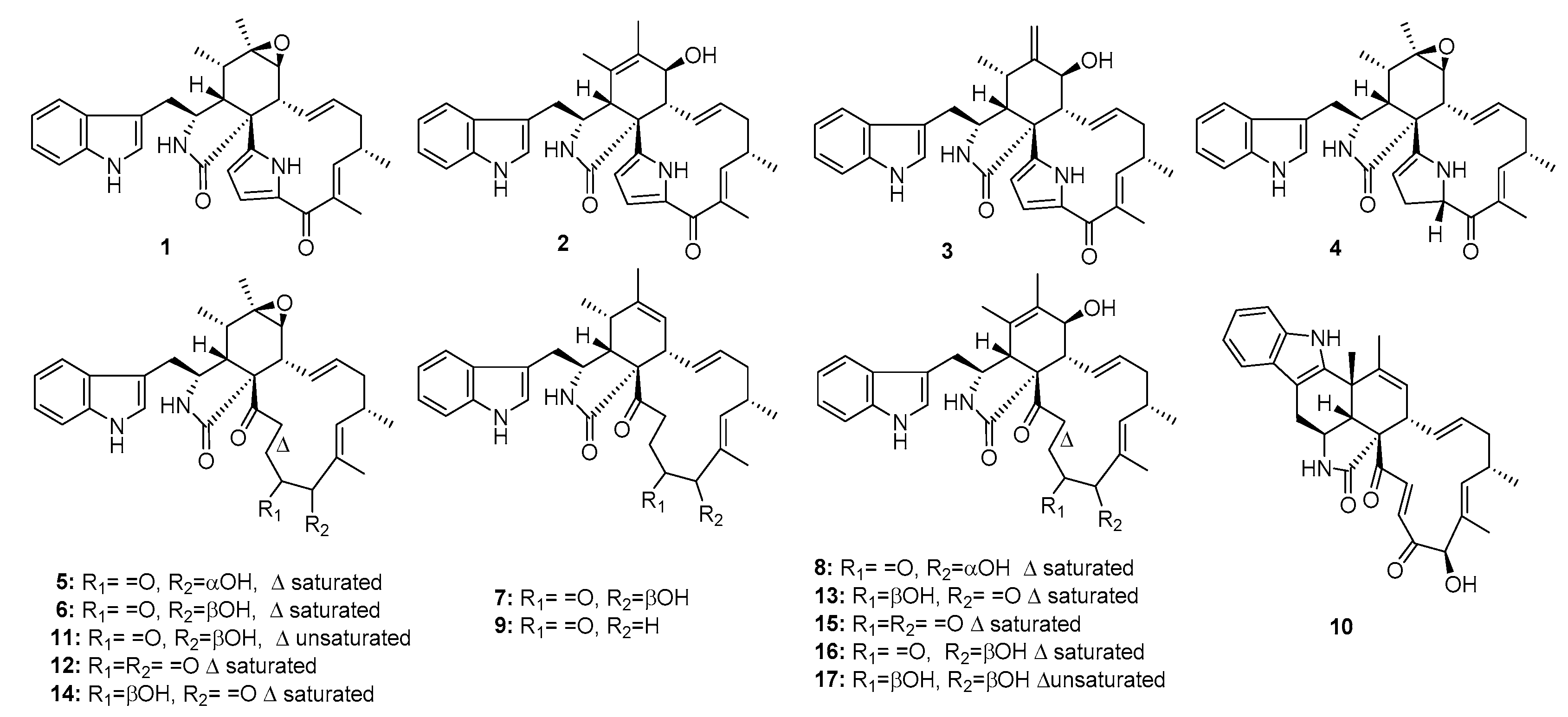
| Penochalasin A (1) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 0.4 μg/mL | [8] | |
| Penochalasin B (2) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 0.3 μg/mL | [8] | |
| Penochalasin C (3) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 0.5 μg/mL | [8] | |
| Penochalasin D (4) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 3.2 μg/mL | [7] | |
| Penochalasin E (5) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 2.1 μg/mL | [7] | |
| Penochalasin F (6) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 1.8 μg/mL | [7] | |
| Penochalasin G (7) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 1.9 μg/mL | [7] | |
| Penochalasin H (8) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 2.8 μg/mL | [7] | |
| Penochalasin I (9) | P. chrysogenum V11 | Mangrove | Indole alkaloid | MDA-MB-435, SGC-7901, A549 | (7.55, 7.32, 16.13) μM | [6] | |
| Penochalasin J (10) | P. chrysogenum V11 | Mangrove | Indole alkaloid | MDA-MB-435, SGC-7901, A549 | (36.68, 37.70, 35.93) μM | [6] | |
| Chaetoglobosin A (11) | P. chrysogenum V11 | Mangrove | Indole alkaloid | P388, MDA-MB-435, SGC-7901, A549 | 0.6 μg/mL (37.56, 7.84, 6.56) μM | Cell-cycle arrest induction, membrane ruffling inhibition, and cell migration | [6,8,9] |
| Chaetoglobosin C (12) | P. chrysogenum V11 | Mangrove | Indole alkaloid | MDA-MB-435, SGC-7901, A549 | (19.97, 15.36, 17.82) μM | [6] | |
| Chaetoglobosin E (13) | P. chrysogenum V11 | Mangrove | Indole alkaloid | A549 | 36.63 μM | [6] | |
| Chaetoglobosin F (14) | P. chrysogenum V11 | Mangrove | Indole alkaloid | P388, MDA-MB-435, SGC-7901, A549 | 0.9 μg/mL, (37.77, 26.53, 27.72) μM | [6,8] | |
| Chaetoglobosin G (15) | P. chrysogenum V11 | Mangrove | Indole alkaloid | MDA-MB-435, SGC-7901, A549 | (38.77, 25.86, 27.63) μM | [6] | |
| Chaetoglobosin O (16) | Penicillium sp. | Marine alga | Indole alkaloid | P388 | 2.4 μg/mL | [7] | |
| Cytoglobosin C (17) | P. chrysogenum V11 | Mangrove | Indole alkaloid | MDA-MB-435, SGC-7901, A549 | (12.58, 8.15, 3.35) μM | [6] | |
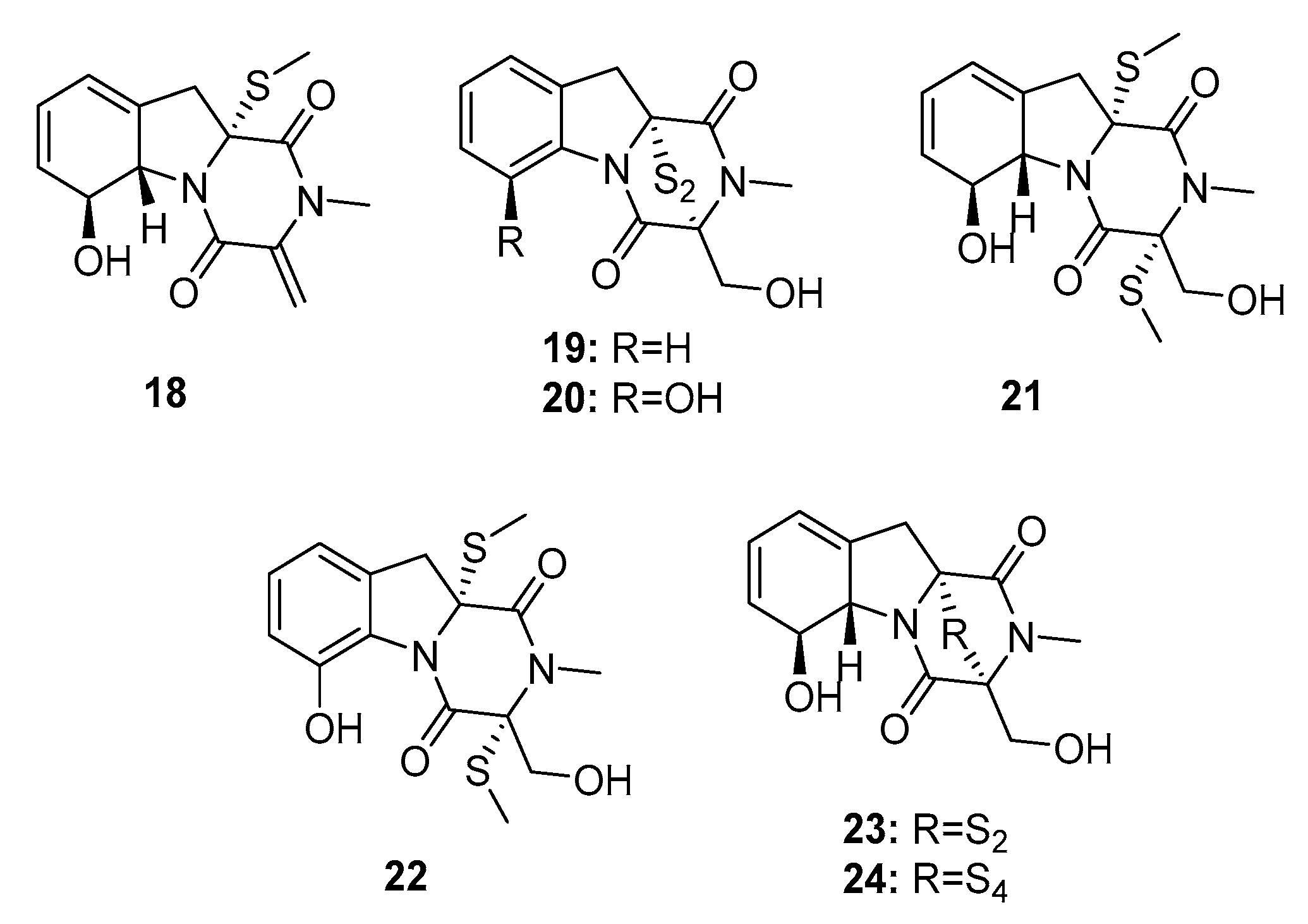
| 18 | Penicillium sp. JMF034 | Deep sea sediment | Diketopiperazine | P388 | 3.4 μM | [12] | |
| 19 | Penicillium sp. JMF034 | Deep sea sediment | Diketopiperazine | P388 | 0.058 μM | HMT G9a (IC50 = 55 μM) | [12] |
| 20 | Penicillium sp. JMF034 | Deep sea sediment | Diketopiperazine | P388 | 0.11 μM | [12] | |
| 21 | Penicillium sp. JMF034 | Deep sea sediment | Diketopiperazine | P388 | 0.11 μM | HMT G9a (IC50 = 58 μM) | [12] |
| 22 | Penicillium sp. JMF034 | Deep sea sediment | Diketopiperazine | P388 | 0.056 μM | HMT G9a (IC50 = 2.6 μM) | [12] |
| Gliotoxin (23) | Penicillium sp. JMF034 | Deep sea sediment | Diketopiperazine | P388 | 0.024 μM | HMT G9a (IC50 = 6.4 μM) Dual inhibitor of farnesyltransferase and geranylgeranyltransferase I | [12,13] |
| Gliotoxin G (24) | Penicillium sp. JMF034 P. brocae MA-231 | Deep sea sediment Mangrove | Diketopiperazine | P388 A2780, A2780 CisR | 0.02 μM (0.664, 0.661) μM | HMT G9a (IC50 = 2.1 μM) | [12] [14] |
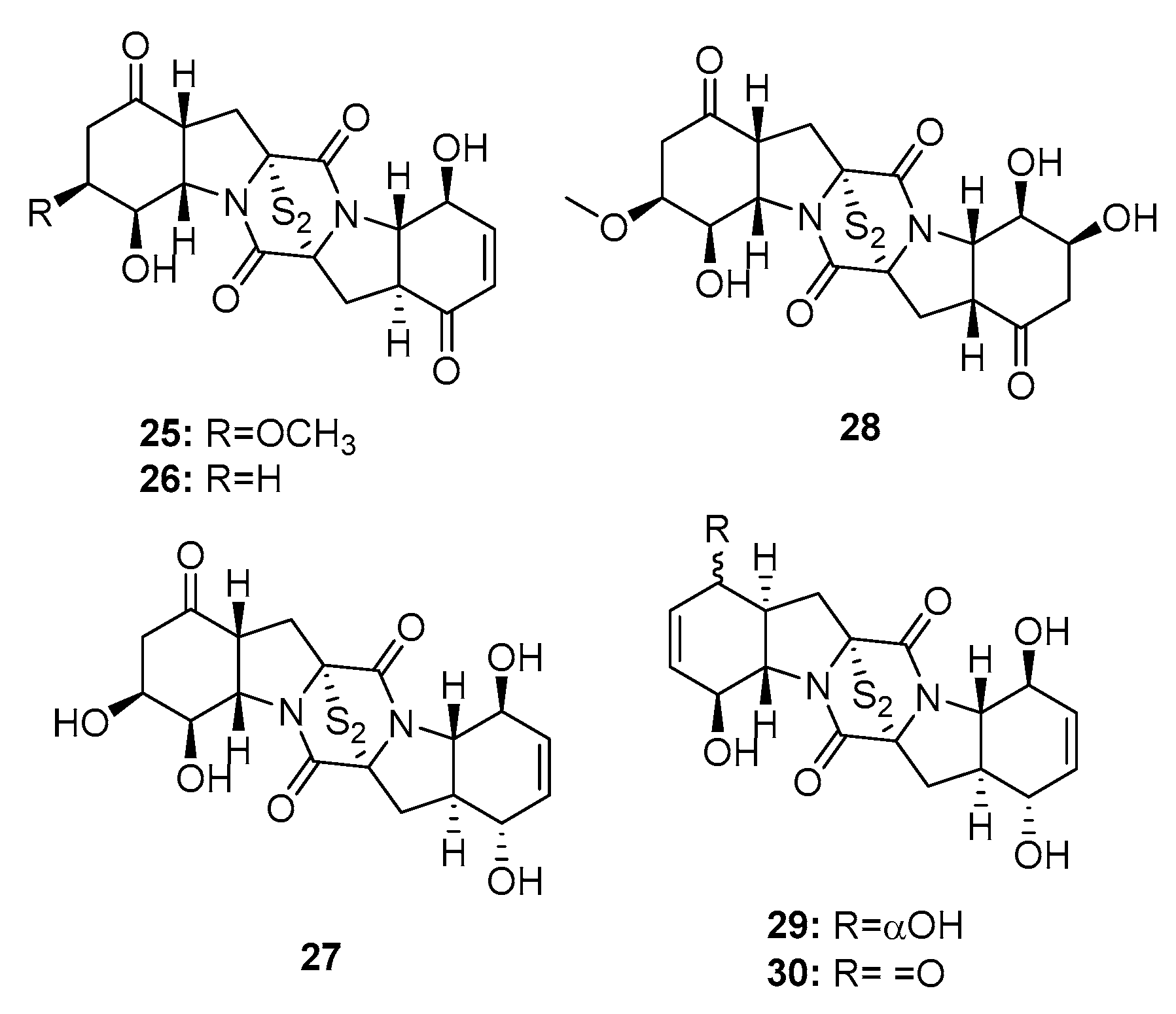
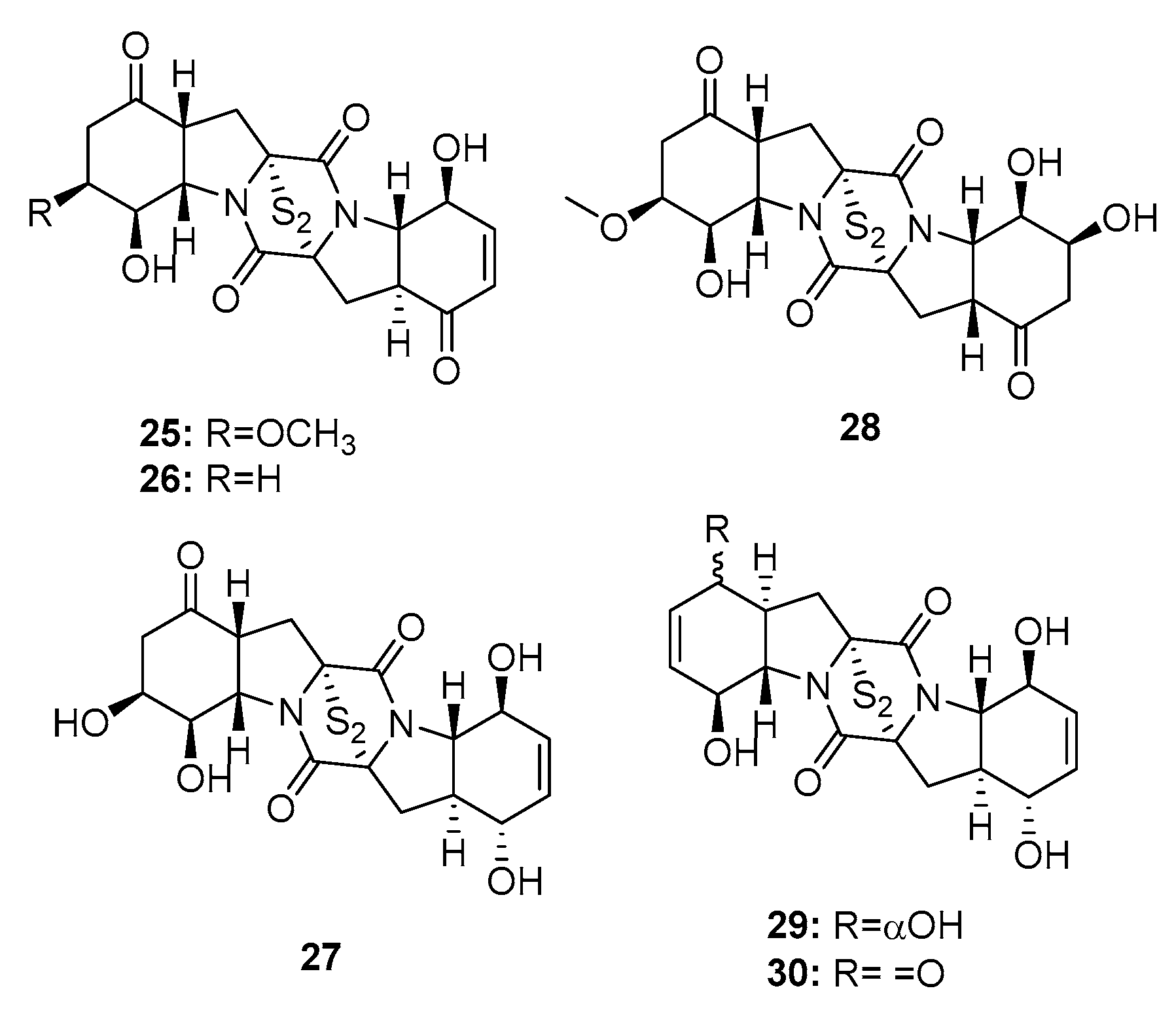
| Brozazine A (25) | P. brocae MA-231 | Mangrove | Diketopiperazine | Du145, Hela, HepG2, MCF-7, NCI-H460, SGC-7901, SW1990, SW480, U251 | (4.2, 6.8, 6.4, 5.5, 4.9, 2.6, 6.0, 2.0, 5.2) μM | [16] | |
| Brozazine B (26) | P. brocae MA-231 | Mangrove | Diketopiperazine | Du145, Hela, HepG2, MCF-7, NCI-H460, SGC-7901, SW1990, SW480, U251 | (3.6, 5.3, 5.5, 6.1, 4.0, 2.4, 6.4, 1.2, 3.5) μM | [16] | |
| Brozazine E (29) | P. brocae MA-231 | Mangrove | Diketopiperazine | Du145, Hela, HepG2, MCF-7, NCI-H460, SGC-7901, SW1990, U251 | (11.2, 4.3, 5.6, 9.0, 12.4, 3.3, 2.1, 6.1) μM | [16] | |
| Brozazine F (30) | P. brocae MA-231 | Mangrove | Diketopiperazine | Du145, Hela, HepG2, MCF-7, NCI-H460, SGC-7901, SW1990, U251 | (1.7, 6.9, 2.9, 3.0, 0.89, 8.0, 5.9, 5.3) μM | [16] | |
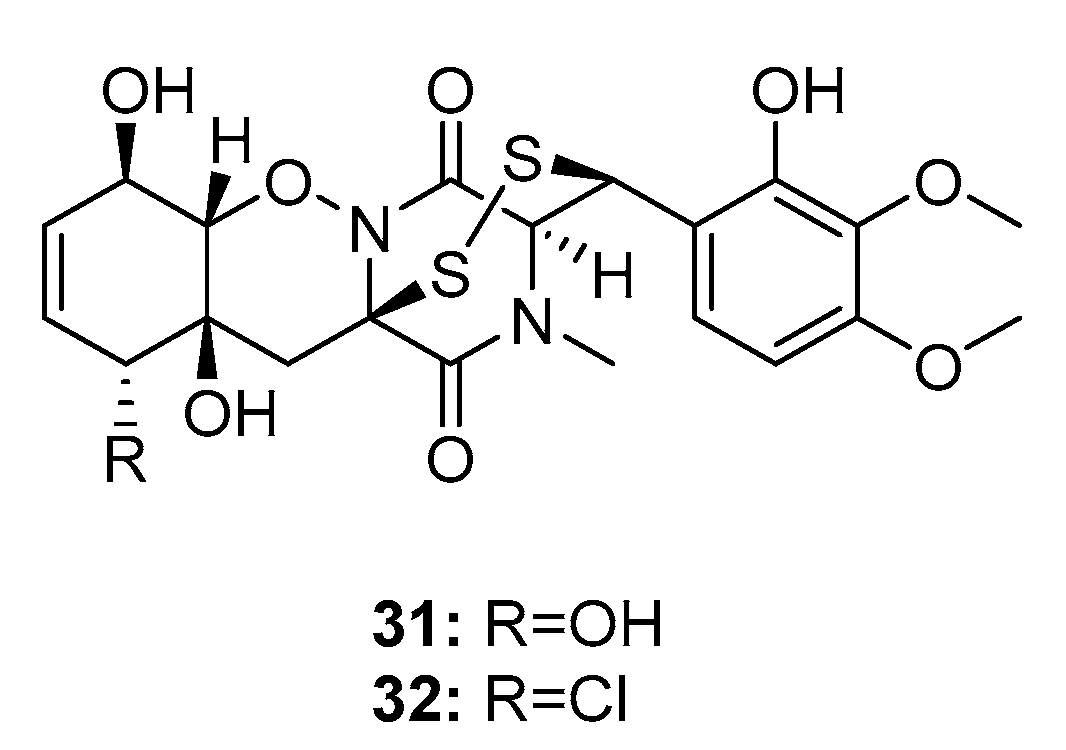
| N-methylpretrichodermam ide B/adametizines A (32) | P. adametzioides AS-53 Penicillium sp. | Marine sponge/sediment/alga | Diketopiperazine | Artemia salina | 4.8 μM | [17,18,19] | |
| L5178Y, 22Rv1, PC-3, LNCaP | (2, 0.51, 5.11, 1.76) μM | ||||||
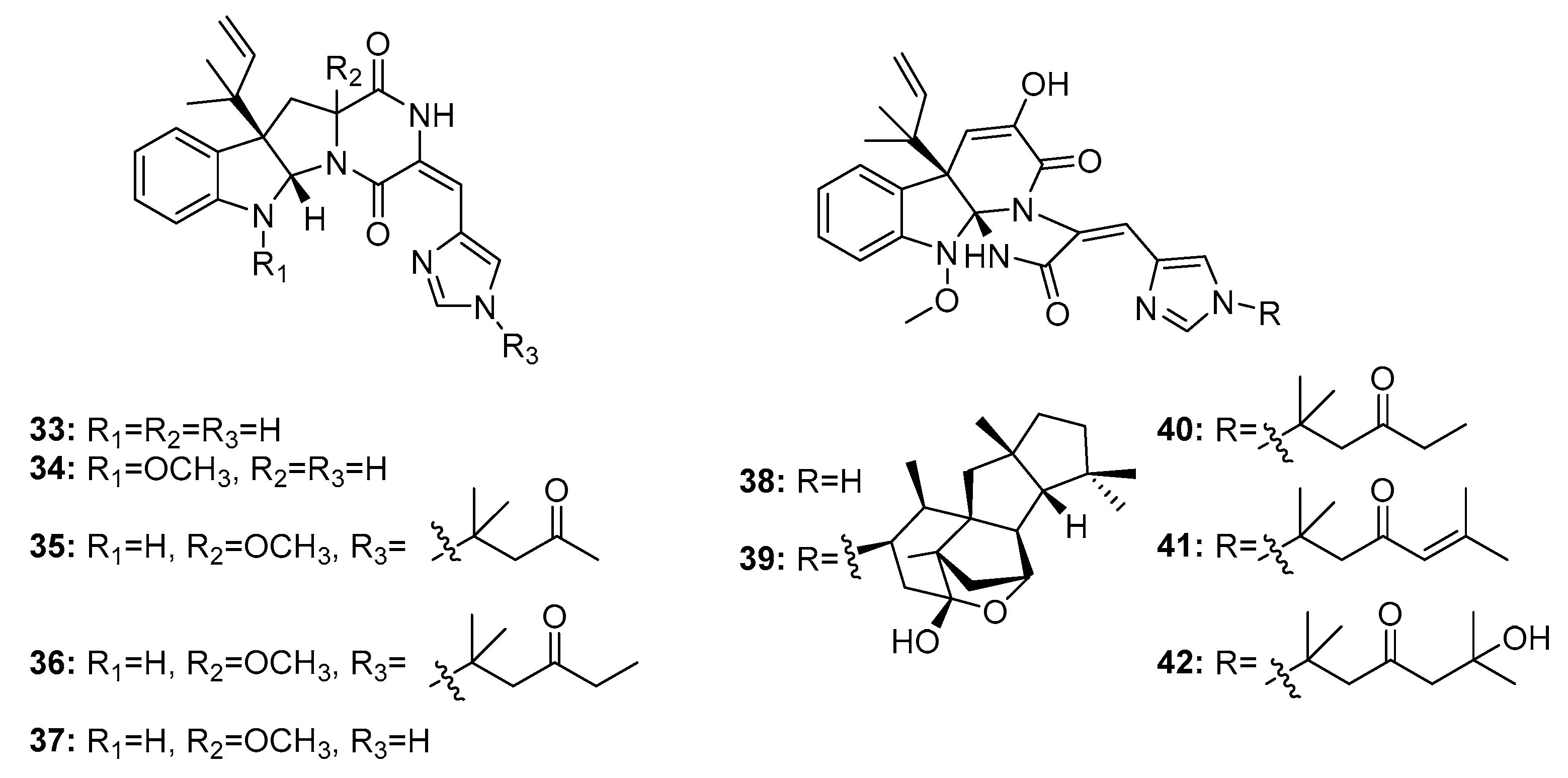
| Roquefortine C (33) | Penicillium sp. | Deep sea sediment | Diketopiperazine | Activate P-glycoprotein and inhibit P450-3A and other haemoproteins | [20,22] | ||
| Roquefortine F (34) | Penicillium sp. | Deep sea sediment | Diketopiperazine | A549, HL-60, BEL-7402, MOLT-4 | (14.0, 33.6, 13.0, 21.2) μM | [22] | |
| Roquefortine G (35) | Penicillium sp. | Deep sea sediment | Diketopiperazine | A549, HL-60 | (42.5, 36.6) μM | [22] | |
| Meleagrin (38) | Penicillium sp. P. commune SD-118 | Deep sea sediment | Indole alkaloid | A549, HL-60 HepG2, NCI-H460, Hela, DU145, MDA-MB-231, | (19.9, 7.4) μM (12.0, 22.0, 20.0, 11.0, 5.0) μg/mL | Arrest the cell cycle through G2/M phase Inhibitor of tubulin polymerization | [21,22,23] |
| Meleagrin B (39) | Penicillium sp. | Deep sea sediment | Indole alkaloid | A549, HL-60, BEL-7402, MOLT-4 | (2.7, 6.7, 1.8, 2.9) μM | [21,22] | |
| Meleagrin C (40) | Penicillium sp. | Deep sea sediment | Indole alkaloid | A549, BEL-7402, MOLT-4 | (9.9, 10.0, 4.7) μM | [22] | |
| Meleagrin D (41) | Penicillium sp. | Deep sea sediment | Indole alkaloid | A549 | 32.2 μM | [21] | |
| Meleagrin E (42) | Penicillium sp. | Deep sea sediment | Indole alkaloid | A549 | 55.9 μM | [21] | |
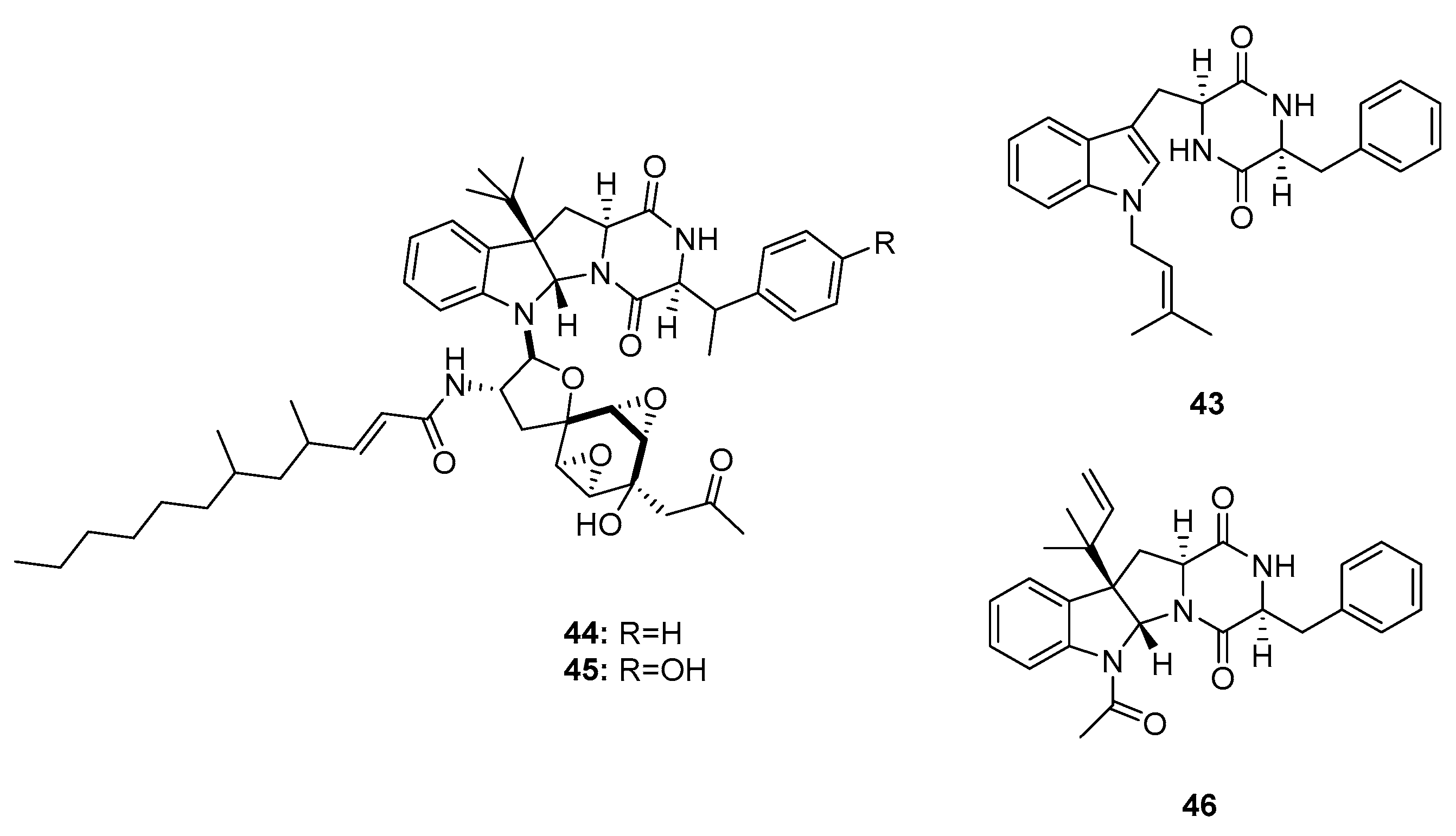
| Penicimutanin (43) | Mutant P. purpurogenum G59 | Marine soil | Diketopiperazine | K562, Hela, MCF-7 | IR% (100 μg/mL): 22.6%, 17.9%, 26.5% | [24] | |
| Penicimutanin A (44) | Mutant P. purpurogenum G59 | Marine soil | Diketopiperazine | K562, HL-60, Hela, BGC-823, MCF-7 | (11.4, 5.4, 9.5, 8.0, 5.4) μM | [24] | |
| Penicimutanin B (45) | Mutant P. purpurogenum G59 | Marine soil | Diketopiperazine | K562, HL-60, Hela, BGC-823, MCF-7 | (19.9, 12.1, 17.7, 16.6, 8.0) μM | [24] | |
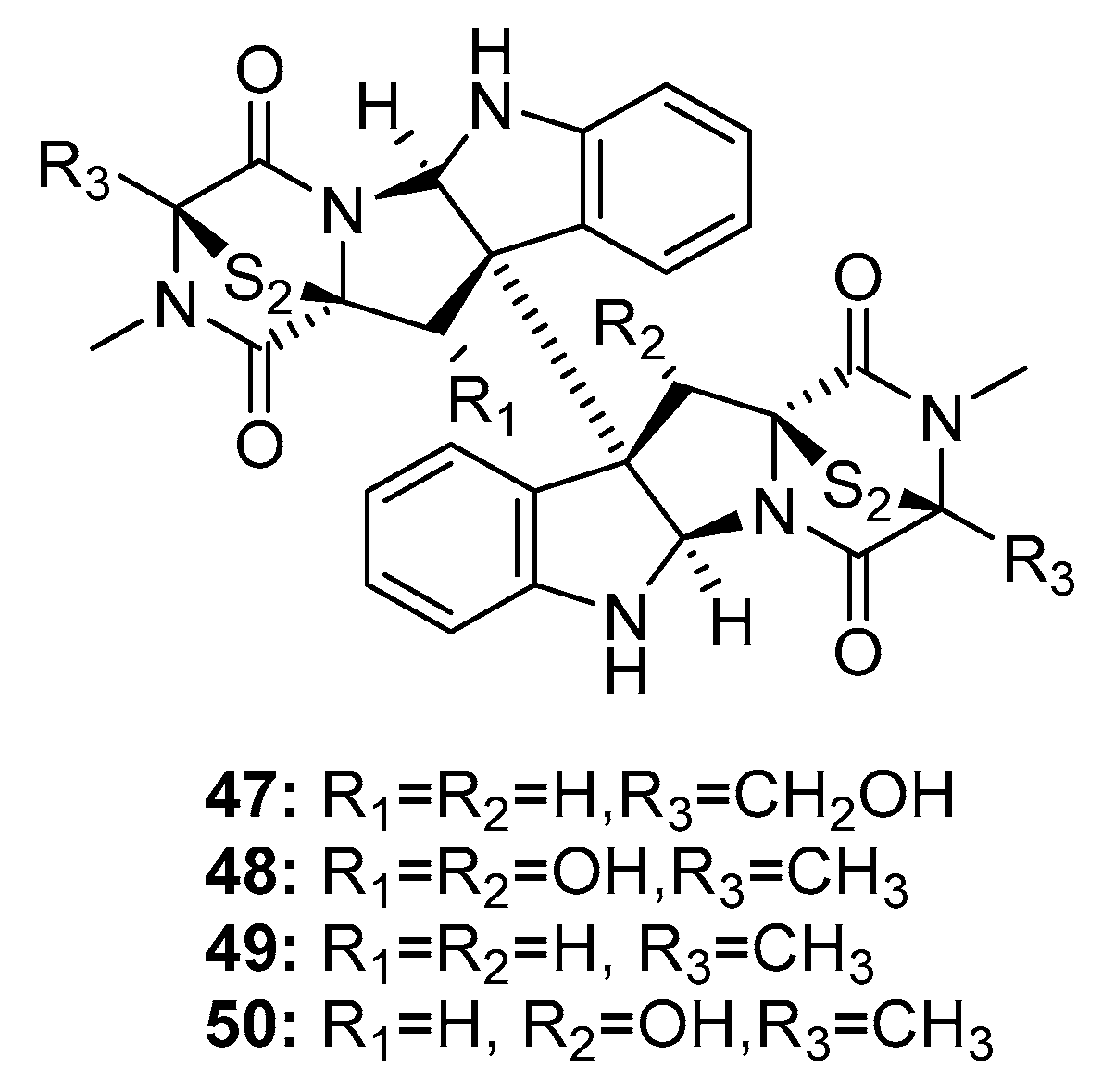
| Fructigenine A (46) | Mutant P. purpurogenum G59 | Marine soil | Diketopiperazine | K562, Hela, MCF-7, BGC-823 | IR% (100μg/mL): 20.8%, 55.3%, 65.6%, 34.8% | [24,25] | |
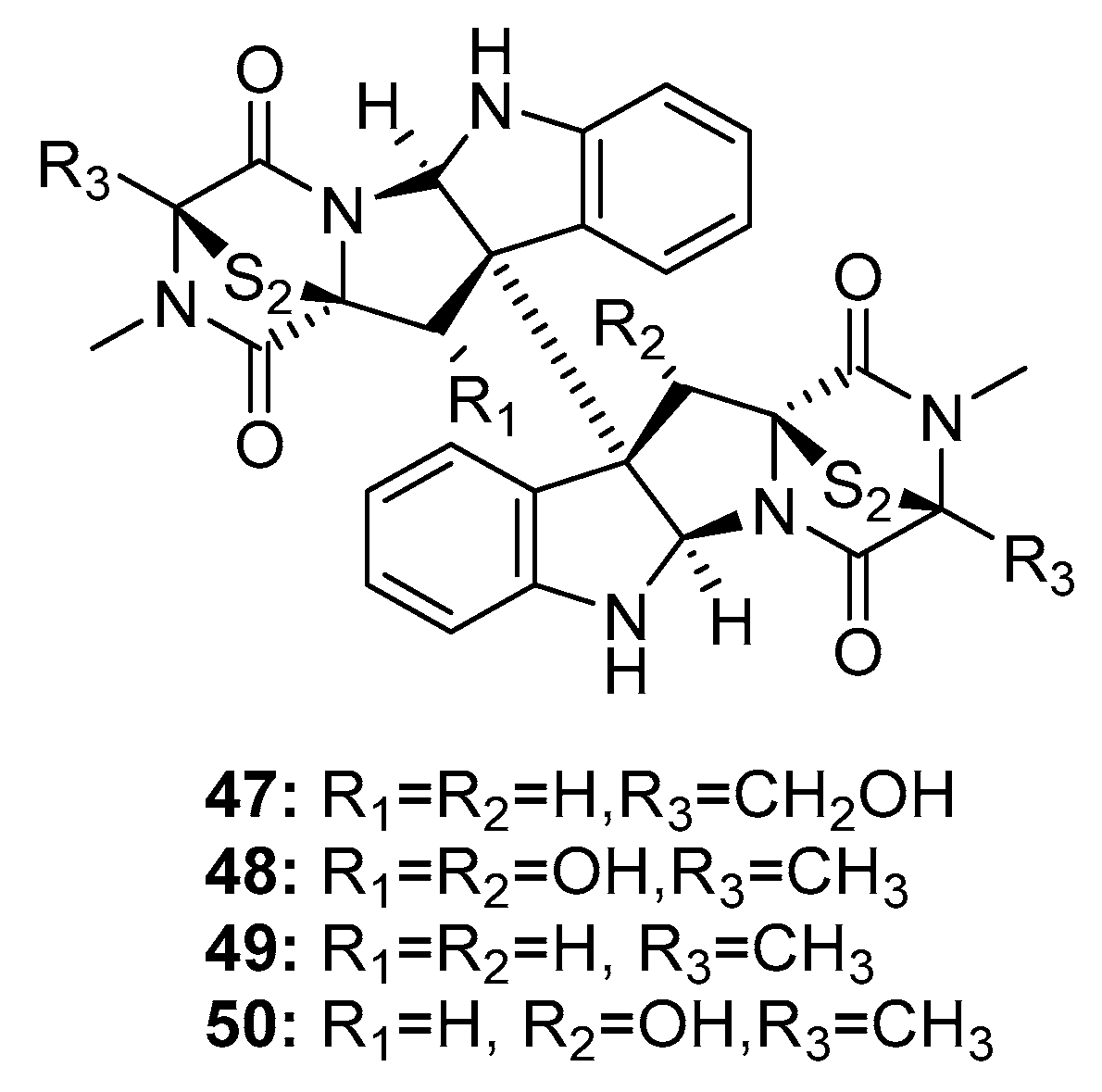
| 11,11′-dideoxyverticillin A (49) | Penicillium sp. | Marine alga | Diketopiperazine | HCT-116 | 30 ng/mL | Induce G2/M arrest through p38 MAPK pathway; Epidermal growth factor receptor tyrosine kinase inhibitor | [28,30,31] |
| 11′-deoxyverticillin A (50) | Penicillium sp. | Marine alga | Diketopiperazine | HCT-116 | 30 ng/mL | [28] | |
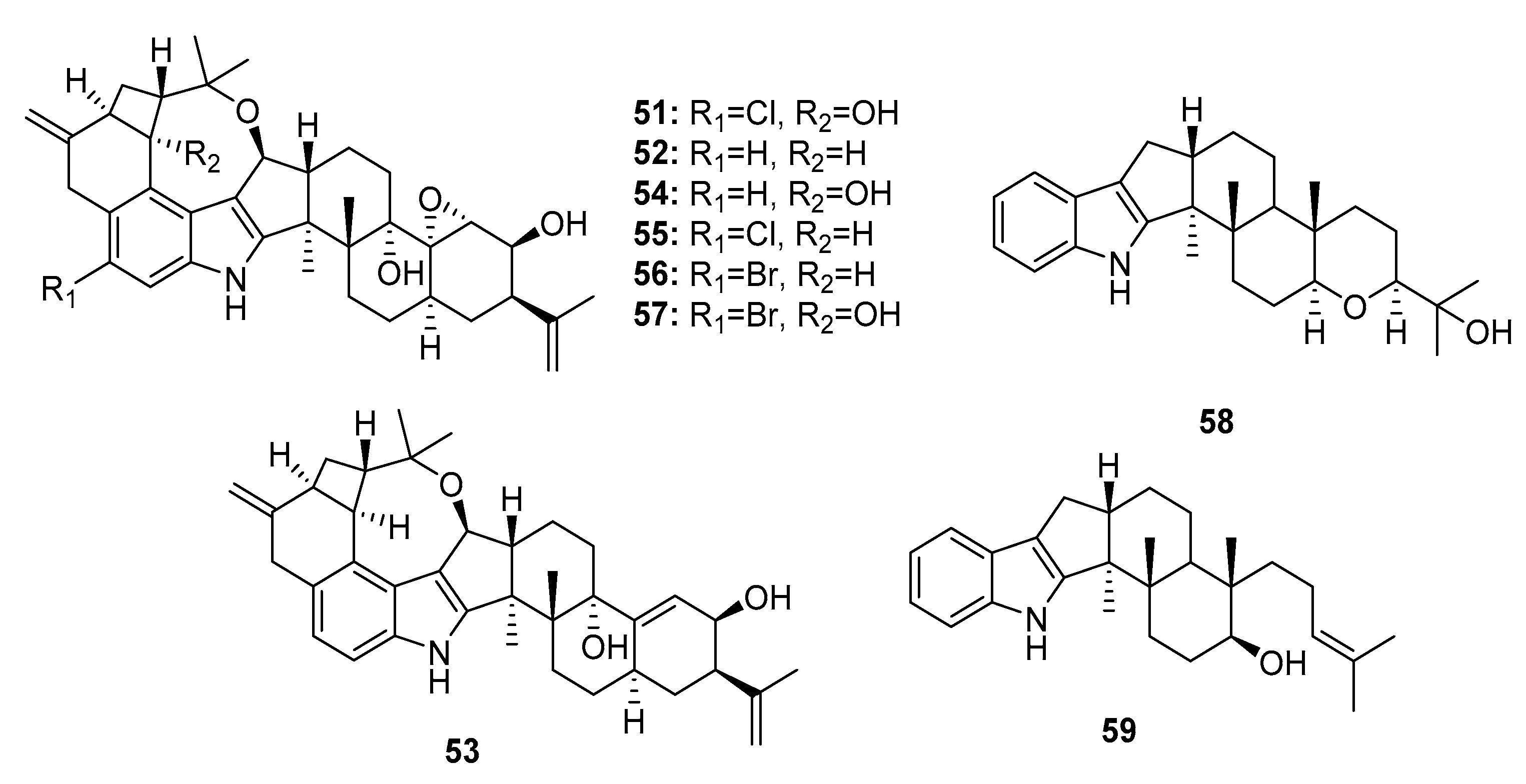
| Penitrem A (51) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) MDA-MB-231 (anti-invasion) | (11.9, 9.8) μM 8.7 μM IR% (15 μM)> 75% | BK channel inhibitor | [34] |
| Penitrem B (52) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF-7, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) | (5.5, 13.7) μM 10.3 μM | [34] | |
| Penitrem D (53) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF-7, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) | (8.3, 29.7) μM 9.2 μM | [34] | |
| Penitrem E (54) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF-7, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) | (17.5, 25.4) μM 20.3 μM | [34] | |
| Penitrem F (55) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF-7, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) | (15.0, 13.8) μM 35.0 μM | [34] | |
| 6-bromopenitrem B (56) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF-7, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) MDA-MB-231 (anti-invasion) | (19.3, 18.8) μM 30.3 μM IR%(15 μM) > 40% | [34] | |
| 6-bromopenitrem E (57) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF-7, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) | (16.7, 8.5) μM 9.6 μM | BK channel inhibitor | [34] |
| Paspaline (58) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF-7, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) | (12.8, 12.4) μM 7.6 μM | [34] | |
| Emnidole SB (59) | P. commune isolate GS20 | Sponge/Sediment | Indole alkaloid | MCF-7, MDA-MB-231 (antiproliferative) MDA-MB-231 (antimigratory) | (10.1, 21.3) μM 19.0 μM | [34,37] | |
| Communesin A (60) | Penicillium sp. | Marine alga/Sediment | Indole alkaloid | P388 | 3.5 μg/mL | [38,40] | |
| Communesin B (61) | Penicillium sp. | Marine alga/Sponge/Sediment | Indole alkaloid | P388, U-937, THP-1, NAMALWA, MOLT-3, SUP-B15 | (0.45, 10.4, 11.4, 9.9, 8.1, 7.2) μg/mL | [38,39,40] | |
| Communesin C (62) | Penicillium sp. | Marine sponge | Indole alkaloid | U-937, THP-1, NAMALWA, MOLT-3, SUP-B15 | (11.3, 13.1, 8.2, 8.6, 10.8) μg/mL | [39] | |
| Communesin D (63) | Penicillium sp. | Marine sponge | Indole alkaloid | U-937, THP-1, NAMALWA, MOLT-3, SUP-B15 | (13.1, 16.2, 14.6, 9.9, 9.0) μg/mL | [39] | |
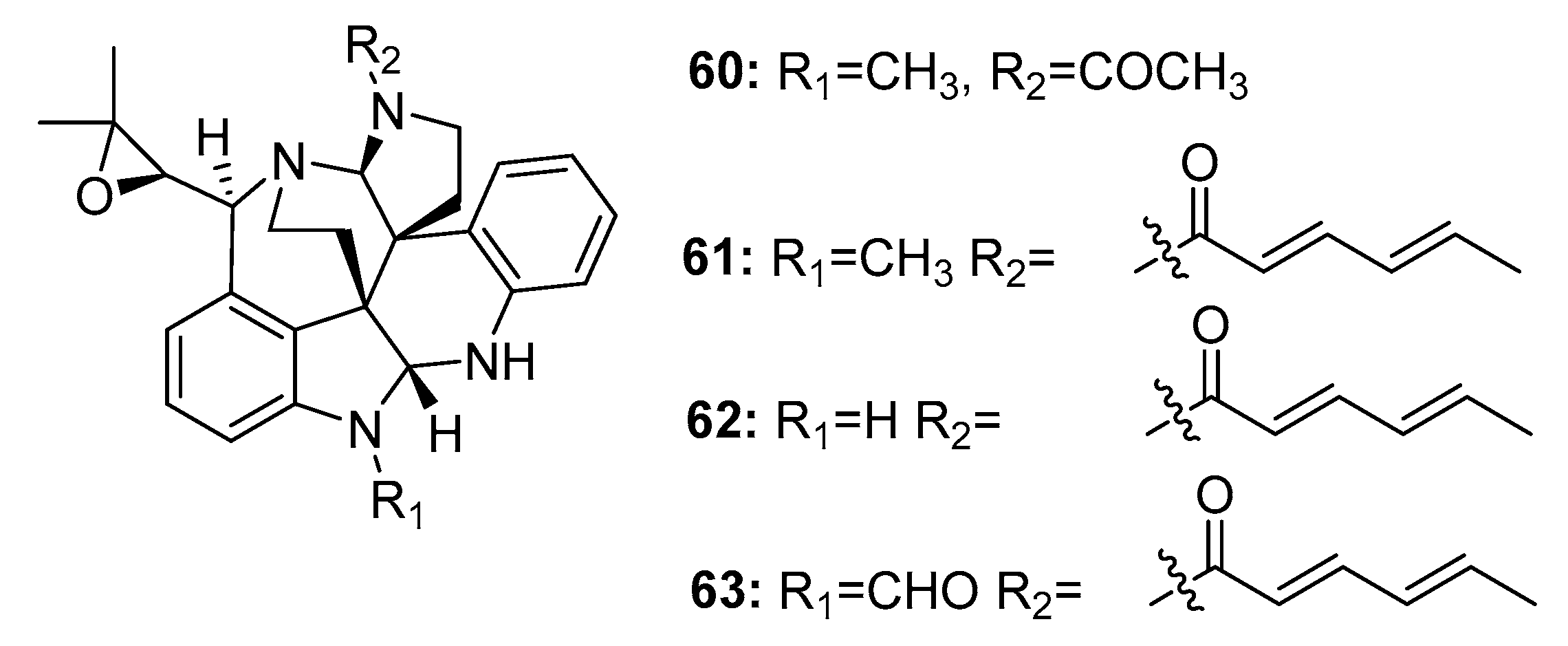
| Penioxamide (64) | P. oxalicum EN-201 | Mangrove | Indole alkaloid | A. salina | 5.6 μM | [42] | |
| 65 | P. brefeldianum SD-273 | Marine sediment | Indole alkaloid | A. salina | 9.4 μM | [43] | |
| Penipaline B (66) | P. paneum SD-44 | Marine sediment | Indole alkaloid | A549, HCT-116 | (20.44, 14.88) μM | [44] | |
| Penipaline C (67) | P. paneum SD-44 | Marine sediment | Indole alkaloid | A549, HCT-116 | (21.54, 18.54) μM | [44] | |
| Terretrione A (68) | P. vinaceum | Marine sponge | 1,4-diazepane alkaloid | MDA-MB-231 | 17.7 μM | [45] | |
| Terretrione C (69) | Penicillium sp.CYE-87 | Marine tunicate | 1,4-diazepane alkaloid | MDA-MB-231 | 17.6 μM | [46] | |
| Terretrione D (70) | Penicillium sp.CYE-87 | Marine tunicate | 1,4-diazepane alkaloid | MDA-MB-231 | 16.5 μM | [46] | |
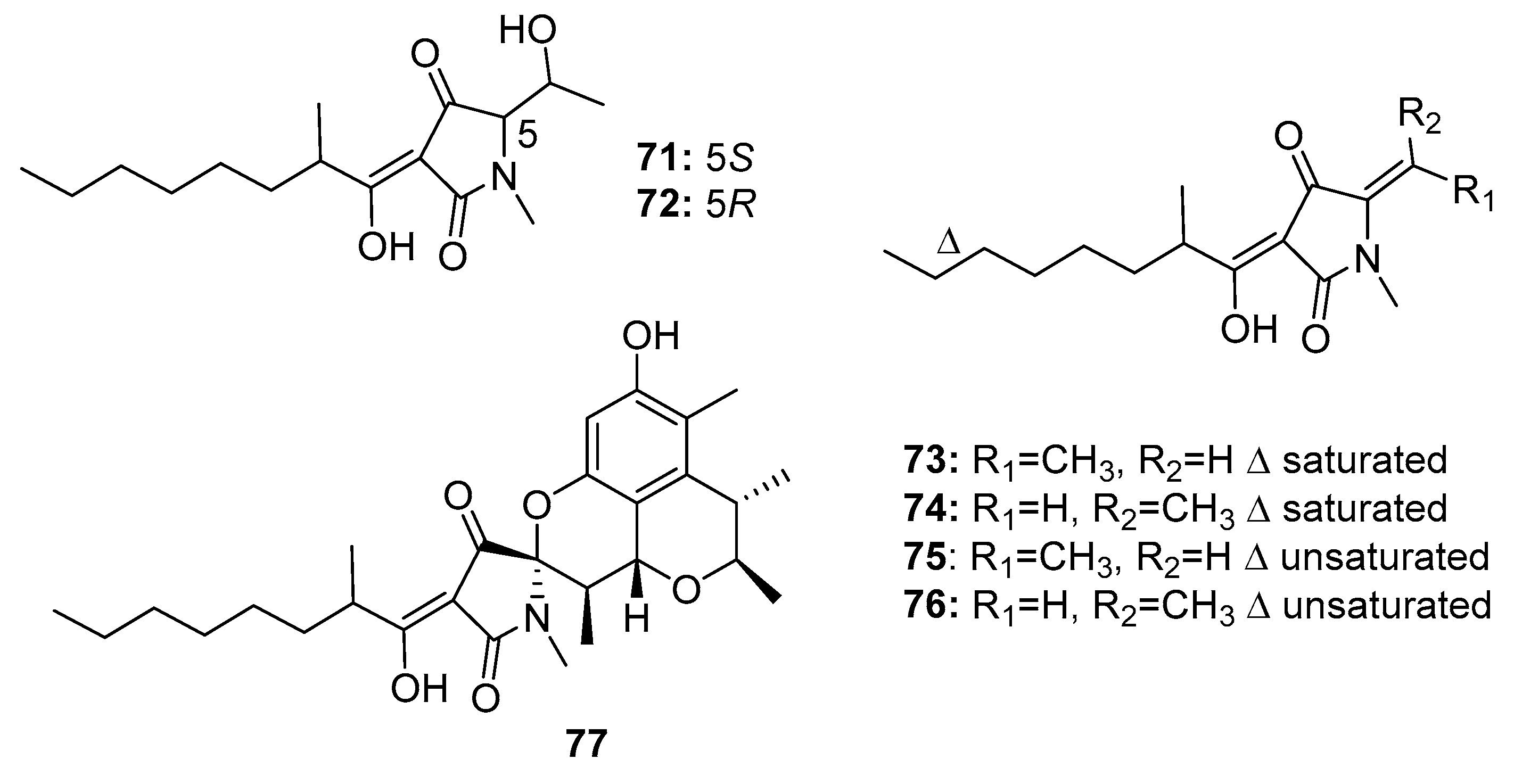
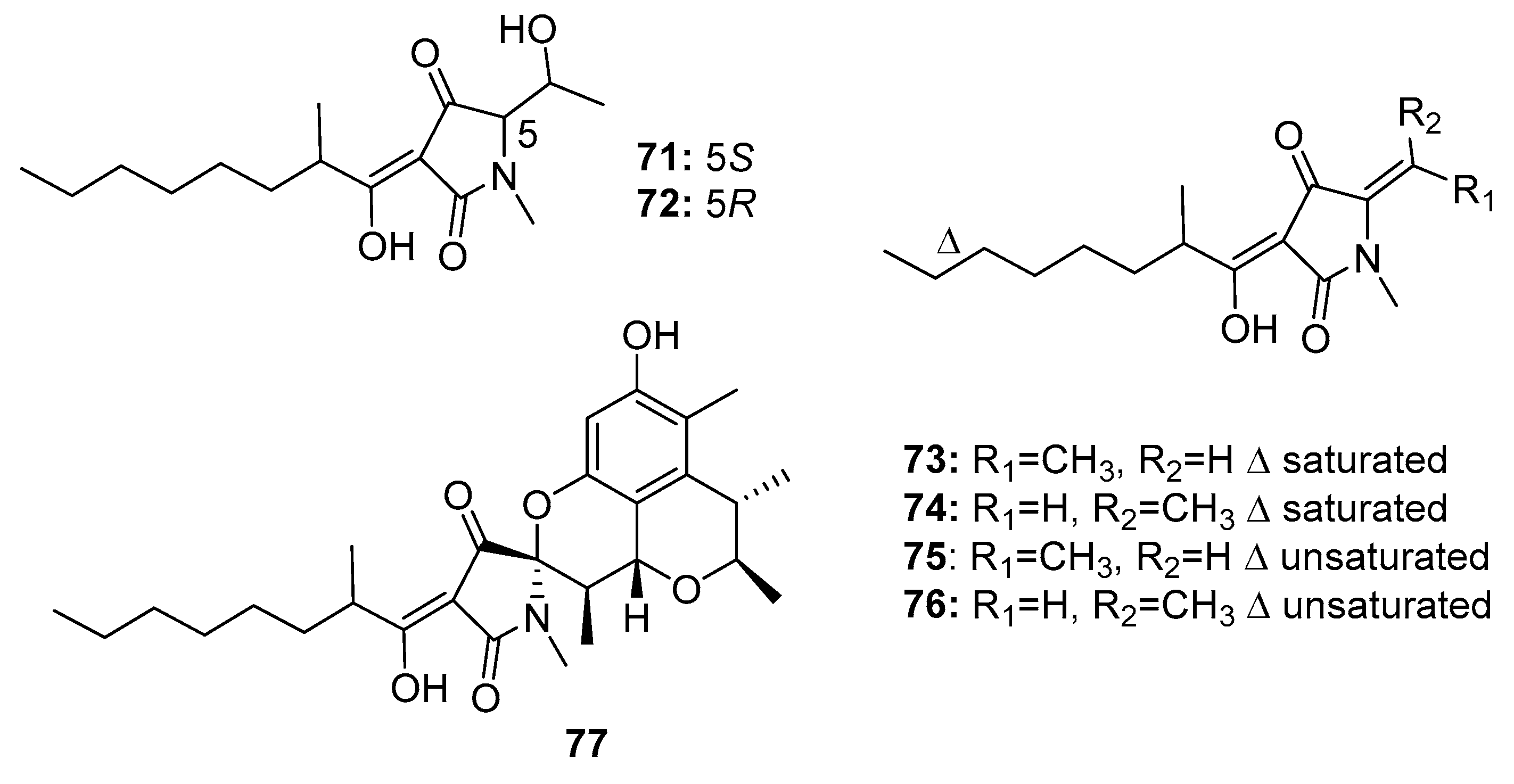
| Penicillenol A1 (71) | Penicillium sp. GQ-7/P. citrinum | Mangrove/Marine sediment | Pyrrolidinone alkaloid | A-375, HL-60, A549, BEL-7402, P388 | 3.2 μg/mL (0.76, 23.8, 13.03, 8.85) μM | [47,49] | |
| Penicillenol A2 (72) | Penicillium sp. GQ-7/P. citrinum | Mangrove/Marine sediment | Pyrrolidinone alkaloid | A-375 HL-60 | 13.8 μg/mL 16.26 μM | [47,49] | |
| Penicillenol B1 (73) | Penicillium sp. GQ-7/P. citrinum | Mangrove/Marine sediment | Pyrrolidinone alkaloid | A-375 HL-60 | 2.8 μg/mL 3.2 μM | [47,49] | |
| Penicillenol B2 (74) | Penicillium sp. GQ-7/P. citrinum | Mangrove/Marine sediment | Pyrrolidinone alkaloid | A-375 HL-60 | 0.97 μg/mL 7.65 μM | [47,49] | |
| Penicillenol D1 (75) | P. citrinum | Marine sediment | Pyrrolidinone alkaloid | A549, HL-60 | (17.2, 18.5) μg/mL | [48] | |
| Penicillenol D2 (76) | P. citrinum | Marine sediment | Pyrrolidinone alkaloid | A549, HL-60 | (12.1, 14.5) μg/mL | [48] | |
| Penitrinine A (77) | P. citrinum | Marine sediment | Pyrrolidinone alkaloid | A-375, SPC-A1, HGC-27 | (20.12, 28.67, 29.49) μM | Upregulate Bax, downregulate Bcl-2, suppress MMP-9 and TIMP-1 | [50] |
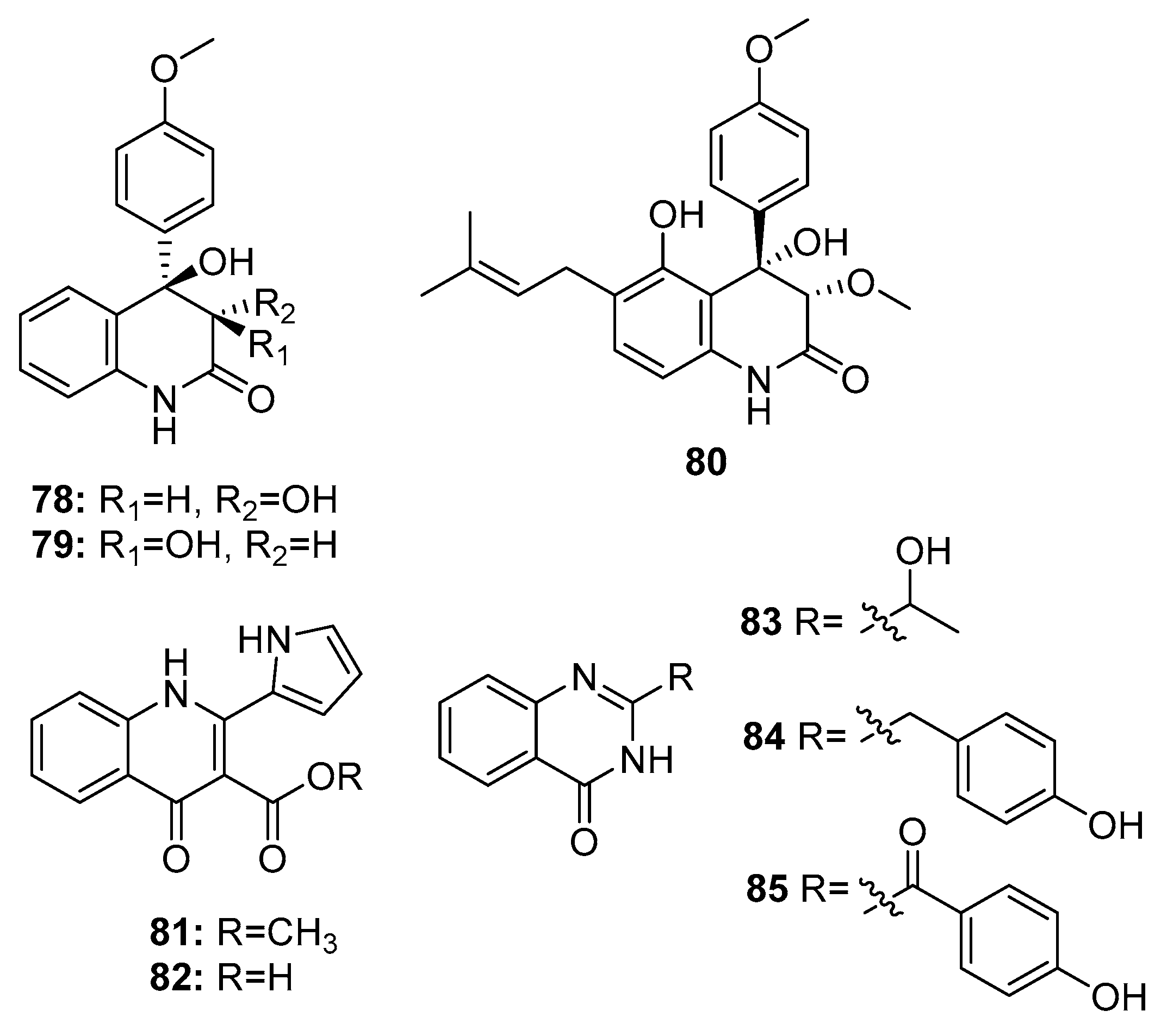
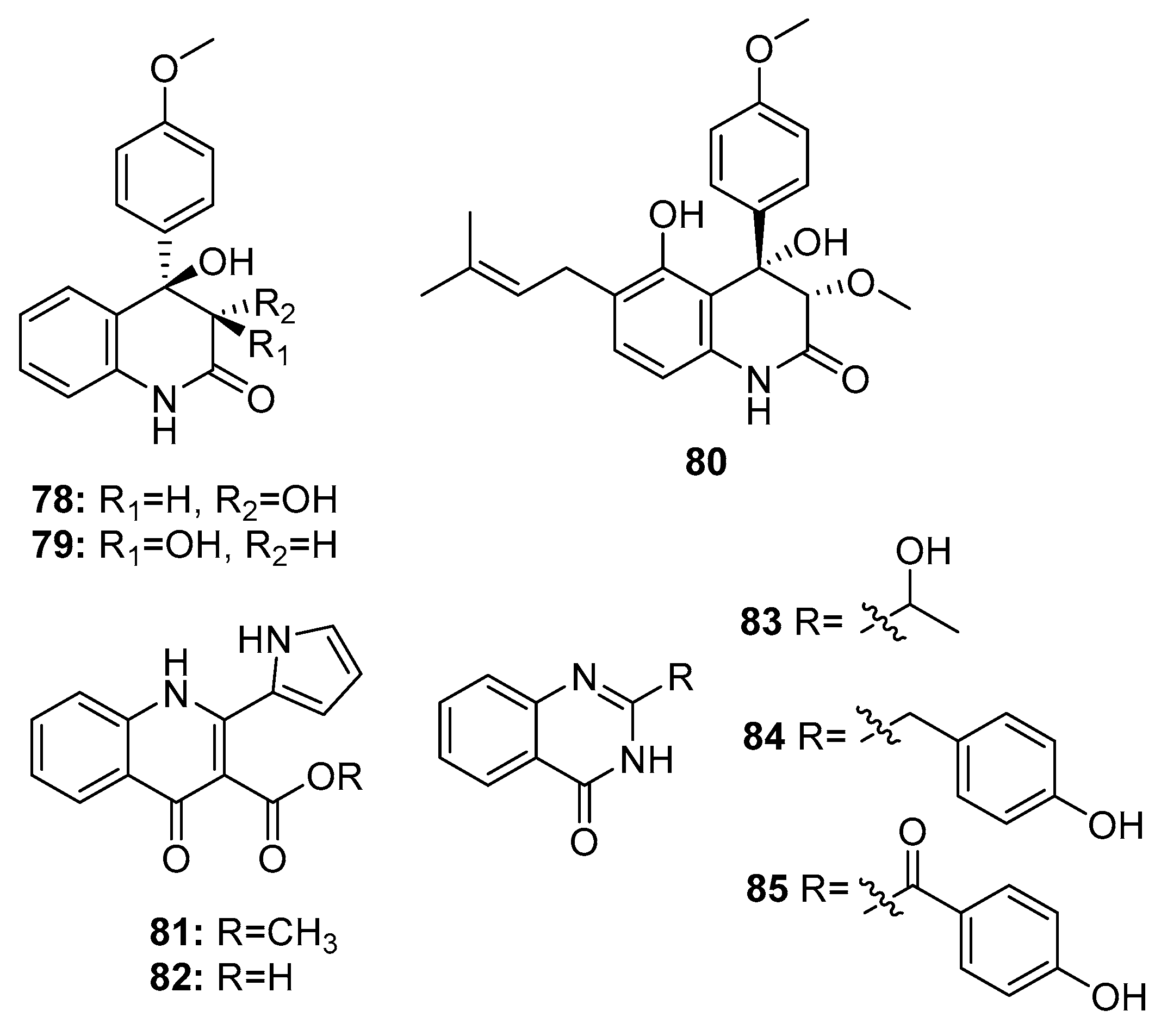
| 78 | P. janczewskii | Sea water | Quinolinone | MDA-MB-231, DU-145, SKOV-3, HT-29, A549, CAKI-1, SK-MEL-2, K562 | IR % (10 μg/mL) = 20~50% | [52] | |
| 79 | P. janczewskii | Sea water | Quinolinone | MDA-MB-231, DU-145, SKOV-3, HT-29, A549, CAKI-1, SK-MEL-2, K562 | IR % (10 μg/mL) = 30~90% | [52] | |
| 80 | P. janczewskii | Sea water | Quinolinone | MDA-MB-231, DU-145, SKOV-3, HT-29 | IR % (10 μg/mL) = 91.6%, 69.2%, 79.8%, 96.0% | [52] | |
| 81 | Penicillium sp. ghq208/Penicillium sp. | Marine sediment/Mangrove | Quinolinone | 95-D, HepG2 | (0.57, 6.5) μg/mL | [53,54] | |
| 82 | Penicillium sp. ghq208 | Marine sediment | Quinolinone | HepG2 | 13.2 μM | [53] | |
| 83 | P. commune SD-118 | Deep sea sediment | Quinazolinone | SW1990 | 20 μg/mL | [23] | |
| 84 | P. chrysogenum EN-118 | Marine alga | Quinazolinone | DU145, A549, Hela | 8 μg/mL | [56] | |
| 85 | P. oxalicum 0312F1 | Marine (not clear) | Quinazolinone | SGC-7901, BEL-7404 | IR % (200 μg/mL) = 30~40% | [55] | |
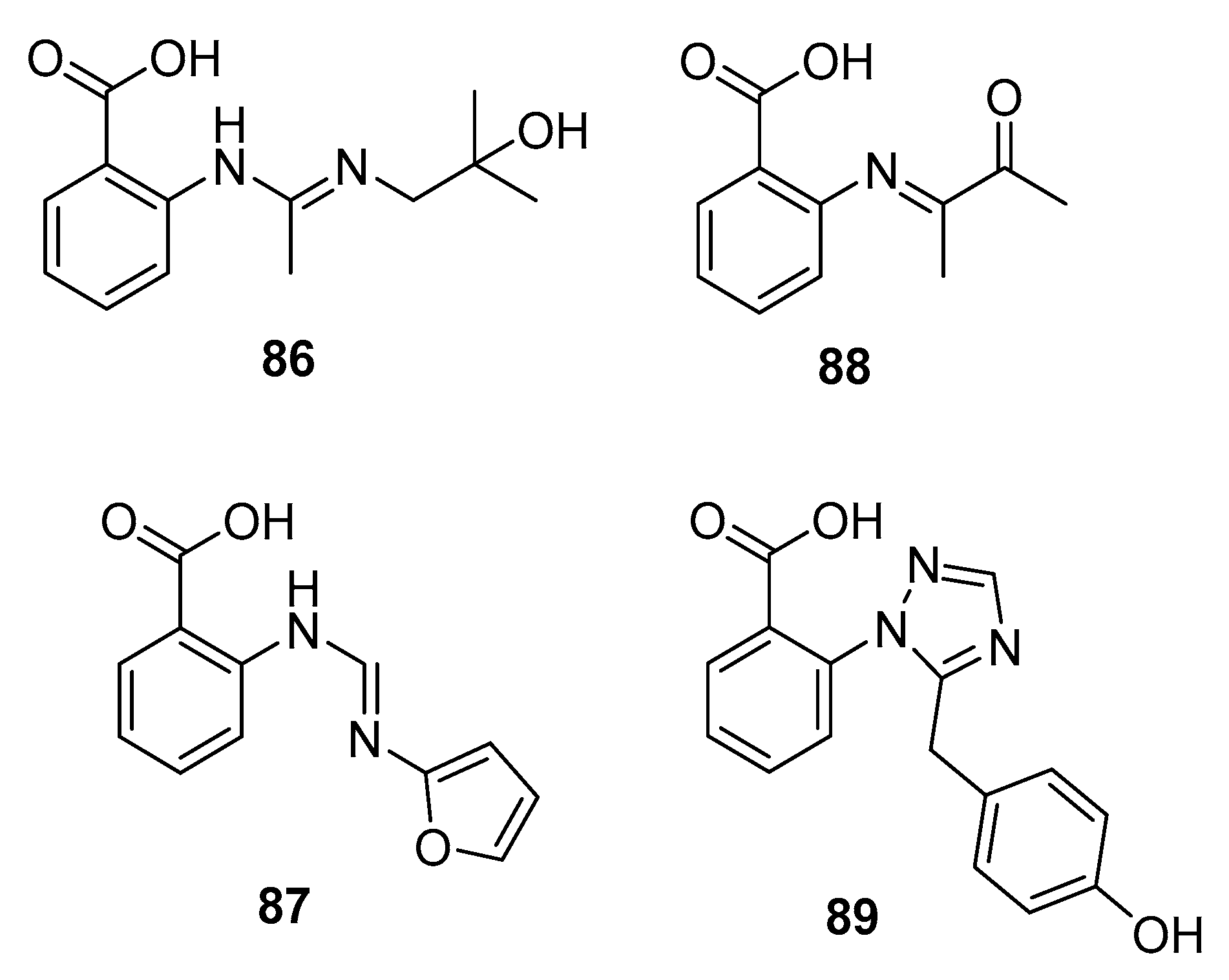
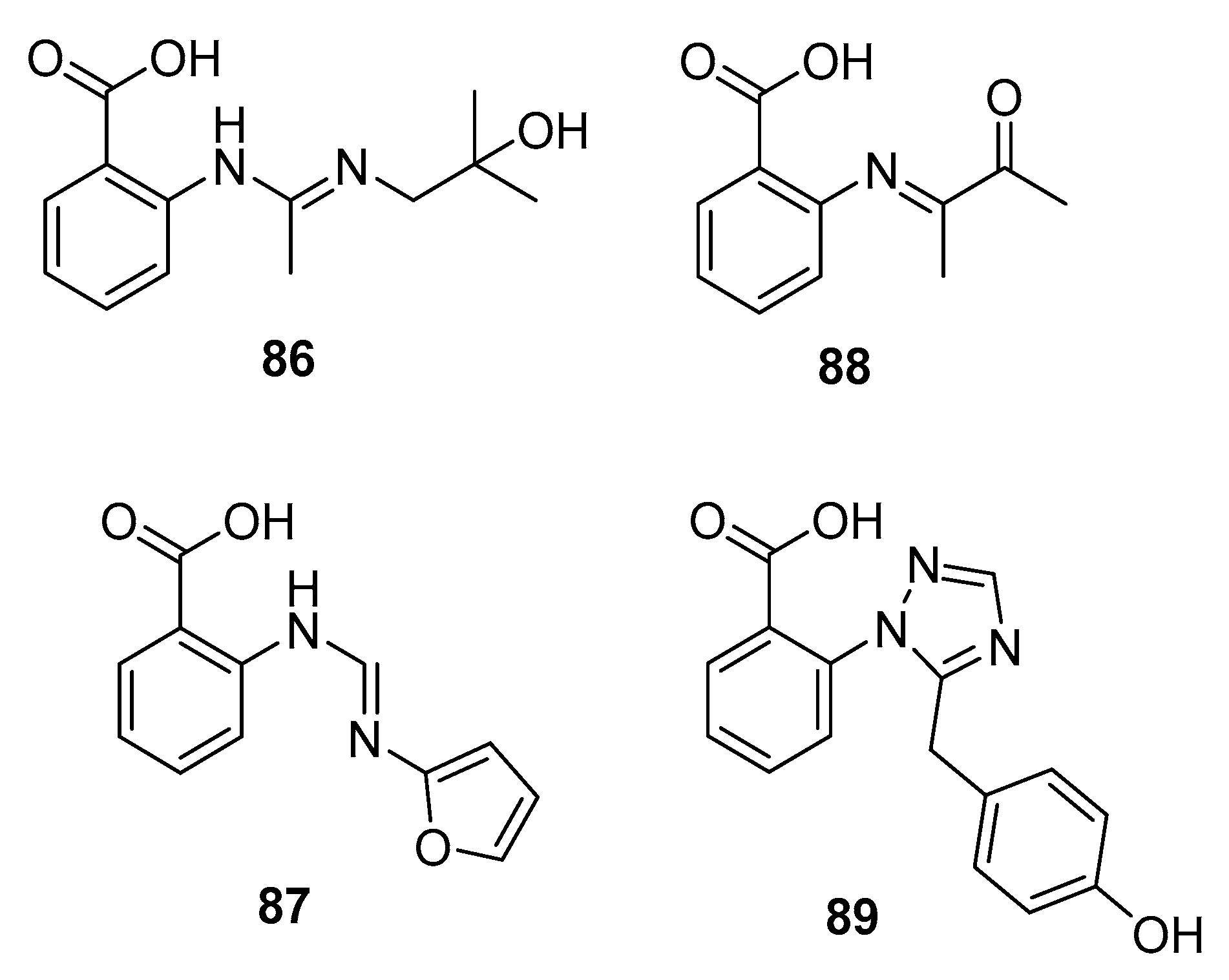
| Penipacid A (86) | P. paneum SD-44 | Deep sea sediment | Amidine alkaloid | RKO | 8.4 μM | [57] | |
| Penipacid E (87) | P. paneum SD-44 | Deep sea sediment | Amidine alkaloid | RKO | 9.7 μM | [57] | |
| 88 | P. paneum SD-44 | Deep sea sediment | Imine alkaloid | Hela | 6.6 μM | [57] | |
| Penipanoid A (89) | P. paneum SD-44 | Deep sea sediment | Triazole alkaloid | SMMC-7721 | 54.2 μM | [58] | |
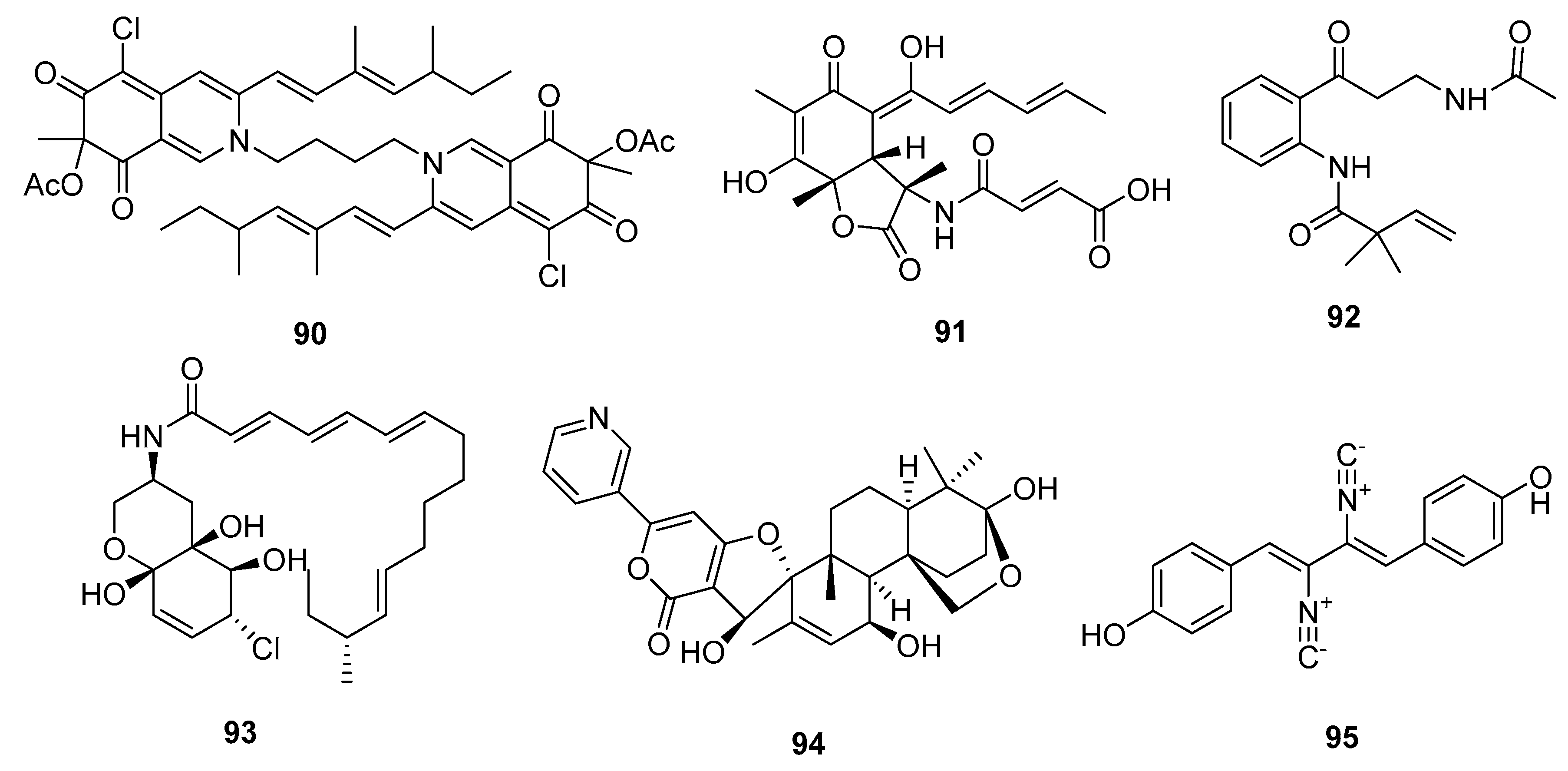
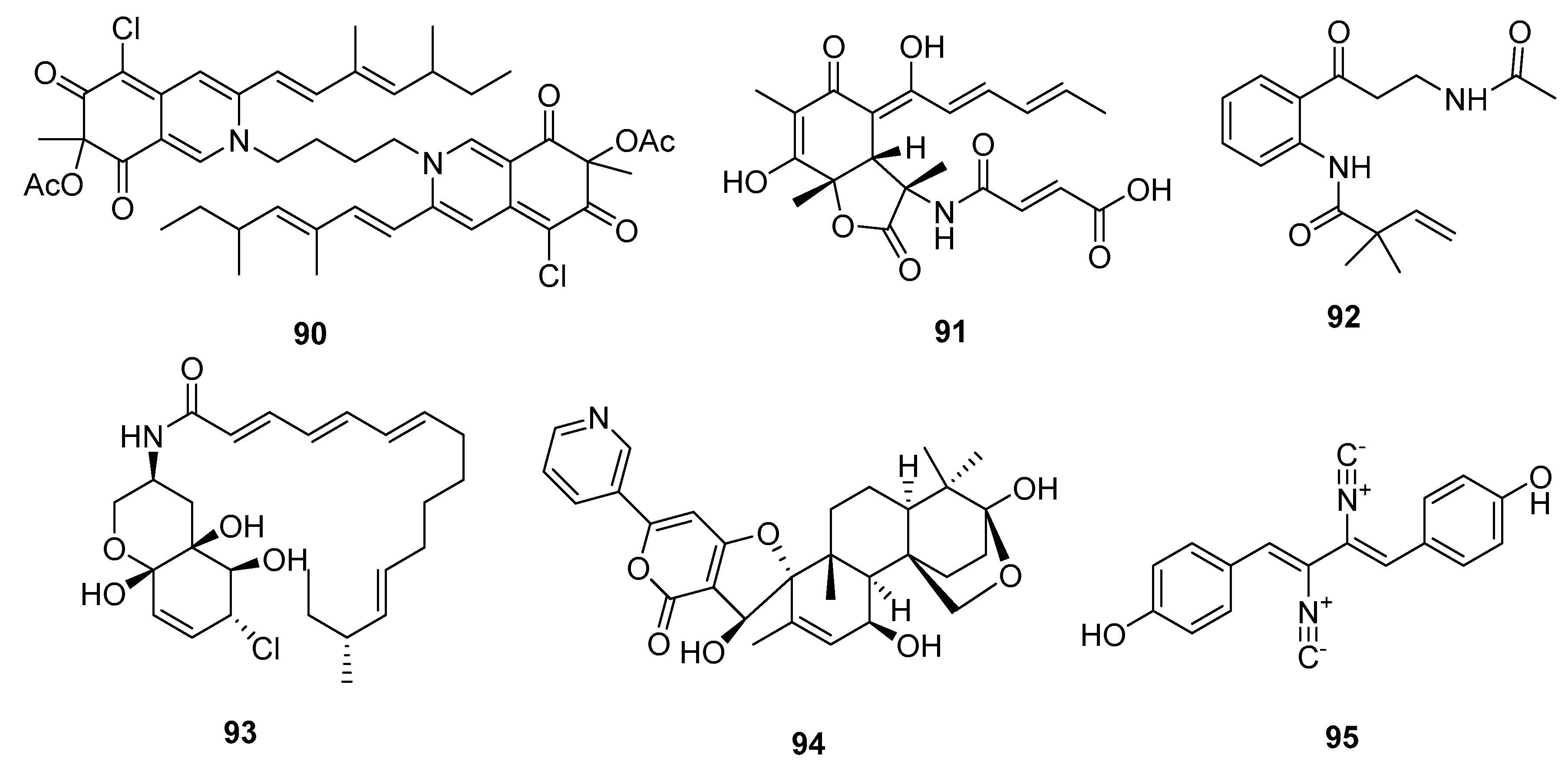
| Bis-sclerotioramin (90) | Penicillium 303# | Mangrove | Azaphilone alkaloid | MDA-MB-231 | 7.13 μM | [59] | |
| Sorbicillactone (91) | P. chrysogenum | Marine sponge | Miscellaneous Alkaloid | L5178Y | 2.2 μg/mL | Selective anti-leukemic | [60] |
| Brocaeloid B (92) | P. brocae | Mangrove | Miscellaneous Alkaloid | A. salina | 36.7 μM | [62] | |
| Varitatin (93) | Mutant P. varibile | Mangrove | Amide alkaloid | HCT-116 | 2.8 μM | IR%(1μM) = 50% and 40% (PDGFR-βand ErbB4) | [63] |
| 18-hydroxydecaturin B (94) | P. oxalicum EN-201 | Mangrove | Pyridinyl-α-pyrone alkaloid | A. salina | 2.3 μM | [42] | |
| Xantocillin X (95) | P. commune SD-118 | Deep sea sediment | Isocyanide alkaloid | MCF-7, HepG2, NCI-H460, Hela, DU145, MDA-MB-231 | (12, 7, 10, 10, 8, 8) μg/mL | Inhibit MEK/EPK pathway and activate class III PI3K/Beclin 1 pathway | [23,67] |
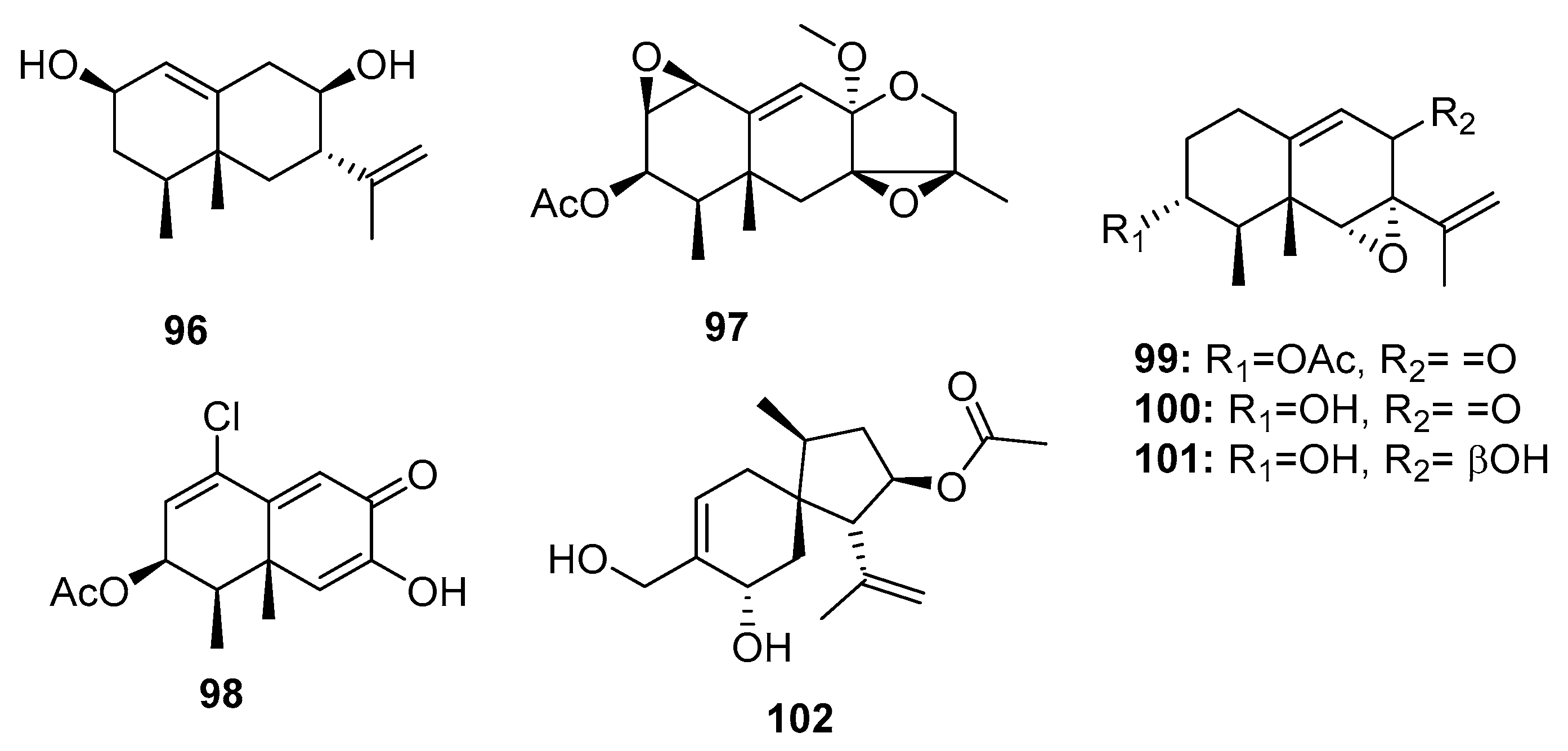
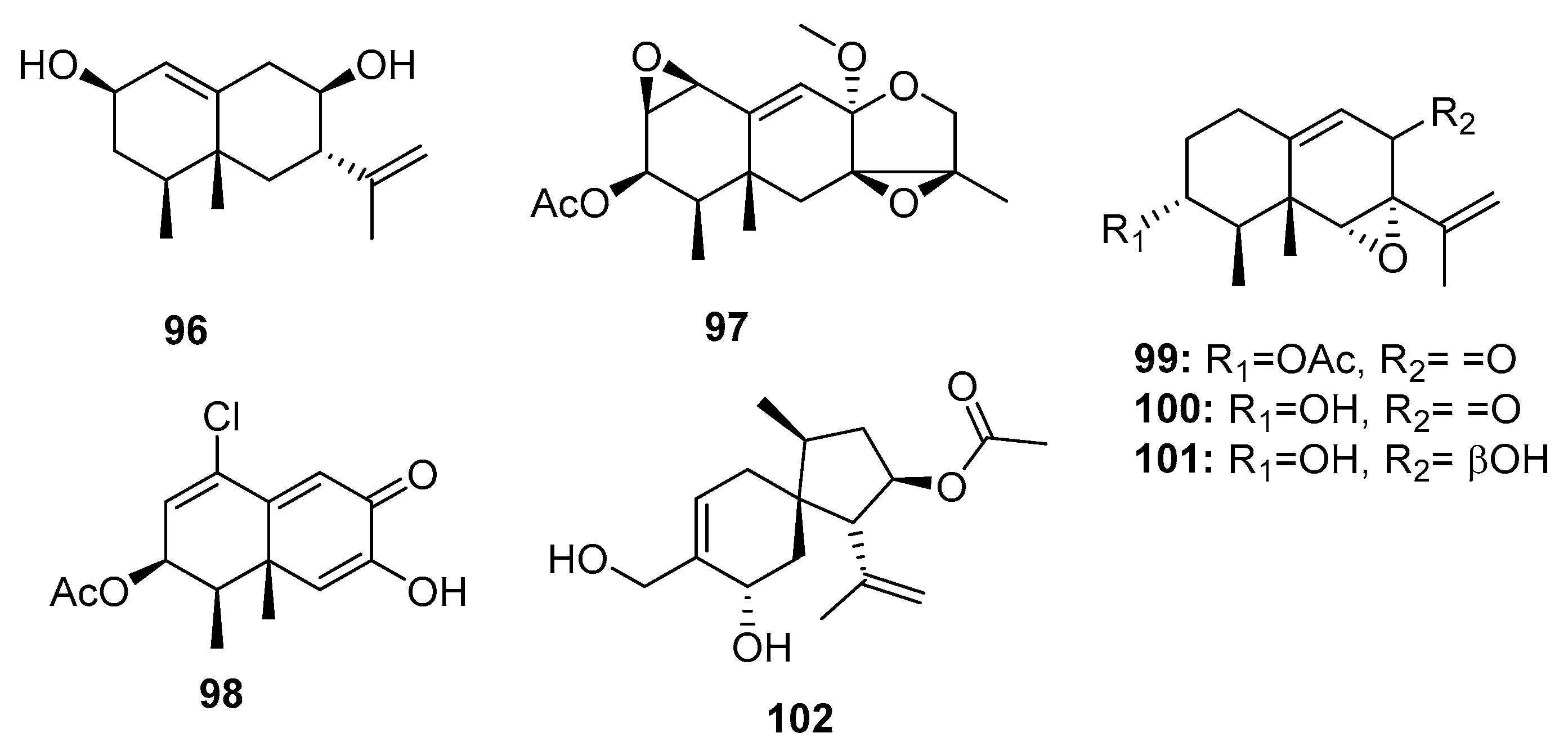
| 96 | Penicillium sp. PR19 N-1 | Marine sludge | Sesquiterpene | HL-60, A549 | (45.8, 82.8) μM | [70] | |
| 97 | Penicillium sp. PR19 N-1 | Marine sludge | Sesquiterpene | HL-60, A549 | (28.3, 5.2) μM | [70] | |
| 98 | Penicillium sp. PR19 N-1 | Marine sludge | Sesquiterpene | HL-60, A549 | (11.8, 12.2) μM | [71] | |
| 99 | Penicillium sp. BL 27-2 | Sea mud | Sesquiterpene | P388, A549, HL-60, BEL-7402 | (0.073, 0.096, 0.065, 4.59) μM | [72] | |
| Sporogen-AO 1 (100) | Penicillium sp. BL 27-2 | Sea mud | Sesquiterpene | P388, A549, HL-60, BEL-7402 | (10.1, 8.81, 10.4, 5.7) μM | [72] | |
| 101 | Penicillium sp. BL 27-2 | Sea mud | Sesquiterpene | P388, A549, HL-60, BEL-7402 | (8.71, 3.51, 7.75, 11.8) μM | [72] | |
| Adametacorenol B (102) | P. adametzioides AS-53 | Marine sponge | Diterpene | NCI-H446 | 5.0 μM | [17] | |
| Conidiogenone B (103) | Penicillium sp. | Deep sea sediment | Diterpene | A549, HL-60 | (40.3, 28.2) μM | [22] | |
| Conidiogenone C (104) | Penicillium sp. | Deep sea sediment | Diterpene | HL-60, BEL-7402 | (0.038, 0.97) μM | [22] | |
| Conidiogenone D (105) | Penicillium sp. | Deep sea sediment | Diterpene | A549, HL-60, BEL-7402, MOLT-4 | (9.3, 5.3, 11.7, 21.1) μM | [22] | |
| Conidiogenone E (106) | Penicillium sp. | Deep sea sediment | Diterpene | A549, HL-60, MOLT-4 | (15.1, 8.5, 25.8) μM | [22] | |
| Conidiogenone F (107) | Penicillium sp. | Deep sea sediment | Diterpene | A549, HL-60, BEL-7402 | (42.2,17.8, 17.1) μM | [22] | |
| Conidiogenone G (108) | Penicillium sp. | Deep sea sediment | Diterpene | A549, HL-60, BEL-7402, MOLT-4 | (8.3, 1.1, 43.2, 4.7) μM | [22] | |
| Brevione I (109) | Penicillium sp. | Deep sea sediment | Diterpene | MCF-7 | 7.44 μM | [73] | |
| Brevione A (110) | Penicillium sp. | Deep sea sediment | Diterpene | MCF-7 | 28.4 μM | [73] | |
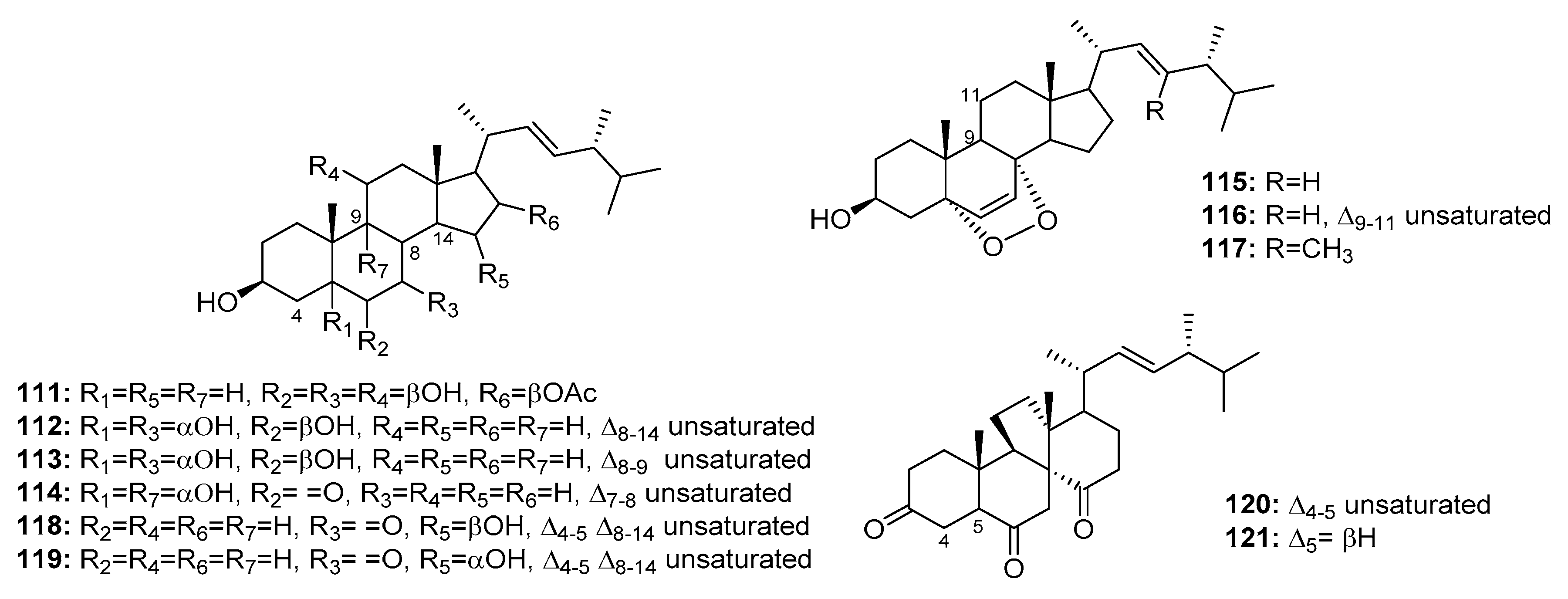
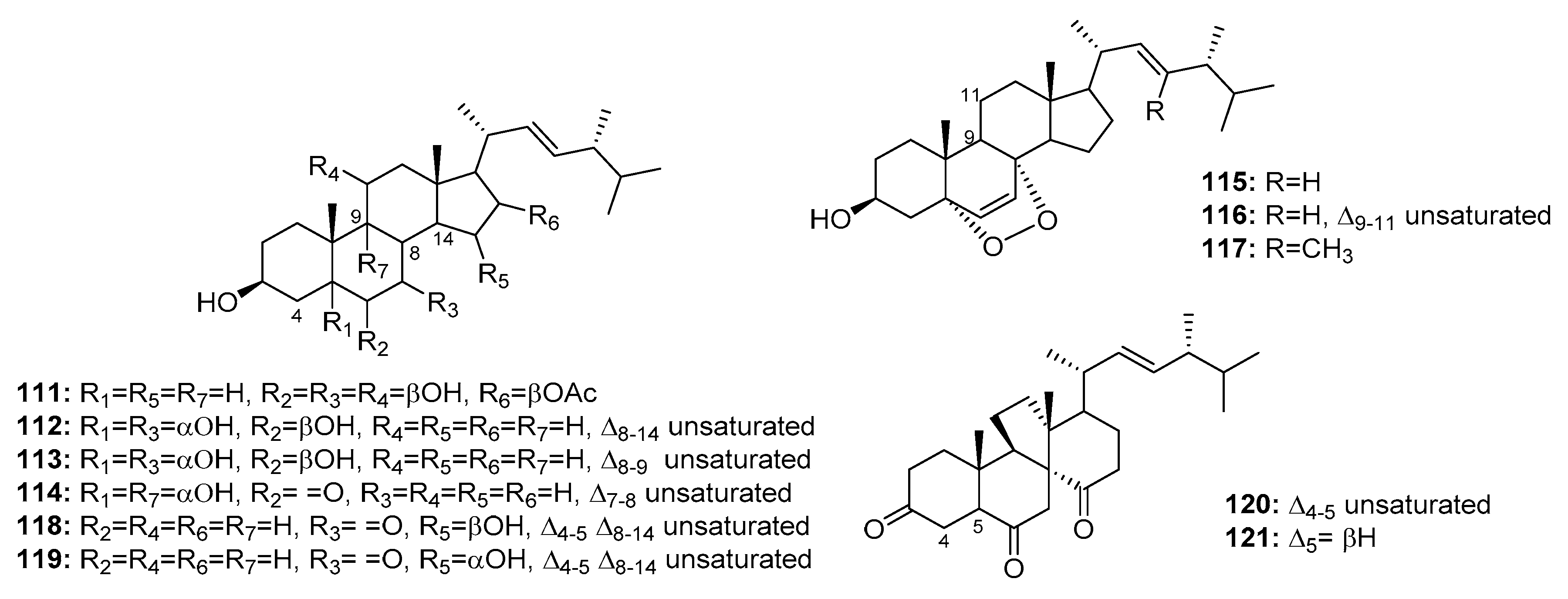
| Penicisteroid A (111) | P. chrysogenum QEN-24S | Marine alga | Steroid | Hela, SW1990, NCI-H460 | (15, 31, 40) μg/mL | [74] | |
| 112 | Penicillium sp. | Marine moss | Steroid | HepG2 | 10.4 μg/mL | [75] | |
| 113 | Penicillium sp. | Marine moss | Steroid | HepG2 | 15.6 μg/mL | [75] | |
| 114 | Penicillium sp. | Marine moss | Steroid | HepG2 | 20.7 μg/mL | [75] | |
| 115 | Penicillium sp. | Marine moss | Steroid | HepG2 | 16.8 μg/mL | [75] | |
| 116 | Penicillium sp. | Marine moss | Steroid | HepG2 | 21.3 μg/mL | [75] | |
| 117 | P. stoloniferum QY2-10 | Sea squirt | Steroid | P388 | 4.07 μM | [76] | |
| 118 | Penicillium sp. | Marine sponge | Steroid | K562 | 5.54 μM | [77] | |
| 119 | Penicillium sp. | Marine sponge | Steroid | K562 | 4.38 μM | [77] | |
| Dankasterone A (120) | Penicillium sp. | Marine sponge | Steroid | HL-60, Hela, K562 | (0.78, 4.11, 7.57) μM | [77] | |
| Dankasterone B (121) | Penicillium sp. | Marine sponge | Steroid | HL-60, Hela, K562 | (3.25, 4.74, 7.89) μM | [77] | |
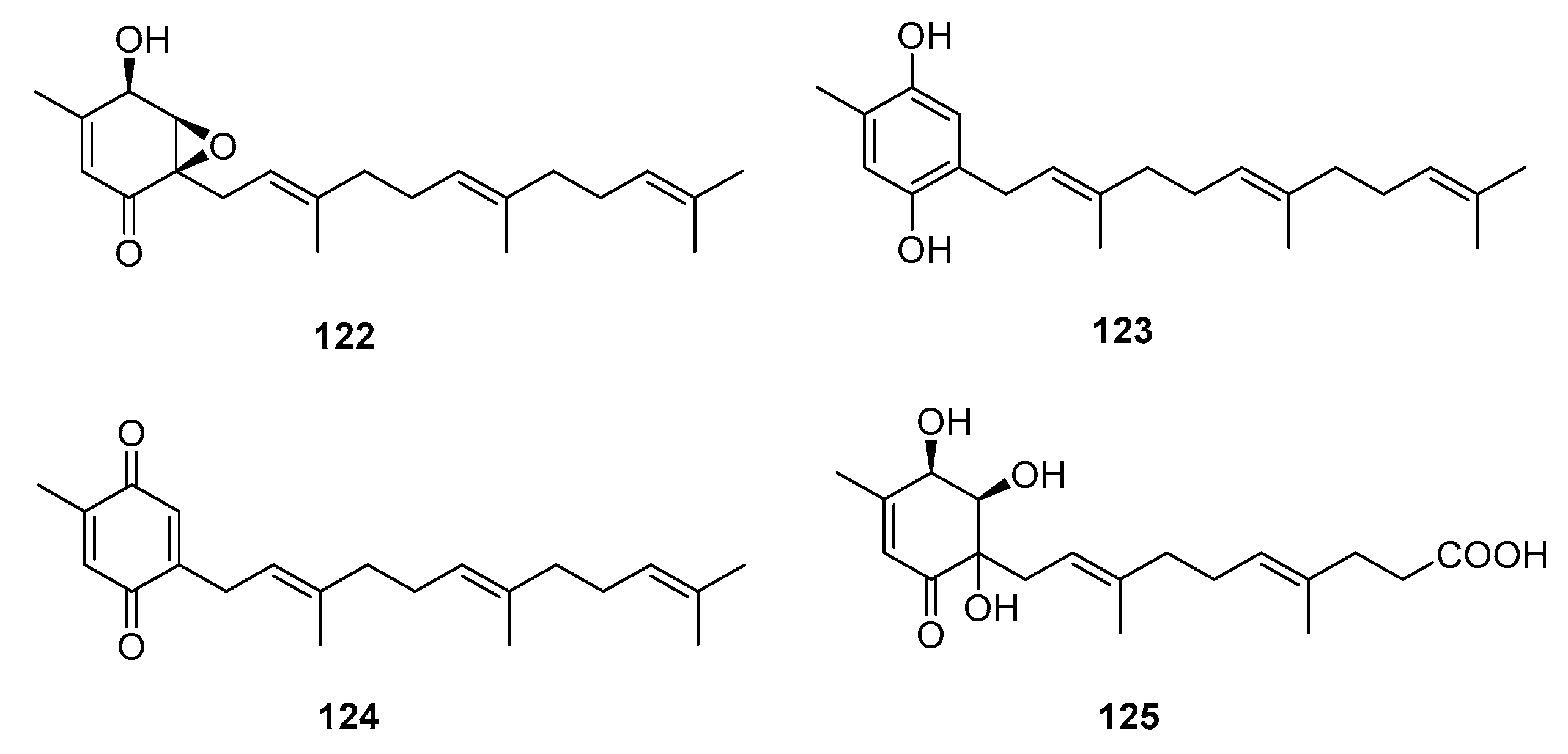
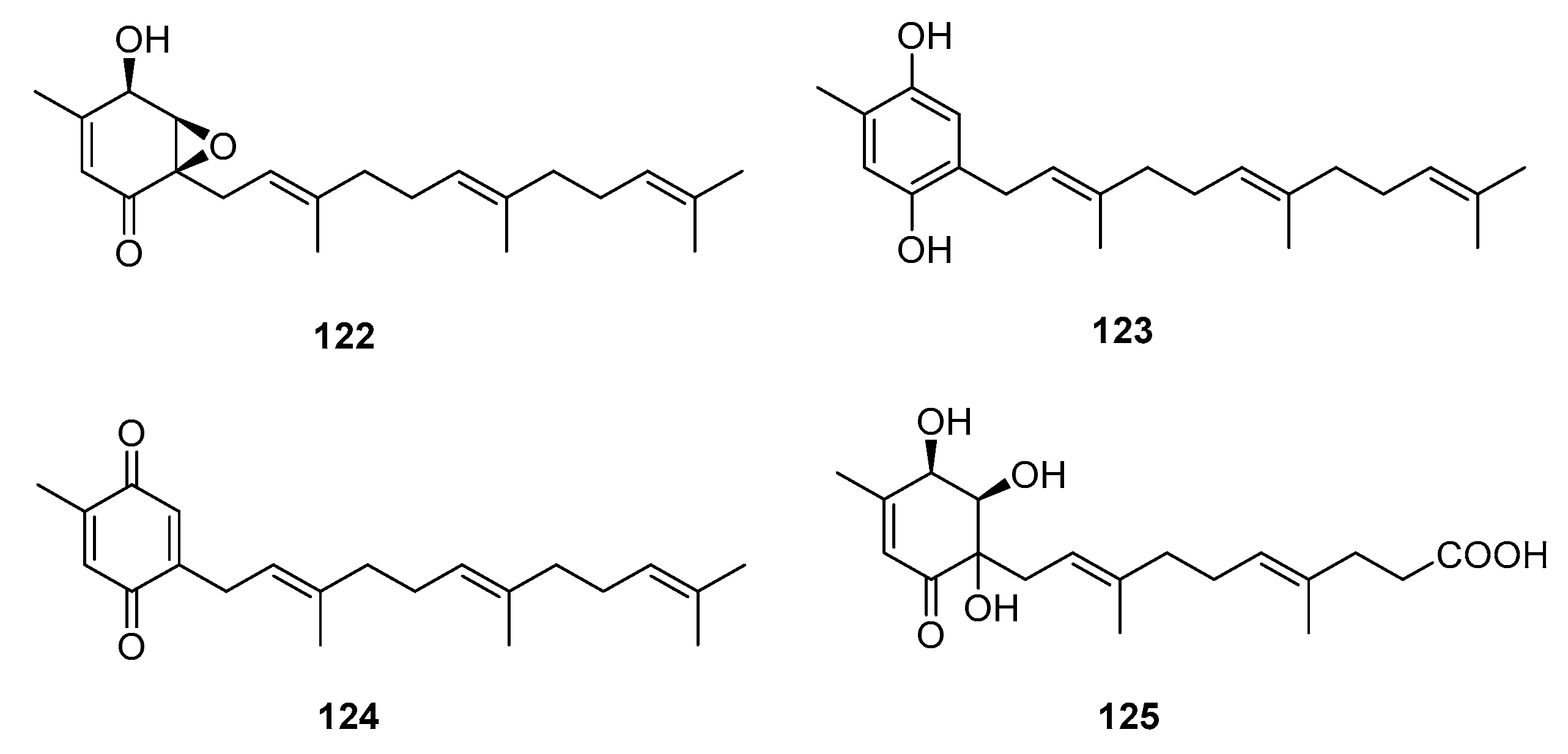
| 7-deacetoxyyanuthone (122) | Penicillium sp. | Marine (not clear) | Meroterpene | A549, SKOV-3, SKMEL-2, XF498, HCT-15 | (7.74, 6.35, 3.86, 10.04, 10.07) μg/mL | [79] | |
| Farnesylbenzenediol (123) | Penicillium sp. | Marine (not clear) | Meroterpene | A549, SKOV-3, SKMEL-2, XF498, HCT-15 | (4.73, 5.31, 4.80, 5.94, 6.11) μg/mL | [79] | |
| Farnesylquinone (124) | Penicillium sp. | Marine (not clear) | Meroterpene | A549, SKOV-3, SKMEL-2, XF498, HCT-15 | (25.44, 37.29, 18.41, 38.07, 42.56) μg/mL | [79] | |
| Penicillone A (125) | Penicillium sp. F11 | Marine (not clear) | Meroterpene | HT1080, Cne2 | (45.8, 46.2) μM | [80] | |
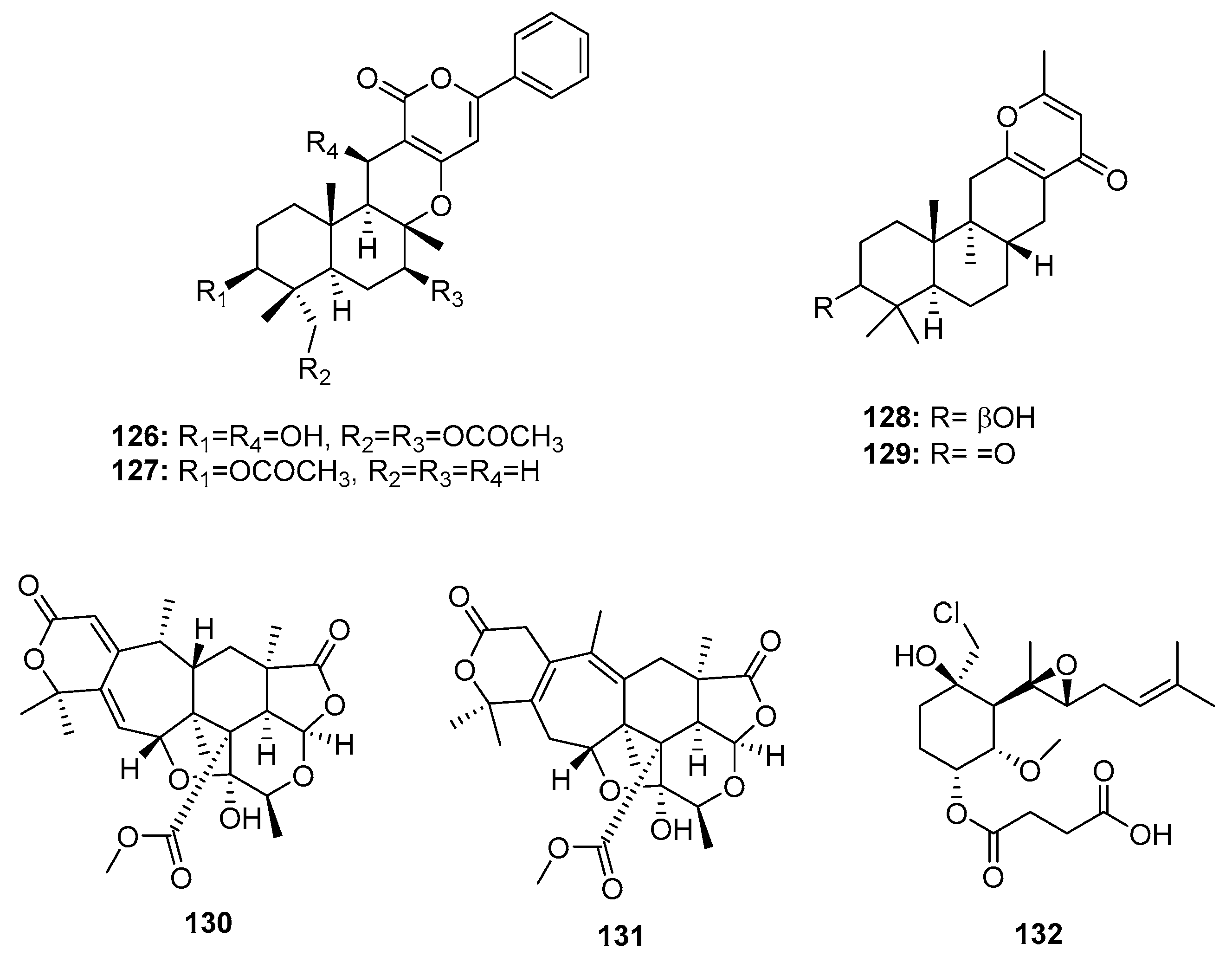
| Phenylpyropene E (126) | P. concentricum ZLQ-69 | Sea water | Sesquiterpene | MGC-803 | 19.1 μM | [81] | |
| Phenylpyropene F (127) | P. concentricum ZLQ-69 | Sea water | Sesquiterpene | MGC-803 | 13.6 μM | [81] | |
| Penicillipyrone A (128) | Penicillium sp. F446 | Marine sediment | Sesquiterpene | K562, A549 | (28, 15) μM | [82] | |
| Penicillipyrone B (129) | Penicillium sp. F446 | Marine sediment | Sesquiterpene | K562, A549 | (50, 17) μM | [82] | |
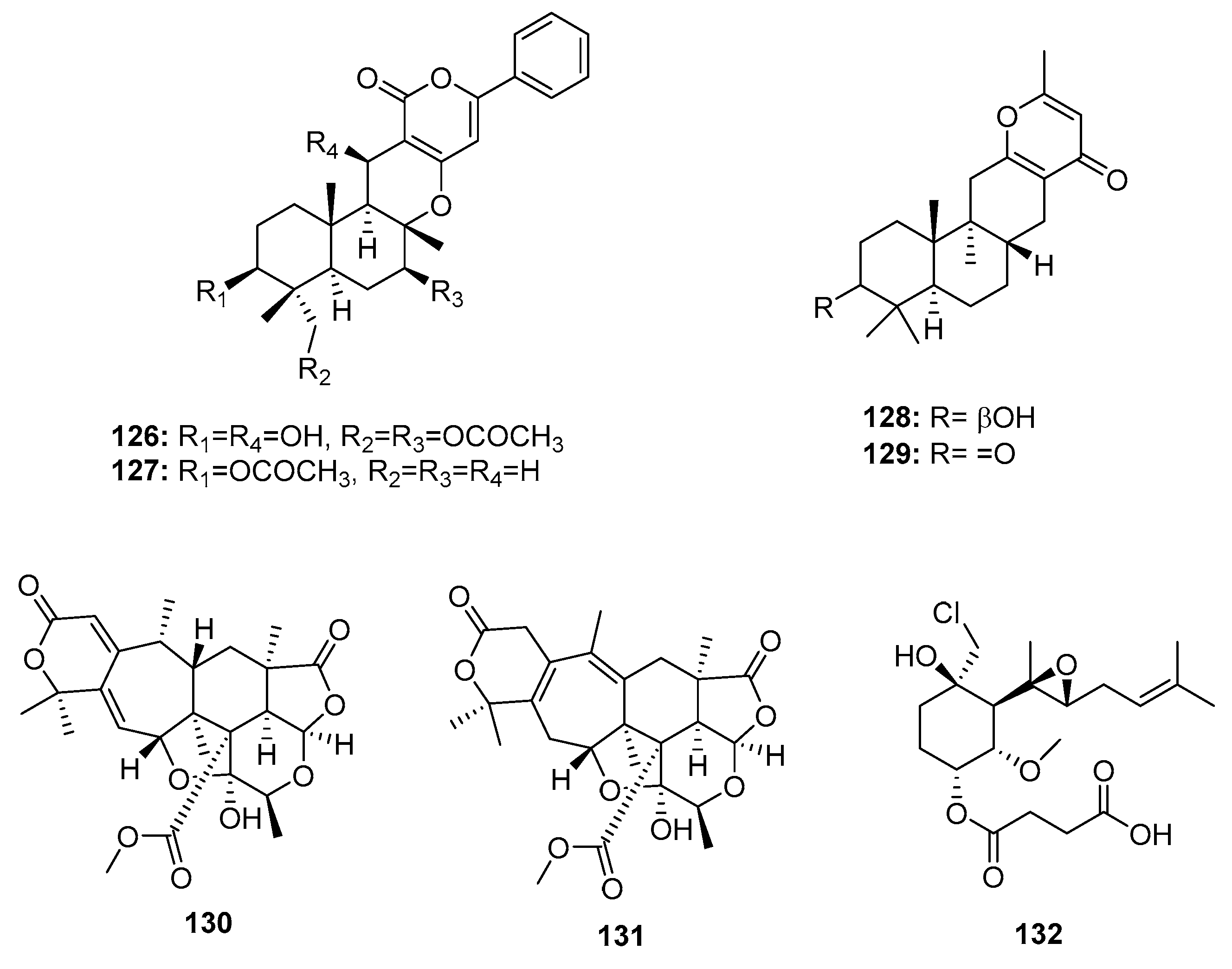
| 130 | Penicillium 303# | Mangrove | Meroterpene | MDA-MB-435, HepG2, HCT-116, A549 | (34.25, 24.56, 33.72, 37.82) μg/mL | [59] | |
| 131 | Penicillium 303# | Mangrove | Meroterpene | MDA-MB-435, HepG2, HCT-116, A549 | (31.32, 23.87, 29.19, 34.06) μg/mL | [59] | |
| Ligerin (132) | Penicillium sp. | Sea water | Merosesquiterpene | POS1, SaOS2, MG63 | (117/78, 137, 1459) nM | [84,85] | |
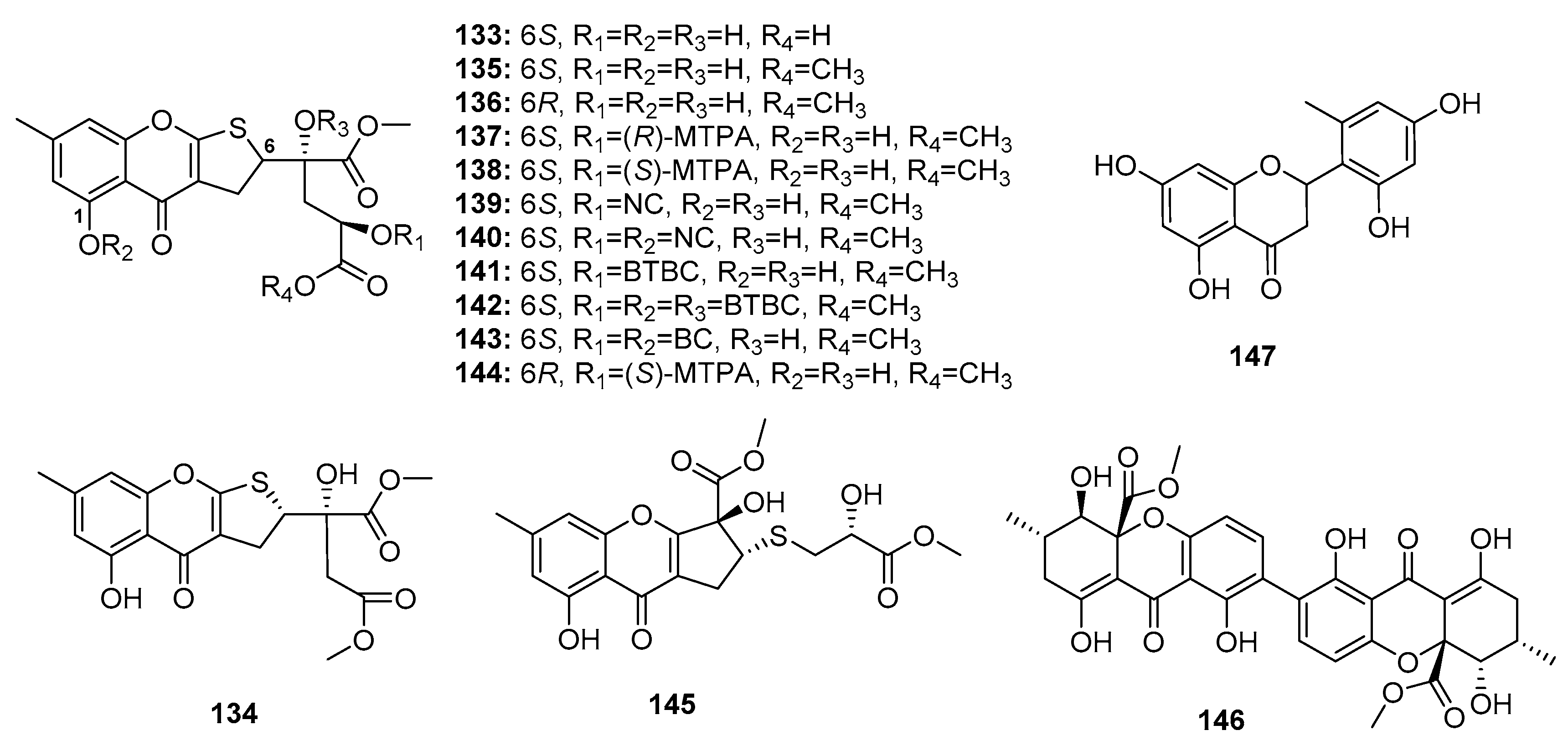
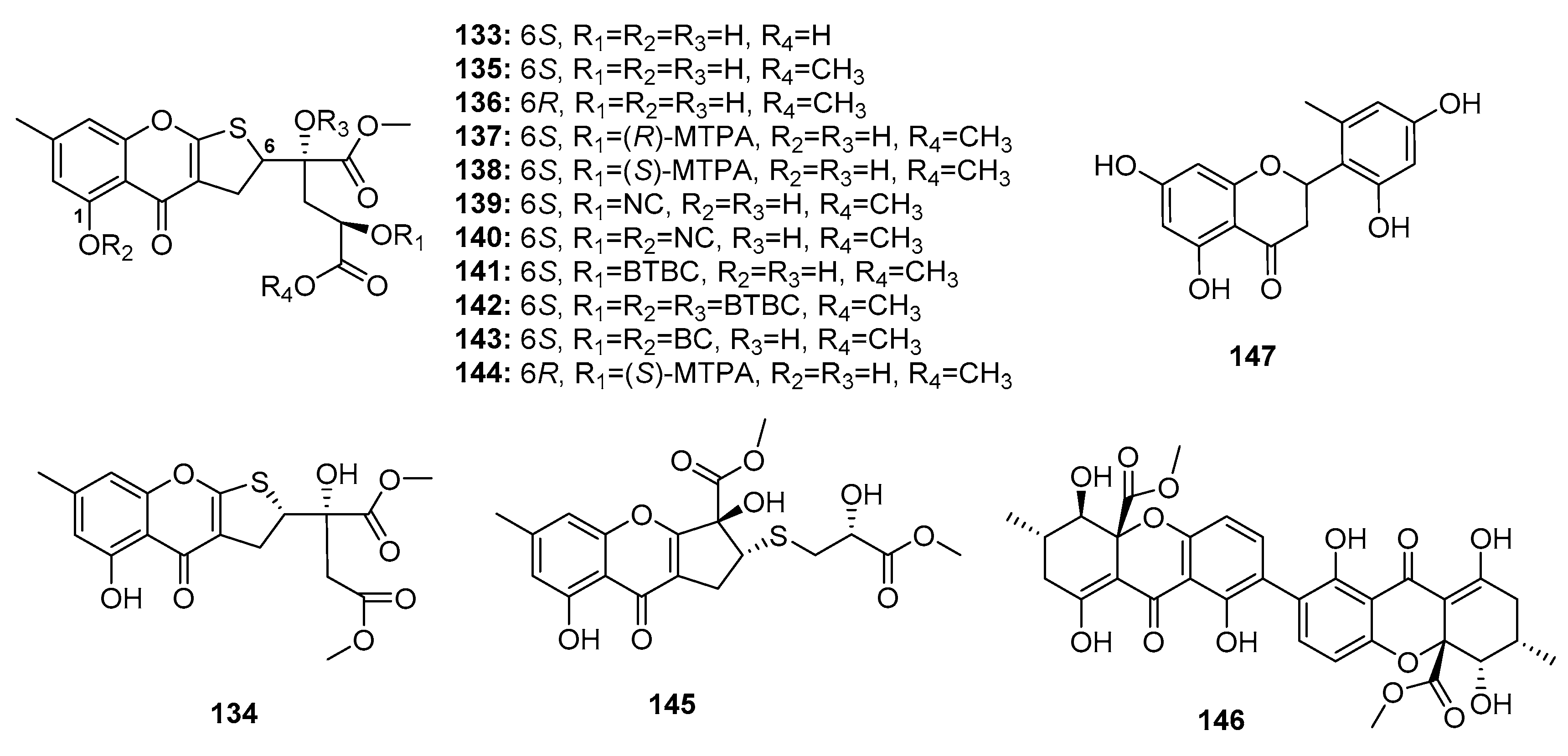
| Oxalicumone D (133) | P. oxalicum SCSGAF 0023 | Marine gorgonian | Chromone | BGC823, MOLT-4 | (10.10, 5.74) μM | [87] | |
| Oxalicumone E (134) | P. oxalicum SCSGAF 0023 | Marine gorgonian | Chromone | H1975, U937, K5652, BGC823, MOLT-4, MCF-7, HL-60, Huh-7 | (5.45, 4.16, 8.80, 1.96, 1.36, 4.32, 2.96, 6.33) μM | [87] | |
| Oxalicumone A (135) | P. oxalicum SCSGAF 0023 | Marine gorgonian | Chromone | H1975, U937, K5652, BGC823, MOLT-4, MCF-7, HL-60, Huh-7, A375, A549, Hela, HepG2, SW-620, L-02 | (10.38, 2.35, 4.53, 4.89, 0.30, 11.30, 2.55, 9.49, 11.7, 41.9, 46.2, 77.8, 22.6, 99.0) μM | [87,88] | |
| Oxalicumone B (136) | P. oxalicum SCSGAF 0023 | Marine gorgonian | Chromone | U937, MOLT-4, HL-60, A375, Hela, SW-620 | (5.00, 2.30, 6.41, 27.8, 60.9, 40.6) μM | [87,88] | |
| Chromosulfine (145) | Mutant P. purpurogenum G59 | Marine (not clear) | Chromone | K562, HL-60, BGC-823, Hela, MCF-7 | (60.8, 16.7, 73.8, 75.4, 59.2) μM | [91] | |
| Secalonic acid F (146) | Penicillium sp. | Deep sea sediment | Xanthone | HL-60 | Modulate Rho GDP dissociation inhibitor 2 Activate caspase 3 and caspase 9 | [92,93] | |
| Penimethavone A (147) | P. chrysogenum | Marine gorgonian | Flavone | Hela, rhabdomyosarcoma | (8.41, 8.18) μM | [94] | |
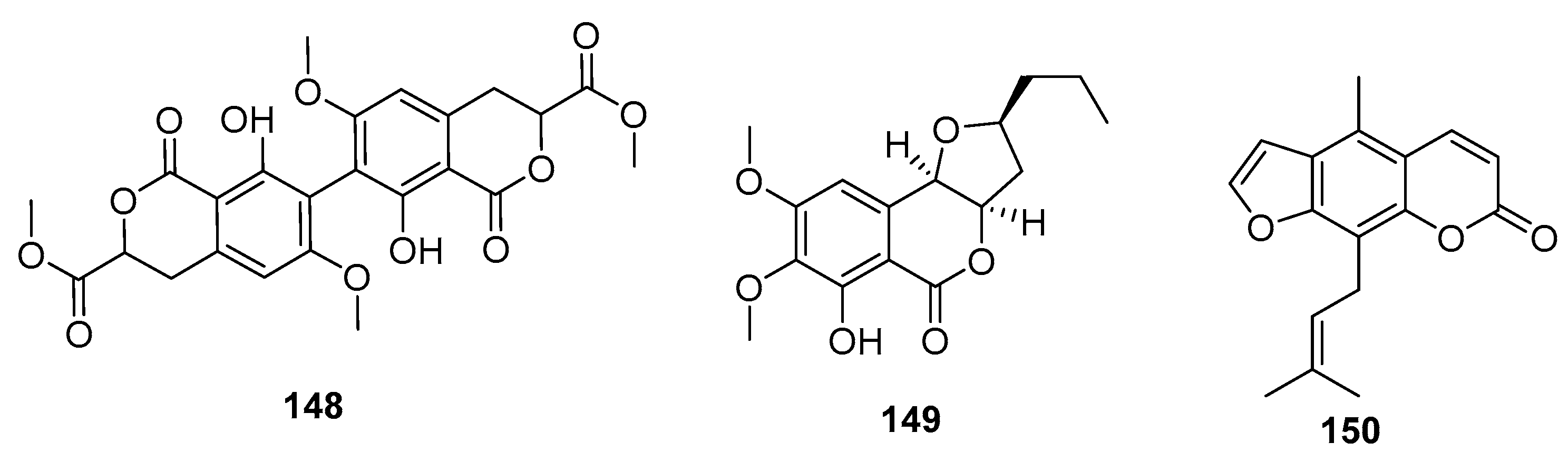
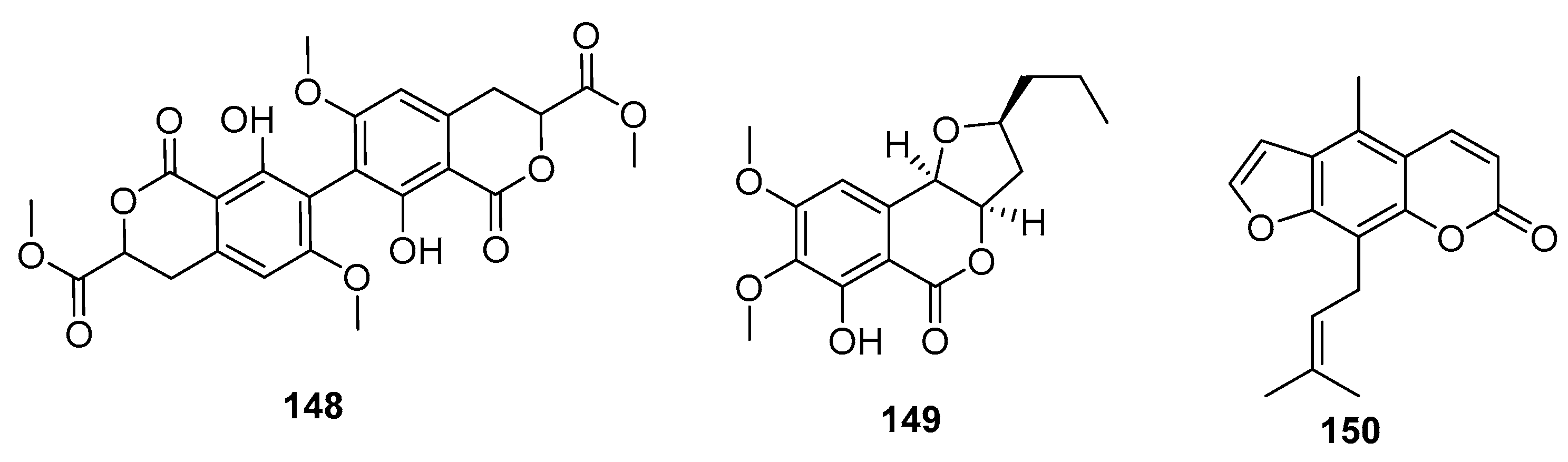
| Bipenicillisorin (148) | P. chrysogenum SCSIO 41001 | Deep sea sediment | Coumarin | K562, A549, Huh-7 | (6.78, 6.94, 2.59) μM | [95] | |
| Monocerin (149) | Penicillium sp. | Marine sponge | Coumarin | L5178Y | 8.4 μM | [96] | |
| 150 | Penicillium sp. ZH16 | Mangrove | Coumarin | KB, KBv200 | (5, 10) μg/mL | [97] | |
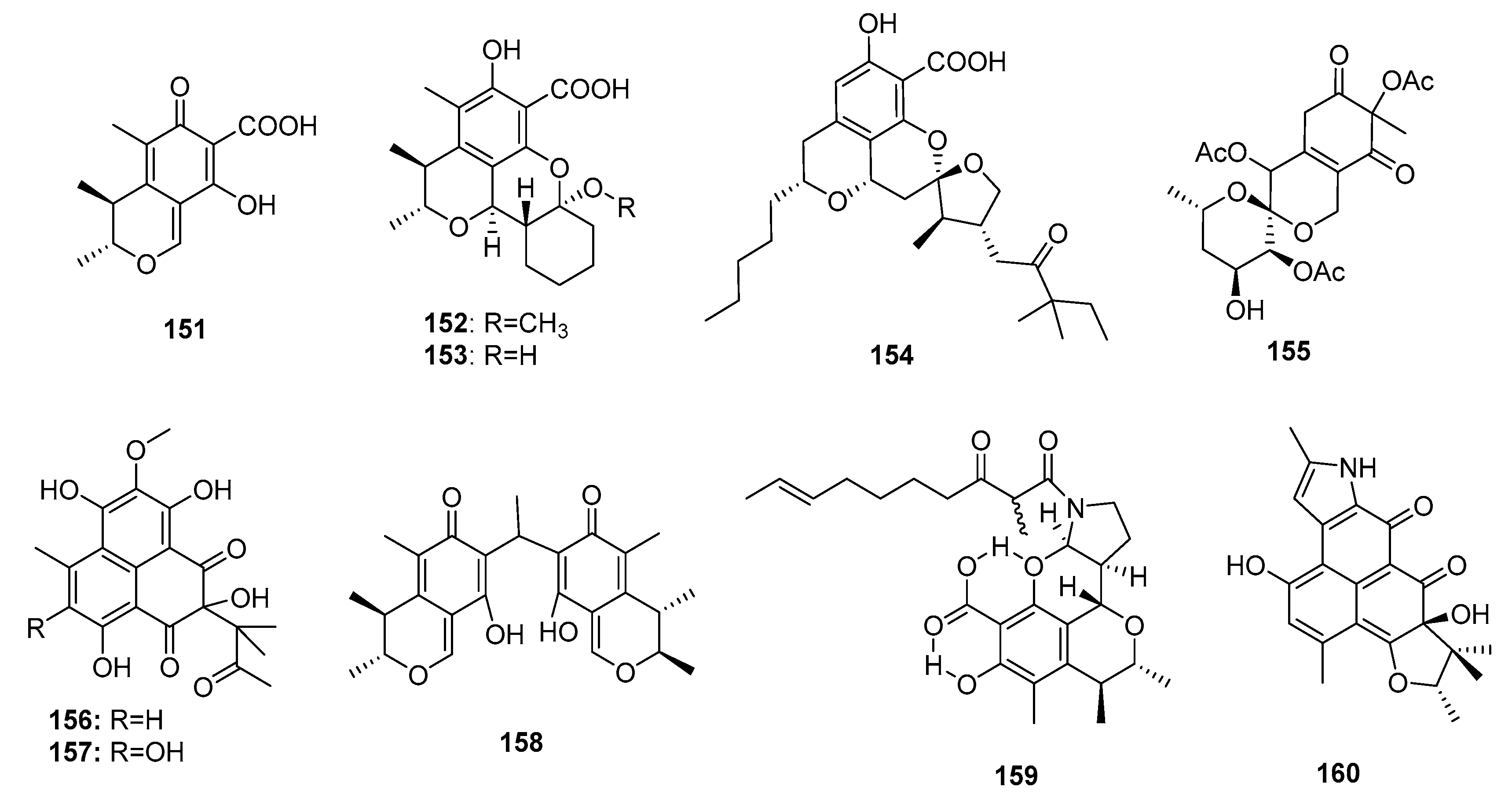
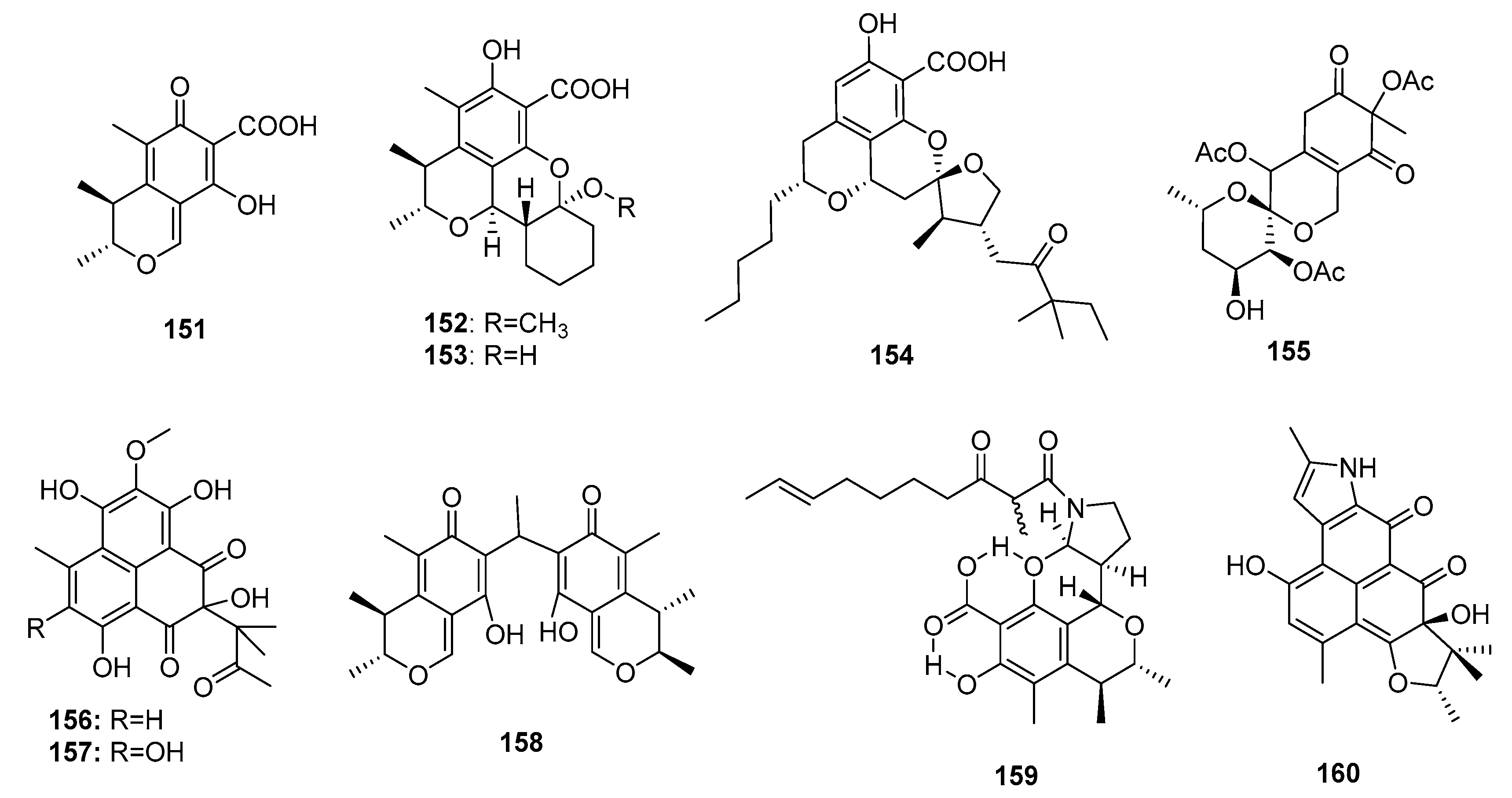
| Citrinin (151) | Penicillium sp FF001 | Marine sponge | Azaphilone polyketide | Inhibit respiration complex III | [99,101] | ||
| Penicitrinol L (152) | P. citrinum | Marine sediment | Azaphilone polyketide | SW-620 | 25.6 μM | [48] | |
| Penicitrinol M (153) | P. citrinum | Marine sediment | Azaphilone polyketide | SW-620 | 20.9 μM | [48] | |
| Berkelic acid (154) | Penicillium sp. | Acid mine lake | Azaphilone polyketide | OVCAR-3 (in NCI60) | 91 nM | Inhibit MMP-3 (GI50 = 1.87 μM) Inhibit caspase 1 (GI50 = 98 μM) | [102] |
| Sargassopenilline C (155) | P. thomii | Marine alga | Azaphilone polyketide | Inhibit the oncogenic nuclear factor AP-1 (IC50 = 15 μM) | [104] | ||
| Sculezonone A (156) | Penicillium sp. | Marine sponge | Azaphilone polyketide | Inhibit both DNA polymerases (α and β) | [105] | ||
| Sculezonone B (157) | Penicillium sp. | Marine sponge | Azaphilone polyketide | Inhibit both DNA polymerases (α and β) | [105] | ||
| Dicitrinone B (158) | P. citrinum | Marine sediment | Azaphilone polyketide | Induce apoptosis through ROS-related caspase pathway | [106] | ||
| Perinadine A (159) | P. citrinum | Marine fish | Azaphilone polyketide | L1210 | 20 μg/mL | [107] | |
| Herqueiazole (160) | Penicillium sp F011 | Marine sediment | Azaphilone polyketide | A549 | 67.3 μM | [108] | |
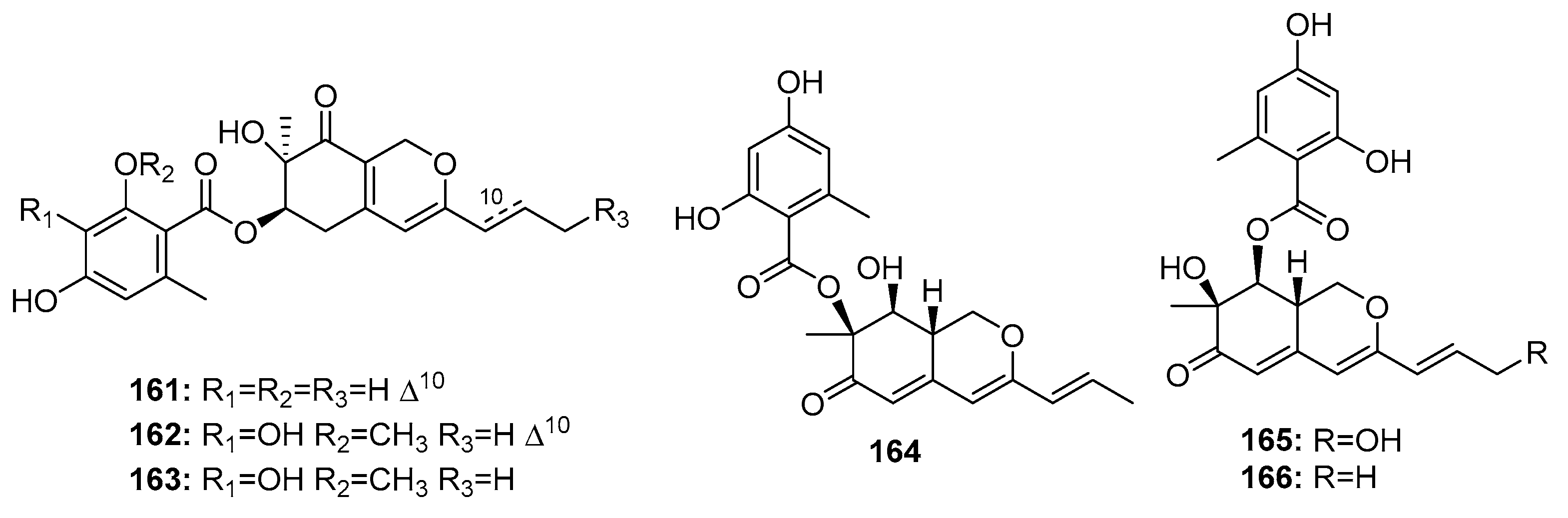
| Comazaphilone D (161) | P. commune QSD-17 | Marine sediment | Azaphilone polyketide | SW1990 | 51 μM | [109] | |
| Comazaphilone E (162) | P. commune QSD-17 | Marine sediment | Azaphilone polyketide | SW1990 | 26 μM | [109] | |
| Comazaphilone F (163) | P. commune QSD-17 | Marine sediment | Azaphilone polyketide | SW1990 | 53 μM | [109] | |
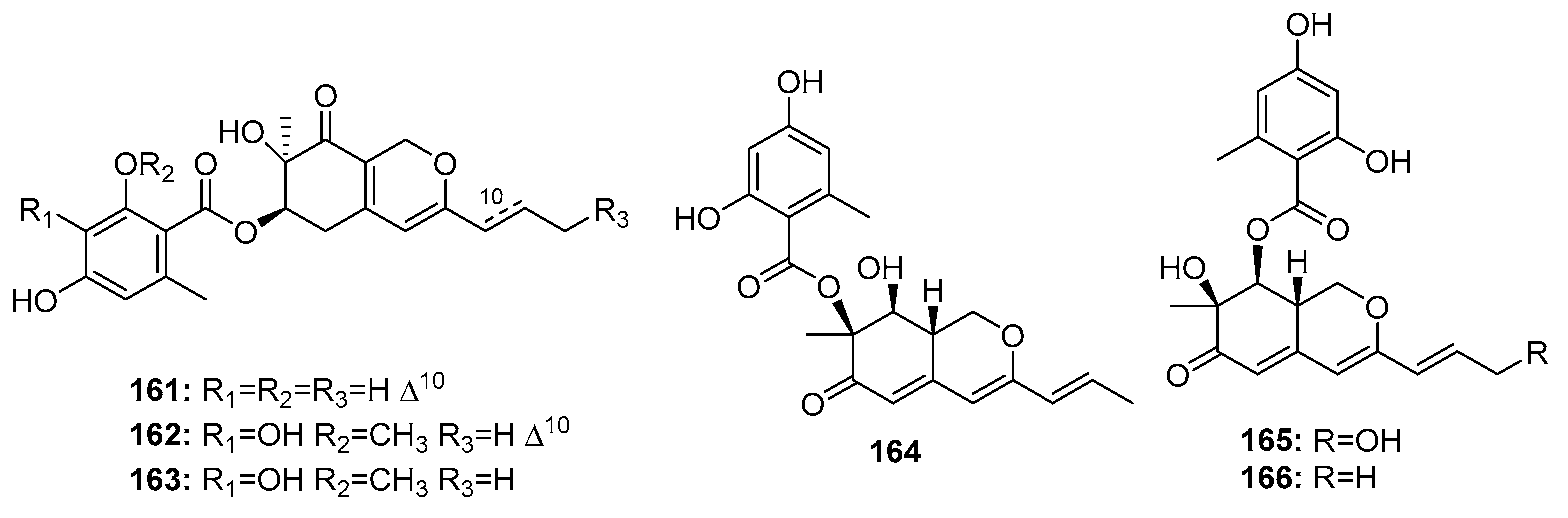
| Pinophilin A (164) | P. pinophilum Hedgcok | Marine seaweed | Azaphilone polyketide | A549, BALL-1, HCT-116, Hela, NUGC-3 | (52.5, 50.2, 51.3, 55.6, 54.7) μM | Inhibit the mammalian DNA polymerases A, B, Y family | [110] |
| Pinophilin B (165) | P. pinophilum Hedgcok | Marine seaweed | Azaphilone polyketide | A549, BALL-1, HCT-116, Hela, NUGC-3 | (93.1, 90.4, 92.5, 99.0, 96.8) μM | Inhibit the mammalian DNA polymerases A, B, Y family | [110] |
| Sch 725680 (166) | P. pinophilum Hedgcok | Marine seaweed | Azaphilone polyketide | A549, BALL-1, HCT-116, Hela, NUGC-3 | (65.7, 62.0, 64.6, 68.8, 66.4) μM | Inhibit the mammalian DNA polymerases A, B, Y family | [110] |
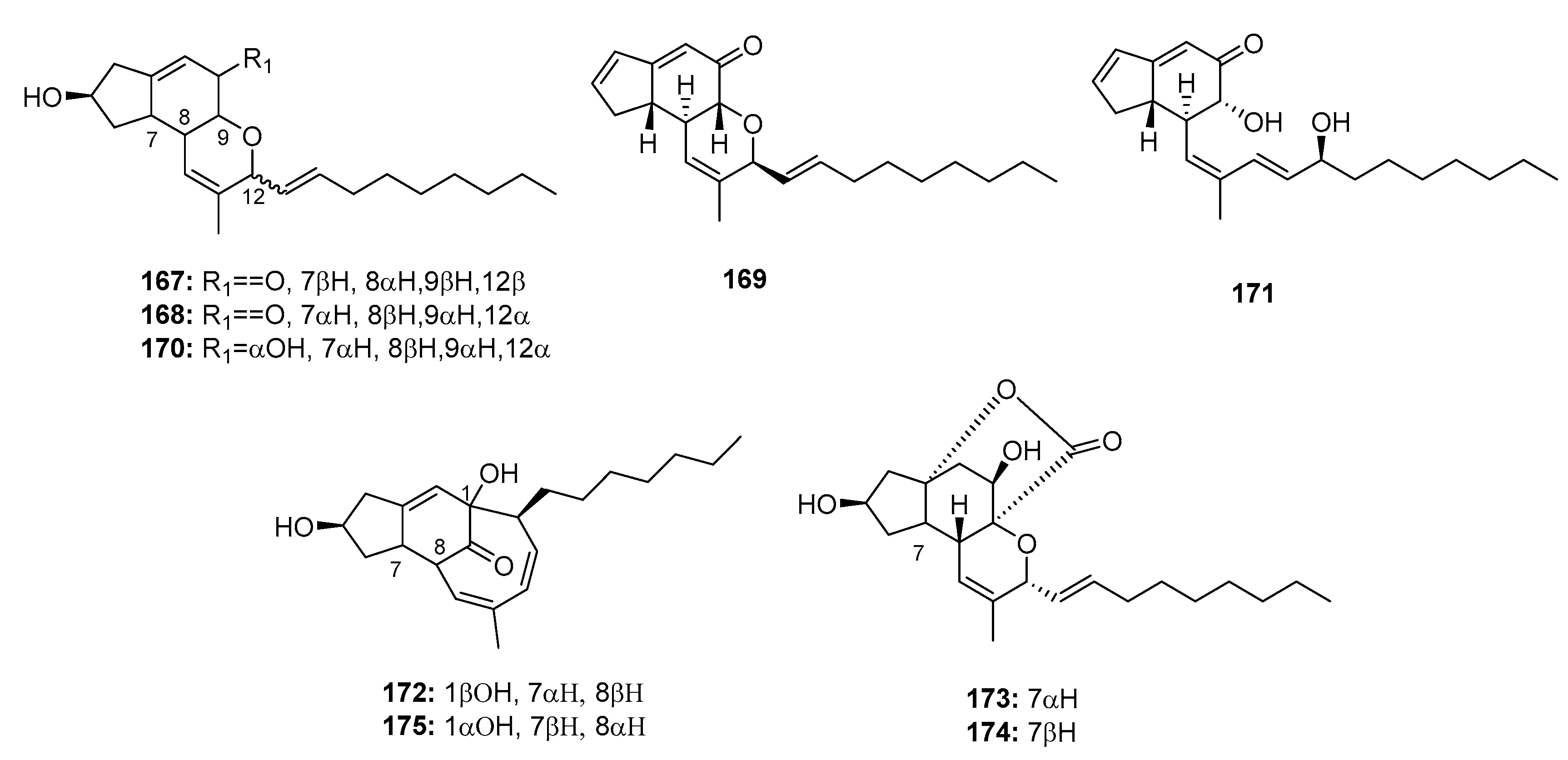
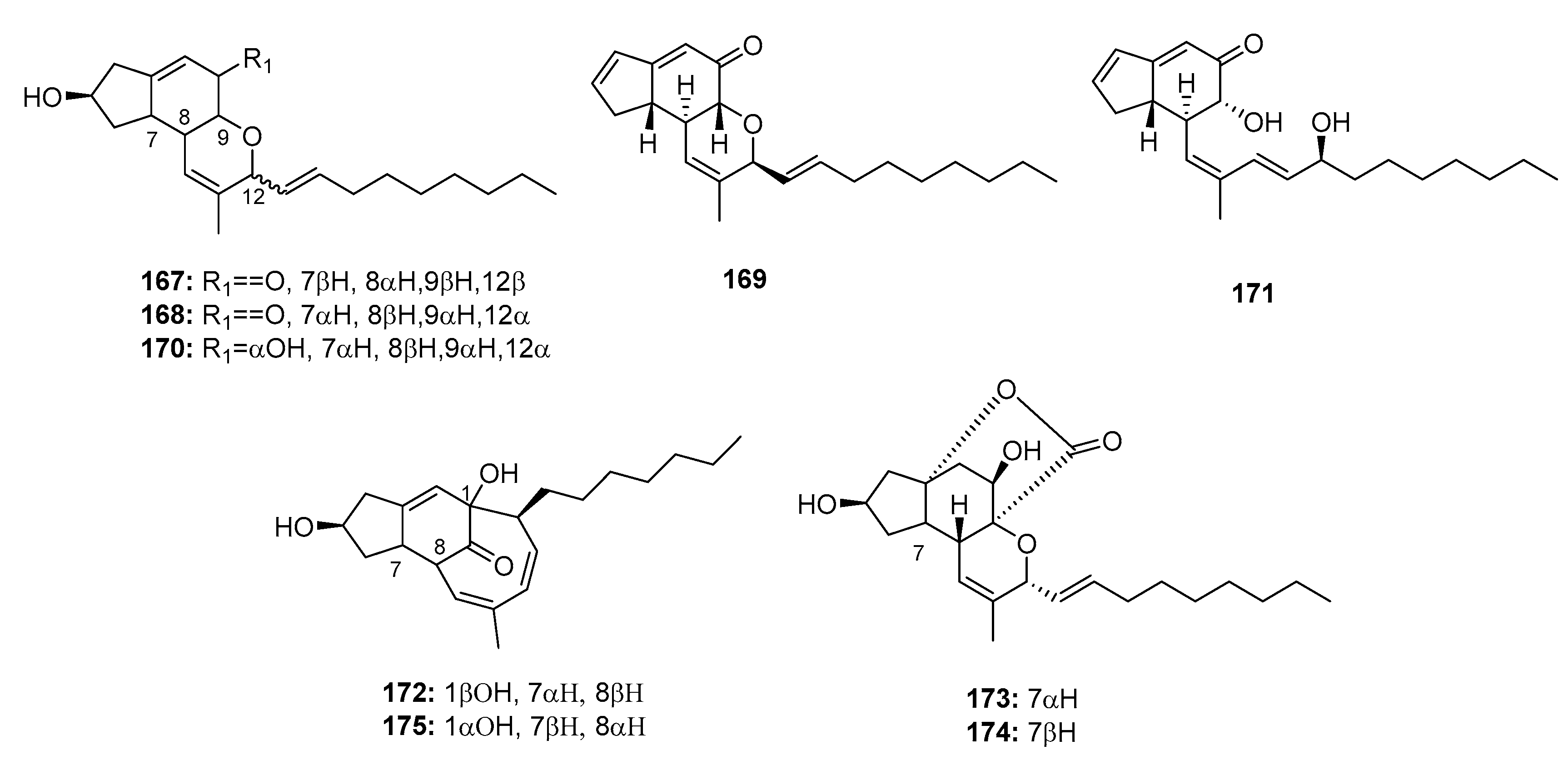
| Penostatin A (167) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388 | 0.8 μg/mL | PTP1B inhibitor (IC50 = 15.87 μM) | [111,113] |
| Penostatin B (168) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388 | 1.2 μg/mL | PTP1B inhibitor (IC50 = 33.65 μM) | [111,113] |
| Penostatin C (169) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388, BSY-1, MCF-7, HCC2998, NCI-H522, DMS114, OVCAR-3, MKN1 | (1.0, 2.0, 1.6, 2.0, 2.5, 1.9, 2.4, 1.7) μg/mL | PTP1B inhibitor (IC50 = 0.37 μM) | [111,113] |
| Penostatin D (170) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388 | 11.0 μg/mL | [111] | |
| Penostatin E (171) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388 | 0.9 μg/mL | [111] | |
| Penostatin F (172) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388 | 1.4 μg/mL | [112] | |
| Penostatin G (173) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388 | 0.5 μg/mL | [112] | |
| Penostatin H (174) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388 | 0.8 μg/mL | [112] | |
| Penostatin I (175) | Penicillium sp. OUPS-79 | Marine alga | Polyketide | P388 | 1.2 μg/mL | [112] | |
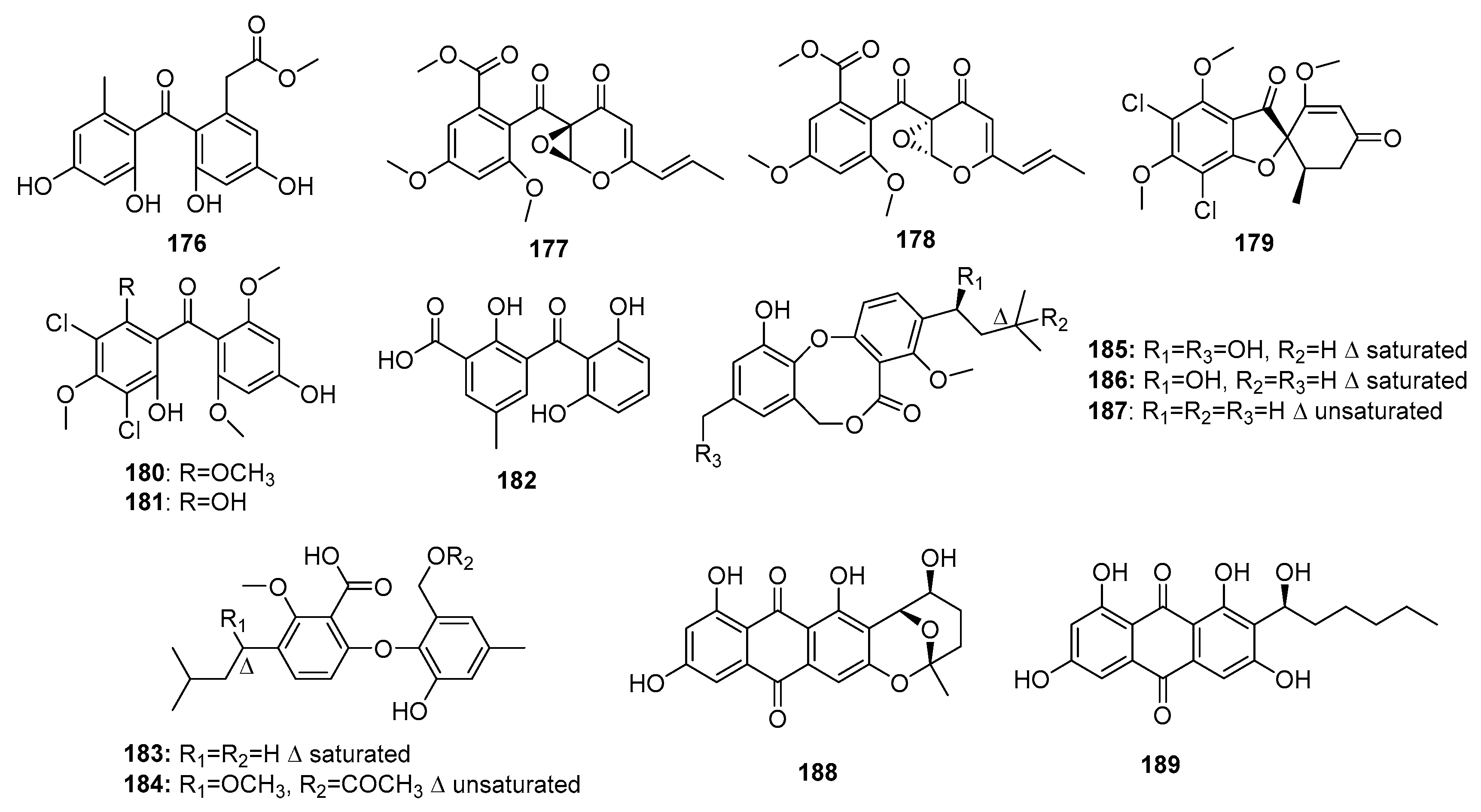
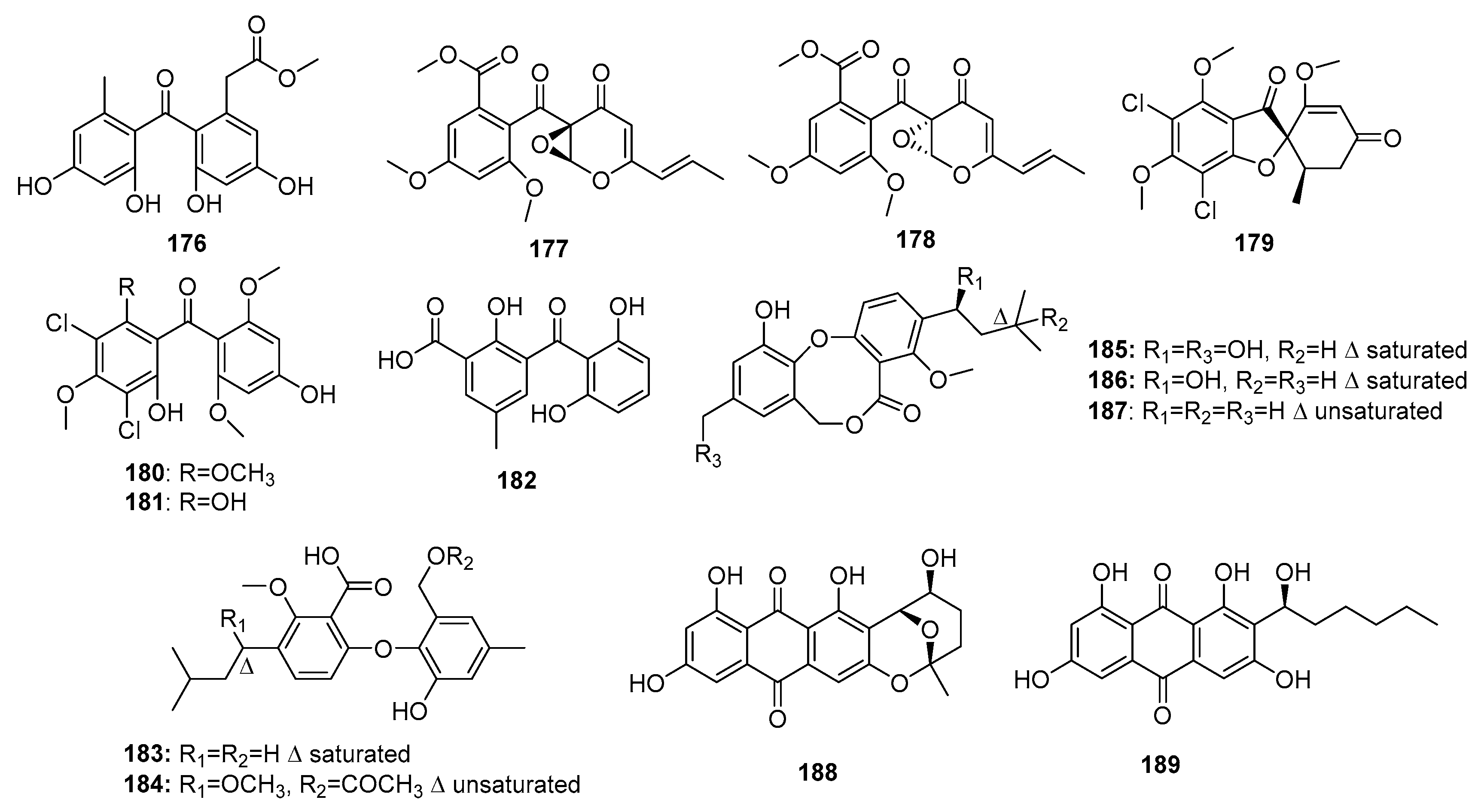
| 176 | P. oxalicum HSY05 | Marine sediment | Phenolic polyketide | DNA topoisomerase I inhibitor | [117] | ||
| 177 | Co-cultured Penicillium sp. WC-29-5 | Mangrove | Phenolic polyketide | H1975 | 3.97 μM | [118] | |
| 178 | Co-cultured Penicillium sp. WC-29-5 | Mangrove | Phenolic polyketide | H1975, HL-60 | (5.73, 3.73) μM | [118] | |
| (+)-5-chlorogriseofulvin (179) | P. canescens MMS460 | Sea water | Phenolic polyketide | KB | IR% (0.6 μM) = 49% | [119] | |
| Griseophenone I (180) | P. canescens MMS460 | Sea water | Phenolic polyketide | KB | IR% (0.6 μM) = 58% | [119] | |
| Griseophenone G (181) | P. canescens MMS460 | Sea water | Phenolic polyketide | KB | IR% (0.6 μM) = 47% | [119] | |
| Iso-monodictyphenone (182) | Penicillium sp. MA-37 | Mangrove | Phenolic polyketide | A. salina | 25.3 μM | [120] | |
| Penikellide A (183) | Penicillium sp. MA-37 | Mangrove | Phenolic polyketide | A. salina | 14.2 μM | [120] | |
| Penikellide B (184) | Penicillium sp. MA-37 | Mangrove | Phenolic polyketide | A. salina | 39.2 μM | [120] | |
| Penicillide (185) | Penicillium sp. ZLN29 | Marine sediment | Phenolic polyketide | HepG2 | (6.7/9.7, 7.8) μM | ACAT and nonpeptide calpain inhibitor | [121,122,124,125] |
| Prepenicillide (186) | Penicillium sp. ZLN29 | Marine sediment | Phenolic polyketide | HepG2, RD | 9.9 μM | [124] | |
| Hydroxypenicillide (187) | P. pinophilum | Marine gorgonian | Phenolic polyketide | Hela | 6.1 μM | [125] | |
| Nidurufin (188) | P. flavidorsum SHK1-27 | Marine sediment | Anthraquinone | K562 | 12.6 μM | Induce cell cycle arrest at G2/M transition | [126] |
| Averantin (189) | P. flavidorsum SHK1-27 | Marine sediment | Anthraquinone | K562 | 12.6 μM | [126] | |
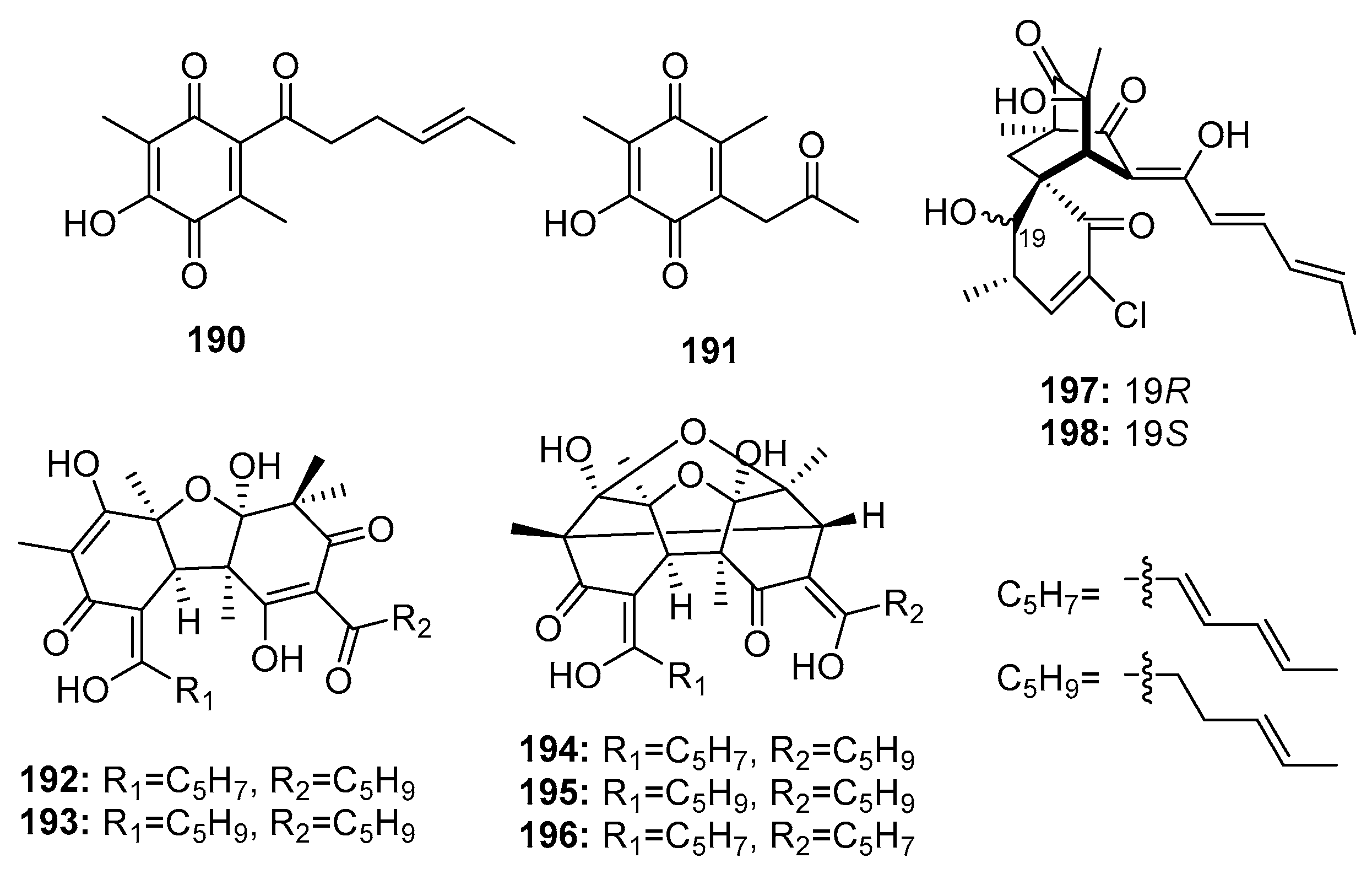
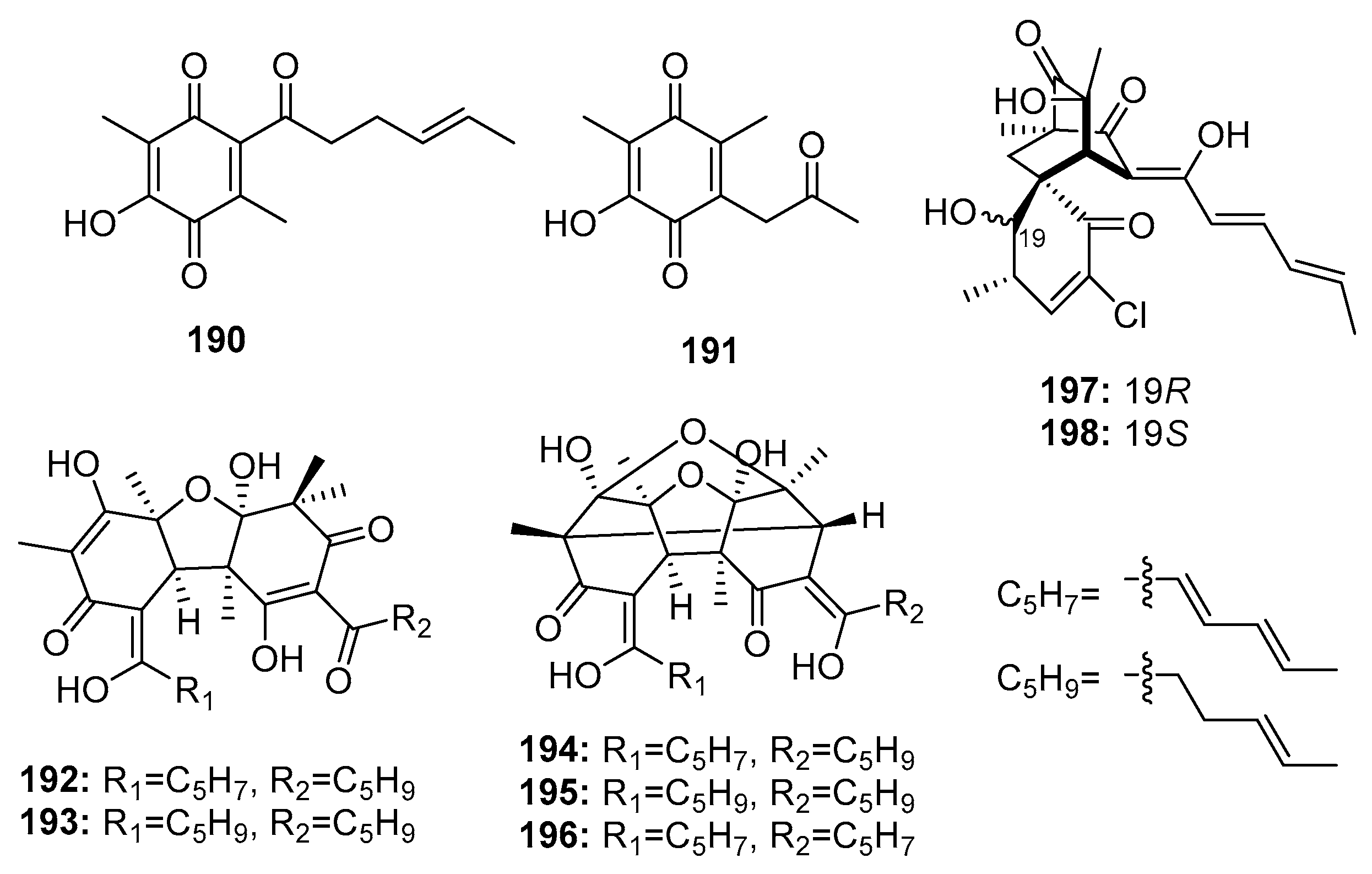
| 190 | P. terrestre | Marine sediment | Polyketide | A549, P388 | (5.3, 15.7) μM | [128] | |
| 191 | P. terrestre | Marine sediment | Polyketide | A549, P388 | (7.6, 10.5) μM | [128] | |
| Dihydrobisvertinolone (192) | P. terrestre | Marine sediment | Polyketide | A549, P388 | (0.52, 1.7) μM | [128] | |
| 193 | P. terrestre | Marine sediment | Polyketide | A549 | 1.4 μM | [128] | |
| 194 | P. terrestre | Marine sediment | Polyketide | A549, P388 | (2.1, 2.8) μM | [129] | |
| 195 | P. terrestre | Marine sediment | Polyketide | A549, P388 | (4.3, 8.8) μM | [129] | |
| Trichodimerol (196) | P. terrestre | Marine sediment | Polyketide | A549, P388 | (4.7, 0.33) μM | [129] | |
| Chloctanspirone A (197) | P. terrestre | Marine sediment | Polyketide | HL-60, A549 | (9.2, 39.7) μM | [131] | |
| Chloctanspirone B (198) | P. terrestre | Marine sediment | Polyketide | HL-60 | 37.8 μM | [131] | |
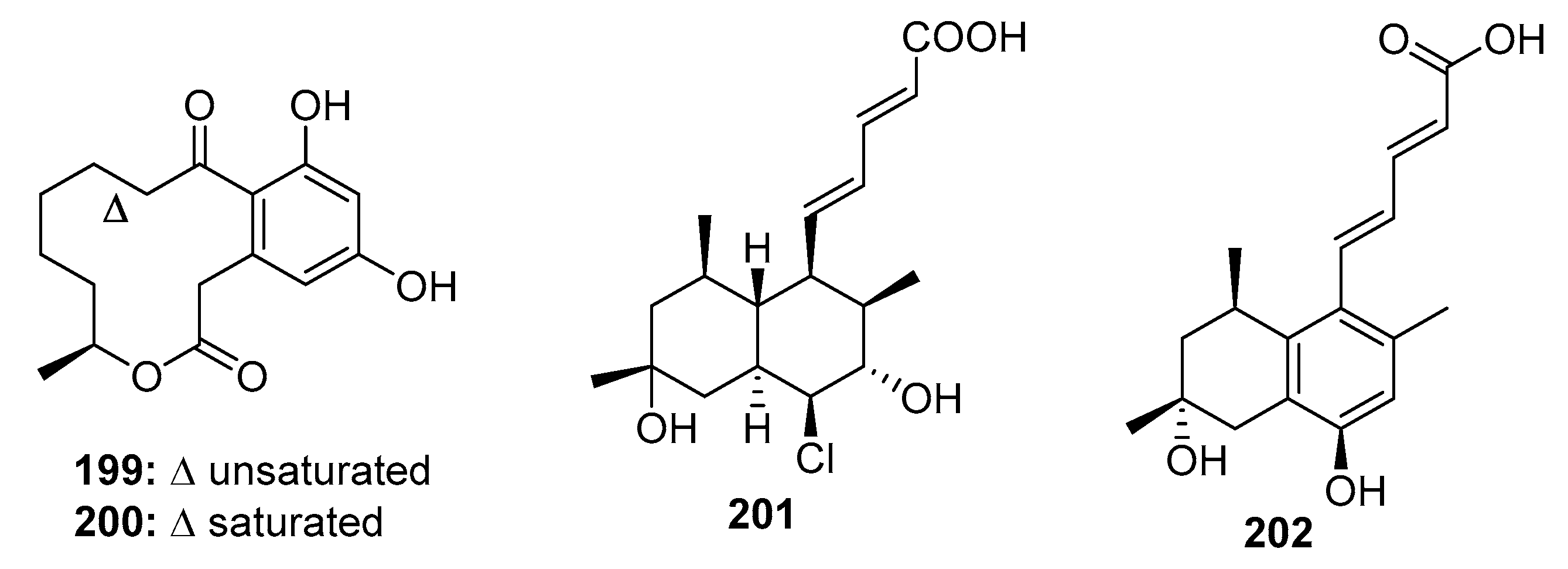
| (10E,15S)-10,11-Dehydrocurvularin (199) | Penicillium sp. DRF2 | Marine sponge | Macrolide | 36 tumor cell lines | 0.28~6 μM | [133,134] | |
| Curvularin (200) | Penicillium sp. DRF2 | Marine sponge | Macrolide | HSP90 inhibitor | [132] | ||
| Tanzawaic acid P (201) | Penicillium sp. CF07370 | Marine sediment | Polyketide | Jurkat, K562, Raji | (28.6, 30.2, 20.3) μM | Active the mitochondrial apoptotic pathway | [137] |
| Tanzawaic acid D (202) | P. steckii | Marine (not clear) | Polyketide | Bind to the FOXO1 which regulates EFGR signaling and stabilizes the FOXO1-DNA conformation | [138] | ||
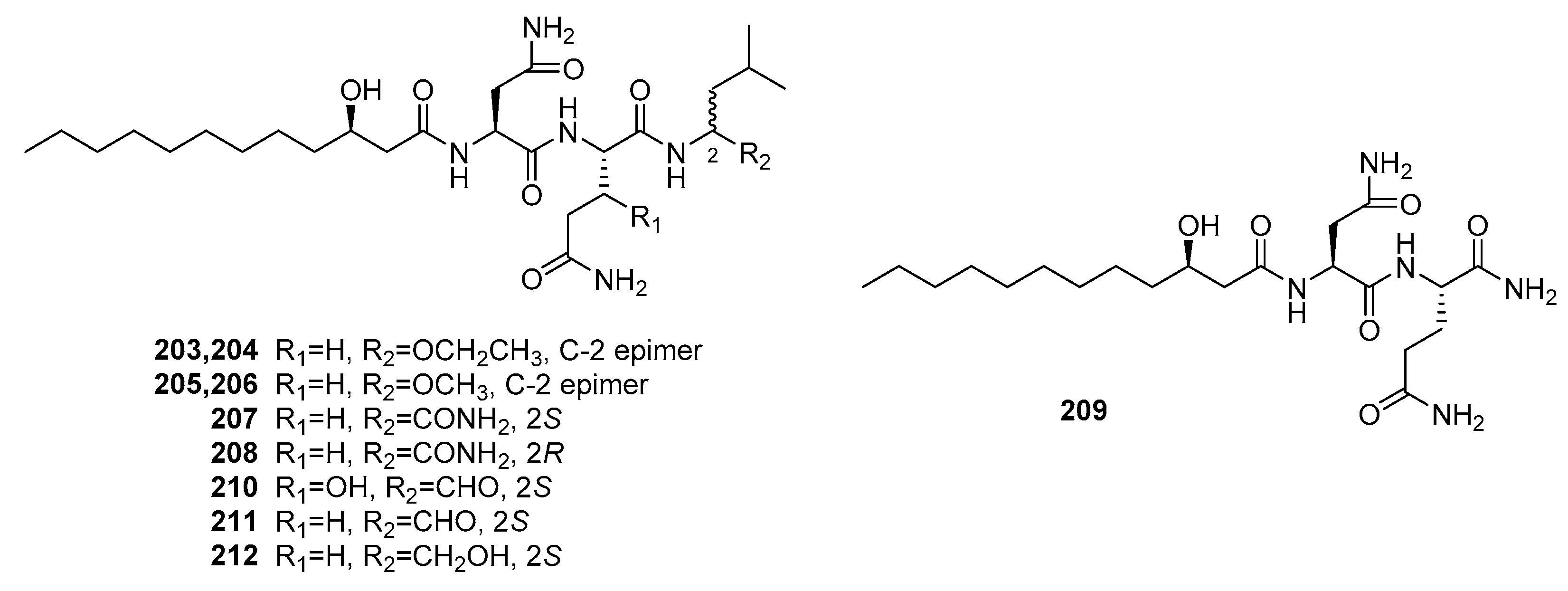
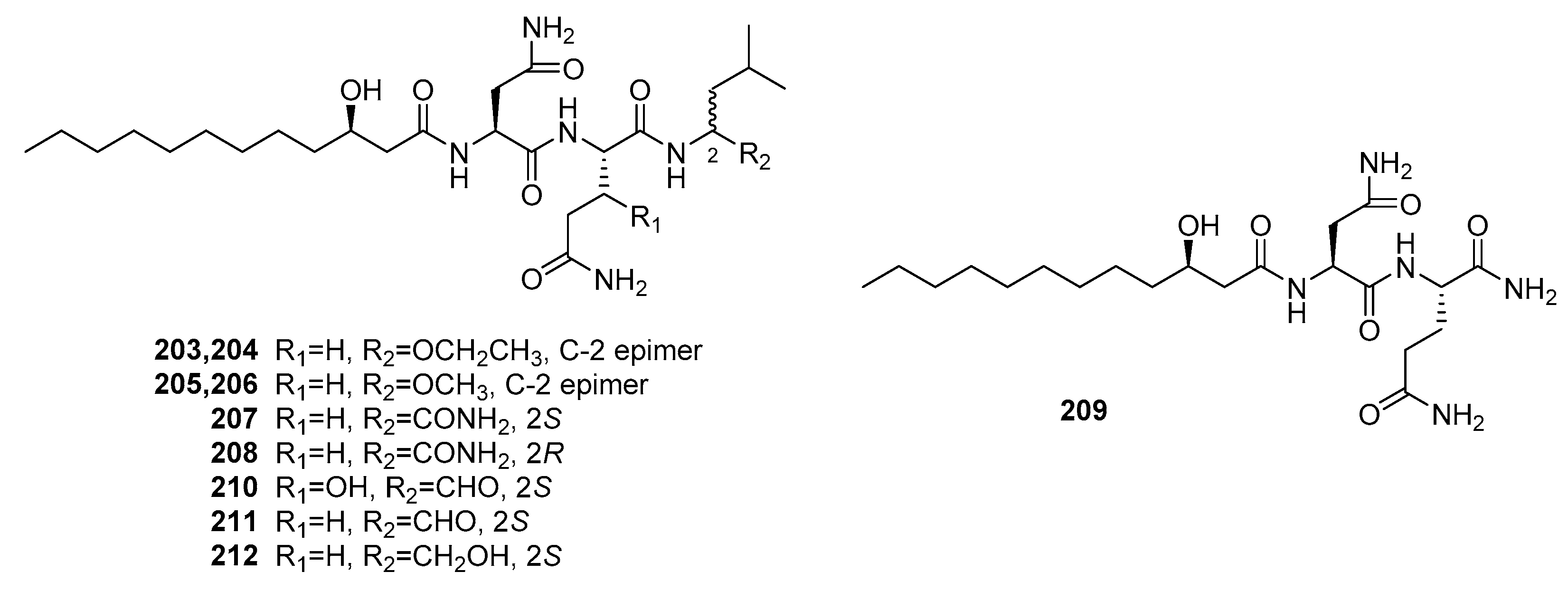
| Penicimutamide A (203) | Mutant P. purpurogenum G59 | Marine soil | Lipopepetide | K562, HL-60, Hela, BGC-823, MCF-7 | IR% (100 μg/mL) = 10~40% | [140] | |
| Penicimutamide B (204) | Mutant P. purpurogenum G59 | Marine soil | Lipopepetide | K562, HL-60, Hela, BGC-823, MCF-7 | IR% (100 μg/mL) = 25~40% | [140] | |
| Penicimutamide C (205) | Mutant P. purpurogenum G59 | Marine soil | Lipopepetide | K562, HL-60, Hela, BGC-823, MCF-7 | IR% (100 μg/mL) = 10~40% | [140] | |
| Penicimutamide D (206) | Mutant P. purpurogenum G59 | Marine soil | Lipopepetide | K562, HL-60, Hela, BGC-823, MCF-7 | IR% (100 μg/mL) = 20~40% | [140] | |
| Penicimutamide E (207) | Mutant P. purpurogenum G59 | Marine soil | Lipopepetide | K562, HL-60, Hela, BGC-823, MCF-7 | IR% (100 μg/mL) = 10~45% | [140] | |
| Penicimutamide F (208) | Mutant P. purpurogenum G59 | Marine soil | Lipopepetide | K562, HL-60, Hela, BGC-823, MCF-7 | IR% (100 μg/mL) = 10~50% | [140] | |
| Penicimutamide G (209) | Mutant P. purpurogenum G59 | Marine soil | Lipopepetide | K562, HL-60, Hela, BGC-823, MCF-7 | IR% (100 μg/mL) = 10~20% | [140] | |
| Fellutamide A (210) | P. fellutanum | Marine fish | Lipopepetide | P388, L1210 | (0.2, 0.8) μg/mL | [139] | |
| Fellutamide B (211) | P. fellutanum | Marine fish | Lipopepetide | P388, L1210 | (0.1, 0.7) μg/mL | [139] | |
| Fellutamide C (212) | Mutant P. purpurogenum G59 | Marine soil | Lipopepetide | K562, HL-60, Hela, BGC-823, MCF-7 | IR% (100 μg/mL) = 30~50% | [140] | |
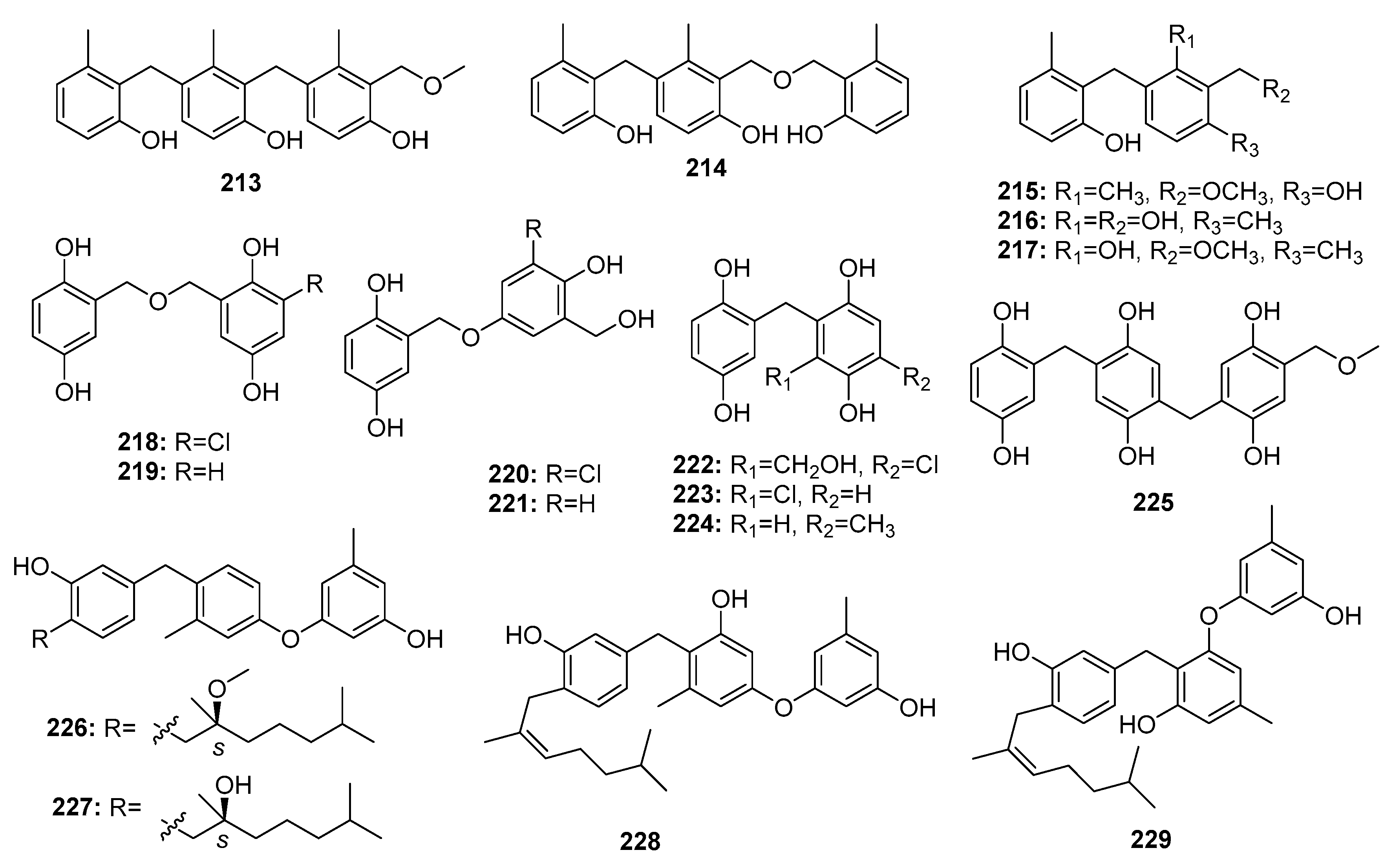
| Peniphenylane A (213) | P. fellutanum HDN14-323 | Deep sea sediment | Polyphenol | Hela | 14.5 μM | [141] | |
| Peniphenylane B (214) | P. fellutanum HDN14-323 | Deep sea sediment | Polyphenol | Hela, HCT-116 | (11.4, 15.8) μM | [141] | |
| Peniphenylane D (215) | P. fellutanum HDN14-323 | Deep sea sediment | Polyphenol | Hela, HL-60, HCT-116 | (9.3, 18.2, 31.7) μM | [141] | |
| Peniphenylane F (216) | P. fellutanum HDN14-323 | Deep sea sediment | Polyphenol | Hela | 29.3 μM | [141] | |
| Peniphenylane G (217) | P. fellutanum HDN14-323 | Deep sea sediment | Polyphenol | Hela, HL-60, HCT-116 | (16.6, 23.2, 24.7) μM | [141] | |
| Terrestol B (218) | P. terrestre | Marine sediment | Polyphenol | HL-60, MOLT-4, A549, BEL-7402 | (6.1, 5.8, 18.3, 62.3) μM | [142] | |
| Terrestol C (219) | P. terrestre | Marine sediment | Polyphenol | HL-60, MOLT-4, A549, BEL-7402 | (5.5, 5.6, 18.2, 57.3) μM | [142] | |
| Terrestol D (220) | P. terrestre | Marine sediment | Polyphenol | HL-60, MOLT-4, A549, BEL-7402 | (5.3, 5.5, 14.3, 38.5) μM | [142] | |
| Terrestol E (221) | P. terrestre | Marine sediment | Polyphenol | HL-60, MOLT-4, A549, BEL-7402 | (54.7, 6.4, 9.6, 59.0) μM | [142] | |
| Terrestol F (222) | P. terrestre | Marine sediment | Polyphenol | HL-60, MOLT-4, A549, BEL-7402 | (55.0, 58.1, 13.8, 63.2) μM | [142] | |
| Terrestol G (223) | P. terrestre | Marine sediment | Polyphenol | HL-60, MOLT-4, A549, BEL-7402 | (5.1, 6.5, 5.7, 6.0) μM | [142] | |
| Terrestol H (224) | P. terrestre | Marine sediment | Polyphenol | HL-60, MOLT-4, A549, BEL-7402 | (6.3, 5.8, 33.8, 61.9) μM | [142] | |
| Terrestol A (225) | P. terrestre | Marine sediment | Polyphenol | HL-60, MOLT-4, A549, BEL-7402 | (33.3, 5.5, 23.5, 57.0) μM | [142] | |
| Expansol A (226) | P. expansum 091006 | Mangrove | Polyphenol | HL-60 | 15.7 μM | [143] | |
| Expansol B (227) | P. expansum 091006 | Mangrove | Polyphenol | HL-60, A549 | (5.4, 1.9) μM | [143,144] | |
| Expansol C (228) | P. expansum 091006 | Mangrove | Polyphenol | HL-60 | 18.2 μM | [143] | |
| Expansol E (229) | P. expansum 091006 | Mangrove | Polyphenol | HL-60 | 20.8 μM | [143] | |
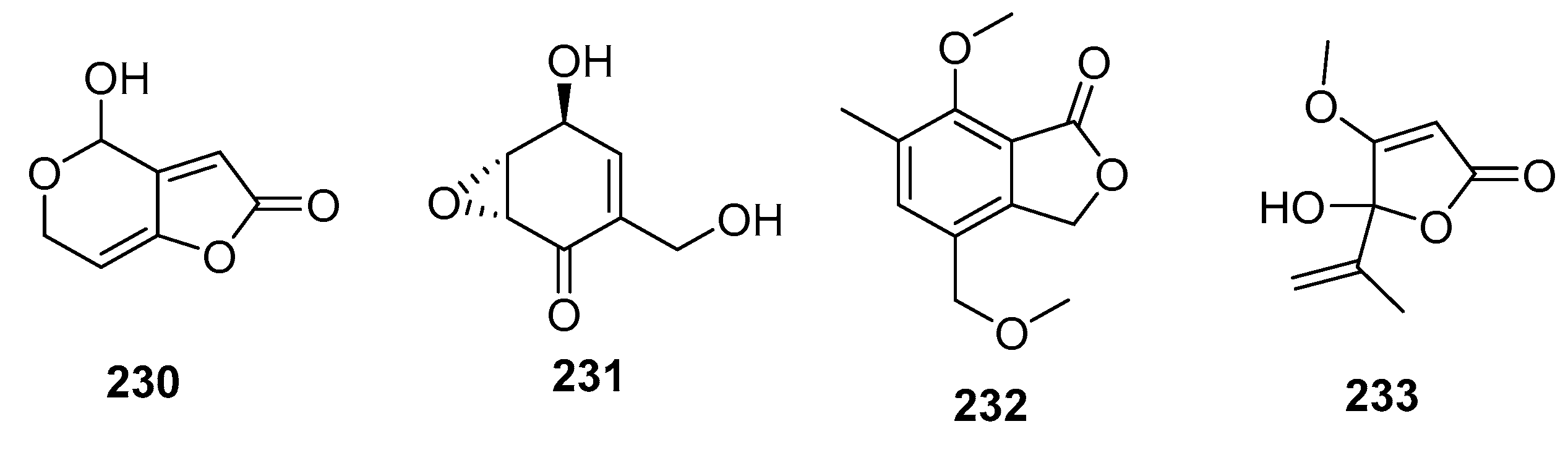
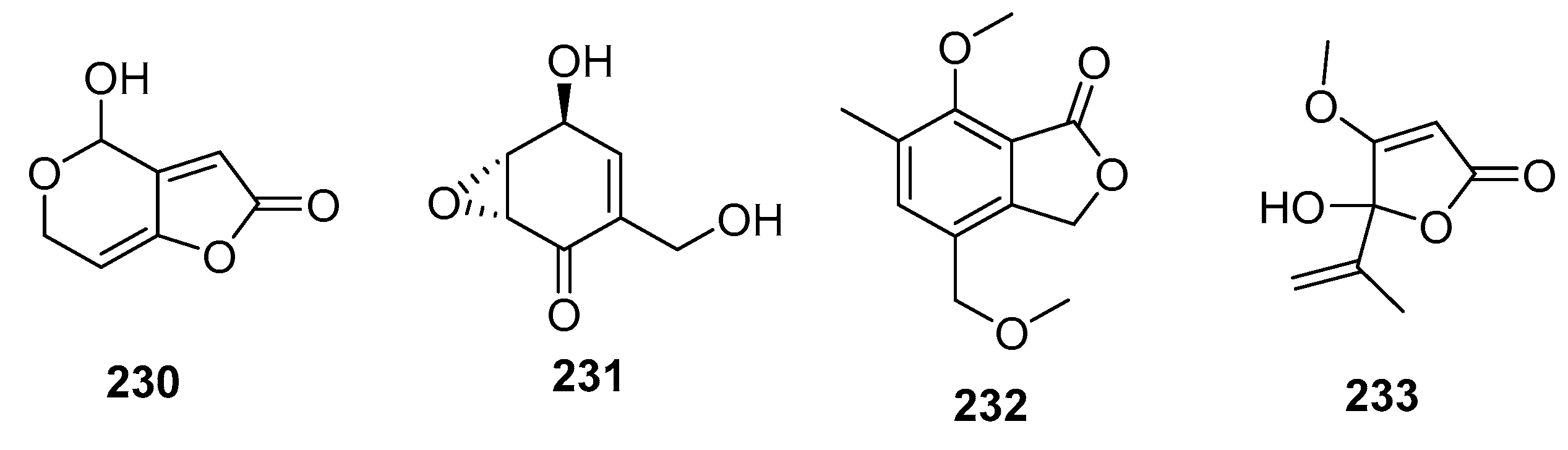
| Patulin (230) | Penicillium sp. | Marine alga | Other | P388, BSY-1, MCF-7, HCC2998, NCI-H522, DMS114, OVCAR-3, MKN1 | (0.06, 0.34, 0.65, 1.54, 0.30, 0.57, 0.37, 0.39) μg/mL | Potassium-uptake inhibitor Ion flux across cell membranes inducer | [111] |
| (+)-Epiepoxydon (231) | Penicillium sp. | Marine alga | Other | P388 | 0.2 μg/mL | [111] | |
| 232 | Penicillium sp. | Mangrove | Other | KB, KBv200 | (6, 10) μg/mL | [145] | |
| Penicillic acid (233) | Penicillium sp. | Sea water | Other | POS1, AT6-1, L299 | (7.8, 29.4, 12.9) μM | [84] |
References
- Lindequist, U. Marine-Derived Pharmaceuticals—Challenges and Opportunities. Biomol. Ther. 2016, 24, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hong, J.; Yin, J.; Moon, H.R.; Liu, Y.; Wei, X.; Oh, D.-C.; Jung, J.H. Dimeric Octaketide Spiroketals from the Jellyfish-Derived Fungus Paecilomyces variotii J08NF-1. J. Nat. Prod. 2015, 78, 2832–2836. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Kim, M.J.; Li, H.; Zhang, P.; Bao, B.; Lee, K.J.; Jung, J.H. Marine-derived Aspergillus species as a source of bioactive secondary metabolites. Mar. Biotechnol. 2013, 15, 499–519. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Song, Y.C.; Chen, J.R.; Xu, C.; Ge, H.M.; Wang, X.T.; Tan, R.X. Chaetoglobosin U, a cytochalasan alkaloid from endophytic Chaetomium globosum IFB-E019. J. Nat. Prod. 2006, 69, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ge, H.M.; Jiao, R.H.; Li, J.; Peng, H.; Wang, Y.R.; Wu, J.H.; Song, Y.C.; Tan, R.X. Cytotoxic chaetoglobosins from the endophyte Chaetomium globosum. Planta Med. 2010, 76, 1910–1914. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Chen, H.; Li, W.; Zhu, X.; Ding, W.; Li, C. Bioactive Chaetoglobosins from the Mangrove Endophytic Fungus Penicillium chrysogenum. Mar. Drugs 2016, 14, 172. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, C.; Yamada, T.; Ito, Y.; Minoura, K.; Numata, A. Cytotoxic cytochalasans from a Penicillium species separated from a marine alga. Tetrahedron 2001, 57, 2997–3004. [Google Scholar] [CrossRef]
- Numata, A.; Takahashi, C.; Ito, Y.; Minoura, K.; Yamada, T.; Matsuda, C.; Nomoto, K. Penochalasins, a novel class of cytotoxic cytochalasans from a Penicillium species separated from a marine alga: Structure determination and solution conformation. J. Chem. Soc. Perkin 1 1996, 3, 239–245. [Google Scholar] [CrossRef]
- Knudsen, P.B.; Hanna, B.; Ohl, S.; Sellner, L.; Zenz, T.; Dohner, H.; Stilgenbauer, S.; Larsen, T.O.; Lichter, P.; Seiffert, M. Chaetoglobosin A preferentially induces apoptosis in chronic lymphocytic leukemia cells by targeting the cytoskeleton. Leukemia 2014, 28, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Schümann, J.; Hertweck, C. Molecular basis of cytochalasan biosynthesis in fungi: Gene cluster analysis and evidence for the involvement of a PKS-NRPS hybrid synthase by RNA silencing. J. Am. Chem. Soc. 2007, 129, 9564–9565. [Google Scholar] [CrossRef] [PubMed]
- McDougall, J. Antiviral action of gliotoxin. Arch. Virol. 1969, 27, 255–267. [Google Scholar] [CrossRef]
- Sun, Y.; Takada, K.; Takemoto, Y.; Yoshida, M.; Nogi, Y.; Okada, S.; Matsunaga, S. Gliotoxin analogues from a marine-derived fungus, Penicillium sp., and their cytotoxic and histone methyltransferase inhibitory activities. J. Nat. Prod. 2011, 75, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Vigushin, D.; Mirsaidi, N.; Brooke, G.; Sun, C.; Pace, P.; Inman, L.; Moody, C.; Coombes, R. Gliotoxin is a dual inhibitor of farnesyltransferase and geranylgeranyltransferase I with antitumor activity against breast cancer in vivo. Med. Oncol. 2004, 21, 21–30. [Google Scholar] [CrossRef]
- Meng, L.-H.; Wang, C.-Y.; Mándi, A.; Li, X.-M.; Hu, X.-Y.; Kassack, M.U.; Kurtán, T.; Wang, B.-G. Three Diketopiperazine Alkaloids with Spirocyclic Skeletons and One Bisthiodiketopiperazine Derivative from the Mangrove-Derived Endophytic Fungus Penicillium brocae MA-231. Org. Lett. 2016, 18, 5304–5307. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.-S.; Guo, Y.-W. Epipolythiodioxopiperazines from fungi: Chemistry and bioactivities. Mini Rev. Med. Chem. 2011, 11, 728–745. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.-H.; Li, X.-M.; Lv, C.-T.; Huang, C.-G.; Wang, B.-G. Brocazines A–F, cytotoxic bisthiodiketopiperazine derivatives from Penicillium brocae MA-231, an endophytic fungus derived from the marine mangrove plant Avicennia marina. J. Nat. Prod. 2014, 77, 1921–1927. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, X.-M.; Meng, L.-H.; Jiang, W.-L.; Xu, G.-M.; Huang, C.-G.; Wang, B.-G. Bisthiodiketopiperazines and acorane sesquiterpenes produced by the marine-derived fungus Penicillium adametzioides AS-53 on different culture media. J. Nat. Prod. 2015, 78, 1294–1299. [Google Scholar] [CrossRef] [PubMed]
- Orfali, R.S.; Aly, A.H.; Ebrahim, W.; Abdel-Aziz, M.S.; Müller, W.E.; Lin, W.; Daletos, G.; Proksch, P. Pretrichodermamide C and N-methylpretrichodermamide B, two new cytotoxic epidithiodiketopiperazines from hyper saline lake derived Penicillium sp. Phytochem. Lett. 2015, 11, 168–172. [Google Scholar] [CrossRef]
- Yurchenko, A.N.; Smetanina, O.F.; Ivanets, E.V.; Kalinovsky, A.I.; Khudyakova, Y.V.; Kirichuk, N.N.; Popov, R.S.; Bokemeyer, C.; Von Amsberg, G.; Chingizova, E.A. Pretrichodermamides D–F from a Marine Algicolous Fungus Penicillium sp. KMM 4672. Mar. Drugs 2016, 14, 122. [Google Scholar] [CrossRef] [PubMed]
- Aninat, C.; Andre, F.; Delaforge, M. Oxidative metabolism by P450 and function coupling to efflux systems: Modulation of mycotoxin toxicity. Food Addit. Contam. 2005, 22, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Feng, T.; Zhao, B.; Li, D.; Cai, S.; Zhu, T.; Wang, F.; Xiao, X.; Gu, Q. Alkaloids from a deep ocean sediment-derived fungus Penicillium sp. and their antitumor activities. J. Antibiot. 2010, 63, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Li, D.; Zhu, T.; Cai, S.; Wang, F.; Xiao, X.; Gu, Q. New alkaloids and diterpenes from a deep ocean sediment derived fungus Penicillium sp. Tetrahedron 2009, 65, 1033–1039. [Google Scholar] [CrossRef]
- Shang, Z.; Li, X.; Meng, L.; Li, C.; Gao, S.; Huang, C.; Wang, B. Chemical profile of the secondary metabolites produced by a deep sea sediment-derived fungus Penicillium commune SD-118. Chin. J. Oceanol. Limnol. 2012, 30, 305–314. [Google Scholar] [CrossRef]
- Fang, S.-M.; Wu, C.-J.; Li, C.-W.; Cui, C.-B. A practical strategy to discover new antitumor compounds by activating silent metabolite production in fungi by diethyl sulphate mutagenesis. Mar. Drugs 2014, 12, 1788–1814. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.-J.; Cui, C.-B.; Li, C.-W.; Wu, C.-J.; Tian, C.-K.; Hua, W. Activation of the dormant secondary metabolite production by introducing gentamicin-resistance in a marine-derived Penicillium purpurogenum G59. Mar. Drugs 2012, 10, 559–582. [Google Scholar] [CrossRef] [PubMed]
- Hauser, D.; Weber, H.; Sigg, H. Isolation and configuration of Chaetocin. Helv. Chim. Acta 1970, 53, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, K.; Sato, K.; Hayakawa, S.; Matsushima, T.; Minato, H. Verticillin A, a new anti-biotic from Verticillium sp. J. Antibiot. 1970, 23, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Son, B.; Jensen, P.; Kauffman, C.; Fenical, W. New Cytotoxic Epidithiodioxopiperazines Related to Verticillin A From A Marine Isolate of the Fungus Penicillium. Nat. Prod. Lett. 1999, 13, 213–222. [Google Scholar] [CrossRef]
- Greiner, D.; Bonaldi, T.; Eskeland, R.; Roemer, E.; Imhof, A. Identification of a specific inhibitor of the histone methyltransferase SU (VAR) 3-9. Nat. Chem. Biol. 2005, 1, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Miao, Z.-H.; Zhao, W.-M.; Ding, J. The p53 pathway is synergized by p38 MAPK signaling to mediate 11,11′-dideoxyverticillin-induced G2/M arrest. FEBS Lett. 2005, 579, 3683–3690. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-X.; Chen, Y.; Guo, X.-N.; Zhang, X.-W.; Zhao, W.-M.; Zhong, L.; Zhou, J.; Xi, Y.; Lin, L.-P.; Ding, J. 11,11′-dideoxy-verticillin: A natural compound possessing growth factor receptor tyrosine kinase-inhibitory effect with anti-tumor activity. Anti-Cancer Drugs 2005, 16, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ashenhurst, J.A.; Movassaghi, M. Total synthesis of (+)-11,11′-dideoxyverticillin A. Science 2009, 324, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Movassaghi, M. Biogenetically-inspired total synthesis of epidithiodiketopiperazines and related alkaloids. Acc. Chem. Res. 2015, 48, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.A.; Houssen, W.E.; Gissendanner, C.R.; Orabi, K.Y.; Foudah, A.I.; El Sayed, K.A. Bioguided discovery and pharmacophore modeling of the mycotoxic indole diterpene alkaloids penitrems as breast cancer proliferation, migration, and invasion inhibitors. MedChemComm 2013, 4, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Sings, H.; Singh, S. Tremorgenic and nontremorgenic 2,3-fused indole diterpenoids. Alkaloids Chem. Biol. 2003, 60, 51–163. [Google Scholar] [PubMed]
- Saikia, S.; Nicholson, M.J.; Young, C.; Parker, E.J.; Scott, B. The genetic basis for indole-diterpene chemical diversity in filamentous fungi. Mycol. Res. 2008, 112, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, J.; Holton, J.; Nolan, C.; Ray, D.; Naik, J.; Mantle, P. The effects of the tremorgenic mycotoxin penitrem A on the rat cerebellum. Vet. Pathol. 1998, 35, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Numata, A.; Takahashi, C.; Ito, Y.; Takada, T.; Kawai, K.; Usami, Y.; Matsumura, E.; Imachi, M.; Ito, T.; Hasegawa, T. Communesins, cytotoxic metabolites of a fungus isolated from a marine alga. Tetrahedron Lett. 1993, 34, 2355–2358. [Google Scholar] [CrossRef]
- Jadulco, R.; Edrada, R.A.; Ebel, R.; Berg, A.; Schaumann, K.; Wray, V.; Steube, K.; Proksch, P. New Communesin Derivatives from the Fungus Penicillium sp. Derived from the Mediterranean Sponge Axinella v errucosa. J. Nat. Prod. 2004, 67, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Vansteelandt, M.; Kerzaon, I.; Blanchet, E.; Tankoua, O.F.; Du Pont, T.R.; Joubert, Y.; Monteau, F.; Le Bizec, B.; Frisvad, J.C.; Pouchus, Y.F. Patulin and secondary metabolite production by marine-derived Penicillium strains. Fungal Biol. 2012, 116, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Crawley, S.L.; Funk, R.L. A synthetic approach to nomofungin/communesin B. Org. Lett. 2003, 5, 3169–3171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, X.-M.; Liu, H.; Li, X.; Wang, B.-G. Two new alkaloids from Penicillium oxalicum EN-201, an endophytic fungus derived from the marine mangrove plant Rhizophora stylosa. Phytochem. Lett. 2015, 13, 160–164. [Google Scholar] [CrossRef]
- An, C.-Y.; Li, X.-M.; Li, C.-S.; Xu, G.-M.; Wang, B.-G. Prenylated indolediketopiperazine peroxides and related homologues from the marine sediment-derived fungus Penicillium brefeldianum SD-273. Mar. Drugs 2014, 12, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Li, C.S.; Li, X.M.; An, C.Y.; Wang, B.G. Prenylated Indole Alkaloid Derivatives from Marine Sediment-Derived Fungus Penicillium paneum SD-44. Helv. Chim. Acta 2014, 97, 1440–1444. [Google Scholar] [CrossRef]
- Asiri, I.A.; Badr, J.M.; Youssef, D.T. Penicillivinacine, antimigratory diketopiperazine alkaloid from the marine-derived fungus Penicillium vinaceum. Phytochem. Lett. 2015, 13, 53–58. [Google Scholar] [CrossRef]
- Shaala, L.A.; Youssef, D.T. Identification and bioactivity of compounds from the fungus Penicillium sp. CYE-87 isolated from a marine tunicate. Mar. Drugs 2015, 13, 1698–1709. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, K.; Zhong, P. Tumonoic Acids K and L, Novel Metabolites from the Marine-Derived Fungus Penicillium citrinum. Heterocycles 2012, 85, 413–419. [Google Scholar] [CrossRef]
- Chen, L.; Zhou, T.; Zhao, Y.-Y. Four new penicitrinols and two new penicillenols from the marine-derived fungus Penicillium citrinum. Heterocycles 2015, 91, 1007–1016. [Google Scholar] [CrossRef]
- Lin, Z.-J.; Lu, Z.-Y.; Zhu, T.-J.; Fang, Y.-C.; Gu, Q.-Q.; Zhu, W.-M. Penicillenols from Penicillium sp. GQ-7, an endophytic fungus associated with Aegiceras corniculatum. Chem. Pharm. Bull. 2008, 56, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.-Y.; Zhou, T.; Zhao, Y.-Y.; Chen, L.; Gong, M.-W.; Xia, Q.-W.; Ying, M.-G.; Zheng, Q.-H.; Zhang, Q.-Q. Antitumor effects and related mechanisms of penicitrinine A, a novel alkaloid with a unique spiro skeleton from the marine fungus Penicillium citrinum. Mar. Drugs 2015, 13, 4733–4753. [Google Scholar] [CrossRef] [PubMed]
- Jafari, E.; Khajouei, M.R.; Hassanzadeh, F.; Hakimelahi, G.H.; Khodarahmi, G.A. Quinazolinone and quinazoline derivatives: Recent structures with potent antimicrobial and cytotoxic activities. Res. Pharm. Sci. 2016, 11, 1–14. [Google Scholar] [PubMed]
- He, J.; Lion, U.; Sattler, I.; Gollmick, F.A.; Grabley, S.; Cai, J.; Meiners, M.; Schünke, H.; Schaumann, K.; Dechert, U. Diastereomeric Quinolinone Alkaloids from the Marine-Derived Fungus Penicillium j anczewskii. J. Nat. Prod. 2005, 68, 1397–1399. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, L.; Zhu, T.; Gu, Q.; Li, D. Unusual pyrrolyl 4-quinolinone alkaloids from the marine-derived fungus Penicillium sp. ghq208. Chem. Pharm. Bull. 2012, 60, 1458–1460. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.-L.; Wang, C.-Y.; Gu, Y.-C.; Wei, M.-Y.; Pan, J.-H.; Deng, D.-S.; She, Z.-G.; Lin, Y.-C. Penicinoline, a new pyrrolyl 4-quinolinone alkaloid with an unprecedented ring system from an endophytic fungus Penicillium sp. Bioorg. Med. Chem. Lett. 2010, 20, 3284–3286. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Li, W.; Wang, J. A novel and other bioactive secondary metabolites from a marine fungus Penicillium oxalicum 0312f1. Nat. Prod. Rep. 2013, 27, 2286–2291. [Google Scholar] [CrossRef] [PubMed]
- An, C.Y.; Li, X.M.; Li, C.S.; Gao, S.S.; Shang, Z.; Wang, B.G. Triazoles and Other N-Containing Metabolites from the Marine-Derived Endophytic Fungus Penicillium chrysogenum EN-118. Helv. Chim. Acta 2013, 96, 682–687. [Google Scholar] [CrossRef]
- Li, C.-S.; Li, X.-M.; Gao, S.-S.; Lu, Y.-H.; Wang, B.-G. Cytotoxic anthranilic acid derivatives from deep sea sediment-derived fungus Penicillium paneum SD-44. Mar. Drugs 2013, 11, 3068–3076. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-S.; An, C.-Y.; Li, X.-M.; Gao, S.-S.; Cui, C.-M.; Sun, H.-F.; Wang, B.-G. Triazole and dihydroimidazole alkaloids from the marine sediment-derived fungus Penicillium paneum SD-44. J. Nat. Prod. 2011, 74, 1331–1334. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, X.; Lin, Y.; Yuan, J.; Lu, Y.; Zhu, X.; Li, J.; Li, M.; Lin, Y.; He, J. Meroterpenes and azaphilones from marine mangrove endophytic fungus Penicillium 303. Fitoterapia 2014, 97, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, G.; Lang, G.; Gulder, T.A.; Tsuruta, H.; Mühlbacher, J.; Maksimenka, K.; Steffens, S.; Schaumann, K.; Stöhr, R.; Wiese, J. The first sorbicillinoid alkaloids, the antileukemic sorbicillactones A and B, from a sponge-derived Penicillium chrysogenum strain. Tetrahedron 2005, 61, 7252–7265. [Google Scholar] [CrossRef]
- Bringmann, G.; Gulder, T.A.; Lang, G.; Schmitt, S.; Stöhr, R.; Wiese, J.; Nagel, K.; Imhoff, J.F. Large-scale biotechnological production of the antileukemic marine natural product sorbicillactone A. Mar. Drugs 2007, 5, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Meng, L.H.; Mándi, A.; Kurtán, T.; Li, X.M.; Liu, Y.; Li, X.; Li, C.S.; Wang, B.G. Brocaeloids A–C, 4-Oxoquinoline and Indole Alkaloids with C-2 Reversed Prenylation from the Mangrove-Derived Endophytic Fungus Penicillium brocae. Eur. J. Org. Chem. 2014, 2014, 4029–4036. [Google Scholar] [CrossRef]
- He, X.; Zhang, Z.; Chen, Y.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Varitatin A, a Highly Modified Fatty Acid Amide from Penicillium variabile Cultured with a DNA Methyltransferase Inhibitor. J. Nat. Prod. 2015, 78, 2841–2845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, C.; Swenson, D.C.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. Novel antiinsectan oxalicine alkaloids from two undescribed fungicolous Penicillium spp. Org. Lett. 2003, 5, 773–776. [Google Scholar] [CrossRef] [PubMed]
- Takatsuki, A.; Suzuki, S.; Ando, K.; Tamura, G.; Arima, K. New antiviral antibiotics; xanthocillin X mono-and dimethylether, and methoxy-xanthocillin X dimethylether. I: Isolation and characterization. J. Antibiot. 1968, 21, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Rothe, W. Vorläufige Mitteilung über eine neues Antibiotikum. Pharmazie 1950, 5, 190. [Google Scholar]
- Zhao, Y.; Chen, H.; Shang, Z.; Jiao, B.; Yuan, B.; Sun, W.; Wang, B.; Miao, M.; Huang, C. SD118-xanthocillin X (1), a novel marine agent extracted from Penicillium commune, induces autophagy through the inhibition of the MEK/ERK pathway. Mar. Drugs 2012, 10, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, F.; Hallock, Y.; Bunkers, G.; Kenfield, D.; Strobel, G.; Yoshida, S. Phytoactive eremophilanes produced by the weed pathogen Drechslera gigantea. Biosci. Biotechnol. Biochem. 1993, 57, 236–239. [Google Scholar] [CrossRef] [PubMed]
- Tirilly, Y.; Kloosterman, J.; Sipma, G.; Kettenes-Van den Bosch, J.J. A fungitoxic sesquiterpene from Hansfordia pulvinata. Phytochemistry 1983, 22, 2082–2083. [Google Scholar] [CrossRef]
- Lin, A.; Wu, G.; Gu, Q.; Zhu, T.; Li, D. New eremophilane-type sesquiterpenes from an Antarctic deep sea derived fungus, Penicillium sp. PR19 N-1. Arch. Pharm. Res. 2014, 37, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lin, A.; Gu, Q.; Zhu, T.; Li, D. Four new chloro-eremophilane sesquiterpenes from an Antarctic deep sea derived fungus, Penicillium sp. PR19N-1. Mar. Drugs 2013, 11, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.F.; Qiao, L.; Lv, A.L.; Pei, Y.H.; Tian, L. Eremophilane sesquiterenes from the marine fungus Penicillium sp. BL27–2. Chin. Chem. Lett. 2008, 19, 562–564. [Google Scholar] [CrossRef]
- Li, Y.; Ye, D.; Shao, Z.; Cui, C.; Che, Y. A sterol and spiroditerpenoids from a Penicillium sp. isolated from a deep sea sediment sample. Mar. Drugs 2012, 10, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-S.; Li, X.-M.; Li, C.-S.; Proksch, P.; Wang, B.-G. Penicisteroids A and B, antifungal and cytotoxic polyoxygenated steroids from the marine alga-derived endophytic fungus Penicillium chrysogenum QEN-24S. Bioorg. Med. Chem. Lett. 2011, 21, 2894–2897. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Tian, L.; Huang, J.; Li, W.; Pei, Y.-H. Cytotoxic sterols from marine-derived fungus Pennicillium sp. Nat. Prod. Rep. 2006, 20, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Xin, Z.-H.; Zhu, T.-J.; Wang, W.-L.; Du, L.; Fang, Y.-C.; Gu, Q.-Q.; Zhu, W.-M. Isocoumarin derivatives from the sea squirt-derived fungus Penicillium stoloniferum QY2-10 and the halotolerant fungus Penicillium notatum B-52. Arch. Pharm. Res. 2007, 30, 816–819. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Shao, C.-L.; Liu, M.; Qi, X.; Wang, C.-Y. Bioactive steroids from a marine-derived fungus Penicillium sp. from the South China Sea. Chem. Nat. Compd. 2014, 3, 568–570. [Google Scholar] [CrossRef]
- Menna, M.; Imperatore, C.; D’Aniello, F.; Aiello, A. Meroterpenes from marine invertebrates: Structures, occurrence, and ecological implications. Mar. Drugs 2013, 11, 1602–1643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Choi, H.D.; Kang, J.S.; Lee, C.-O.; Son, B.W. New polyoxygenated farnesylcyclohexenones, deacetoxyyanuthone A and its hydro derivative from the marine-derived fungus Penicillium sp. J. Nat. Prod. 2003, 66, 1499–1500. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, P.; Tang, X.-X.; Yi, Z.-W.; Qiu, Y.-K.; Wu, Z. Two new compounds from marine-derived fungus Penicillium sp. F11. J. Asian Nat. Prod. Res. 2012, 14, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Zhang, L.; Fu, J.; Che, Q.; Li, D.; Gu, Q.; Zhu, T. Phenylpyropenes E and F: New meroterpenes from the marine-derived fungus Penicillium concentricum ZLQ-69. J. Antibiot. 2015, 68, 748–751. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Lee, J.-H.; You, M.; Choi, T.J.; Park, W.; Lee, S.K.; Oh, D.-C.; Oh, K.-B.; Shin, J. Penicillipyrones A and B, meroterpenoids from a marine-derived Penicillium sp. fungus. J. Nat. Prod. 2014, 77, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Hanson, F.; Eble, T. An antiphage agent isolated from Aspergillus sp. J. Bacteriol. 1949, 58, 527–529. [Google Scholar] [PubMed]
- Vansteelandt, M.; Blanchet, E.; Egorov, M.; Petit, F.; Toupet, L.; Bondon, A.; Monteau, F.; Le Bizec, B.; Thomas, O.P.; Pouchus, Y.F. Ligerin, an antiproliferative chlorinated sesquiterpenoid from a marine-derived Penicillium strain. J. Nat. Prod. 2013, 76, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, E.; Vansteelandt, M.; Bot, R.L.; Egorov, M.; Guitton, Y.; Pouchus, Y.F.; Grovel, O. Synthesis and antiproliferative activity of ligerin and new fumagillin analogs against osteosarcoma. Eur. J. Med. Chem. 2014, 79, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Keri, R.S.; Budagumpi, S.; Pai, R.K.; Balakrishna, R.G. Chromones as a privileged scaffold in drug discovery: A review. Eur. J. Med. Chem. 2014, 78, 340–374. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Luo, J.-F.; Qin, X.-C.; Xu, X.-Y.; Zhang, X.-Y.; Tu, Z.-C.; Qi, S.-H. Dihydrothiophene-condensed chromones from a marine-derived fungus Penicillium oxalicum and their structure-bioactivity relationship. Bioorg. Med. Chem. Lett. 2014, 24, 2433–2436. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-L.; Bao, J.; Liu, K.-S.; Zhang, X.-Y.; He, F.; Wang, Y.-F.; Nong, X.-H.; Qi, S.-H. Cytotoxic dihydrothiophene-condensed chromones from the marine-derived fungus Penicillium oxalicum. Planta Med. 2013, 79, 1474–1479. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Q.-L.; Nong, X.-H.; Zhang, X.-Y.; Xu, X.-Y.; Qi, S.-H.; Wang, Y.-F. Oxalicumone A, a new dihydrothiophene-condensed sulfur chromone induces apoptosis in leukemia cells through endoplasmic reticulum stress pathway. Eur. J. Pharmacol. 2016, 783, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Guo, K.; Wang, X.; Chen, H.; Min, J.; Qi, S.; Zhao, W.; Li, W. Toxicity study of oxalicumone A, derived from a marine-derived fungus Penicillium oxalicum, in cultured renal epithelial cells. Mol. Med. Rep. 2017, 15, 2611–2619. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Cui, C.-B.; Li, C.-W.; Peng, J.-X.; Gu, Q.-Q. Chromosulfine, a novel cyclopentachromone sulfide produced by a marine-derived fungus after introduction of neomycin resistance. RSC Adv. 2016, 6, 43975–43979. [Google Scholar] [CrossRef]
- Li, N.; Yi, Z.; Wang, Y.; Zhang, Q.; Zhong, T.; Qiu, Y.; Wu, Z.; Tang, X. Differential proteomic analysis of HL60 cells treated with secalonic acid F reveals caspase 3-induced cleavage of Rho GDP dissociation inhibitor 2. Oncol. Rep. 2012, 28, 2016–2022. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Sun, H.; Liu, D.; Zhang, J.; Zhang, J.; Yan, M.; Pan, X. Secalonic acid-F inhibited cell growth more effectively than 5-fluorouracil on hepatocellular carcinoma in vitro and in vivo. Neoplasma 2017, 64, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.-M.; Wang, C.-Y.; Gu, Y.-C.; Shao, C.-L. Penimethavone A, a flavone from a gorgonian-derived fungus Penicillium chrysogenum. Nat. Prod. Rep. 2016, 30, 2274–2277. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, J.; Wang, Z.; Lin, X.; Zhao, B.; Kaliaperumal, K.; Liao, X.; Tu, Z.; Li, J.; Xu, S. Structurally diverse secondary metabolites from a deep sea-derived fungus Penicillium chrysogenum SCSIO 41001 and their biological evaluation. Fitoterapia 2017, 117, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Aktas, N.; Konuklugil, B.; Mandi, A.; Daletos, G.; Lin, W.; Dai, H.; Kurtan, T.; Proksch, P. A new fusarielin analogue from Penicillium sp. isolated from the Mediterranean sponge Ircinia oros. Tetrahedron Lett. 2015, 56, 5317–5320. [Google Scholar] [CrossRef]
- Huang, Z.; Yang, J.; Cai, X.; She, Z.; Lin, Y. A new furanocoumarin from the mangrove endophytic fungus Penicillium sp.(ZH16). Nat. Prod. Rep. 2012, 26, 1291–1295. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.C.; Raistrick, H. On the production and chemical constitution of a new yellow colouring matter, citrinin, produced from glucose by Penicillium citrinum Thom. Philos. Trans. R. Soc. Lond. 1931, 220, 269–295. [Google Scholar] [CrossRef]
- Chagas, G.M.; Campello, A.P.; Klüppel, M.; Lúcia, W. Mechanism of citrinin-induced dysfunction of mitochondria. I. Effects on respiration, enzyme activities and membrane potential of renal cortical mitochondria. J. Appl. Toxicol. 1992, 12, 123–129. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cox, R.J. The molecular steps of citrinin biosynthesis in fungi. Chem. Sci. 2016, 7, 2119–2127. [Google Scholar] [CrossRef]
- Subramani, R.; Kumar, R.; Prasad, P.; Aalbersberg, W. Cytotoxic and antibacterial substances against multi-drug resistant pathogens from marine sponge symbiont: Citrinin, a secondary metabolite of Penicillium sp. Asian Pac. J. Trop. Biomed. 2013, 3, 291–296. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B.; Kelly, K. Berkelic acid, a novel spiroketal with selective anticancer activity from an acid mine waste fungal extremophile. J. Org. Chem. 2006, 71, 5357–5360. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhou, J.; Snider, B.B. Synthesis of (−)-Berkelic Acid. Angew. Chem. Int. Ed. 2009, 48, 1283–1286. [Google Scholar] [CrossRef] [PubMed]
- Zhuravleva, O.I.; Sobolevskaya, M.P.; Afiyatullov, S.S.; Kirichuk, N.N.; Denisenko, V.A.; Dmitrenok, P.S.; Yurchenko, E.A.; Dyshlovoy, S.A. Sargassopenillines A–G, 6,6-spiroketals from the alga-derived fungi Penicillium thomii and Penicillium lividum. Mar. Drugs 2014, 12, 5930–5943. [Google Scholar] [CrossRef] [PubMed]
- Perpelescu, M.; Kobayashi, J.I.; Furuta, M.; Ito, Y.; Izuta, S.; Takemura, M.; Suzuki, M.; Yoshida, S. Novel phenalenone derivatives from a marine-derived fungus exhibit distinct inhibition spectra against eukaryotic DNA polymerases. Biochemistry 2002, 41, 7610–7616. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gong, M.-W.; Peng, Z.-F.; Zhou, T.; Ying, M.-G.; Zheng, Q.-H.; Liu, Q.-Y.; Zhang, Q.-Q. The marine fungal metabolite, dicitrinone B, induces A375 cell apoptosis through the ROS-related caspase pathway. Mar. Drugs 2014, 12, 1939–1958. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Tsuda, M.; Sekiguchi, M.; Mikami, Y.; Kobayashi, J.I. Perinadine A, a Novel Tetracyclic Alkaloid from Marine-Derived Fungus Penicillium citrinum. Org. Lett. 2005, 7, 4261–4264. [Google Scholar] [CrossRef] [PubMed]
- Julianti, E.; Lee, J.-H.; Liao, L.; Park, W.; Park, S.; Oh, D.-C.; Oh, K.-B.; Shin, J. New polyaromatic metabolites from a marine-derived fungus Penicillium sp. Org. Lett. 2013, 15, 1286–1289. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-S.; Li, X.-M.; Zhang, Y.; Li, C.-S.; Cui, C.-M.; Wang, B.-G. Comazaphilones A–F, azaphilone derivatives from the marine sediment-derived fungus Penicillium commune QSD-17. J. Nat. Prod. 2011, 74, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Myobatake, Y.; Takeuchi, T.; Kuramochi, K.; Kuriyama, I.; Ishido, T.; Hirano, K.; Sugawara, F.; Yoshida, H.; Mizushina, Y. Pinophilins A and B, inhibitors of mammalian A-, B-, and Y-family DNA polymerases and human cancer cell proliferation. J. Nat. Prod. 2012, 75, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, C.; Minoura, K.; Oka, T.; Ohta, T.; Hagishita, S.; Numata, A. Absolute stereostructures of novel cytotoxic metabolites, penostatins A–E, from a Penicillium species separated from an Enteromorpha alga. Tetrahedron 1999, 55, 14353–14368. [Google Scholar] [CrossRef]
- Iwamoto, C.; Minoura, K.; Hagishita, S.; Nomoto, K.; Numata, A. Penostatins F–I, novel cytotoxic metabolites from a Penicillium species separated from an Enteromorpha marine alga. J. Chem. Soc. Perkin 1 1998, 3, 449–456. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Yang, C.-G.; Wei, P.-Y.; Li, L.; Luo, D.-Q.; Zheng, Z.-H.; Lu, X.-H. Penostatin derivatives, a novel kind of protein phosphatase 1B inhibitors isolated from solid cultures of the entomogenous fungus Isaria tenuipes. Molecules 2014, 19, 1663–1671. [Google Scholar] [CrossRef] [PubMed]
- Snider, B.B.; Liu, T. Total Synthesis of (±)-Deoxypenostatin A. Approaches to the Syntheses of Penostatins A and B. J. Org. Chem. 2000, 65, 8490–8498. [Google Scholar] [CrossRef] [PubMed]
- Barriault, L.; Ang, P.J.; Lavigne, R.M. Rapid Assembly of the Bicyclo [5.3.1] undecenone Core of Penostatin F: A Successive Diels-Alder/Claisen Reaction Strategy with an Efficient Stereochemical Relay. Org. Lett. 2004, 6, 1317–1319. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.M.; Townsend, C.A. New insights into the formation of fungal aromatic polyketides. Nat. Rev. Microbiol. 2010, 8, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wang, H.-F.; Zhang, L.-H.; Liu, F.; He, F.-J.; Bai, J.; Hua, H.-M.; Chen, G.; Pei, Y.-H. New compound with DNA Topo I inhibitory activity purified from Penicillium oxalicum HSY05. Nat. Prod. Rep. 2015, 29, 2197–2202. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Zhuang, Y.; Kong, F.; Zhang, C.; Zhu, W. Phenolic polyketides from the co-cultivation of marine-derived Penicillium sp. WC-29–5 and Streptomyces fradiae 007. Mar. Drugs 2014, 12, 2079–2088. [Google Scholar] [CrossRef] [PubMed]
- Roullier, C.; Guitton, Y.; Valery, M.; Amand, S.; Prado, S.; Robiou du Pont, T.; Grovel, O.; Pouchus, Y.F. Automated detection of natural halogenated compounds from LC-MS profiles–Application to the isolation of bioactive chlorinated compounds from marine-derived fungi. Anal. Chem. 2016, 88, 9143–9150. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Li, X.-M.; Li, C.-S.; Wang, B.-G. Diphenyl ether and benzophenone derivatives from the marine mangrove-derived fungus Penicillium sp. MA-37. Phytochem. Lett. 2014, 9, 22–25. [Google Scholar] [CrossRef]
- Nishida, H.; Tomoda, H.; Cao, J.; Araki, S.; Okuda, S.; Omura, S. Purpactins, new inhibitors of Acyl-CoA: Cholesterol acyltransferase produced by Penicillium purpurogenum. J. Antibiot. 1991, 44, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.-C.; Lee, H.; Chun, H.; Kho, Y. Penicillide, a nonpeptide calpain inhibitor, produced by Penicillium sp. J. Microbiol. Biotechnol. 1998, 8, 188–190. [Google Scholar]
- Salituro, G.M.; Pettibone, D.J.; Clineschmidt, B.V.; Williamson, J.M.; Zink, D.L. Potent, non-peptidic oxytocin receptor antagonists from a natural source. Bioorg. Med. Chem. Lett. 1993, 3, 337–340. [Google Scholar] [CrossRef]
- Gao, H.; Zhou, L.; Li, D.; Gu, Q.; Zhu, T.J. New Cytotoxic Metabolites from the Marine-Derived Fungus Penicillium sp. ZLN29. Helv. Chim. Acta 2013, 96, 514–519. [Google Scholar] [CrossRef]
- Zhao, D.-L.; Shao, C.-L.; Zhang, Q.; Wang, K.-L.; Guan, F.-F.; Shi, T.; Wang, C.-Y. Azaphilone and diphenyl ether derivatives from a gorgonian-derived strain of the fungus Penicillium pinophilum. J. Nat. Prod. 2015, 78, 2310–2314. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Liu, W.-W. Nidurufin as a new cell cycle inhibitor from marine-derived fungus Penicillium flavidorsum SHK1-27. Arch. Pharm. Res. 2011, 34, 901–905. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, G.J.; Murphy, R.A., Jr.; Cava, M.P. Aflatoxin precursors: Total synthesis of (.+−.)-averufin and (.+−.)-nidurufin. J. Org. Chem. 1985, 50, 5533–5537. [Google Scholar] [CrossRef]
- Liu, W.; Gu, Q.; Zhu, W.; Cui, C.; Fan, G. Two new benzoquinone derivatives and two new bisorbicillinoids were isolated from a marine-derived fungus Penicillium terrestre. J. Antibiot. 2005, 58, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Gu, Q.; Zhu, W.; Cui, C.; Fan, G. Dihydrotrichodimerol and tetrahydrotrichodimerol, two new bisorbicillinoids, from a marine-derived Penicillium terrestre. J. Antibiot. 2005, 58, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Harned, A.M.; Volp, K.A. The sorbicillinoid family of natural products: Isolation, biosynthesis, and synthetic studies. Nat. Prod. Rep. 2011, 28, 1790–1810. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, L.; Zhu, T.; Kurtán, T.; Mándi, A.; Zhao, Z.; Li, J.; Gu, Q. Chloctanspirones A and B, novel chlorinated polyketides with an unprecedented skeleton, from marine sediment derived fungus Penicillium terrestre. Tetrahedron 2011, 67, 7913–7918. [Google Scholar] [CrossRef]
- Janin, Y.L. Heat shock protein 90 inhibitors. A text book example of medicinal chemistry? J. Med. Chem. 2005, 48, 7503–7512. [Google Scholar] [CrossRef] [PubMed]
- De Castro, M.V.; Ióca, L.P.; Williams, D.E.; Costa, B.Z.; Mizuno, C.M.; Santos, M.F.; de Jesus, K.; Ferreira, E.V.L.; Seleghim, M.H.; Sette, L.D. Condensation of Macrocyclic Polyketides Produced by Penicillium sp. DRF2 with Mercaptopyruvate Represents a New Fungal Detoxification Pathway. J. Nat. Prod. 2016, 79, 1668–1678. [Google Scholar] [CrossRef] [PubMed]
- Greve, H.; Schupp, P.J.; Eguereva, E.; Kehraus, S.; Kelter, G.; Maier, A.; Fiebig, H.H.; König, G.M. Apralactone A and a New Stereochemical Class of Curvularins from the Marine Fungus Curvularia sp. Eur. J. Org. Chem. 2008, 30, 5085–5092. [Google Scholar] [CrossRef] [PubMed]
- Getino, M.; Fernández-López, R.; Palencia-Gándara, C.; Campos-Gómez, J.; Sánchez-López, J.M.; Martínez, M.; Fernández, A.; de la Cruz, F. Tanzawaic acids, a chemically novel set of bacterial conjugation inhibitors. PLoS ONE 2016, 11, e0148098. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J.; Pil, G.B.; Heo, S.-J.; Lee, H.-S.; Lee, J.S.; Lee, Y.-J.; Lee, J.; Won, H.S. Anti-inflammatory activity of tanzawaic acid derivatives from a marine-derived fungus Penicillium steckii 108YD142. Mar. Drugs 2016, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Martínez, F.; José, M.; Díaz-Marrero, A.R.; Darias, J.; Cerella, C.; Diederich, M.; Cueto, M. Tanzawaic acids isolated from a marine-derived fungus of the genus Penicillium with cytotoxic activities. Org. Biomol. Chem. 2015, 13, 7248–7256. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ai, X.; Hou, J.; Ye, X.; Liu, R.; Shen, S.; Li, Z.; Lu, S. Integrated discovery of FOXO1–DNA stabilizers from marine natural products to restore chemosensitivity to anti-EGFR-based therapy for metastatic lung cancer. Mol. Biosyst. 2017, 13, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Shigemori, H.; Wakuri, S.; Yazawa, K.; Nakamura, T.; Sasaki, T.; Kobayashi, J.I. Fellutamides A and B, cytotoxic peptides from a marine fish-possessing fungus Penicillium fellutanum. Tetrahedron 1991, 47, 8529–8534. [Google Scholar] [CrossRef]
- Wu, C.-J.; Li, C.-W.; Cui, C.-B. Seven new and two known lipopeptides as well as five known polyketides: The activated production of silent metabolites in a marine-derived fungus by chemical mutagenesis strategy using diethyl sulphate. Mar. Drugs 2014, 12, 1815–1838. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Guo, W.; He, X.; Che, Q.; Zhu, T.; Gu, Q.; Li, D. Peniphenylanes A–G from the Deep sea-Derived Fungus Penicillium fellutanum HDN14-323. Planta Med. 2016, 82, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Fang, Y.; Zhu, T.; Gu, Q.; Zhu, W. Gentisyl alcohol derivatives from the marine-derived fungus Penicillium terrestre. J. Nat. Prod. 2007, 71, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lu, Z.; Liu, P.; Wang, Y.; Li, J.; Hong, K.; Zhu, W. Cytotoxic Polyphenols from the Fungus Penicillium expansum 091 006 Endogenous with the Mangrove Plant Excoecaria agallocha. Planta Med. 2012, 78, 1861–1866. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Zhu, H.; Fu, P.; Wang, Y.; Zhang, Z.; Lin, H.; Liu, P.; Zhuang, Y.; Hong, K.; Zhu, W. Cytotoxic polyphenols from the marine-derived fungus Penicillium expansum. J. Nat. Prod. 2010, 73, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, R.; Qiu, S.X.; She, Z.; Lin, Y. A new isobenzofuranone from the mangrove endophytic fungus Penicillium sp.(ZH58). Nat. Prod. Rep. 2013, 27, 1902–1905. [Google Scholar] [CrossRef] [PubMed]
- Kornprobst, J.-M.; La Barre, S. New Trends in Marine Natural Products. J. Oceanogr. Mar. Res. 2014, 2, 1000e109. [Google Scholar]
- Zeng, Y.; Wang, H.; Kamdem, R.S.; Orfali, R.S.; Dai, H.; Makhloufi, G.; Janiak, C.; Liu, Z.; Proksch, P. A new cyclohexapeptide, penitropeptide and a new polyketide, penitropone from the endophytic fungus Penicillium tropicum. Tetrahedron Lett. 2016, 57, 2998–3001. [Google Scholar] [CrossRef]
- Koul, M.; Singh, S. Penicillium spp.: Prolific producer for harnessing cytotoxic secondary metabolites. Anti-Cancer Drugs 2017, 28, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Brady, S.F.; Simmons, L.; Kim, J.H.; Schmidt, E.W. Metagenomic approaches to natural products from free-living and symbiotic organisms. Nat. Prod. Rep. 2009, 26, 1488–1503. [Google Scholar] [CrossRef] [PubMed]




© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Su, M.; Song, S.-J.; Jung, J.H. Marine-Derived Penicillium Species as Producers of Cytotoxic Metabolites. Mar. Drugs 2017, 15, 329. https://doi.org/10.3390/md15100329
Liu S, Su M, Song S-J, Jung JH. Marine-Derived Penicillium Species as Producers of Cytotoxic Metabolites. Marine Drugs. 2017; 15(10):329. https://doi.org/10.3390/md15100329
Chicago/Turabian StyleLiu, Sen, Mingzhi Su, Shao-Jiang Song, and Jee H. Jung. 2017. "Marine-Derived Penicillium Species as Producers of Cytotoxic Metabolites" Marine Drugs 15, no. 10: 329. https://doi.org/10.3390/md15100329
APA StyleLiu, S., Su, M., Song, S. -J., & Jung, J. H. (2017). Marine-Derived Penicillium Species as Producers of Cytotoxic Metabolites. Marine Drugs, 15(10), 329. https://doi.org/10.3390/md15100329