Degradation of Polysaccharides from Grateloupia filicina and Their Antiviral Activity to Avian Leucosis Virus Subgroup J

 ,
, 

Abstract
:1. Introduction
2. Results
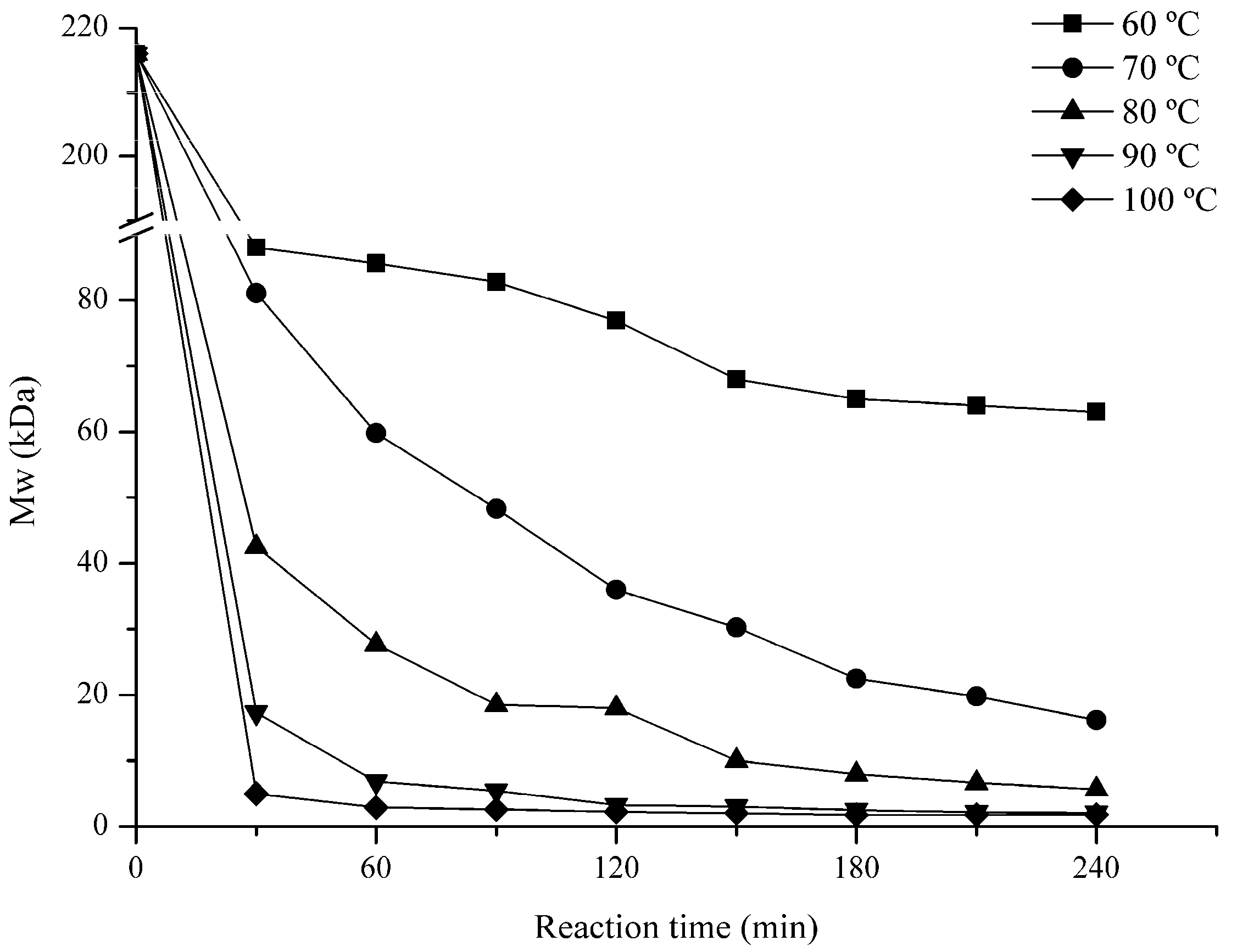
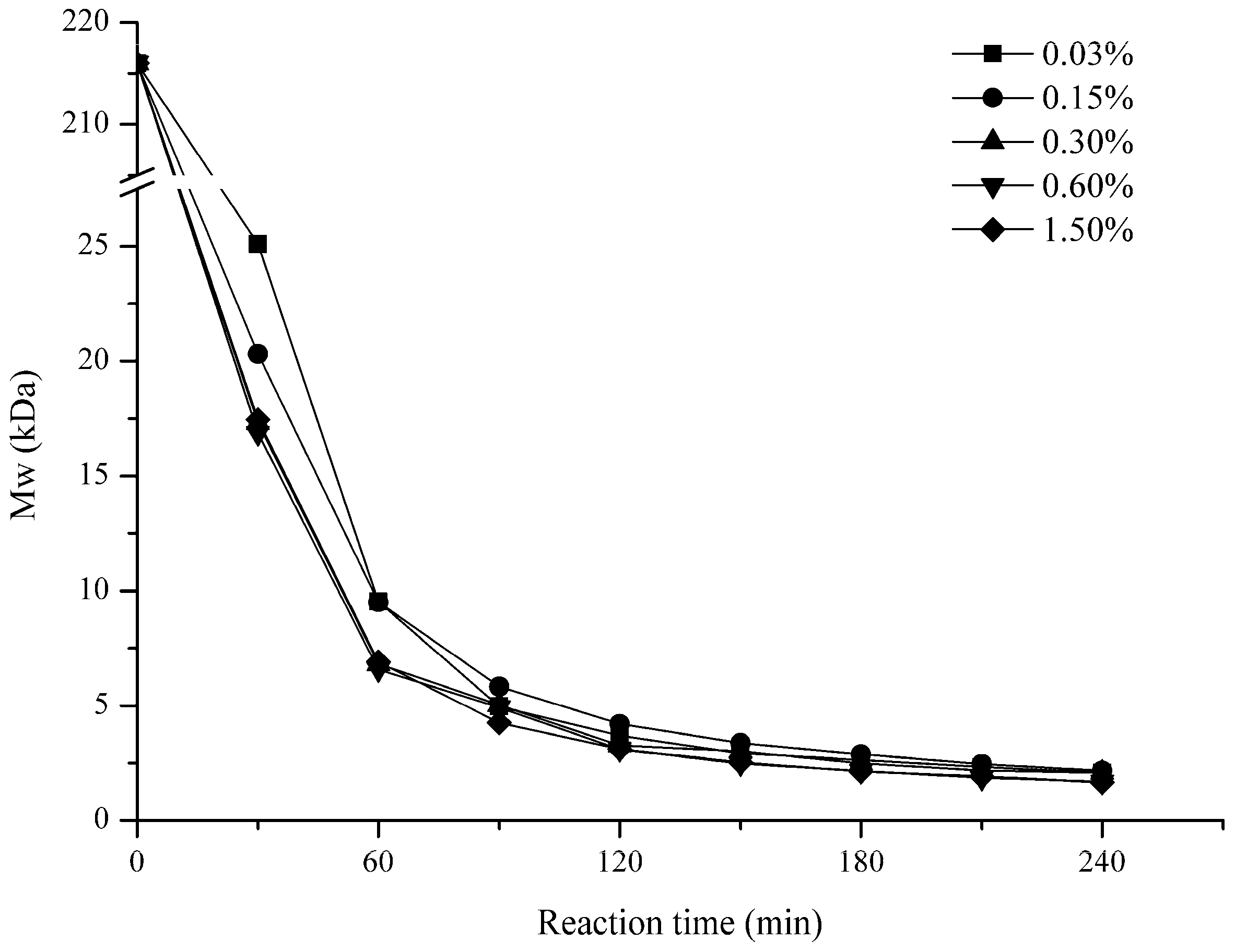
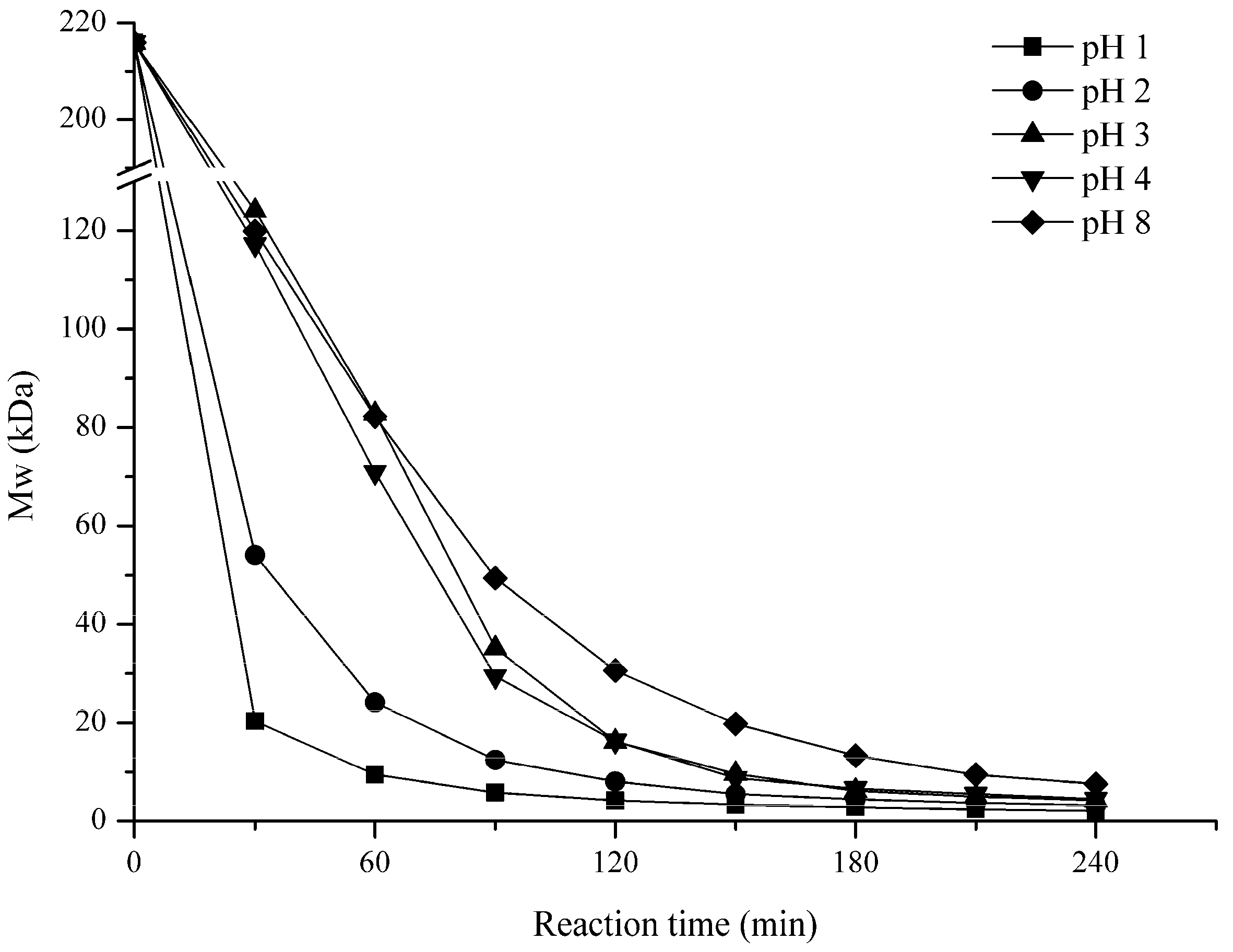
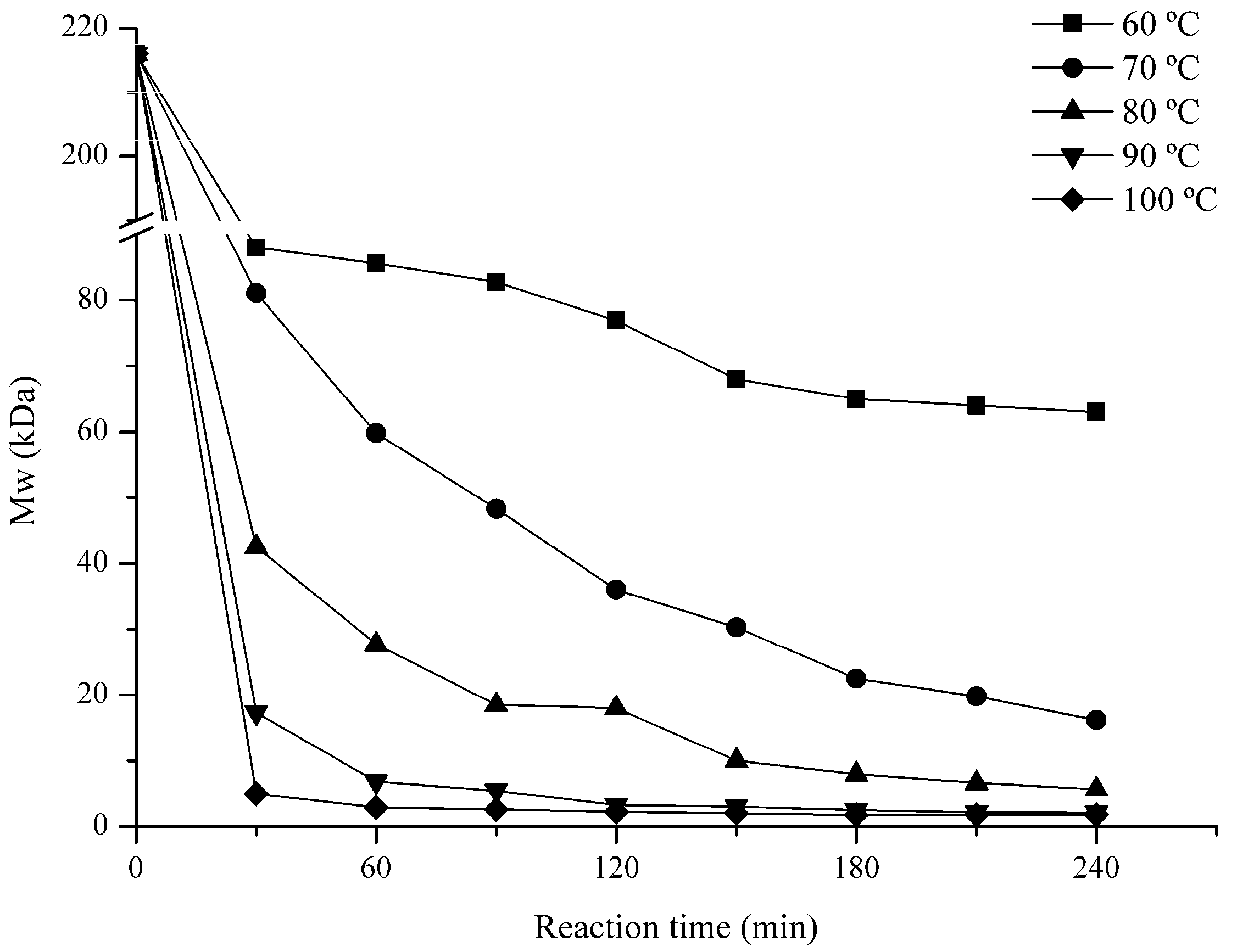
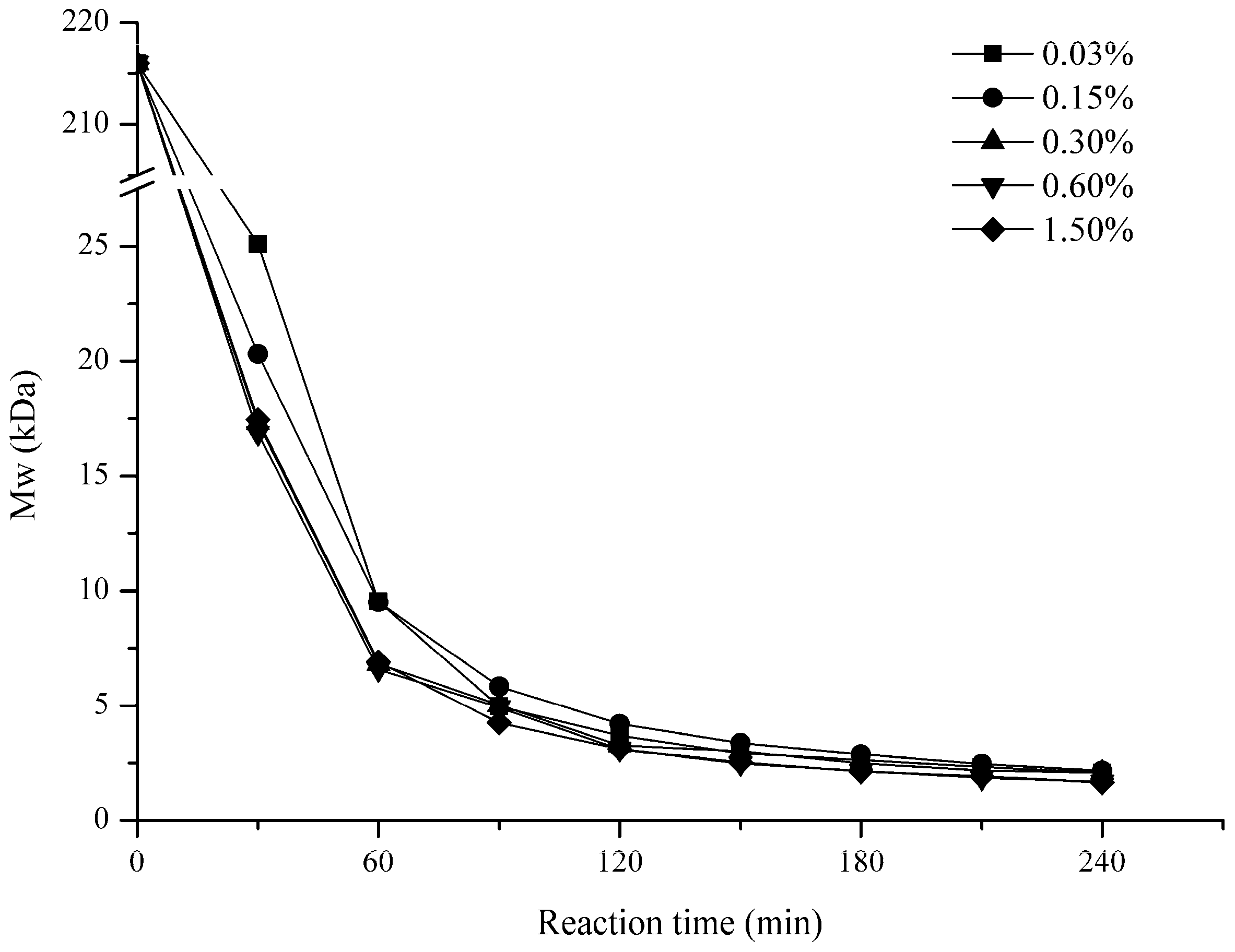
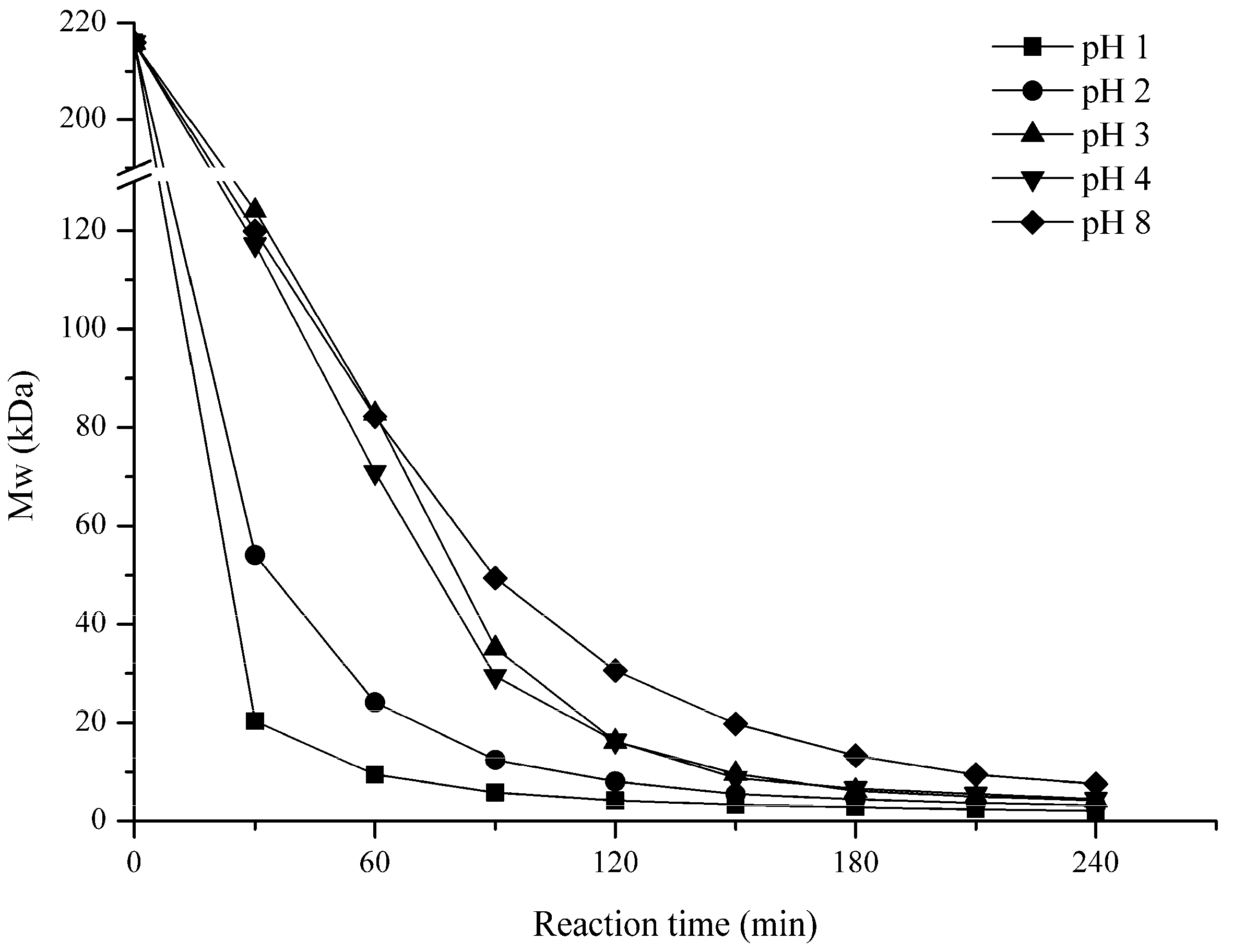
2.1. Influence of Degradation Conditions on the Mws of Polysaccharides
2.1.1. Effect of Temperature
2.1.2. Effect of H2O2 Concentration
2.1.3. Effect of pH
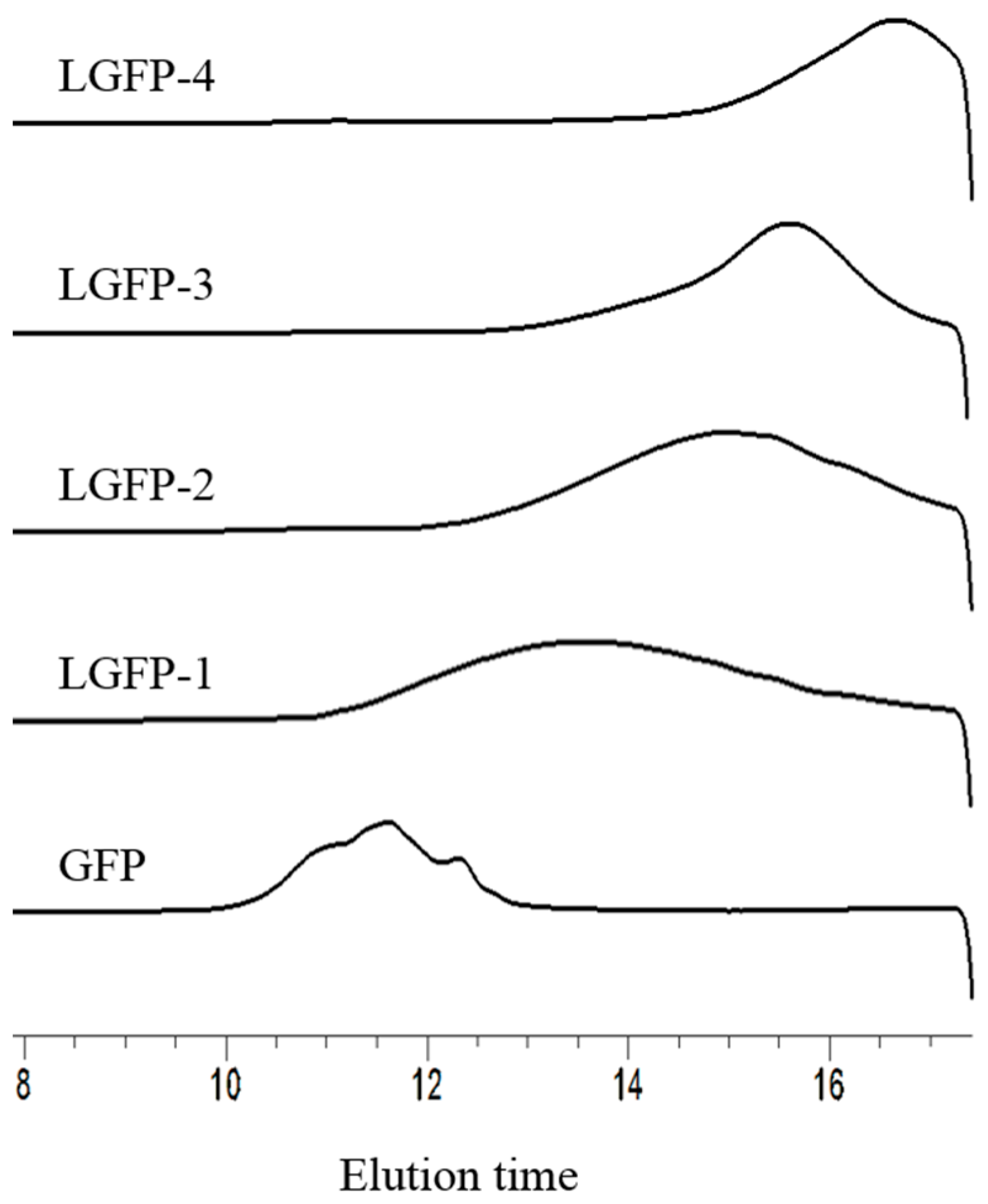
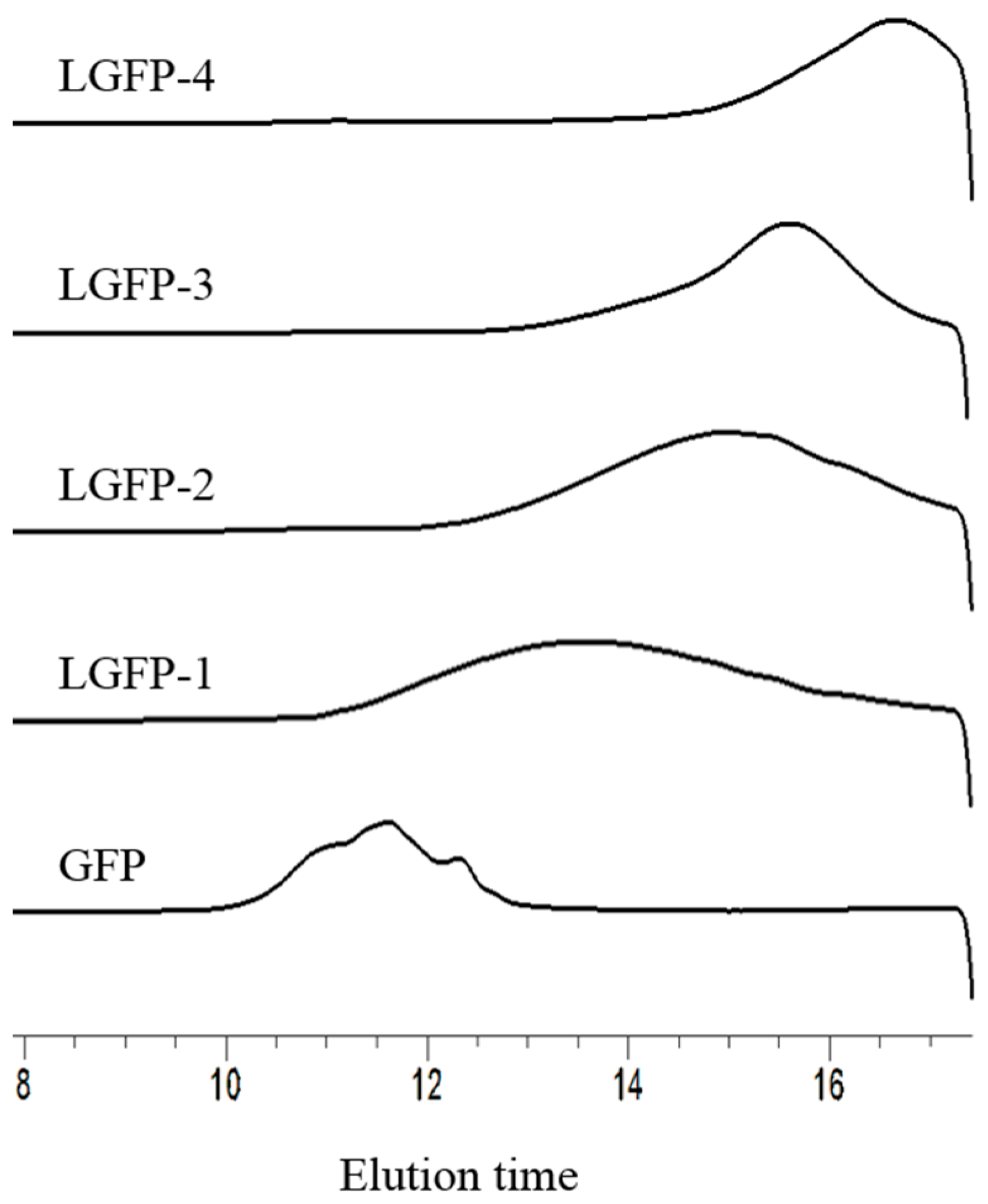
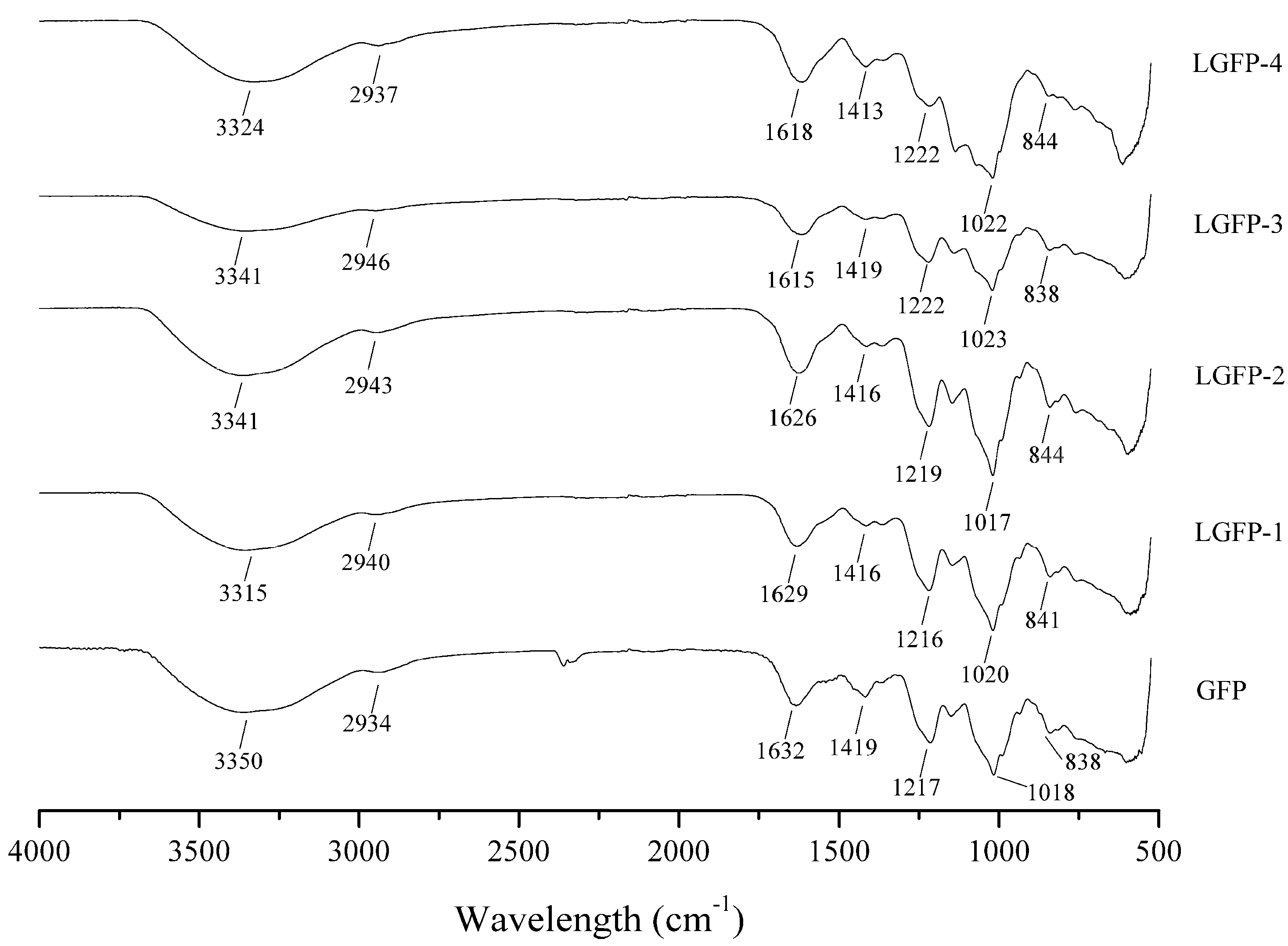
2.2. Preparation and Chemical Characterization of LGFP
2.2.1. Preparation of LGFP
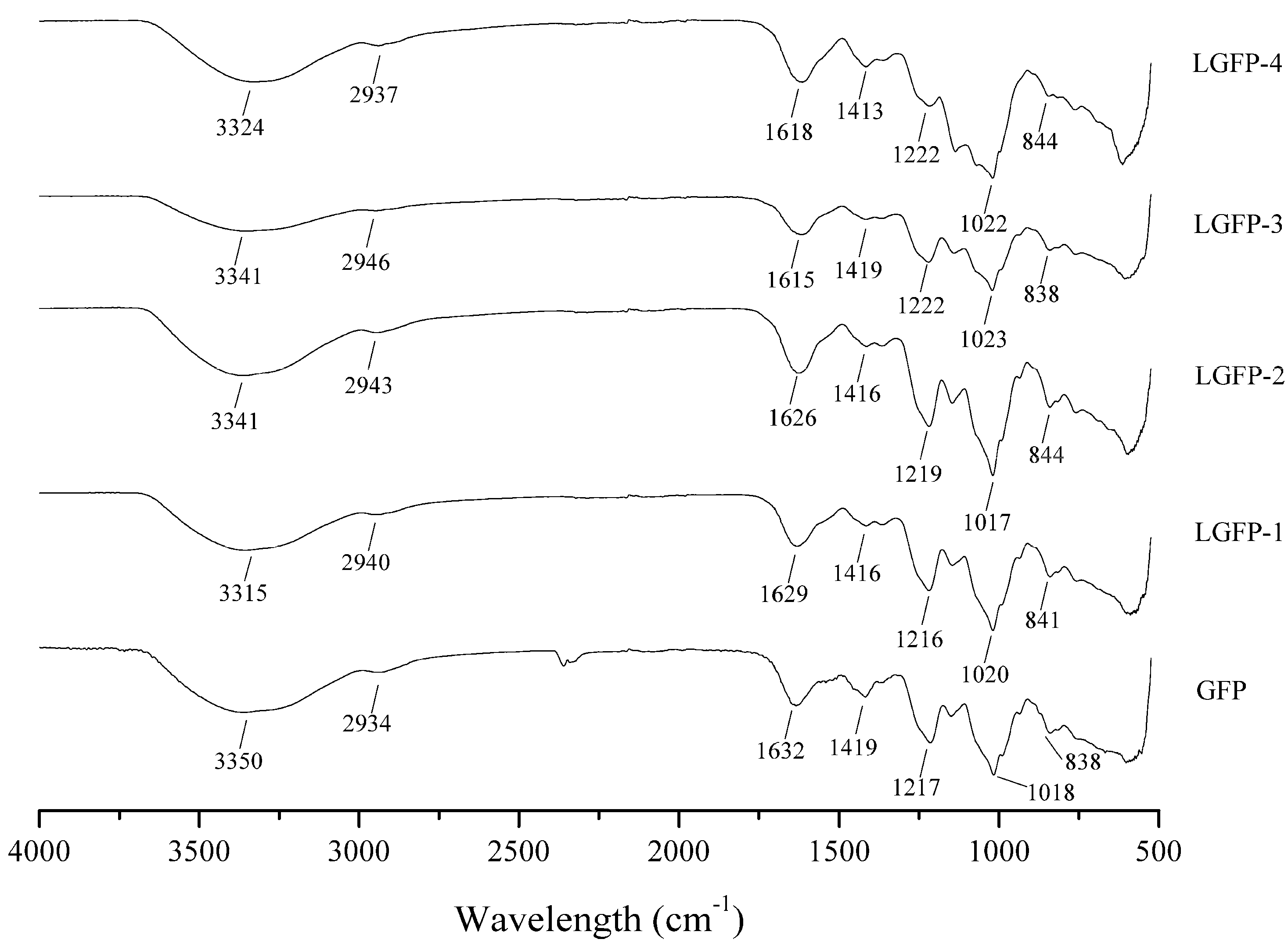
2.2.2. Chemical Characterization
2.3. Cytotoxic Activity
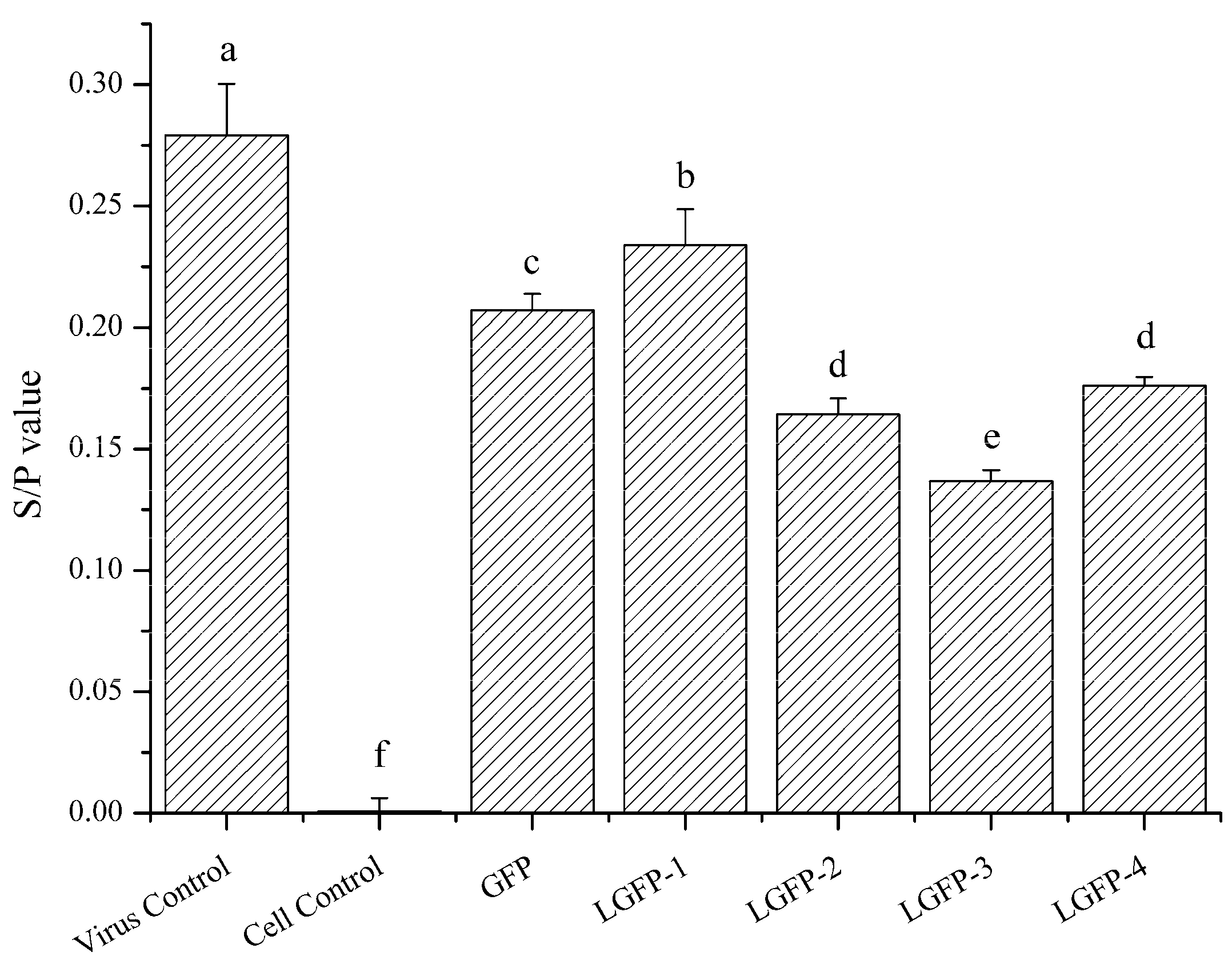
2.4. Anti-ALV-J Activity In Vitro
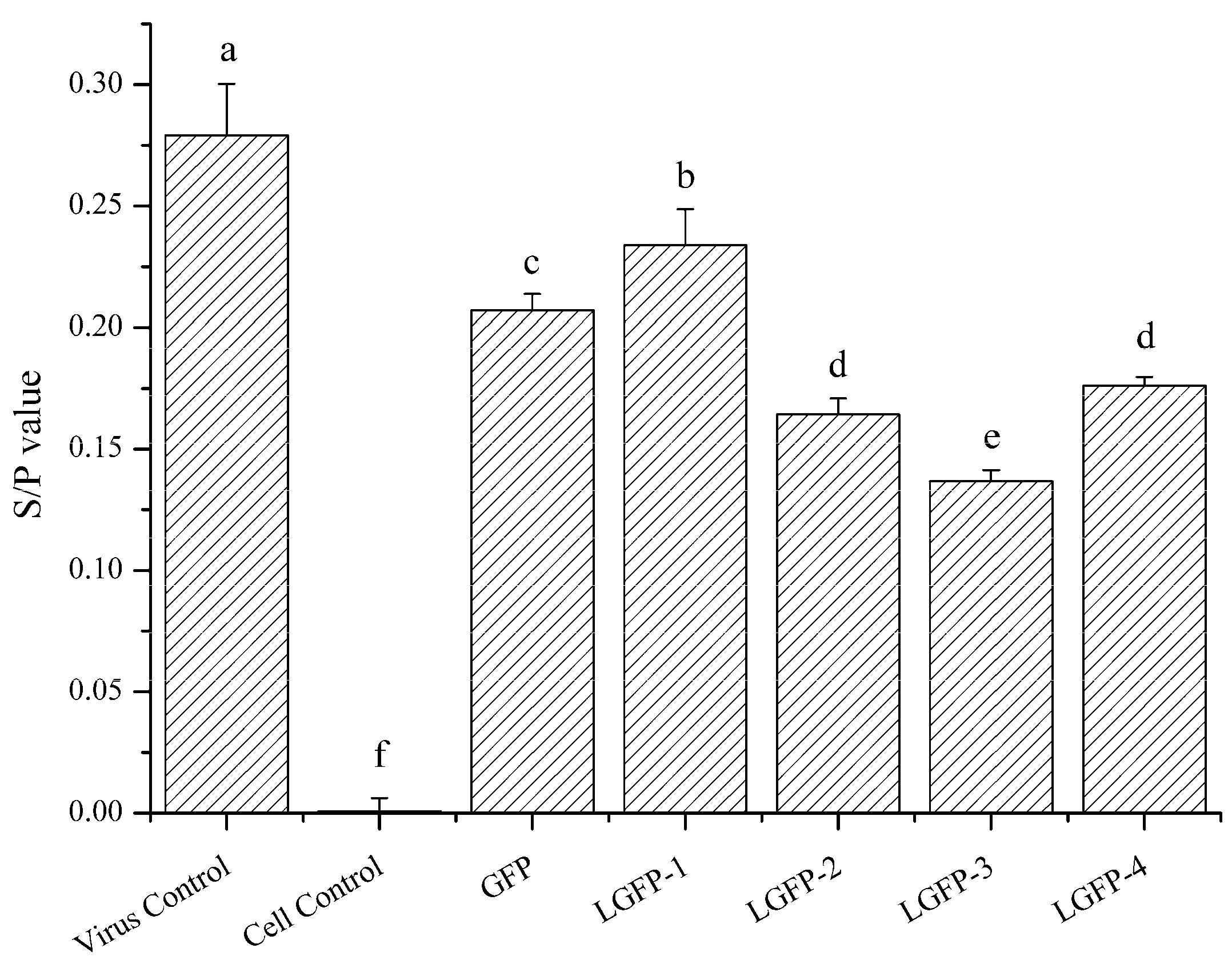
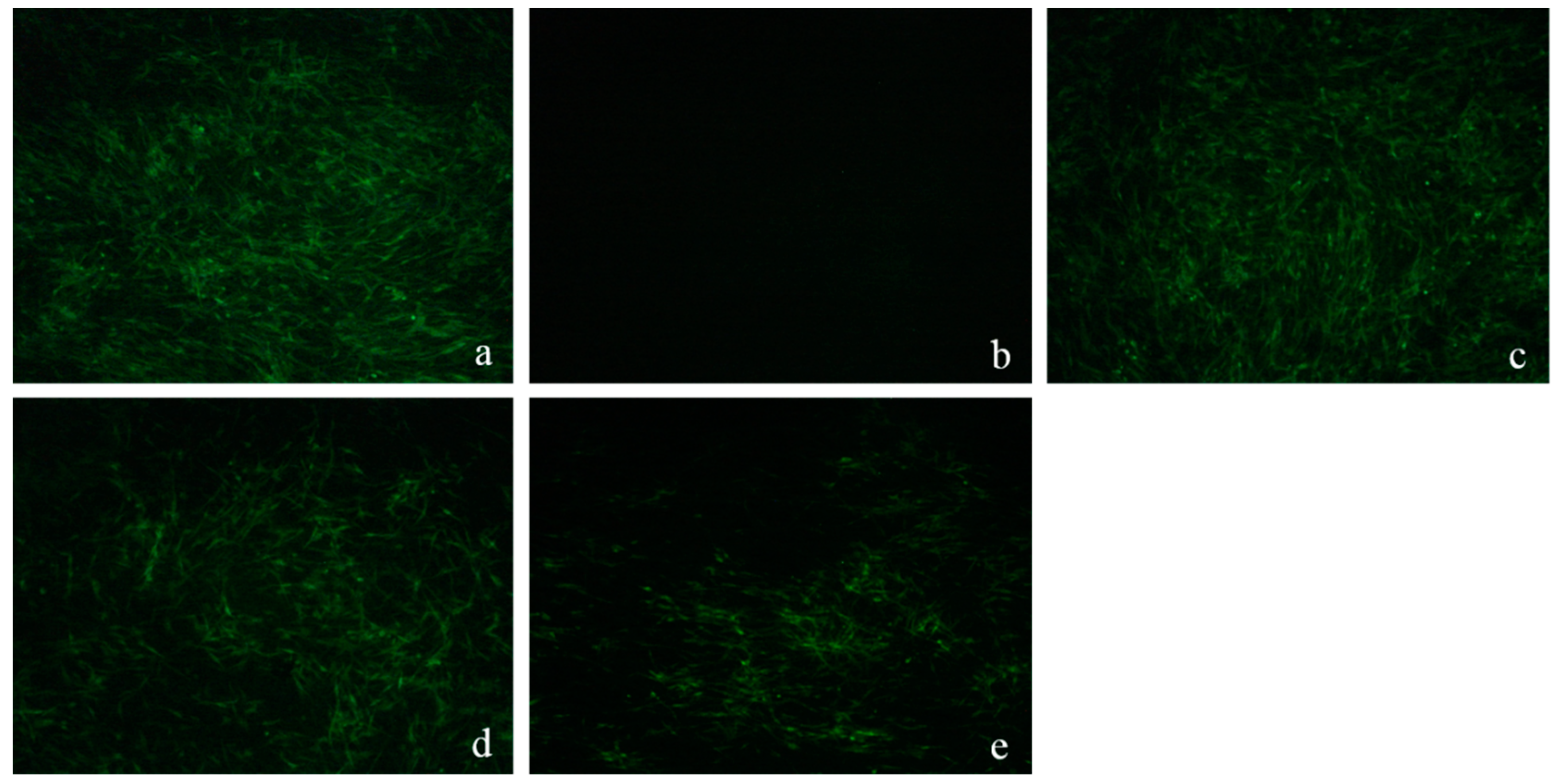
2.4.1. ALV-Specific Antigen Detection
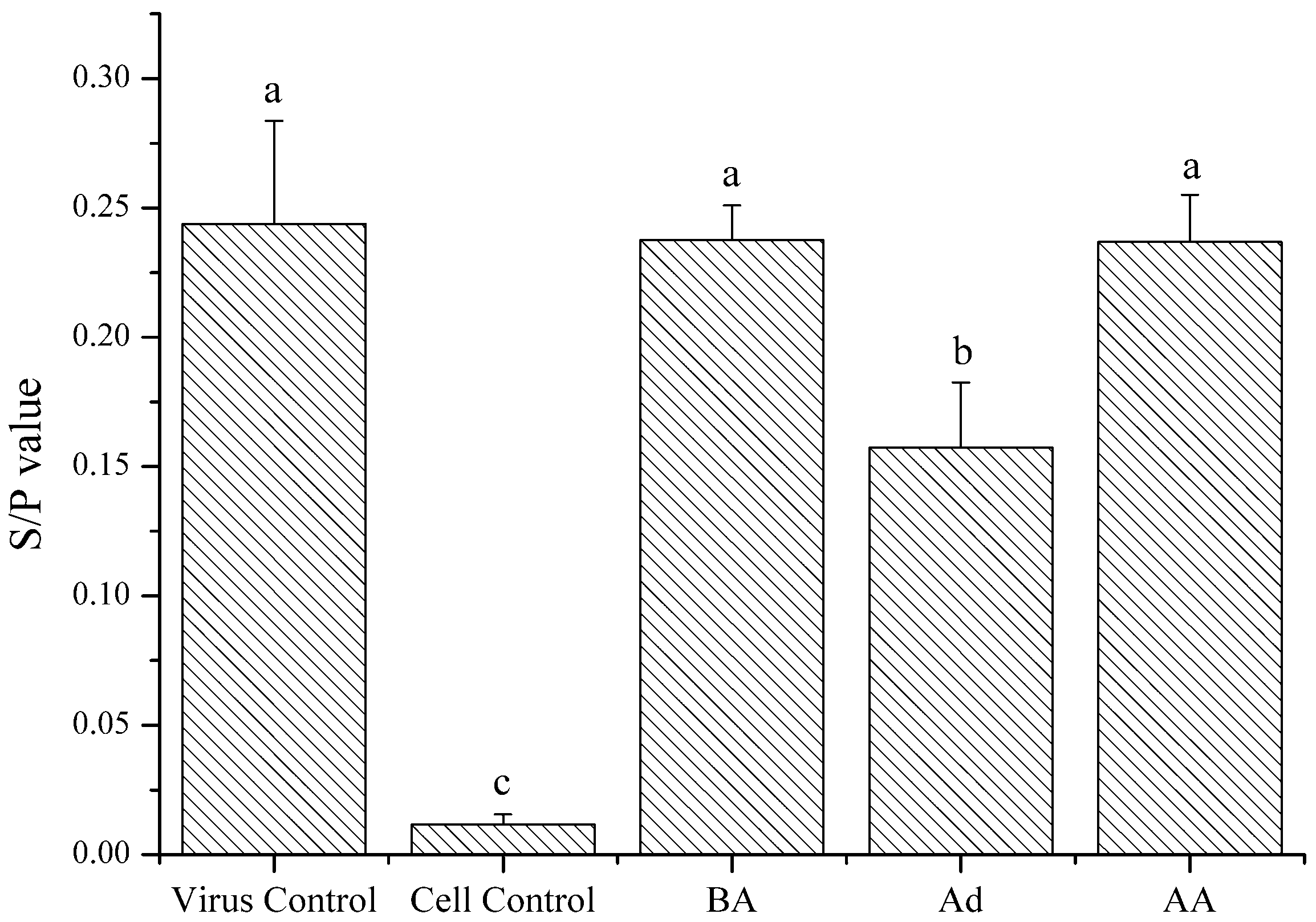
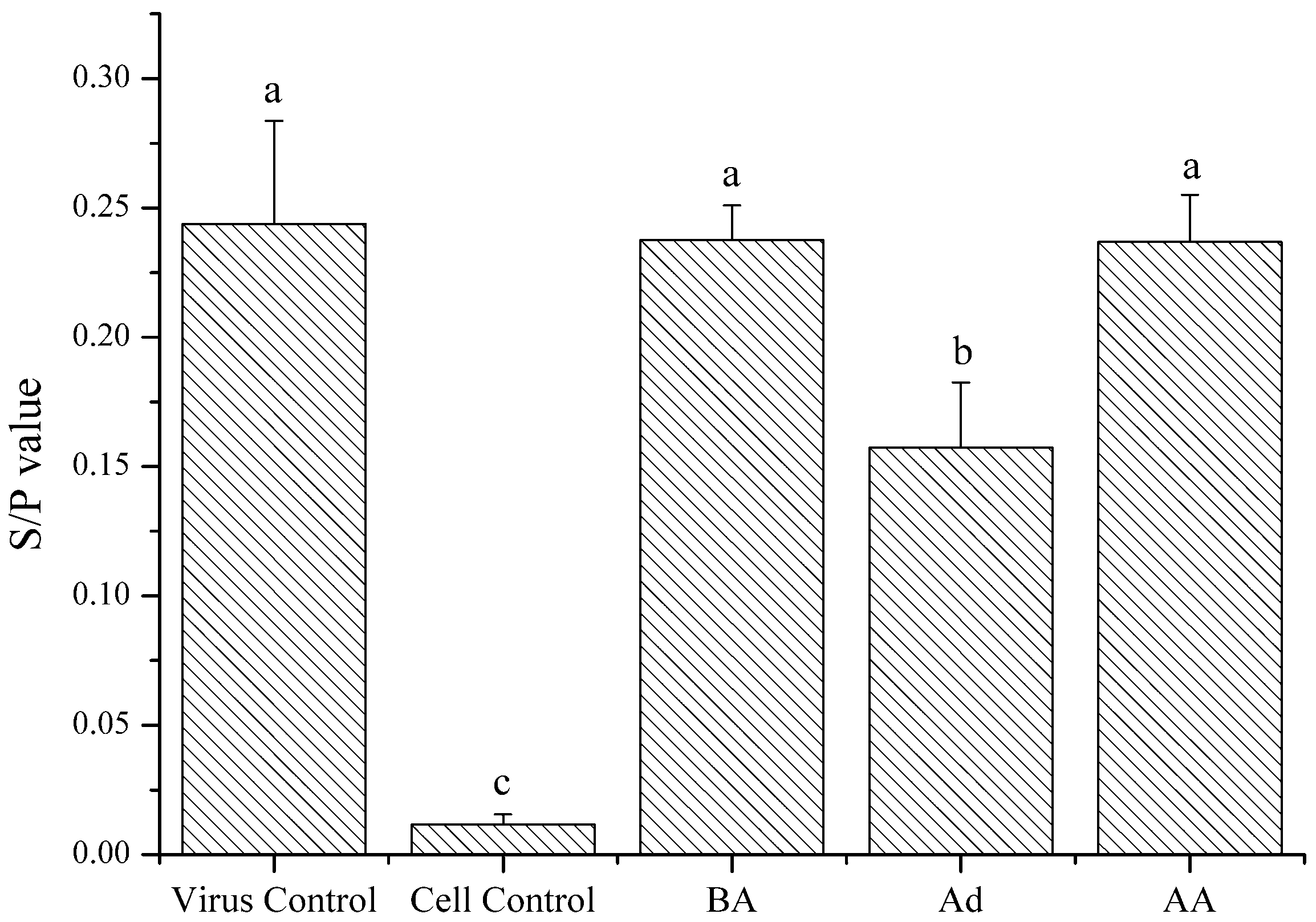
2.4.2. Action Phase of Polysaccharides
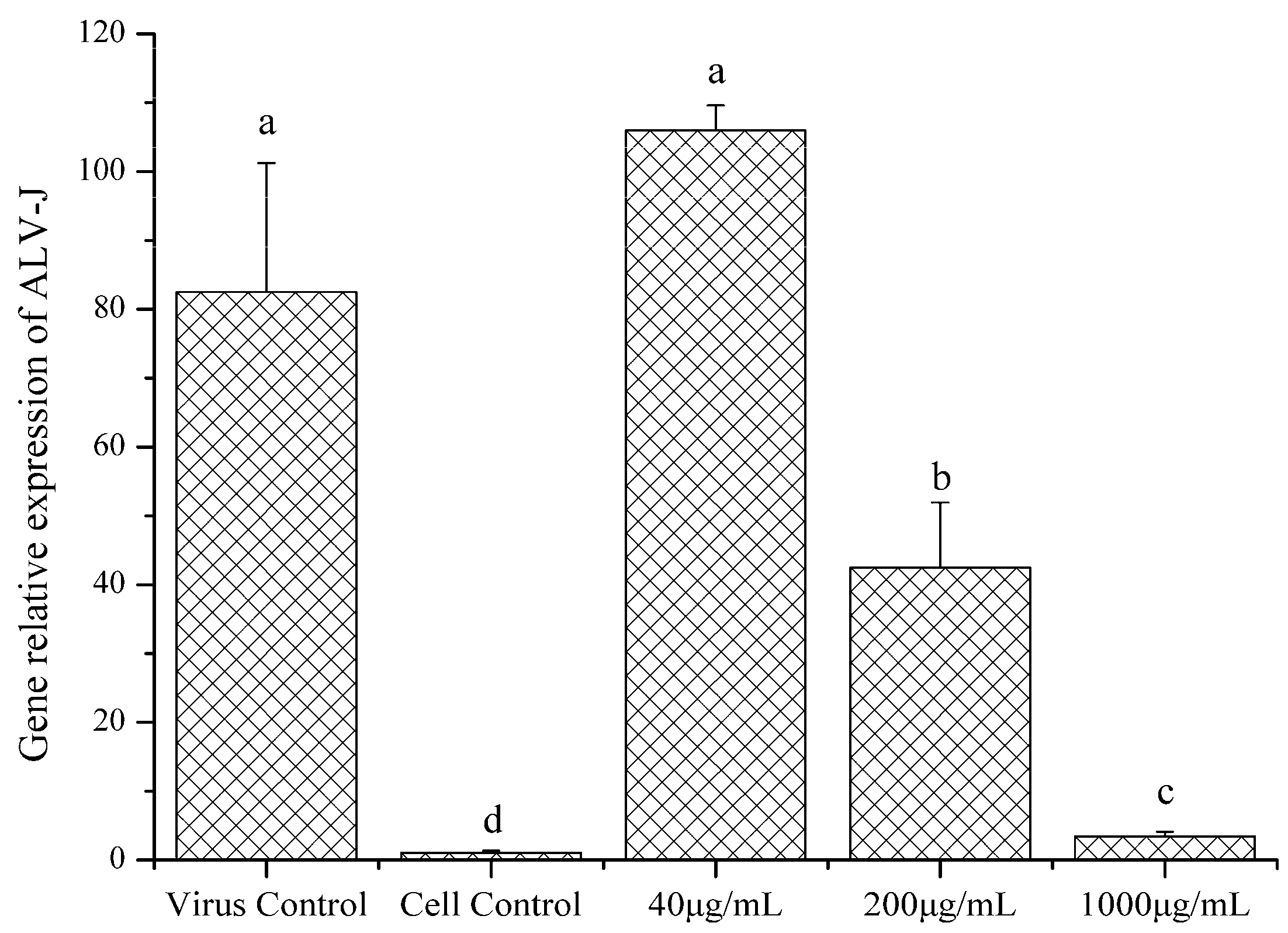
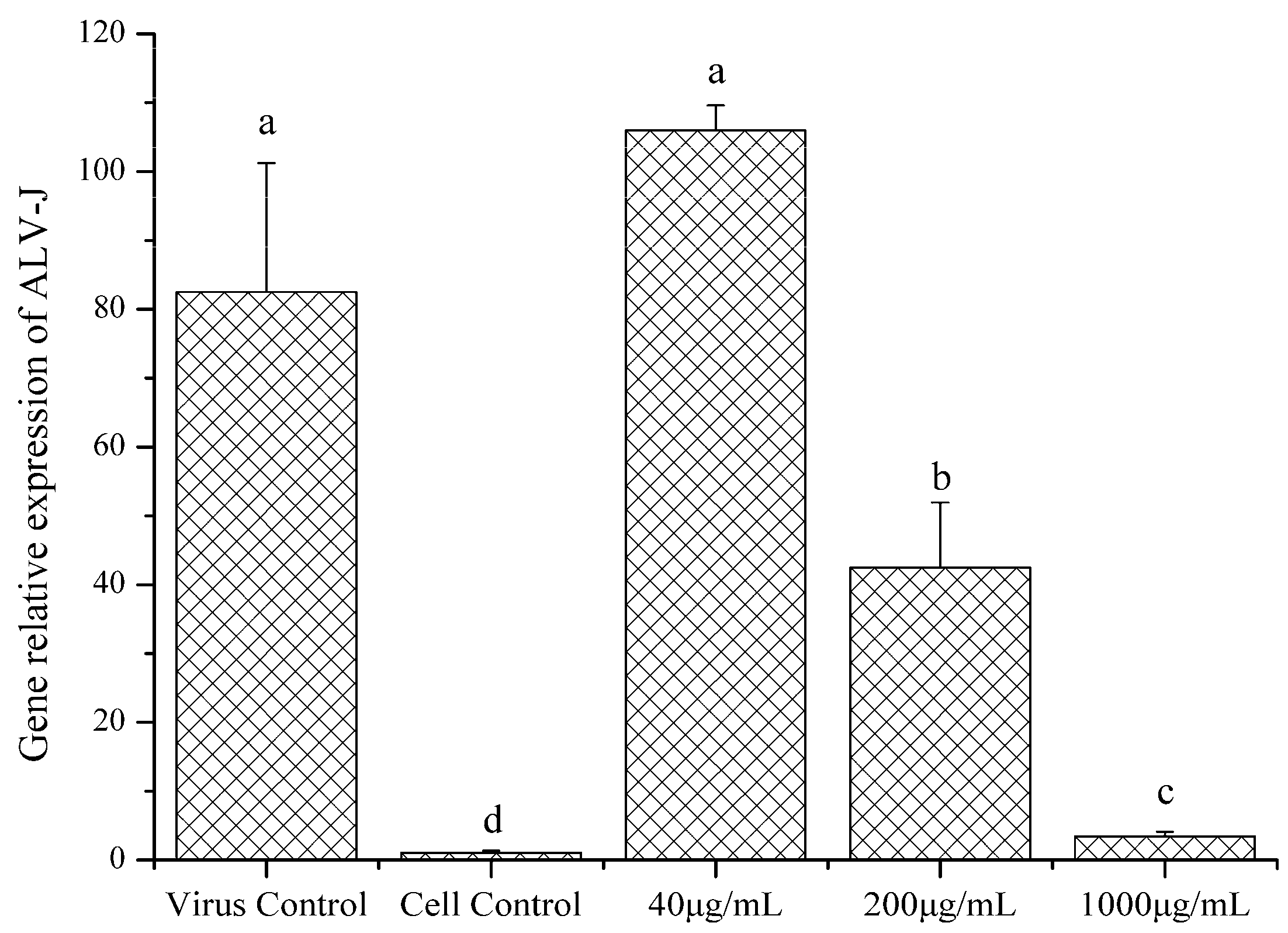
2.4.3. Gene Relative Expression of ALV-J
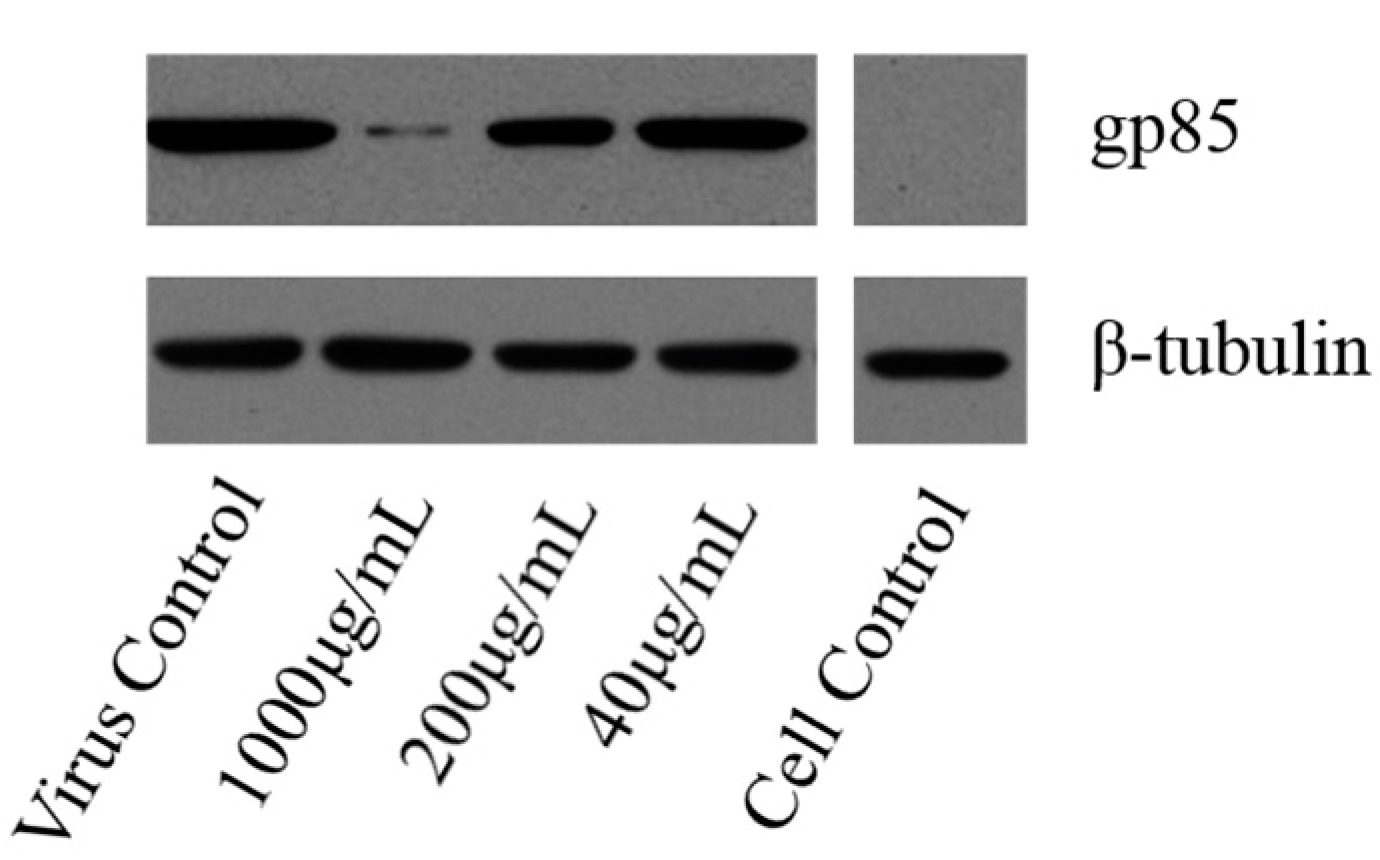
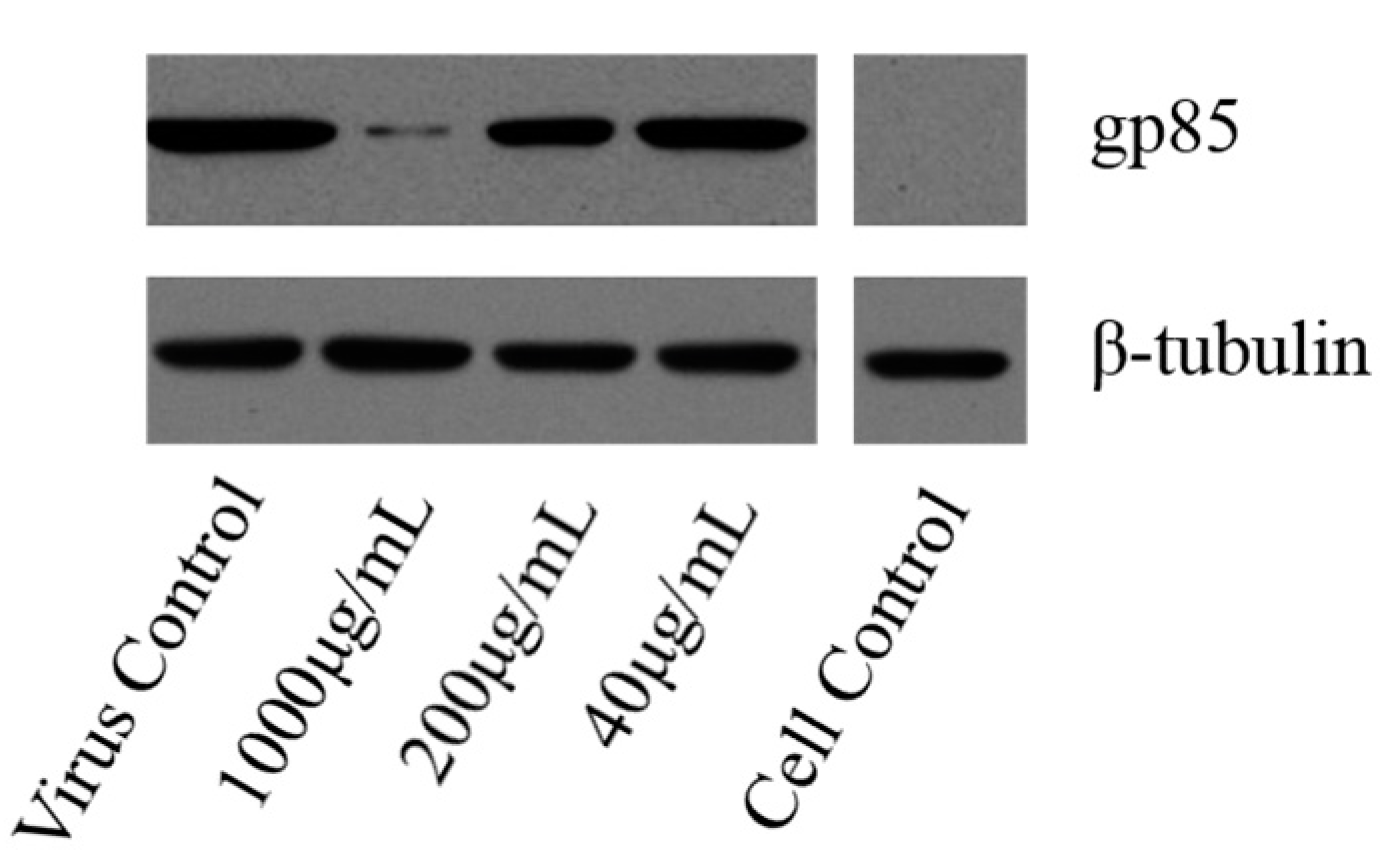
2.4.4. Western-Blot and Indirect Immunofluorescence Assay (IFA) Analysis
3. Discussion
4. Materials and Methods
4.1. Seaweeds Samples and Reagents
4.2. Extraction of GFP
4.3. Influence of H2O2 Oxidation on GFP Degradation and Preparation of LGFP
4.4. Chemical Characterization
4.5. Cell Lines, Cell Culture, Virus and Antibodies
4.6. Determination of Cytotoxic Activity
4.7. Anti-ALV-J Activity In Vitro
4.7.1. ALV Specific Antigen Detection
4.7.2. Detection of the Polysaccharides Action Phase
4.7.3. Viral Gene Relative Expression and Protein Expression
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yu, C.L.; Wei, K.; Liu, L.P.; Yang, S.F.; Hu, L.P.; Zhao, P.; Meng, X.Y.; Shao, M.X.; Wang, C.W.; Zhu, L.J.; et al. Taishan Pinus massoniana pollen polysaccharide inhibits subgroup J avian leucosis virus infection by directly blocking virus infection and improving immunity. Sci. Rep. 2017, 7, 44353. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wen, S.; Wang, S.; Hao, P.; Cheng, Z.; Liu, Y.; Zhao, P.; Liu, J. gp85 protein vaccine adjuvanted with silica nanoparticles against ALV-J in chickens. Vaccine 2017, 35, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.F.; Fadly, A.M.; Hunt, H.D. Hypervariability in the envelope genes of subgroup J avian leukosis viruses obtained from different farms in the United States. Virology 2000, 272, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Dou, W.W.; Li, H.M.; Cheng, Z.Q.; Zhao, P.; Liu, J.Z.; Cui, Z.Z.; Liu, H.G.; Jing, W.F.; Guo, H.J. Maternal antibody induced by recombinant gp85 protein vaccine adjuvanted with CpG-ODN protects against ALV-J early infection in chickens. Vaccine 2013, 31, 6144–6149. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Dong, J.; Waterhouse, G.I. N.; Cheng, Z.; Ai, S. Electrochemical immunosensor with nanocellulose-Au composite assisted multiple signal amplification for detection of avian leukosis virus subgroup J. Biosens. Bioelectron. 2018, 101 (Suppl. C), 110–115. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.F.; Cui, Z.Z. Evolution of gp85 gene of subgroup J avian leukosis virus under the selective pressure of antibodies. Sci. China Ser. C-Life Sci. 2006, 49, 227–234. [Google Scholar] [CrossRef]
- De Clerck, O.; Gavio, B.; Fredericq, S.; Barbara, I.; Coppejans, E. Systematics of Grateloupia filicina (Halymeniaceae, Rhodophyta), based on rbcL sequence analyses and morphological evidence, including the reinstatement of G. minima and the description of G. capensis sp. nov. J. Phycol. 2005, 41, 391–410. [Google Scholar]
- Denis, C.; Morancais, M.; Gaudin, P.; Fleurence, J. Effect of enzymatic digestion on thallus degradation and extraction of hydrosoluble compounds from Grateloupia turuturu. Bot. Mar. 2009, 52, 262–267. [Google Scholar] [CrossRef]
- Nikapitiya, C.; De Zoysa, M.; Jeon, Y.J.; Lee, J. Isolation of sulfated anticoagulant compound from fermented red seaweed Grateloupia filicina. J. World Aquac. Soc. 2007, 38, 407–417. [Google Scholar] [CrossRef]
- Yu, Q.; Yan, J.; Wang, S.; Ji, L.; Ding, K.; Vella, C.; Wang, Z.; Hu, Z. Antiangiogenic effects of GFP08, an agaran-type polysaccharide isolated from Grateloupia filicina. Glycobiology 2012, 22, 1343–1352. [Google Scholar] [CrossRef] [PubMed]
- Ye, D.Y.; Jiang, Z.B.; Zheng, F.C.; Wang, H.M.; Zhang, Y.M.; Gao, F.F.; Chen, P.H.; Chen, Y.C.; Shi, G.G. Optimized extraction of polysaccharides from Grateloupia livida (Harv.) Yamada and biological activities. Molecules 2015, 20, 16817–16832. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.B.; Chen, Y.C.; Yao, F.; Chen, W.Z.; Zhong, S.P.; Zheng, F.C.; Shi, G.G. Antioxidant, antibacterial and antischistosomal activities of extracts from Grateloupia livida (Harv). Yamada. PLoS ONE 2013, 8, e80413. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, K.; Mateu, C.G.; Mandal, P.; Pujol, C.A.; Damonte, E.B.; Ray, B. Galactan sulfate of Grateloupia indica: Isolation, structural features and antiviral activity. Phytochemistry 2007, 68, 1428–1435. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Bligh, S.W.; Shi, S.S.; Wang, Z.T.; Hu, Z.B.; Crowder, J.; Branford-White, C.; Vella, C. Structural features and anti-HIV-1 activity of novel polysaccharides from red algae Grateloupia longifolia and Grateloupia filicina. Int. J. Biol. Macromol. 2007, 41, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Ramnani, P.; Chitarrari, R.; Tuohy, K.; Grant, J.; Hotchkiss, S.; Philp, K.; Campbell, R.; Gill, C.; Rowland, I. In vitro fermentation and prebiotic potential of novel low molecular weight polysaccharides derived from agar and alginate seaweeds. Anaerobe 2012, 18, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.Q.; Wang, L.; Zhou, Y. Immunomodulation and antitumor activities of different-molecular-weight polysaccharides from Porphyridium cruentum. Carbohydr. Polym. 2012, 87, 1206–1210. [Google Scholar] [CrossRef]
- Li, B.; Liu, S.; Xing, R.; Li, K.; Li, R.; Qin, Y.; Wang, X.; Wei, Z.; Li, P. Degradation of sulfated polysaccharides from Enteromorpha prolifera and their antioxidant activities. Carbohydr. Polym. 2013, 92, 1991–1996. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Guo, F.; Hu, J.; Zhang, L.; Xue, C.; Zhang, Z.; Li, B. Antithrombotic activity of oral administered low molecular weight fucoidan from Laminaria Japonica. Thromb. Res. 2016, 144, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.X.; Chen, F.; Li, F. Preparation and potential in vivo anti-influenza virus activity of low molecular-weight kappa-carrageenans and their derivatives. J. Appl. Polym. Sci. 2013, 127, 2110–2115. [Google Scholar] [CrossRef]
- Yamada, T.; Ogamo, A.; Saito, T.; Uchiyama, H.; Nakagawa, Y. Preparation of O-acylated low-molecular-weight carrageenans with potent anti-HIV activity and low anticoagulant effect. Carbohydr. Polym. 2000, 41, 115–120. [Google Scholar] [CrossRef]
- Lee, J.B.; Takeshita, A.; Hayashi, K.; Hayashi, T. Structures and antiviral activities of polysaccharides from Sargassum trichophyllum. Carbohydr. Polym. 2011, 86, 995–999. [Google Scholar] [CrossRef]
- Kim, W.J.; Choi, J.W.; Jang, W.J.; Kang, Y.S.; Lee, C.W.; Synytsya, A.; Park, Y.I. Low-molecular weight mannogalactofucans prevent herpes simplex virus type 1 infection via activation of Toll-like receptor 2. Int. J. Biol. Macromol. 2017, 103, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, H.M.; Qin, G.Y. Structure characterization and antioxidant activity of polysaccharides from Chinese quince seed meal. Food Chem. 2017, 234, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Kolsi, R.B.; Ben Salah, H.; Jardak, N.; Chaaben, R.; Jribi, I.; El Feki, A.; Rebai, T.; Jamoussi, K.; Allouche, N.; Blecker, C.; et al. Sulphated polysaccharide isolated from Sargassum vulgare: Characterization and hypolipidemic effects. Carbohydr. Polym. 2017, 170, 148–159. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Lee, B.-J.; Kim, Y.A.; Lee, H.-J.; Joo, Y.H. Effects of several salt marsh plants on mouse spleen and thymus cell proliferation using MTT assay. Ocean Sci. J. 2005, 40, 209–212. [Google Scholar] [CrossRef]
- Song, L.; Chen, X.L.; Liu, X.D.; Zhang, F.B.; Hu, L.F.; Yue, Y.; Li, K.C.; Li, P.C. Characterization and comparison of the structural features, immune-modulatory and anti-avian influenza virus activities conferred by three algal sulfated polysaccharides. Mar. Drugs 2016, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, J.D.; Zhang, X.S.; Hao, C.; Zhao, X.L.; Jiao, G.L.; Shan, X.D.; Tai, W.J.; Yu, G.L. Inhibition of influenza a virus infection by fucoidan targeting viral neuraminidase and cellular EGFR pathway. Sci. Rep. 2017, 7, 10760. [Google Scholar] [CrossRef] [PubMed]
- Bouhlal, R.; Haslin, C.; Chermann, J.-C.; Colliec-Jouault, S.; Sinquin, C.; Simon, G.; Cerantola, S.; Riadi, H.; Bourgougnon, N. Antiviral activities of sulfated polysaccharides isolated from Sphaerococcus coronopifolius (Rhodophytha, Gigartinales) and Boergeseniella thuyoides (Rhodophyta, Ceramiales). Mar. Drugs 2011, 9, 1187–1209. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, S.; Wang, J.; Song, L.; Xing, R.; Liu, S.; Yu, H.; Li, P. Sulfated polysaccharides isolated from cloned Grateloupia filicina and their anticoagulant activity. BioMed Res. Int. 2015, 2015, 612352. [Google Scholar] [PubMed]
- Athukorala, Y.; Jung, W.-K.; Park, P.-J.; Lee, Y.-J.; Kim, S.-K.; Vasanthan, T.; No, H.-K.; Jeon, Y.-J. Evaluation of biomolecular interactions of sulfated polysaccharide isolated from Grateloupia filicina on blood coagulation factors. J. Microbiol. Biotechnol. 2008, 18, 503–511. [Google Scholar] [PubMed]
- Qin, C.Q.; Du, Y.M.; Xiao, L. Effect of hydrogen peroxide treatment on the molecular weight and structure of chitosan. Polym. Degrad. Stabil. 2002, 76, 211–218. [Google Scholar] [CrossRef]
- Parovuori, P.; Hamunen, A.; Forssell, P.; Autio, K.; Poutanen, K. Oxidation of potato starch by hydrogen-peroxide. Starch-Starke 1995, 47, 19–23. [Google Scholar] [CrossRef]
- Zeronian, S.H.; Inglesby, M.K. Bleaching of cellulose by hydrogen peroxide. Cellulose 1995, 2, 265–272. [Google Scholar] [CrossRef]
- Hien, N.Q.; Phu, D.V.; Duy, N.N.; Nguyen, T.K.L. Degradation of chitosan in solution by gamma irradiation in the presence of hydrogen peroxide. Carbohydr. Polym. 2012, 87, 935–938. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, J.; Jin, W.H.; Zhang, H.; Zhang, Q.B. Degradation of Laminaria japonica fucoidan by hydrogen peroxide and antioxidant activities of the degradation products of different molecular weights. Carbohydr. Polym. 2012, 87, 153–159. [Google Scholar] [CrossRef]
- Iqbal, S.; Marchetti, R.; Aman, A.; Silipo, A.; Ul Qader, S.A.; Molinaro, A. Enzymatic and acidic degradation of high molecular weight dextran into low molecular weight and its characterizations using novel Diffusion-ordered NMR spectroscopy. Int. J. Biol. Macromol. 2017, 103, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.R.; Dong, W.X.; Yu, C.M.; He, Z.Q.; Lv, Y.L.; Sun, Y.H.; Feng, X.Y.; Li, N.; Lee, L.F.; Li, M. Occurrence of avian leukosis virus subgroup J in commercial layer flocks in China. Avian Pathol. 2004, 33, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.Q.; Zhang, L.; Liu, S.D.; Zhang, L.J.; Cui, Z.Z. Emerging of fibro subgroup J in a flock of Chinese local breed. Acta Microbiol. Sin. 2005, 45, 584–587. [Google Scholar]
- Sun, S.H.; Cui, Z.Z. Epidemiological and pathological studies of subgroup J avian leukosis virus infections in Chinese local “yellow” chickens. Avian Pathol. 2007, 36, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, K.; Smith, L.M.; Howes, K.; Payne, L.N. Antigenic variants of J subgroup avian leukosis virus: sequence analysis reveals multiple changes in the env gene. J. Gen. Virol. 1998, 79, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Zavala, G.; Cheng, S.; Jackwood, M.W. Molecular epidemiology of avian leukosis virus subgroup j and evolutionary history of its 3′ untranslated region. Avian Dis. Dig. 2007, 51, 942–953. [Google Scholar] [CrossRef]
- Cao, Y.G.; Hao, Y.; Li, Z.H.; Liu, S.T.; Wang, L.X. Antiviral activity of polysaccharide extract from Laminaria japonica against respiratory syncytial virus. Biomed. Pharmacother. 2016, 84, 1705–1710. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Avila, L.M.; Elizondo-Gonzalez, R.; Rodriguez-Santillan, P.; Aguilar-Briseno, J.A.; Ricque-Marie, D.; Rodriguez-Padilla, C.; Cruz-Suarez, L.E. Innocuity and anti-Newcastle-virus-activity of Cladosiphon okamuranus fucoidan in chicken embryos. Poult. Sci. 2016, 95, 2795–2802. [Google Scholar] [CrossRef] [PubMed]
- Gheda, S.F.; El-Adawi, H.I.; El-Deeb, N.M. Antiviral profile of brown and red seaweed polysaccharides against hepatitis C virus. Iran. J. Pharm. Res. 2016, 15, 483–491. [Google Scholar] [PubMed]
- Bourgougnon, N.; Lahaye, M.; Quemener, B.; Chermann, J.C.; Rimbert, M.; Cormaci, M.; Furnari, G.; Kornprobst, J.M. Annual variation in composition and in vitro anti-HIV-1 activity of the sulfated glucuronogalactan from Schizymenia dubyi (Rhodophyta, Gigartinales). J. Appl. Phycol. 1996, 8, 155–161. [Google Scholar] [CrossRef]
- Chen, Y.; Song, M.Y.; Wang, Y.X.; Xiong, W.; Zeng, L.; Zhang, S.B.; Xu, M.Y.; Du, H.X.; Liu, J.G.; Wang, D.Y.; et al. The anti-DHAV activities of Astragalus polysaccharide and its sulfate compared with those of BSRPS and its sulfate. Carbohydr. Polym. 2015, 117, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Ji, Y.B.; Qi, Z.; Cui, D.; Xin, G.S.; Wang, B.; Cao, Y.; Wang, D.D. Anti-tumor activity of sulfated polysaccharides from Sargassum fusiforme. Saudi Pharm. J. 2017, 25, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.H.; Wang, Z.J.; Shen, M.Y.; Nie, S.P.; Gong, B.; Li, H.S.; Zhao, Q.; Li, W.J.; Xie, M.Y. Sulfated modification, characterization and antioxidant activities of polysaccharide from Cyclocarya paliurus. Food Hydrocoll. 2016, 53, 7–15. [Google Scholar] [CrossRef]
- Li, J.; Chi, Z.; Yu, L.; Jiang, F.; Liu, C. Sulfated modification, characterization, and antioxidant and moisture absorption/retention activities of a soluble neutral polysaccharide from Enteromorpha prolifera. Int. J. Biol. Macromol. 2017, 105, 1544–1553. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kawai, Y.; Seno, N.; Anno, K. A modified method for chondrosulfatase assay. Anal. Biochem. 1969, 32, 314–321. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Q.; Wang, J.; Shi, X.; Zhang, Z. Analysis of the monosaccharide composition of fucoidan by precolumn derivation HPLC. Chin. J. Oceanol. Limn. 2009, 27, 578–582. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Shandiz, S.A.S.; Khosravani, M.; Mohammadi, S.; Noorbazargan, H.; Mirzaie, A.; Inanlou, D.N.; Jalali, M.D.; Jouzaghkar, H.; Baghbani-Arani, F.; Keshavarz-Pakseresht, B. Evaluation of imatinib mesylate (Gleevec) on KAI1/CD82 gene expression in breast cancer MCF-7 cells using quantitative real-time PCR. Asian Pac. J. Trop. Biomed. 2016, 6, 159–163. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Temperature (°C) | pH | H2O2 (%) | Time (min) | Yield (%) |
|---|---|---|---|---|---|
| LGFP-1 | 90 | 4 | 0.3 | 75 | 73.4 |
| LGFP-2 | 90 | 4 | 0.3 | 135 | 46.28 |
| LGFP-3 | 90 | 4 | 0.3 | 210 | 42.99 |
| LGFP-4 | 90 | 4 | 0.3 | 240 | 40.38 |
| Sample | Total Sugar (%) | Sulfate (%) | Protein (%) | Mw (kDa) | Monosaccharides Composition (Molar Ratio) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Man | Rha | Glc A | Glc | Gal | Xyl | Fuc | |||||
| GFP | 55.22 ± 0.96 | 21.52 ± 0.04 | 1.45 ± 0.08 | 216.7 ± 1.2 | 0.00 | 0.01 | 0.03 | 0.01 | 1 | 0.03 | 0.03 |
| LGFP-1 | 49.74 ± 1.22 | 20.99 ± 0.28 | 0.96 ± 0.01 | 40.2 ± 0.3 | 0.01 | 0.01 | 0.03 | 0.01 | 1 | 0.03 | 0.02 |
| LGFP-2 | 47.21 ± 1.32 | 16.86 ± 0.10 | 0.45 ± 0.02 | 14.0 ± 0.3 | 0.01 | 0.01 | 0.05 | 0.01 | 1 | 0.03 | 0.02 |
| LGFP-3 | 44.73 ± 2.43 | 18.33 ± 0.43 | 0.18 ± 0.01 | 8.7 ± 0.2 | 0.01 | 0.01 | 0.05 | 0.01 | 1 | 0.03 | 0.01 |
| LGFP-4 | 41.00 ± 0.25 | 17.01 ± 0.44 | 0.29 ± 0.00 | 2.7 ± 0.1 | 0.01 | 0.02 | 0.04 | 0.01 | 1 | 0.02 | 0.02 |
| Concentration (mg/mL) | 2 | 1 | 0.5 | 0.25 | 0.125 | 0.0625 | 0.03125 |
|---|---|---|---|---|---|---|---|
| GFP | 0.95 ± 0.02 | 1.00 ± 0.06 | 1.03 ± 0.07 | 1.08 ± 0.01 | 1.07 ± 0.04 | 1.05 ± 0.09 | 1.02 ± 0.06 |
| LGFP-1 | 0.96 ± 0.02 | 1.00 ± 0.05 | 1.05 ± 0.04 | 1.05 ± 0.08 | 1.01 ± 0.04 | 0.99 ± 0.04 | 1.00 ± 0.04 |
| LGFP-2 | 0.9 4± 0.05 | 1.06 ± 0.04 | 1.01 ± 0.02 | 1.05 ± 0.04 | 1.05 ± 0.05 | 1.01 ± 0.04 | 1.03 ± 0.07 |
| LGFP-3 | 0.9 5± 0.02 | 1.02 ± 0.05 | 1.04 ± 0.10 | 0.99 ± 0.03 | 1.03 ± 0.03 | 0.99 ± 0.05 | 1.03 ± 0.04 |
| LGFP-4 | 0.96 ± 0.10 | 0.98 ± 0.03 | 1.03 ± 0.06 | 1.01 ± 0.05 | 1.01 ± 0.03 | 1.04 ± 0.07 | 1.09 ± 0.03 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Chen, X.; Cheng, Z.; Liu, S.; Yu, H.; Wang, X.; Li, P. Degradation of Polysaccharides from Grateloupia filicina and Their Antiviral Activity to Avian Leucosis Virus Subgroup J. Mar. Drugs 2017, 15, 345. https://doi.org/10.3390/md15110345
Sun Y, Chen X, Cheng Z, Liu S, Yu H, Wang X, Li P. Degradation of Polysaccharides from Grateloupia filicina and Their Antiviral Activity to Avian Leucosis Virus Subgroup J. Marine Drugs. 2017; 15(11):345. https://doi.org/10.3390/md15110345
Chicago/Turabian StyleSun, Yuhao, Xiaolin Chen, Ziqiang Cheng, Song Liu, Huahua Yu, Xueqin Wang, and Pengcheng Li. 2017. "Degradation of Polysaccharides from Grateloupia filicina and Their Antiviral Activity to Avian Leucosis Virus Subgroup J" Marine Drugs 15, no. 11: 345. https://doi.org/10.3390/md15110345
APA StyleSun, Y., Chen, X., Cheng, Z., Liu, S., Yu, H., Wang, X., & Li, P. (2017). Degradation of Polysaccharides from Grateloupia filicina and Their Antiviral Activity to Avian Leucosis Virus Subgroup J. Marine Drugs, 15(11), 345. https://doi.org/10.3390/md15110345