The Anti-Angiogenic Activity of a Cystatin F Homologue from the Buccal Glands of Lampetra morii

Abstract
:1. Introduction
2. Results
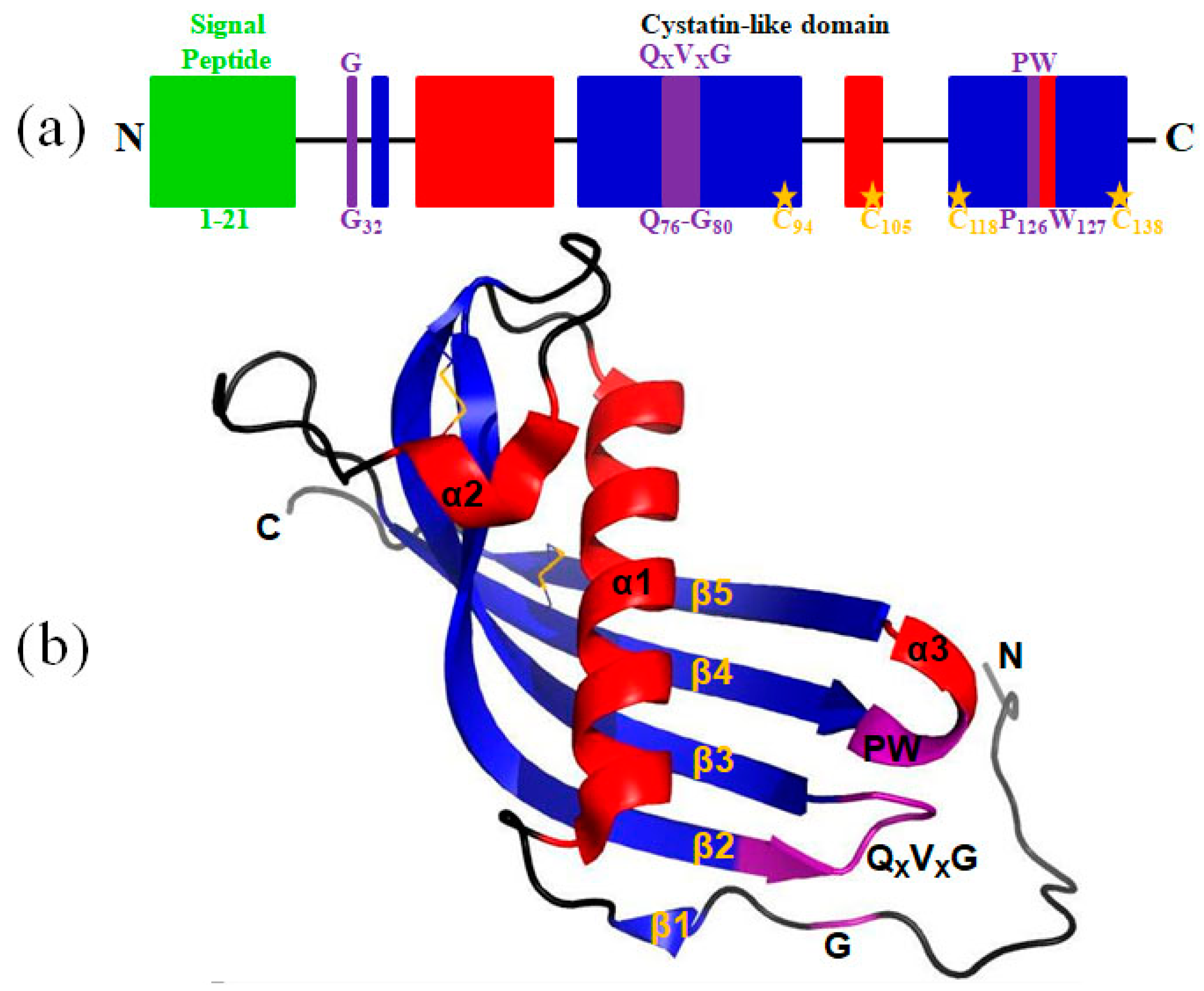
2.1. A Cystatin F Homologue was Identified from the Buccal Glands of L. morii
2.2. Sequence Alignment and Phylogenetic Tree
2.3. Lm-cystatin F was Expressed as a His-Tag Fusion Protein
2.4. rLm-cystatin F Blocked the Activity of Papain and the Proliferation of HUVECs
2.5. rLm-cystatin F Suppressed the Adhesive, Migrated, and Invasive Processes of HUVECs
2.6. rLm-cystatin F Reduced the Abilities of Tube Formation from HUVECs
3. Discussion
4. Materials and Methods
4.1. Cloning of a Cystatin F Homologue (Lm-cystatin F) from the Buccal Glands of L. morii
4.2. Sequence Analysis, Alignment, and Phylogenetic Tree Construction
4.3. Expression, Purification, and Identification of rLm-cystatin F
4.4. Enzyme Activity Assay
4.5. HUVECs’ Culture and MTT Assay
4.6. Adhesion, Migration, and Invasion Assays
4.7. Anti-Angiogenic Activity Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ochieng, J.; Chaudhuri, G. Cystatin superfamily. J. Health Care Poor Underserved 2010, 21, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.T.; Lee, S.J.; Kang, M.A.; Park, J.E.; Kim, B.Y.; Yoon, D.Y.; Yang, Y.; Lee, C.H.; Yeom, Y.I.; Choe, Y.K.; et al. Cystatin SN neutralizes the inhibitory effect of cystatin C on cathepsin B activity. Cell Death Dis. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Zavasnik-Bergant, T.; Repnik, U.; Schweiger, A.; Romih, R.; Jeras, M.; Turk, V.; Kos, J. Differentiation- and maturation-dependent content, localization, and secretion of cystatin C in human dendritic cells. J. Leukoc Biol. 2005, 78, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Zavasnik-Bergant, T. Cystatin protease inhibitors and immune functions. Front. Biosci. 2008, 13, 4625–4637. [Google Scholar] [CrossRef] [PubMed]
- Magister, S.; Kos, J. Cystatins in immune system. J. Cancer 2013, 4, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Ao, J.; Li, Q.; Yang, Z.; Mu, Y. A cystatin F homologue from large yellow croaker (Larimichthys crocea) inhibits activity of multiple cysteine proteinases and Ii chain processing in vitro. Fish Shellfish Immunol. 2016, 48, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Mathews, P.M.; Levy, E. Cystatin C in aging and in Alzheimer’s disease. Ageing Res. Rev. 2016, 32, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Ding, Y.; Li, X.; Wu, X. Cystatin C is a disease-associated protein subject to multiple regulation. Immunol. Cell Biol. 2015, 93, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.S.; Park, S.; Lee, K.W.; Madhi, H.; Park, S.G.; Lee, H.G.; Cho, Y.Y.; Yoo, J.; Dong Kim, K. Extracellular cystatin SN and cathepsin B prevent cellular senescence by inhibiting abnormal glycogen accumulation. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gevorgyan, M.M.; Voronina, N.P.; Goncharova, N.V.; Kozaruk, T.V.; Russkikh, G.S.; Bogdanova, L.A.; Korolenko, T.A. Cystatin C as a marker of progressing cardiovascular events during coronary heart disease. Bull. Exp. Biol. Med. 2017, 162, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Horka, H.; Staudt, V.; Klein, M.; Taube, C.; Reuter, S.; Dehzad, N.; Andersen, J.F.; Kopecky, J.; Schild, H.; Kotsyfakis, M.; et al. The tick salivary protein sialostatin L inhibits the Th9-derived production of the asthma-promoting cytokine IL-9 and is effective in the prevention of experimental asthma. J. Immunol. 2012, 188, 2669–2676. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.; Valdés, J.J.; Kotsyfakis, M. The role of cystatins in tick physiology and blood feeding. Ticks Tick Borne Dis. 2012, 3, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavašnik-Bergant, T.; Vidmar, R.; Sekirnik, A.; Fonović, M.; Salát, J.; Grunclová, L.; Kopáček, P.; Turk, B. Salivary tick cystatin OmC2 targets lysosomal cathepsins S and C in human dendritic cells. Front. Cell Infect. Microbiol. 2017, 7, 288. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, T.H.S.; Lu, S.; Gonzalez, B.R.G.; Torquato, R.J.S.; Tanaka, A.S. Characterization of a novel cystatin type 2 from Rhipicephalus microplus midgut. Biochimie 2017, 140, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Chmelař, J.; Kotál, J.; Langhansová, H.; Kotsyfakis, M. Protease inhibitors in tick saliva: The role of serpins and cystatins in tick-host-pathogen interaction. Front. Cell Infect. Microbiol. 2017, 7, 216. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Tang, N.; Wan, R.; Qi, Y.; Lin, X.; Lin, J. Recombinant snake venom cystatin inhibits the growth, invasion and metastasis of B16F10 cells and MHCC97H cells in vitro and in vivo. Toxicon 2011, 57, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Xie, Q.; Wang, X.; Li, X.; Chen, Y.; Lin, X.; Lin, J. Inhibition of invasion and metastasis of MHCC97H cells by expression of snake venom cystatin through reduction of proteinases activity and epithelial-mesenchymal transition. Arch. Pharm. Res. 2011, 34, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Tang, N.; Wan, R.; Qi, Y.; Lin, X.; Lin, J. Recombinant snake venom cystatin inhibits tumor angiogenesis in vitro and in vivo associated with downregulation of VEGF-A165, Flt-1 and bFGF. Anticancer Agents Med. Chem. 2013, 13, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Tang, N.; Lin, Y.; Wang, X.; Lin, X.; Lin, J. Recombinant adenovirus snake venom cystatin inhibits the growth, invasion, and metastasis of B16F10 cells in vitro and in vivo. Melanoma Res. 2013, 23, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.A.; Madio, B.; Jin, J.; Undheim, E.A.; Fry, B.G.; King, G.F. Melt with this kiss: Paralyzing and liquefying venom of the assassin bug Pristhesancus plagipennis (Hemiptera: Reduviidae). Mol. Cell. Proteomics. 2017, 16, 552–566. [Google Scholar] [CrossRef] [PubMed]
- Pla, D.; Petras, D.; Saviola, A.J.; Modahl, C.M.; Sanz, L.; Pérez, A.; Juárez, E.; Frietze, S.; Dorrestein, P.C.; Mackessy, S.P.; et al. Transcriptomics-guided bottom-up and top-down venomics of neonate and adult specimens of the arboreal rear-fanged Brown Treesnake, Boiga irregularis, from Guam. J. Proteomics 2017, 174, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, Z.; Ao, J.; Chen, X. Molecular and functional characterization of a novel stefin analogue in large yellow croaker (Pseudosciaena crocea). Dev. Comp. Immunol. 2009, 33, 1268–1277. [Google Scholar] [CrossRef] [PubMed]
- Premachandra, H.K.; Wan, Q.; Elvitigala, D.A.; De Zoysa, M.; Choi, C.Y.; Whang, I.; Lee, J. Genomic characterization and expression profiles upon bacterial infection of a novel cystatin B homologue from disk abalone (Haliotis discus discus). Dev. Comp. Immunol. 2012, 38, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.P.; Hu, Y.H.; Sun, L. Scophthalmus maximus cystatin B enhances head kidney macrophage-mediated bacterial killing. Dev. Comp. Immunol. 2010, 34, 1237–1241. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gou, M.; Han, J.; Yuan, X.; Li, Y.; Li, T.; Jiang, Q.; Xiao, R.; Li, Q. Proteomic analysis of buccal gland secretion from fasting and feeding lampreys (Lampetra morii). Proteome Sci. 2018, 16, 9. [Google Scholar] [CrossRef] [PubMed]
- Perišić Nanut, M.; Sabotič, J.; Švajger, U.; Jewett, A.; Kos, J. Cystatin F affects natural killer cell cytotoxicity. Front. Immunol. 2017, 8, 1459. [Google Scholar] [CrossRef] [PubMed]
- Schüttelkopf, A.W.; Hamilton, G.; Watts, C.; van Aalten, D.M. Structural basis of reduction-dependent activation of human cystatin F. J. Biol. Chem. 2006, 281, 16570–16575. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Lu, L.; Zheng, Y.; Liu, X.; Xiao, R.; Wang, J.; Li, Q. Novel anticandidal activity of a recombinant Lampetra japonica RGD3 protein. J. Microbiol. Biotechnol. 2014, 24, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Brillard-Bourdet, M.; Nguyên, V.; Ferrer-di Martino, M.; Gauthier, F.; Moreau, T. Purification and characterization of a new cystatin inhibitor from Taiwan cobra (Naja naja atra) venom. Biochem. J. 1998, 331, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Liu, Y.; Duan, D.; Gou, M.; Wang, H.; Wang, J.; Li, Q.; Xiao, R. Anti-angiogenic activities of CRBGP from buccal glands of lampreys (Lampetra japonica). Biochimie 2016, 123, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Zhang, Z.; Wang, H.; Han, Y.; Gou, M.; Li, B.; Duan, D.; Wang, J.; Liu, X.; Li, Q. Identification and characterization of a cathepsin D homologue from lampreys (Lampetra japonica). Dev. Comp. Immunol. 2015, 49, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.J.; Barrett, A.J. A cystatin-like cysteine proteinase inhibitor from venom of the African puff adder (Bitis arietans). Biochem. J. 1987, 246, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Kotsyfakis, M.; Sá-Nunes, A.; Francischetti, I.M.; Mather, T.N.; Andersen, J.F.; Ribeiro, J.M. Antiinflammatory and immunosuppressive activity of sialostatin L, a salivary cystatin from the tick Ixodes scapularis. J. Biol. Chem. 2006, 281, 26298–26307. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Gai, X.; Wang, L.; Song, L.; Zhang, H.; Qiu, L.; Wang, M.; Siva, V.S. Identification and characterization of a Cystatin gene from Chinese mitten crab Eriocheir sinensis. Fish Shellfish Immunol. 2010, 29, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, X.; Cao, J.; Zhou, Y.; Gong, H.; Zhang, H.; Li, X.; Zhou, J. Characterization of a secreted cystatin from the tick Rhipicephalus haemaphysaloides. Exp. Appl. Acarol. 2015, 67, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, G.; Li, Z.; Cheng, Y.; Gao, C.; Zeng, L.; Chen, J.; Yan, L.; Sun, X.; Guo, L.; et al. Molecular cloning, recombinant expression and antifungal activity of BnCPI, a Cystatin in Ramie (Boehmeria nivea L.). Genes (Basel) 2017, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, L.; Liu, X.; Wang, S.; Ehsan, M.; Yan, R.; Song, X.; Xu, L.; Li, X. Characterization of a secreted cystatin of the parasitic nematode Haemonchus contortus and its immune-modulatory effect on goat monocytes. Parasit. Vectors. 2017, 10, 425. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, K.; Kanemori, M.; Yanagi, H.; Yura, T. Overexpression of trigger factor prevents aggregation of recombinant proteins in Escherichia coli. Appl. Environ. Microbiol. 2000, 66, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Lu, L.; Liu, X.; Li, Q. Anti-angiogenic activities of Lj-RGD3 toxin protein from Lampetra japonica and its mutation protein Lj-112 without RGD motifs. Sheng Wu Gong Cheng Xue Bao 2011, 27, 1428–1437. [Google Scholar] [PubMed]
- Li, Y.; Zhu, H.; Wei, X.; Li, H.; Yu, Z.; Zhang, H.; Liu, W. LPS induces HUVEC angiogenesis in vitro through miR-146a-mediated TGF-β1 inhibition. Am. J. Transl. Res. 2017, 9, 591–600. [Google Scholar] [PubMed]
- Shin, M.R.; Kang, S.K.; Kim, Y.S.; Lee, S.Y.; Hong, S.C.; Kim, E.C. TNF-α and LPS activate angiogenesis via VEGF and SIRT1 signalling in human dental pulp cells. Int. Endod. J. 2015, 48, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Yamakawa, Y.; Omori-Satoh, T. Primary structure of the antihemorrhagic factor in serum of the Japanese Habu: a snake venom metalloproteinase inhibitor with a double-headed cystatin domain. J. Biochem. 1992, 112, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Valente, R.H.; Dragulev, B.; Perales, J.; Fox, J.W.; Domont, G.B. BJ46a, a snake venom metalloproteinase inhibitor. Isolation, characterization, cloning and insights into its mechanism of action. Eur. J. Biochem. 2001, 268, 3042–3052. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.; St Pierre, L.; Trabi, M.; Johnson, L.A.; de Jersey, J.; Masci, P.P.; Lavin, M.F. Cloning and characterisation of novel cystatins from elapid snake venom glands. Biochimie 2011, 93, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Otto, H.H.; Schirmeister, T. Cysteine proteases and their inhibitors. Chem. Rev. 1997, 97, 133–172. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo Azevedo, F.; Santanna, L.P.; Bóbbo, V.C.; Libert, E.A.; Araújo, E.P.; Abdalla Saad, M.; Lima, M.H.M. Evaluating the effect of 3% papain gel application in cutaneous wound healing in mice. Wounds 2017, 29, 96–101. [Google Scholar] [PubMed]
- Premzl, A.; Turk, V.; Kos, J. Intracellular proteolytic activity of cathepsin B is associated with capillary-like tube formation by endothelial cells in vitro. J. Cell. Biochem. 2006, 97, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Li, Q.; Han, J.; Gou, M.; Zheng, Y.; Li, B.; Xiao, R.; Wang, J. rLj-RGD3 induces apoptosis via the mitochondrial-dependent pathway and inhibits adhesion, migration and invasion of human HeyA8 cells via FAK pathway. Int. J. Biol. Macromol. 2017, 96, 652–668. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Amino Acids | Species | Accession Number | Identity (%) |
|---|---|---|---|---|
| Mammals | 145 | Homo sapiens | CAD52872.1 | 33 |
| Mammals | 144 | Mus musculus | NP_034107.2 | 32 |
| Mammals | 145 | Hipposideros armiger | XP_019488972 | 31 |
| Aves | 147 | Gallus gallus | NP_001186323 | 32 |
| Aves | 147 | Columba livia | PKK20598.1 | 31 |
| Aves | 147 | Buceros rhinoceros silvestris | XP_010137948.1 | 30 |
| Amphibians | 149 | Xenopus laevis | NP_001091281 | 32 |
| Amphibians | 145 | Alligator mississippiensis | KYO18358.1 | 38 |
| Amphibians | 145 | Chrysemys picta bellii | XP_005293586 | 35 |
| Amphibians | 145 | Pelodiscus sinensis | XP_006136954 | 31 |
| Reptiles | 145 | Eublepharis macularius | JAC94872 | 32 |
| Reptiles | 154 | Python regius | JAC94922 | 31 |
| Fishes | 143 | Oncorhynchus mykiss | XP_021420522.1 | 35 |
| Fishes | 143 | Salmo salar | NP_001134364.1 | 34 |
| Fishes | 133 | Esox lucius | NP_001297968.1 | 38 |
| Fishes | 128 | Danio rerio | NP_001082882 | 36 |
| Fishes | 140 | Takifugu rubripes | XP_011601516 | 32 |
| Agnathans | 152 | Lampetra morii | MG902948 | 100 |
| Nematodas | 136 | Necator americanus | ETN77353.1 | 27 |
| Nematodas | 141 | Ancylostoma duodenale | KIH58790.1 | 26 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Li, B.; Wang, J.; Xiao, R. The Anti-Angiogenic Activity of a Cystatin F Homologue from the Buccal Glands of Lampetra morii. Mar. Drugs 2018, 16, 477. https://doi.org/10.3390/md16120477
Zhu M, Li B, Wang J, Xiao R. The Anti-Angiogenic Activity of a Cystatin F Homologue from the Buccal Glands of Lampetra morii. Marine Drugs. 2018; 16(12):477. https://doi.org/10.3390/md16120477
Chicago/Turabian StyleZhu, Mingru, Bowen Li, Jihong Wang, and Rong Xiao. 2018. "The Anti-Angiogenic Activity of a Cystatin F Homologue from the Buccal Glands of Lampetra morii" Marine Drugs 16, no. 12: 477. https://doi.org/10.3390/md16120477
APA StyleZhu, M., Li, B., Wang, J., & Xiao, R. (2018). The Anti-Angiogenic Activity of a Cystatin F Homologue from the Buccal Glands of Lampetra morii. Marine Drugs, 16(12), 477. https://doi.org/10.3390/md16120477