Terpenoids in Marine Heterobranch Molluscs


Abstract
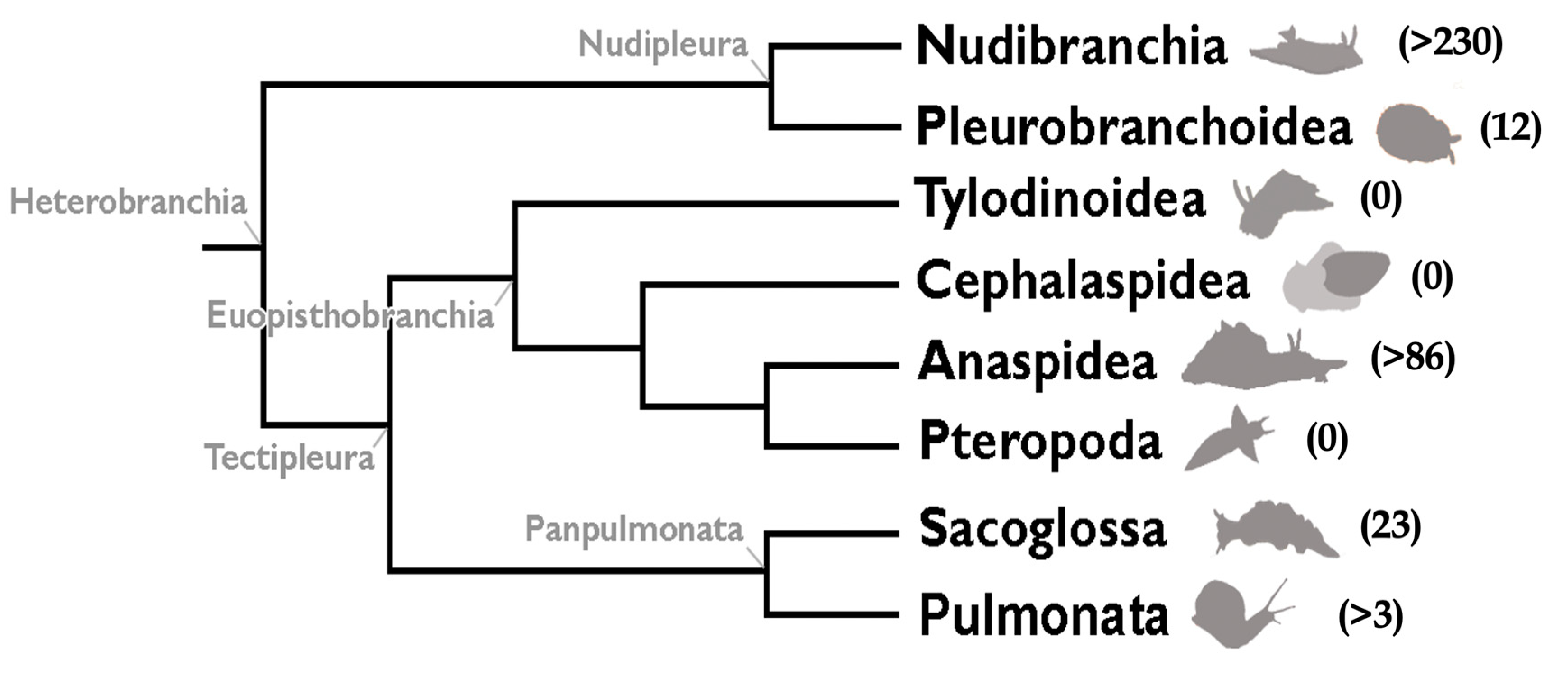
:1. Background
2. Monoterpenoids
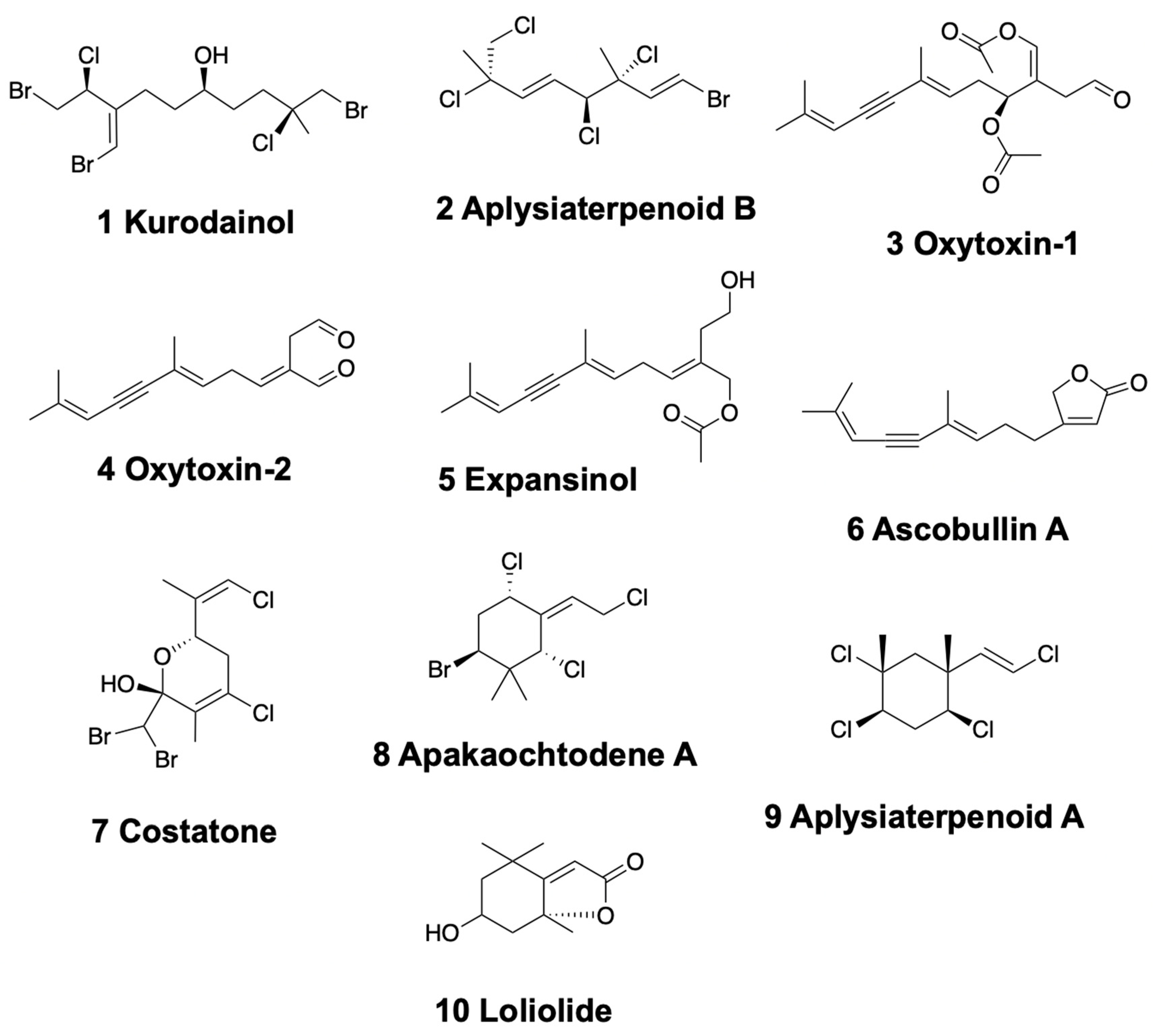
2.1. Linear Monoterpenoids
2.2. Monocyclic Monoterpenoids
3. Sesquiterpenoids
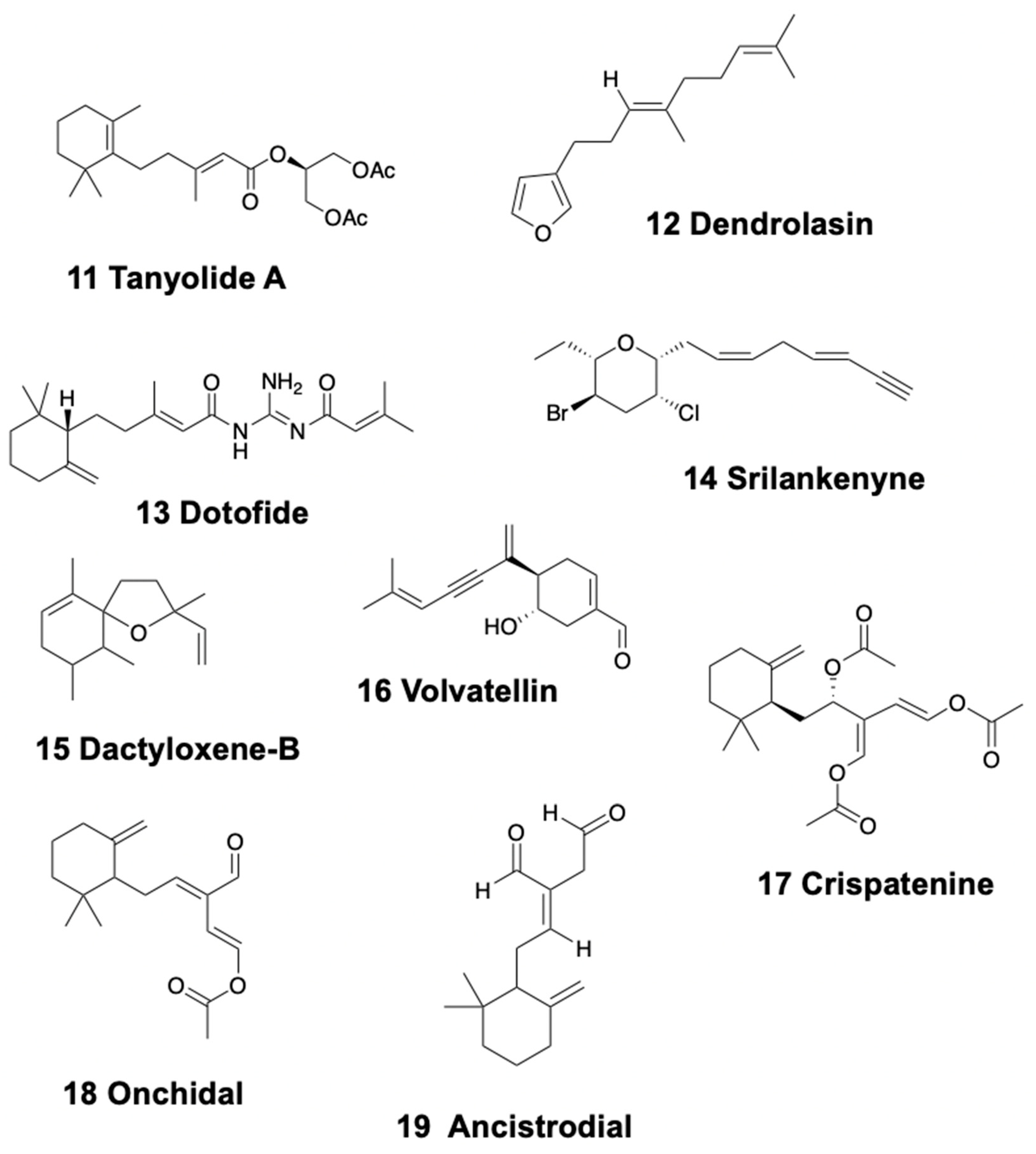
3.1. Linear Sesquiterpenoids
3.2. Monocyclic Sesquiterpenoids
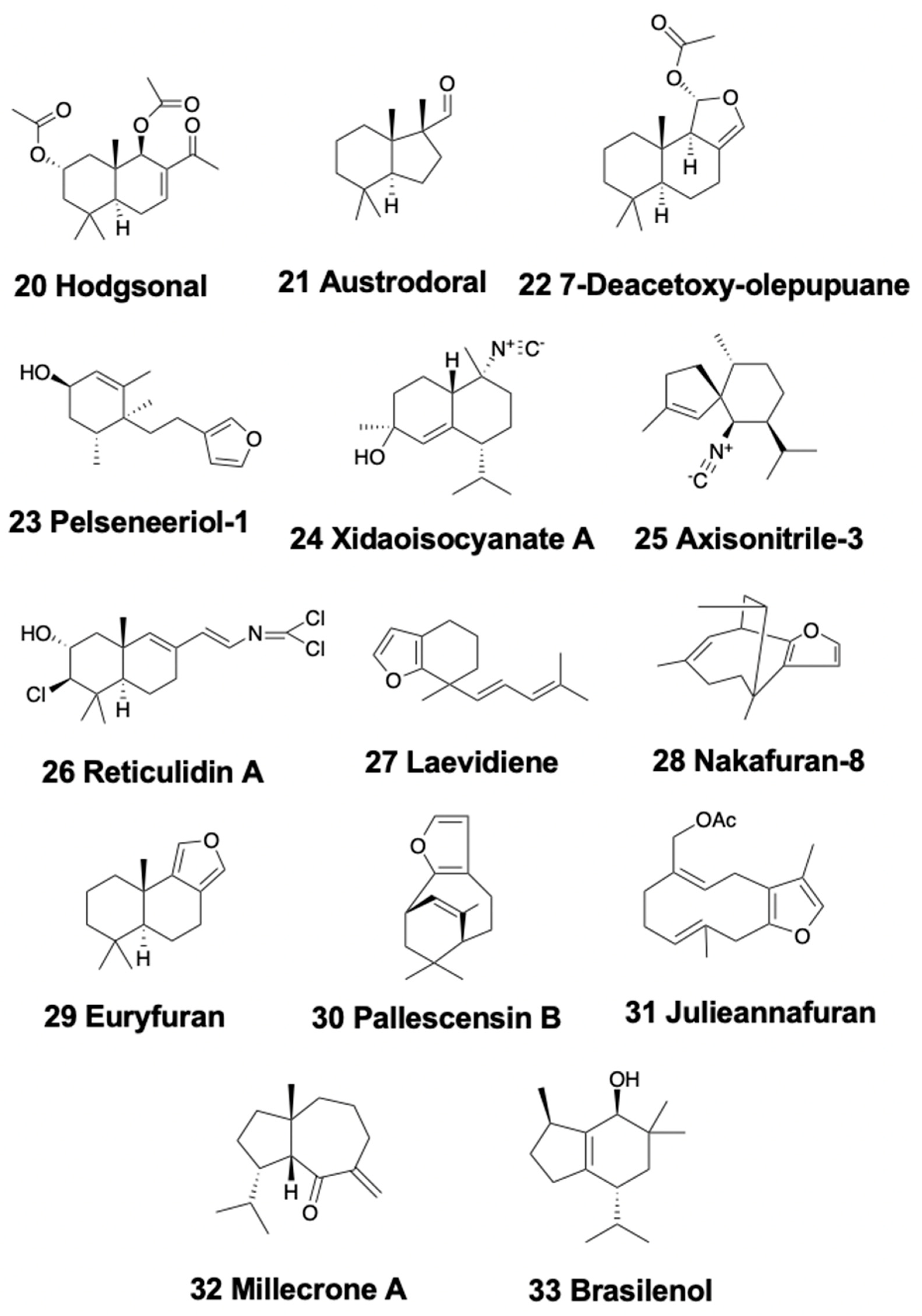
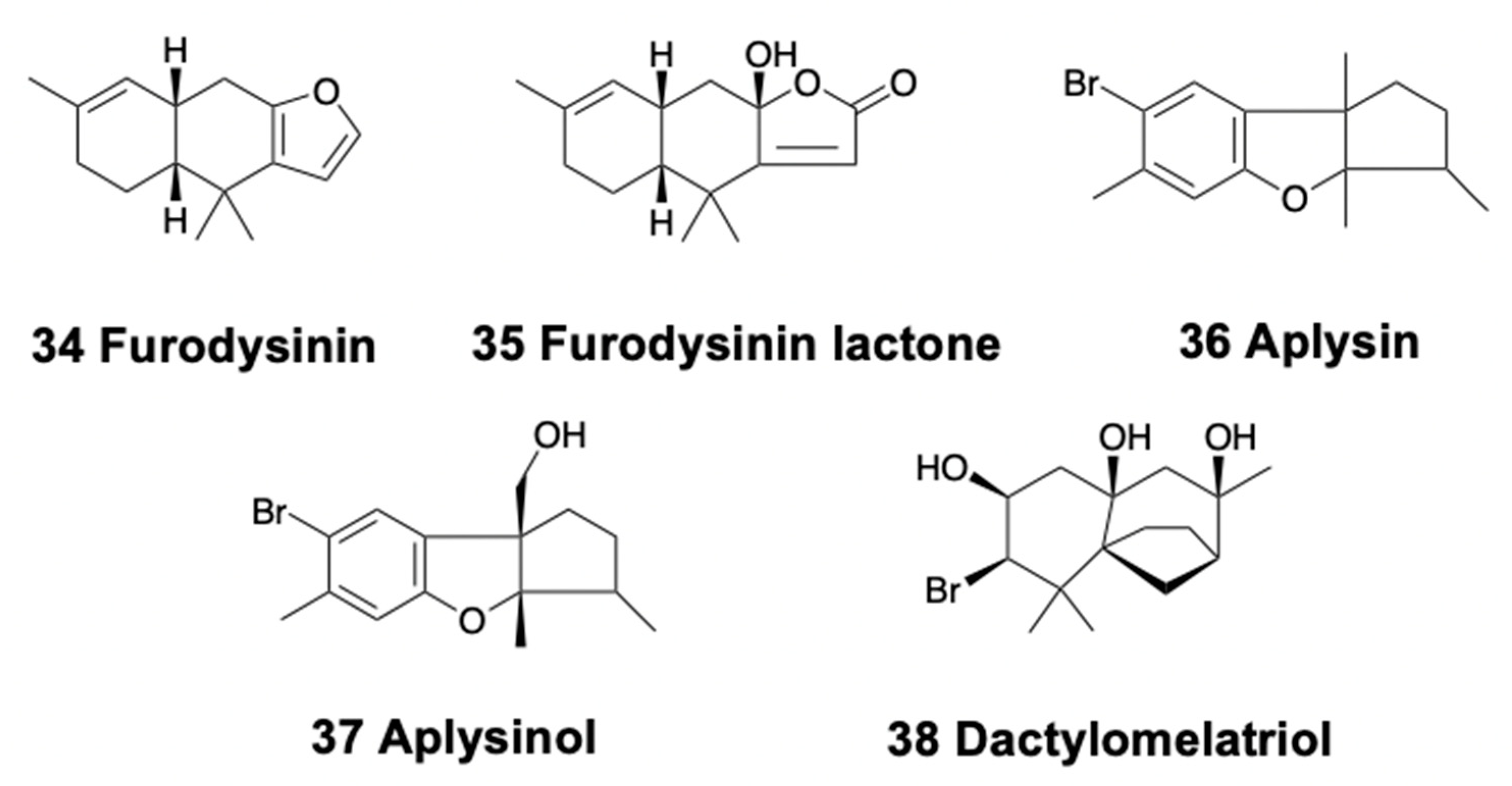
3.3. Bicyclic Sesquiterpenoids
3.4. Tricyclic Sesquiterpenoids
4. Diterpenoids
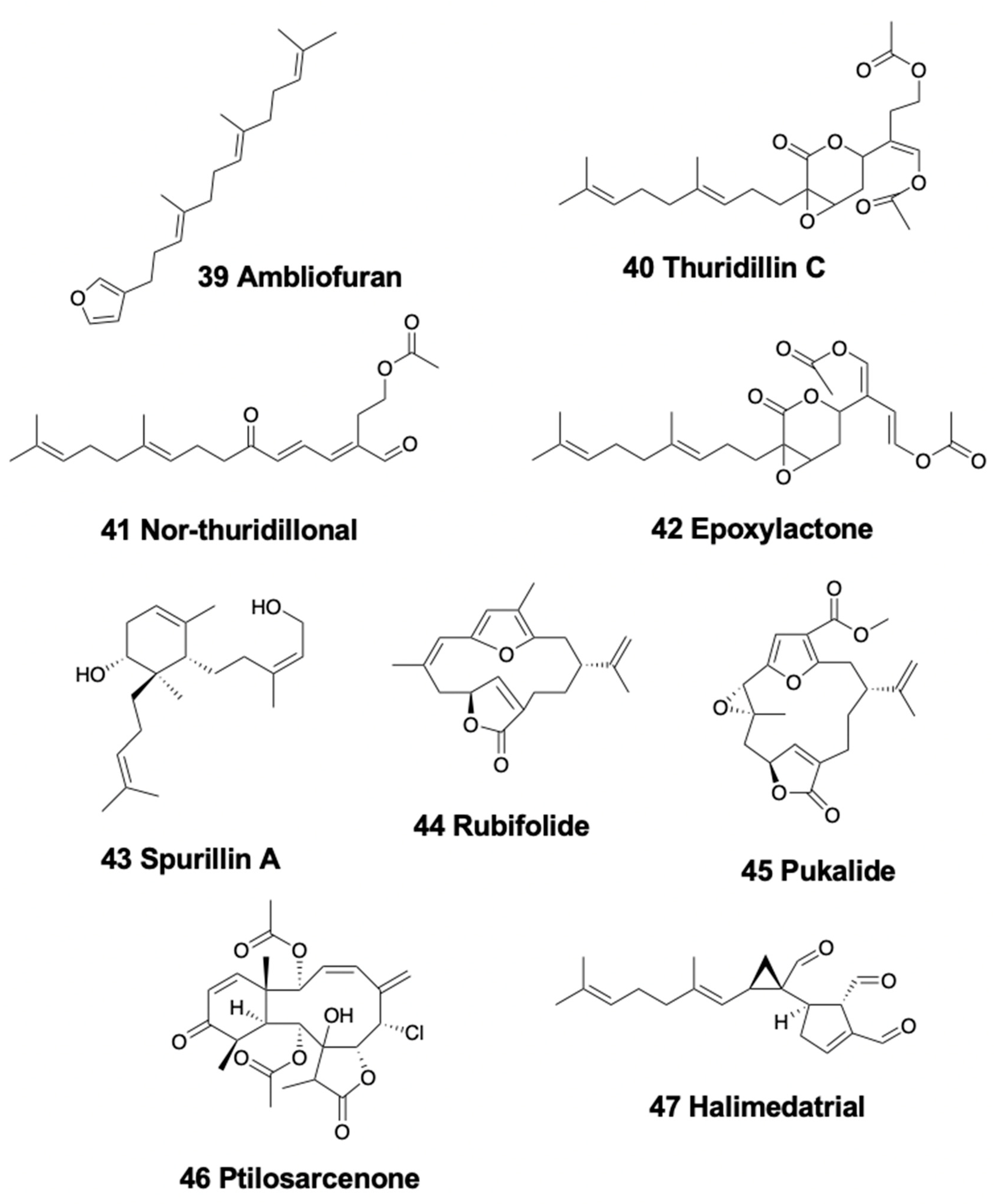
4.1. Linear Diterpenoids
4.2. Monocyclic Diterpenoids
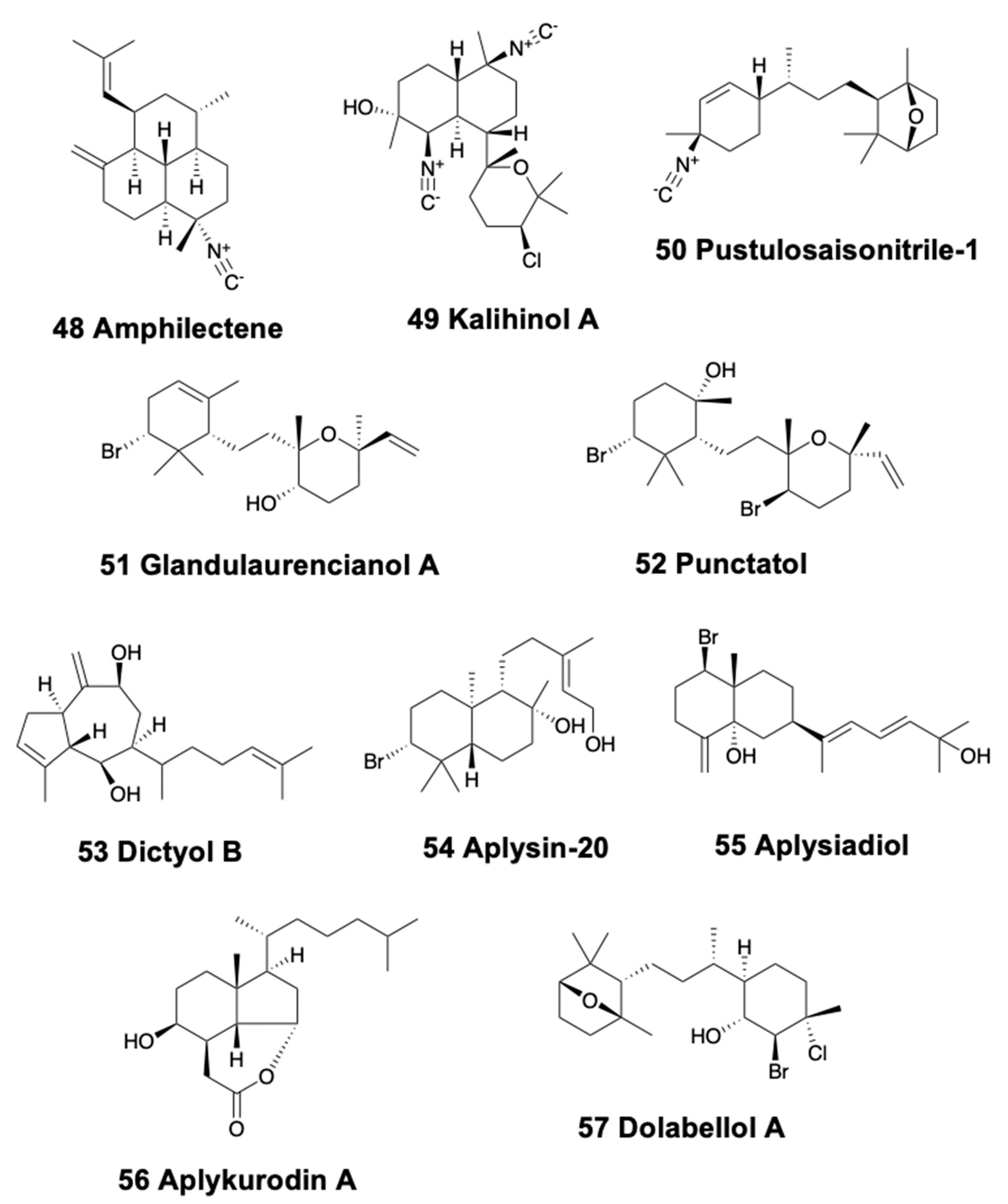
4.3. Bicyclic Diterpenoids
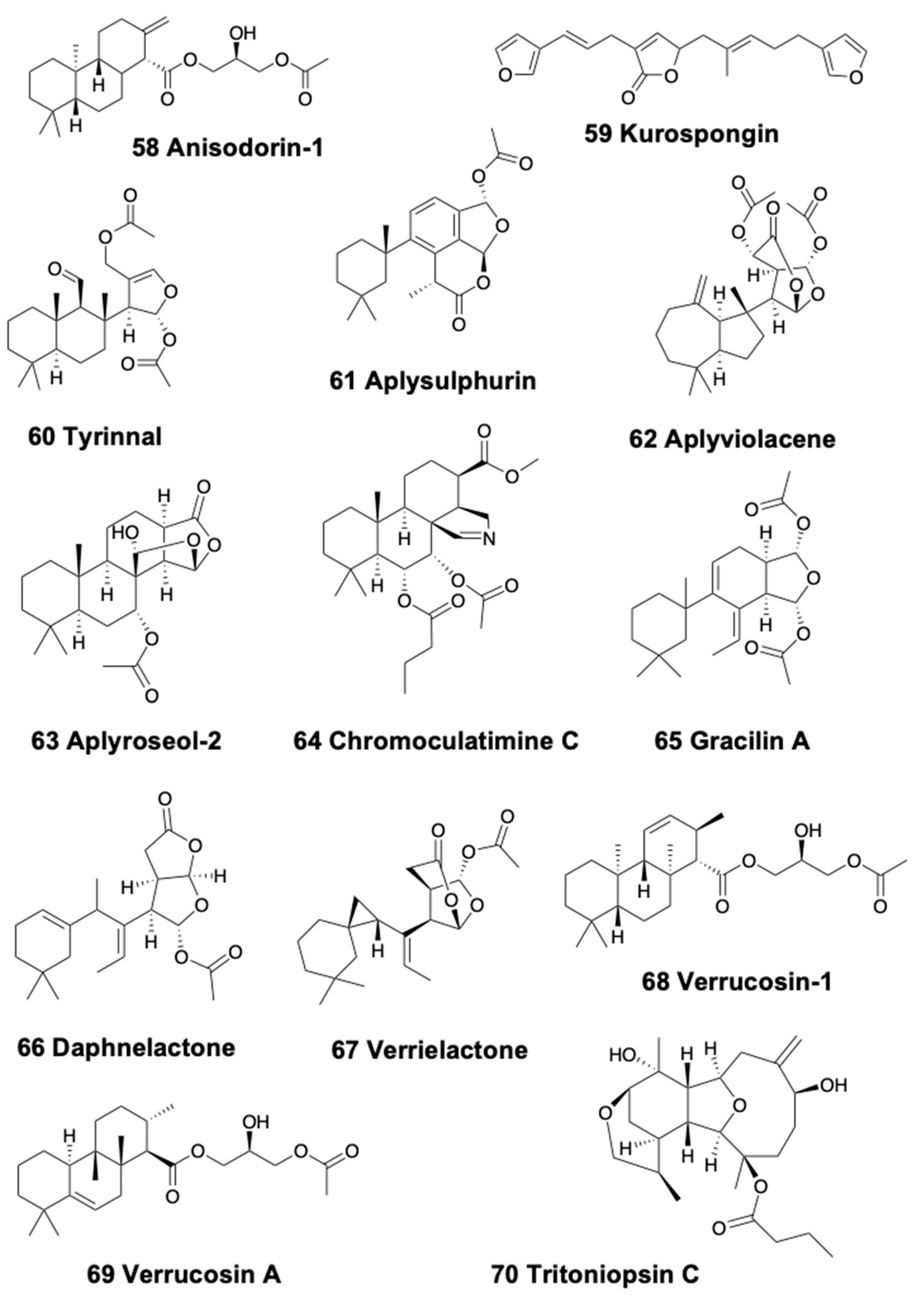
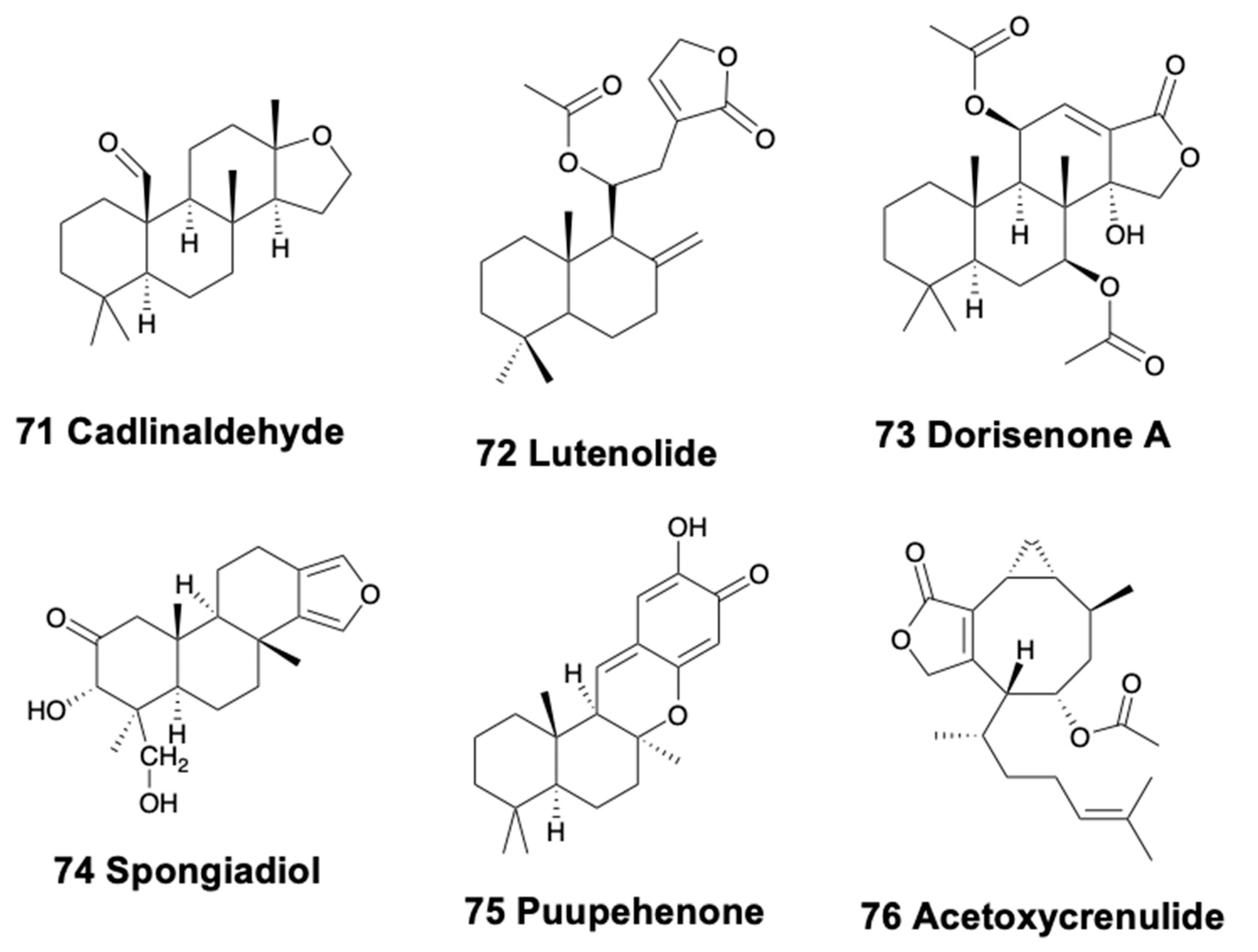
4.4. Tricyclic Diterpenoids
4.5. Tetracyclic Diterpenoids
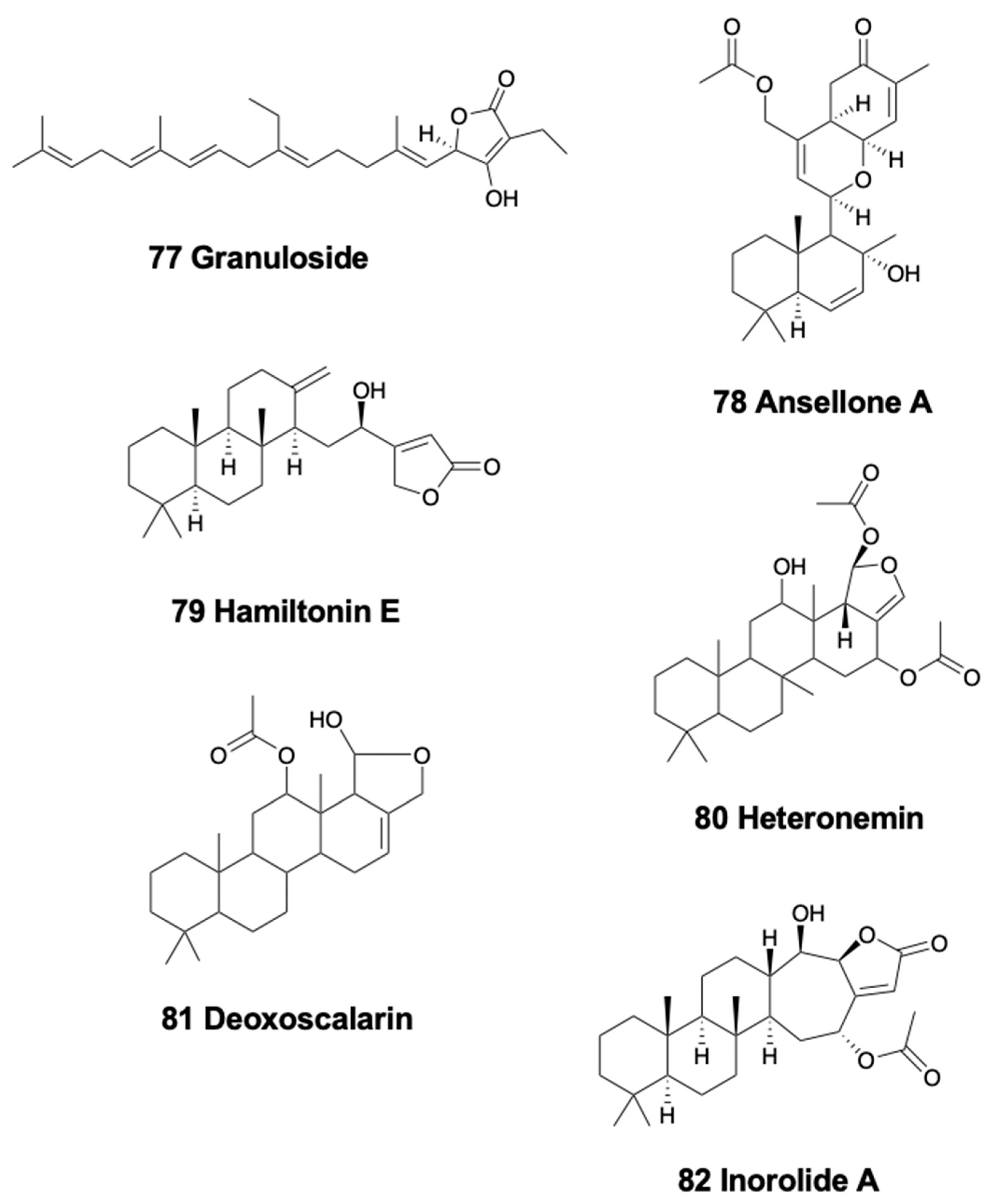
5. Sesterterpenoids
5.1. Linear Sesterterpenoids
5.2. Bicyclic Sesterterpenoids
5.3. Tetracyclic Sesterterpenoids
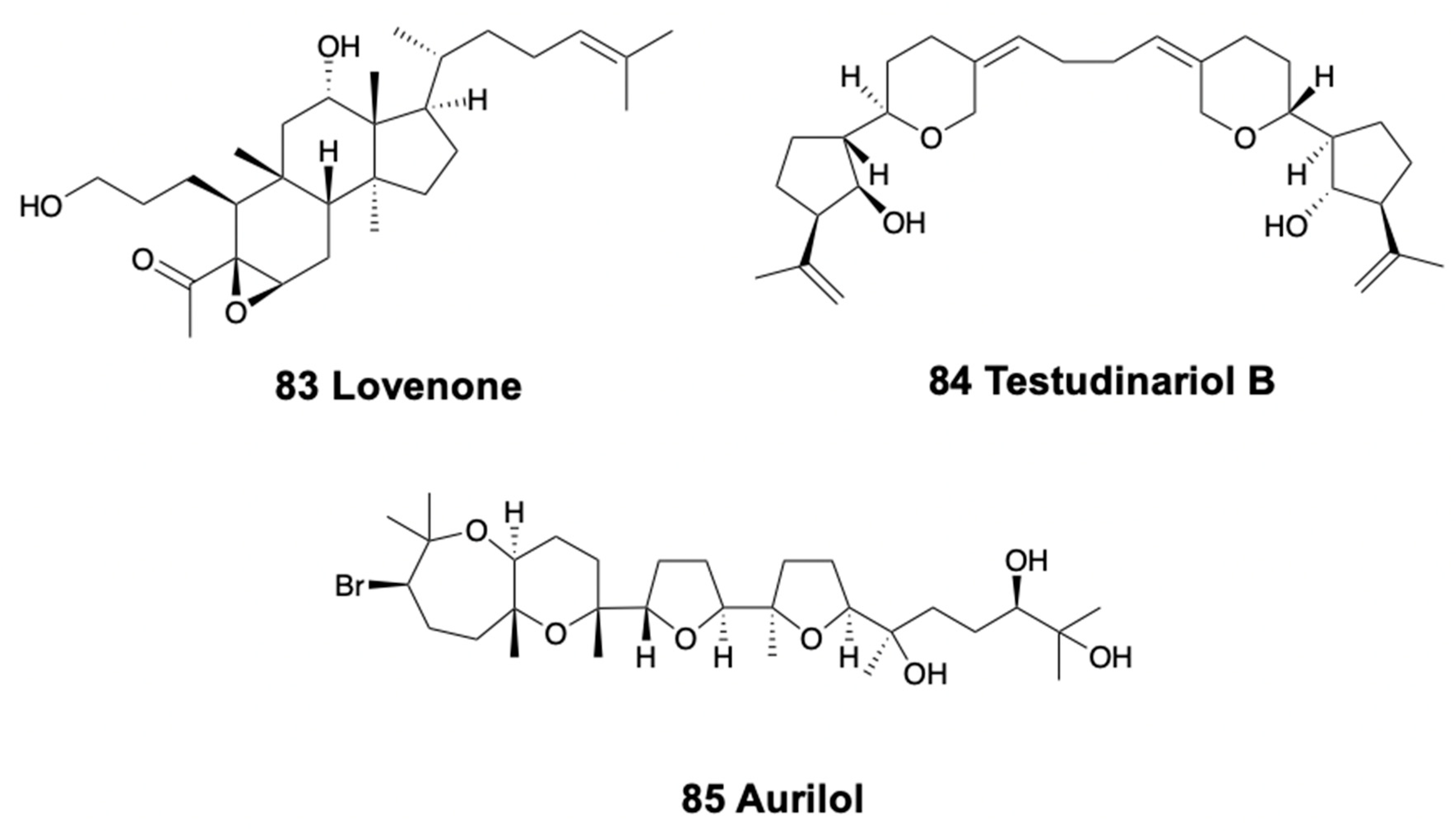
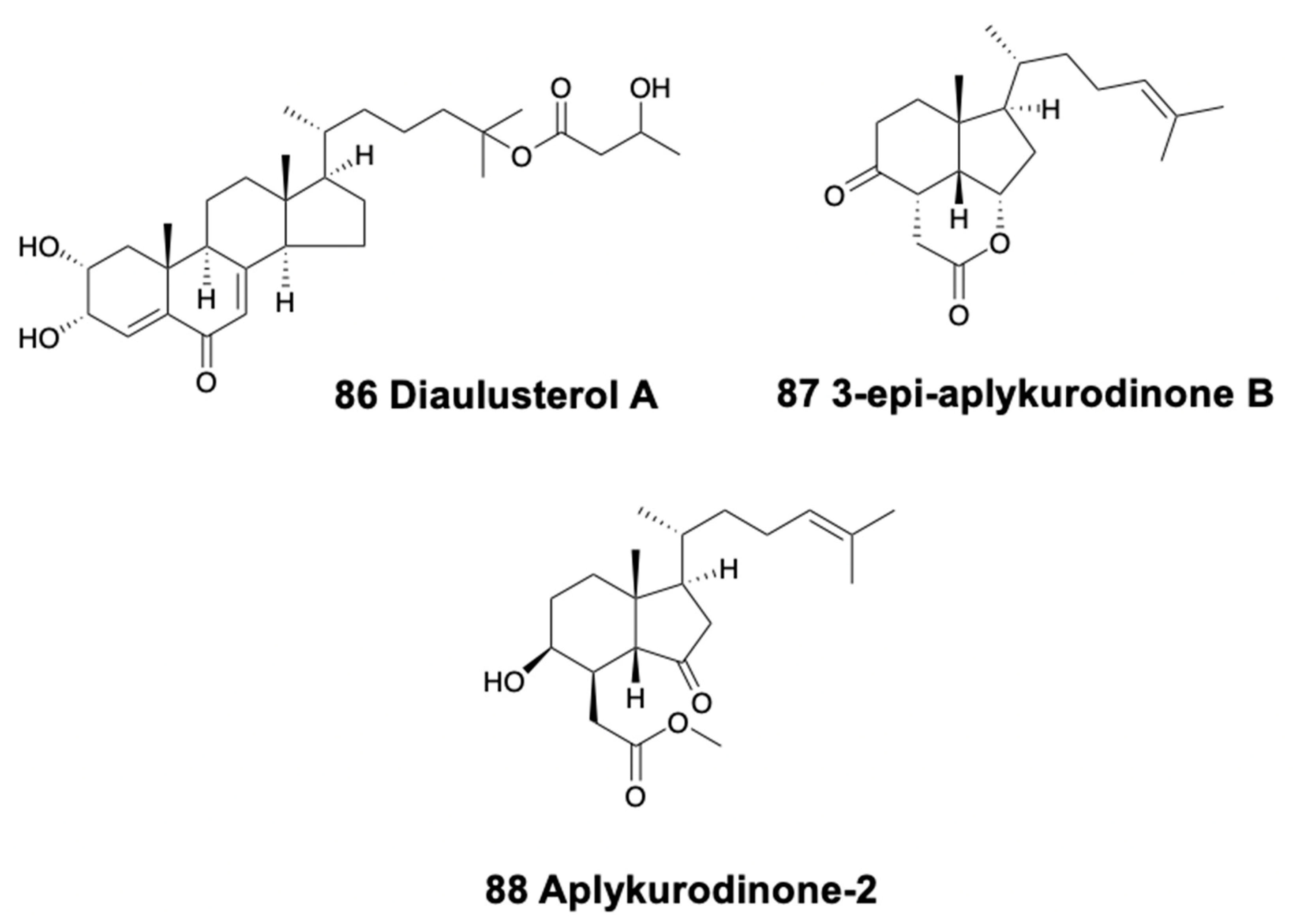
6. Triterpenoids
7. Carotenoids (Tetraterpenoids)
8. Steroids
9. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Blunt, J.W.; Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2018, 35, 8–53. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila, C.; Núñez-Pons, L.; Moles, J. From the tropics to the poles: Chemical defensive strategies in sea slugs (Mollusca: Heterobranchia). In Chemical Ecology: The Ecological Impacts of Marine Natural Products; Puglisi, M.P., Becerro, M.A., Eds.; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Puglisi, M.P.; Becerro, M.A. Chemical Ecology: The Ecological Impacts of Marine Natural Products; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Hare, S.R.; Farnham, J.M.; Tantillo, D.J. Putative biosynthetic cycloadditions en route to the diterpenoid (+)-chatancin. Tetrahedron 2017, 73, 4227–4232. [Google Scholar] [CrossRef]
- Davis, E.M.; Croteau, R. Cyclization enzymes in the biosynthesis of monoterpenes, sesquiterpenes, and diterpenes. Topics Curr. Chem. 2000, 209, 53–95. [Google Scholar]
- Kumari, I.; Ahmed, M.; Akhter, Y. Evolution of catalytic microenvironment governs substrate and product diversity in trichodiene synthase and other terpene fold enzymes. Biochimie 2017, 144, 9–20. [Google Scholar] [CrossRef]
- Pazouki, L.; Niinemets, Ü. Multi-substrate terpene synthases: Their occurrence and physiological significance. Front. Plant Sci. 2016, 7, 1019. [Google Scholar] [CrossRef] [Green Version]
- Gil Ruíz, P. Productos Naturales; Nafarroako Unibertsitate Publikoa: Nafarroa, Spain, 2002. [Google Scholar]
- Blunt, J.W.; Munro, M.H.G. Dictionary of Marine Natural Products; Chapman and Hall/CRC: Boca Raton, FL, USA, 2008. [Google Scholar]
- Hegazy, M.E.F.; Mohamed, T.A.; Alhammady, M.A.; Shaheen, A.M.; Reda, E.H.; Elshamy, A.I.; Aziz, M.; Paré, P.W. Molecular architecture and biomedical leads of terpenes from Red Sea marine invertebrates. Mar. Drugs 2015, 13, 3154–3181. [Google Scholar] [CrossRef]
- Gross, H.; König, G.M. Terpenoids from marine organisms: Unique structures and their pharmacological potential. Phytochem. Rev. 2006, 5, 115–141. [Google Scholar] [CrossRef]
- Grosso, C.; Valentão, P.; Ferreres, F.; Andrade, P.B. Alternative and efficient extraction methods for marine-derived compounds. Mar. Drugs 2015, 13, 3182–3230. [Google Scholar] [CrossRef] [Green Version]
- Bayona, L.M.; Videnova, M.; Choi, Y.H. Increasing metabolic diversity in marine sponges extracts by controlling extraction parameters. Mar. Drugs 2018, 16, 393. [Google Scholar] [CrossRef] [Green Version]
- Avila, C. Natural products of opisthobranch molluscs: A biological review. Oceanogr. Mar. Biol. Annu. Rev. 1995, 33, 487–559. [Google Scholar]
- Costa Leal, M.; Madeira, C.; Brandão, C.A.; Puga, J.; Calado, R. Bioprospecting of marine invertebrates for new natural products—A chemical and zoogeographical perspective. Molecules 2012, 17, 9842–9854. [Google Scholar] [CrossRef] [PubMed]
- Schrödl, M.; Jörger, K.M.; Klussmann-Kolb, A.; Wilson, N.G. Bye bye ‘Opisthobranchia’! A review on the contribution of mesopsammic sea slugs to euthyneuran systematics. Thalassas 2011, 27, 101–112. [Google Scholar]
- Medina, M.; Lal, S.; Vallès, Y.; Takaoka, T.L.; Dayrat, B.A.; Boore, J.L.; Gosliner, T. Crawling through time: Transition of snails to slugs dating back to the Paleozoic, based on mitochondrial phylogenomics. Mar. Genomics 2011, 4, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Wägele, H.; Klussmann-Kolb, A.; Verbeek, E.; Schrödl, M. Flashback and foreshadowing—A review of the taxon Opisthobranchia. Org. Div. Evol. 2014, 14, 133–149. [Google Scholar] [CrossRef] [Green Version]
- Zapata, F.; Wilson, N.G.; Howison, M.; Andrade, S.C.; Jörger, K.M.; Schrödl, M.; Goetz, F.E.; Giribet, G.; Dunn, C.W. Phylogenomic analyses of deep gastropod relationships reject Orthogastropoda. Proc. Royal Soc. B Biol. Sci. 2014, 281, 20141739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WoRMS. World Register of Marine Species, Database. Available online: http://www.marinespecies.org (accessed on 29 January 2020).
- Avila, C. Molluscan natural products as biological models: Chemical ecology, histology, and laboratory culture. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 1–23. [Google Scholar]
- McClintock, J.B.; Baker, P.J. (Eds.) Marine Chemical Ecology; CRC Marine Science Series Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Thompson, T.E. Acidic allomones in marine organisms. J. Mar. Biol. Assoc. UK 1988, 68, 499–517. [Google Scholar] [CrossRef]
- Wägele, H.; Ballesteros, M.; Avila, C. Defensive grandular structures in opistobranch molluscs—From histology to ecology. Oceanogr. Mar. Biol. Annu. Rev. 2006, 44, 197–276. [Google Scholar]
- Wägele, H.; Knezevic, K.; Moustafa, A.Y. Distribution and morphology of defensive acid-secreting glands in Nudipleura (Gastropoda: Heterobranchia), with an emphasis on Pleurobranchomorpha. J. Moll. Stud. 2017, 83, 422–433. [Google Scholar] [CrossRef]
- Cimino, G.; Ghiselin, M.T. Chemical defense and the evolution of opisthobranch gastropods. Proc. California Acad. Sci. 2009, 60, 175–422. [Google Scholar]
- Ianora, A.; Boersma, M.; Casotti, R.; Fontana, A.; Harder, J.; Hoffmann, F.; Pavia, H.; Potin, P.; Poulet, S.A.; Toth, G. New trends in marine chemical ecology. Estuar. Coasts 2006, 29, 531–551. [Google Scholar] [CrossRef] [Green Version]
- Paul, V.J.; Arthur, K.E.; Ritson-Williams, R.; Ross, C.; Sharp, K. Chemical defenses: From compounds to communities. Biol. Bull. 2007, 213, 226–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila, C. A preliminary catalogue of natural substances of opisthobranch molluscs from Western Mediterranean and near Atlantic. Sci. Mar. 1992, 56, 373–382. [Google Scholar]
- Avila, C. Substancias Naturales de Moluscos Opistobranquios: Estudio de Su Estructura, Origen y Función En Ecosistemas Bentónicos. Ph.D. Thesis, Universitat de Barcelona, Barcelona, Spain, 1993; p. 546. [Google Scholar]
- Avila, C.; Durfort, M. Histology of epithelia and mantle glands of selected species of doridacean mollusks with chemical defensive strategies. Veliger 1996, 39, 148–163. [Google Scholar]
- Puglisi, M.P.; Sneed, J.M.; Sharp, K.H.; Ritson-Williams, R.; Paul, V.J. Marine chemical ecology in benthic environments. Nat. Prod. Rep. 2014, 31, 1510–1553. [Google Scholar] [CrossRef] [PubMed]
- Avila, C.; Taboada, S.; Núñez-Pons, L. Antarctic marine chemical ecology: What is next? Mar. Ecol. 2008, 29, 1–71. [Google Scholar] [CrossRef]
- Paul, V.J. Ecological Roles of Marine Natural Products; Comstock Publishications Association: Ithaka, NY, USA, 1992. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef]
- Haefner, B. Drugs from the deep: Marine natural products as drug candidates. Drug Discov. Today 2003, 8, 536–544. [Google Scholar] [CrossRef]
- Clardy, J.; Walsh, C. Lessons from natural molecules. Nature 2004, 432, 829–837. [Google Scholar] [CrossRef]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nature Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. New horizons for old drugs and drug leads. J. Nat. Prod. 2014, 77, 703–723. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Marine-sourced anti-cancer and cancer pain control agents in clinical and late preclinical development. Mar. Drugs 2014, 12, 255–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faircloth, G.; Cuevas, M.C. Kahalalide F and ES285: Potent anticancer agents from marine molluscs. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 363–379. [Google Scholar]
- Kigoshi, H.; Kita, M. Antitumor effects of sea hare-derived compounds in cancer. In Handbook of Anticancer Drugs from Marine Origin; Kim, S.-K., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2015; pp. 701–739. [Google Scholar]
- Garson, M.J. Marine mollusks from Australia and New Zealand: Chemical and ecological studies. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 159–174. [Google Scholar]
- Davies-Coleman, M.T. Secondary metabolites from the marine gastropod molluscs of Antarctica, Southern Africa and South America. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 133–157. [Google Scholar]
- Wahidulla, S.; Guo, Y.W.; Fakhr, I.M.I.; Mollo, E. Chemical diversity in opisthobranch molluscs from scarcely investigated Indo-Pacific areas. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 175–198. [Google Scholar]
- Miyamoto, T. Selected bioactive compounds from Japanese anaspideans and nudibranchs. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 199–214. [Google Scholar]
- Wang, J.R.; He, W.F.; Guo, Y.W. Chemistry, chemoecology, and bioactivity of the South China Sea opisthobranch molluscs and their dietary organisms. J. Asian Nat. Prod. Res. 2013, 15, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Andersen, R.J.; Desjardine, K.; Woods, K. Skin chemistry of nudibranchs from the West Coast of North America. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 277–301. [Google Scholar]
- Tian, Y.; Li, Y.-L.; Zhao, F.-C. Secondary metabolites from polar organisms. Mar. Drugs 2017, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, H.; Sakai, R.; Jimbo, M. Bioactive molecules from sea hares. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 215–239. [Google Scholar]
- Darias, J.; Cueto, M.; Díaz-Marrero, A.R. The chemistry of marine pulmonate gastropods. In Molluscs: Progress in Molecular and Subcellular Biology; Cimino, G., Gavagnin, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43, pp. 105–131. [Google Scholar]
- Dean, L.J.; Prinsep, M.R. The chemistry and chemical ecology of nudibranchs. Nat. Prod. Rep. 2017, 34, 1359–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimino, G.; Gavagnin, M. (Eds.) Molluscs: Progress in Molecular and Subcellular Biology; Springer: Berlin/Heidelberg, Germany, 2006; Volume 43. [Google Scholar]
- Benkendorff, K. Molluscan biological and chemical diversity: Secondary metabolites and medicinal resources produced by marine molluscs. Biol. Rev. 2010, 85, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Kandyuk, R.P. Sterols and their functional role in Mollusks (a review). Hydrobiol. J. 2006, 42, 56–66. [Google Scholar] [CrossRef]
- Garson, M. Marine natural products as antifeedants. In Comprehensive Natural Products II. Chemistry and Biology; Mander, L., Liu, H.W., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2010; pp. 503–537. [Google Scholar]
- Cimino, G.; Ghiselin, M.T. Marine natural products chemistry as an evolutionary narrative. In Marine Chemical Ecology; McClintock, J.B., Baker, P.J., Eds.; CRC Marine Science Series Press: Boca Raton, FL, USA, 2001; pp. 115–154. [Google Scholar]
- McClintock, J.B.; Amsler, C.D.; Baker, B.J. Overview of the chemical ecology of benthic marine invertebrates along the Western Antarctic peninsula. Integr. Comp. Biol. 2010, 50, 967–980. [Google Scholar] [CrossRef]
- Núñez-Pons, L.; Avila, C. Natural products mediating ecological interactions in Antarctic benthic communities: A mini-review of the known molecules. Nat. Prod. Rep. 2015, 32, 1114–1130. [Google Scholar] [CrossRef]
- Katayama, A.; Ina, K.; Nozaki, H.; Nakayama, M. Structural elucidation of kurodainol, a novel halogenated monoterpene from sea hare (Aplysia kurodai). Agric. Biol. Chem. 1982, 46, 859–860. [Google Scholar] [CrossRef]
- Kusumi, T.; Uchida, H.; Inouye, Y.; Ishitsuka, M.; Yamamoto, H.; Kakisawa, H. Novel cytotoxic monoterpenes having a halogenated tetrahydropyran from Aplysia kurodai. J. Org. Chem. 1987, 52, 4597–4600. [Google Scholar] [CrossRef]
- Miyamoto, T.; Higuchi, R.; Marubayashi, N.; Komori, T. Two new polyhalogenated monoterpenes from the sea hare Aplysia kurodai. Liebigs Ann. Chem. 1988, 12, 1191–1193. [Google Scholar] [CrossRef]
- Imperato, F.; Minale, L.; Riccio, R. Constituents of the digestive gland of molluscs of the genus Aplysia. II. Halogenated monoterpenes from Aplysia limacina. Experientia 1977, 33, 1273–1274. [Google Scholar] [CrossRef]
- Quiñoá, E.; Castedo, L.; Riguera, R. The halogenated monoterpenes of Aplysia punctata. Comp. Study. Comp. Biochem. Physiol. B 1989, 92, 99–101. [Google Scholar] [CrossRef]
- Ortega, M.J.; Zubía, E.; Salvá, J. New polyhalogenated monoterpenes from the sea hare Aplysia punctata. J. Nat. Prod. 1997, 60, 482–484. [Google Scholar] [CrossRef]
- Gavagnin, M.; Marín, A.; Castelluccio, F.; Villani, G.; Cimino, G. Defensive relationships between Caulerpa prolifera and its shelled sacoglossan predators. J. Exp. Mar. Biol. Ecol. 1994, 175, 197–210. [Google Scholar] [CrossRef]
- Cimino, G.; Crispino, A.; Di Marzo, V.; Gavagnin, M.; Ros, J.D. Oxytoxins, bioactive molecules produced by the marine opisthobranch mollusc Oxynoe olivacea from a diet-derived precursor. Experientia 1990, 46, 767–770. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Montanaro, D.; Ortea, J.; Cimino, G. Chemical studies of Caribbean sacoglossans: Dietary relationships with green algae and ecological implications. J. Chem. Ecol. 2000, 26, 1563–1578. [Google Scholar] [CrossRef]
- Fontana, A.; Ciavatta, M.L.; Mollo, E.; Naik, C.D.; Wahidulla, S.; D’Sousa, L.; Cimino, G. Volvatellin, cauerpenyne-related product from the sacoglossan Volvatella sp. J. Nat. Prod. 1999, 62, 931–933. [Google Scholar] [CrossRef]
- Fischel, J.L.; Lemee, R.; Formento, P.; Caldani, C.; Moll, J.L.; Pesando, D.; Meinesz, A.; Grelier, P.; Pietra, P.; Guerriero, A. Cell growth inhibitory effects of caulerpenyne, a sesquiterpenoid from the marine algae Caulerpa taxifolia. Anticancer Res. 1995, 15, 2155–2160. [Google Scholar]
- Brunelli, M.; Garcia-Gil, M.; Mozzachiodi, R.; Roberto, M.; Scuri, R.; Traina, G.; Zaccardi, M.L. Neurotoxic effects of caulerpenyne. Prog. Neuropsychopharmacol. Biol. Psychiatry 2000, 24, 939–954. [Google Scholar] [CrossRef]
- Richter, P.; Schubert, G.; Schaible, A.M.; Cavas, L.; Werz, O.; Pohnert, G. Caulerpenyne and related bis-enol esters are novel-type inhibitors of human 5-lipoxygenase. ChemMedChem 2014, 9, 1655–1659. [Google Scholar] [CrossRef] [PubMed]
- Mehra, R.; Bhushan, S.; Bast, F.; Singh, S. Marine macroalga Caulerpa: Role of its metabolites in modulating cancer signaling. Mol. Biol. Rep. 2019, 46, 3545–3555. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.R. Evolution of the Sacoglossa (Mollusca, Opisthobranchia) and the ecological associations with their food plants. Evol. Ecol. 1997, 11, 301–335. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; López Gresa, M.P.; Gavagnin, M.; Manzo, E.; Mollo, E.; D’Souza, L.; Cimino, G. New caulerpenyne-derived metabolites of an Elysia sacoglossan from the south Indian coast. Molecules 2006, 11, 808–816. [Google Scholar] [CrossRef]
- Stierle, D.B.; Wing, R.M.; Sims, J.J. Marine natural products, XI Costatone and costatolide, new halogenated monoterpenes from the red seaweed, Plocamium costatum. Tetrahedron Lett. 1976, 49, 4455–4458. [Google Scholar] [CrossRef]
- Grkovic, T.; Appleton, D.R.; Copp, B.R. Chemistry and chemical ecology of some of the common opisthobranch molluscs found on the shores of NE New Zealand. Chem. New Zeal. 2005, 69, 12–15. [Google Scholar]
- Jongaramruong, J.; Blackman, A.J.; Skelton, B.W.; White, A.H. Chemical relationships between the sea hare Aplysia parvula and the red seaweed Laurencia filiformis from Tasmania. Austral. J. Chem. 2002, 55, 275–280. [Google Scholar] [CrossRef]
- Ginsburg, D.W.; Paul, V.J. Chemical defenses in the sea hare Aplysia parvula: Importance of diet and sequestration of algal secondary metabolites. Mar. Ecol. Progr. Ser. 2001, 215, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Ireland, C.; Faulkner, D.J.; Finer, J.; Clardy, J. A novel diterpene from Dollabella californica. J. Amer. Chem. Soc. 1976, 98, 4664–4665. [Google Scholar] [CrossRef]
- Ireland, C.; Faulkner, D.J. Diterpenes from Dolabella californica. J. Org. Chem. 1977, 42, 3157–3162. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Ode, R.h.; Herald, C.L.; von Dreele, R.B.; Michel, C. The isolation and structure of dolatriol. J. Amer. Chem. Soc. 1976, 98, 4677–4678. [Google Scholar] [CrossRef]
- Pettit, G.R.; Herald, C.L.; Ode, R.H.; Brown, P.; Gust, D.J.; Michel, C. The isolation of loliolide from an Indian Ocean opisthobranch mollusc. J. Nat. Prod. 1980, 43, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Manzo, E.; Carbone, M.; Mollo, E.; Irace, C.; Di Pascale, A.; Li, Y.; Ciavatta, M.L.; Cimino, G.; Guo, Y.-W.; Gavagnin, M. Structure and synthesis of a unique isonitrile lipid isolated from the marine mollusk Actinocyclus papillatus. Org. Lett. 2011, 13, 1897–1899. [Google Scholar] [CrossRef]
- Guo, Y.-W.; Gavagnin, M.; Carbone, M.; Mollo, E.; Cimino, G. Recent Sino-Italian collaborative studies on marine organisms from the South China Sea. Pure Appl. Chem. 2012, 84, 1391–1405. [Google Scholar] [CrossRef]
- Carbone, M.; Ciavatta, M.L.; Mathieu, V.; Ingels, A.; Kiss, R.; Pascale, P.; Mollo, E.; Ungur, N.; Guo, Y.-W.; Gavagnin, M. Marine terpenoid diacylguanidines: Structure, synthesis, and biological evaluation of naturally occurring actinofide and synthetic analogues. J. Nat. Prod. 2017, 80, 1339–1346. [Google Scholar] [CrossRef]
- Krug, P.J.; Boyd, K.G.; Faulkner, D.J. Isolation and synthesis of tanyolides A and B, metabolites of the nudibranch Sclerodoris tanya. Tetrahedron 1995, 51, 11063–11074. [Google Scholar] [CrossRef]
- Fontana, A.; Muniaín, C.; Cimino, G. First chemical study of patagonian nudibranchs: A new seco-11, 12-spongiane, tyrinnal, from the defensive organs of Tyrinna nobilis. J. Nat. Prod. 1998, 61, 1027–1029. [Google Scholar] [CrossRef]
- Mudianta, W.; White, A.M.; Suciati, P.L.K.; Krishnaraj, R.R.; Winters, A.E.; Mollo, E.; Cheney, K.L.; Garson, M.J. Chemoecological studies on marine natural products: Terpene chemistry from marine mollusks. Pure Appl. Chem. 2014, 86, 995–1002. [Google Scholar] [CrossRef] [Green Version]
- Putz, A.; Kehraus, S.; Díaz-Agras, G.; Wägele, H.; König, G.M. Dotofide, a guanidine-interrupted terpenoid from the marine slug Doto pinnatifida (Gastropoda, Nudibranchia). Eur. J. Org. Chem. 2011, 3733–3737. [Google Scholar] [CrossRef]
- De Silva, E.D.; Schwartz, R.E.; Scheuer, P.J. Srilankyene, a new metabolite from the sea hare Aplysia oculifera. J. Org. Chem. 1983, 48, 395–396. [Google Scholar] [CrossRef]
- Schulte, G.R.; Chung, M.C.H.; Scheuer, P.J. Two bicyclic C15 enynes from the sea hare Aplysia oculifera. J. Org. Chem. 1981, 46, 3870–3873. [Google Scholar] [CrossRef]
- McDonald, F.J.; Campbell, D.C.; Vanderah, D.J.; Schmitz, F.J.; Washecheck, D.M.; Burks, J.E.; van der Helm, D. Marine natural products. Dactylyne, an acetylenic dibromochloro ether from the sea hare Aplysia dactylomela. J. Org. Chem. 1975, 40, 665–666. [Google Scholar]
- Vanderah, D.J.; Schmitz, F.J. Marine natural products: Isodactylyne, a halogenated acetylenic ether from the sea hare Aplysia dactylomela. J. Org. Chem. 1976, 41, 3480–3481. [Google Scholar] [CrossRef]
- Gopichand, Y.; Schmitz, F.J.; Shelly, J.; Rahman, A.; van der Helm, D. Marine natural products: Halogenated acetylenic ethers from the sea hare Aplysia dactylomela. J. Org. Chem. 1981, 46, 5192–5197. [Google Scholar] [CrossRef]
- Schmitz, F.J.; McDonald, F.J. Marine natural products: Dactyloxene-B, a sesquiterpene ether from the sea hare, Aplysia dactylomela. Tetrahedron Lett. 1974, 29, 2541–2544. [Google Scholar] [CrossRef]
- Schmitz, F.J.; McDonald, F.J.; Vanderah, D.J. Marine natural products: Sesquiterpenes alcohols and ethers from the sea hare Aplysia dactylomela. J. Org. Chem. 1978, 43, 4220–4225. [Google Scholar] [CrossRef]
- Schmitz, F.J.; Michaud, D.P.; Hollenbeak, K.H. Marine natural products: Dihydroxydeodactol monoacetate, a halogenated sesquiterpene ether from the sea hare Aplysia dactylomela. J. Org. Chem. 1980, 45, 1525–1528. [Google Scholar] [CrossRef]
- Mancini, I.; Guella, G.; Pietra, F. Highly diastereoselective, biogenetically patterned synthesis of (+)-(1S,6R)-volvatellin (=(+)-(4R,5S)-5Hydroxy4-(5-methyl-1-methylenehex-4-en-2-ynyl)cyclohex-1-ene-1-carbaldehyde). Helv. Chim. Acta 2000, 83, 694–701. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Cimino, G.; Ortea, J. A new γ-dihydropyrone-propionate from the Caribbean ascoglossan Tridachia crispata. Tetrahedron Lett. 1996, 37, 4259–4262. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Castelluccio, F.; Montanaro, D.; Ortea, J.; Cimino, G. A novel dietary sesquiterpene from the marine sacoglossan Tridachia crispata. Nat. Prod. Lett. 1997, 10, 151–156. [Google Scholar] [CrossRef]
- Ireland, C.; Faulkner, D.J. The defensive secretion of the opisthobranch mollusc Onchidella binneyi. Bioorg. Chem. 1978, 7, 125–131. [Google Scholar] [CrossRef]
- Iken, K.; Avila, C.; Ciavatta, M.L.; Fontana, A.; Cimino, G. Hodgsonal, a new drimane sesquiterpene from the mantle of the Antarctic nudibranch Bathydoris hodgsoni. Tetrahedron Lett. 1998, 39, 5635–5638. [Google Scholar] [CrossRef]
- Avila, C.; Iken, K.; Fontana, A.; Cimino, G. Chemical ecology of the Antarctic nudibranch Bathydoris hodgsoni Eliot, 1907: Defensive role and origin of its natural products. J. Exp. Mar. Biol. Ecol. 2000, 252, 27–44. [Google Scholar] [CrossRef]
- Gavagnin, M.; De Napoli, A.; Castelluccio, F.; Cimino, G. Austrodorin-A and-B: First tricyclic diterpenoid 2′-monoglyceryl esters from an Antarctic nudibranch. Tetrahedron Lett. 1999, 40, 8471–8475. [Google Scholar] [CrossRef]
- Davies-Coleman, M.T.; Faulkner, D.J. New diterpenoic acid glycerides from the Antarctic nudibranch Austrodoris kerguelensis. Tetrahedron 1991, 47, 9743–9750. [Google Scholar] [CrossRef]
- Gavagnin, M.; Trivellone, E.; Castelluccio, F.; Cimino, G.; Cattaneo-Vietti, R. Glyceryl ester of a new halimane diterpenoic acid from the skin of the antarctic nudibranch Austrodoris kerguelenensis. Tetrahedron Lett. 1995, 36, 7319–7322. [Google Scholar] [CrossRef]
- Gavagnin, M.; Carbone, M.; Mollo, E.; Cimino, G. Austrodoral and austrodoric acid: Nor-sesquiterpenes with a new carbon skeleton from the Antarctic nudibranch Austrodoris kerguelenensis. Tetrahedron Lett. 2003, 44, 1495–1498. [Google Scholar] [CrossRef]
- Gavagnin, M.; Carbone, M.; Mollo, E.; Cimino, G. Further chemical studies on the Antarctic nudibranch Austrodoris kerguelenensis: New terpenoid acylglycerols and revision of the previous stereochemistry. Tetrahedron 2003, 59, 5579–5583. [Google Scholar] [CrossRef]
- Fontana, A.; Ciavatta, M.L.; Miyamoto, T.; Spinella, A.; Cimino, G. Biosynthesis of drimane terpenoids in dorid molluscs: Pivotal role of 7-deacetoxyolepupuane in two species of Dendrodoris nudibranchs. Tetrahedron 1999, 55, 5937–5946. [Google Scholar] [CrossRef]
- Sakio, Y.; Hirano, Y.J.; Hayashi, M.; Komiyama, K.; Ishibashi, M. Dendocarbins A–N, new drimane sesquiterpenes from the nudibranch Dendrodoris carbunculosa. J. Nat. Prod. 2001, 64, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Gavagnin, M.; Mollo, E.; Calado, G.; Fahey, S.; Ghiselin, M.T.; Ortea, J.; Cimino, G. Chemical studies of porostome nudibranchs: Comparative and ecological aspects. Chemoecology 2001, 11, 131–136. [Google Scholar] [CrossRef]
- Avila, C.; Cimino, G.; Crispino, A.; Spinella, A. Drimane sesquiterpenoids in Mediterranean Dendrodoris nudibranchs: Anatomical distribution and biological role. Experientia 1991, 47, 306–310. [Google Scholar] [CrossRef]
- Gaspar, H.; Gavagnin, M.; Calado, G.; Castelluccio, F.; Mollo, E.; Cimino, G. Pelseneeriol-1 and-2: New furanosesquiterpene alcohols from porostome nudibranch Doriopsilla pelseneeri. Tetrahedron 2005, 61, 11032–11037. [Google Scholar] [CrossRef]
- Spinella, A.; Álvarez, L.A.; Avila, C.; Cimino, G. New acetoxy-ent-pallescensin-A sesquiterpenoids from the skin of the porostome nudibranch Doriopsilla areolata. Tetrahedron Lett. 1994, 35, 8665–8668. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Castelluccio, F.; Ghiselin, M.T.; Calado, G.; Cimino, G. Can molluscs biosynthesize typical sponge metabolites? The case of the nudibranch Doriopsilla areolata. Tetrahedron 2001, 57, 8913–8916. [Google Scholar]
- Fontana, A.; Tramice, A.; Cutignano, A.; d’Ippolito, G.; Gavagnin, M.; Cimino, G. Terpene biosynthesis in the nudibranch Doriopsilla areolata. J. Org. Chem. 2003, 68, 2405–2409. [Google Scholar] [CrossRef]
- Gaspar, H.; Cutignano, A.; Ferreira, T.; Calado, G.; Cimino, G.; Fontana, A. Biosynthetic evidence supporting the generation of terpene chemodiversity in marine mollusks of the genus Doriopsilla. J. Nat. Prod. 2008, 71, 2053–2056. [Google Scholar] [CrossRef]
- Long, J.D.; Hay, M.E. Fishes learn aversions to a nudibranch’s chemical defense. Mar. Ecol. Progr. Ser. 2006, 307, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Fusetani, N.; Wolstenholme, H.J.; Matsunaga, S.; Hirota, H. Two new sesquiterpene isonitriles from the nudibranch Phyllidia pustulosa. Tetrahedron Lett. 1991, 32, 7291–7294. [Google Scholar] [CrossRef]
- Okino, T.; Yoshimura, E.; Hirota, H.; Fusetani, N. New antifouling sesquiterpenes from four nudibranchs of the family Phyllidiidae. Tetrahedron 1996, 52, 9447–9454. [Google Scholar] [CrossRef]
- Hirota, H.; Okino, T.; Yoshimura, E.; Fusetani, N. Five new antifouling sesquiterpenes from two marine sponges of the genus Axinyssa and the nudibranch Phyllidia pustulosa. Tetrahedron 1998, 54, 13971–13980. [Google Scholar] [CrossRef]
- Cimino, G.; Fontana, A.; Gavagnin, M. Marine opisthobranch molluscs: Chemistry and ecology in sacoglossan and dorids. Curr. Org. Chem. 1999, 3, 327–372. [Google Scholar]
- Garson, M.J.; Simpson, J.S. Marine isocyanides and related natural products—Structure, biosynthesis and ecology. Nat. Prod. Rep. 2004, 21, 164–179. [Google Scholar] [CrossRef]
- Dumdei, E.J.; Flowers, A.E.; Garson, M.J.; Moore, C.J. The biosynthesis of sesquiterpene isocyanides and isothiocyanates in the marine sponge Acanthella cavernosa (Dendy); evidence for dietary transfer to the dorid nudibranch Phyllidiella pustulosa. Comp. Biochem. Physiol. 1997, 118, 1385–1392. [Google Scholar] [CrossRef]
- Manzo, E.; Ciavatta, M.L.; Gavagnin, M.; Mollo, E.; Guo, Y.-W.; Cimino, G. Isocyanide terpene metabolites of Phyllidiella pustulosa, a nudibranch from the South China Sea. J. Nat. Prod. 2004, 67, 1701–1704. [Google Scholar] [CrossRef]
- Shimomura, M.; Miyaoka, H.; Yamada, Y. Absolute configuration of marine diterpenoid kalihinol A. Tetrahedron Lett. 1999, 40, 8015–8017. [Google Scholar] [CrossRef]
- Lyakhova, E.G.; Kolesnikova, S.A.; Kalinovskii, A.I.; Stonik, V.A. Secondary metabolites of the Vietnamese nudibranch mollusc Phyllidiella pustulosa. Chem. Nat. Comp. 2010, 46, 534–538. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, W.-T.; Li, S.-W.; Ye, J.-Y.; Huan, X.-J.; Gavagnin, M.; Yao, L.-G.; Wang, H.; Miao, Z.-H.; Li, X.-W.; et al. Cytotoxic nitrogenous terpenoids from two South China Sea nudibranchs Phyllidiella pustulosa, Phyllidia coelestis, and their sponge-prey Acanthella cavernosa. Mar. Drugs 2019, 17, 56. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.D. GC-MS and NMR analysis of Phyllidiella pustulosa and one of its dietary sources, the sponge Phakellia carduus. Comp. Biochem. Physiol. 2003, 134, 307–313. [Google Scholar] [CrossRef]
- König, G.M.; Wright, A.D.; Franzblau, S.G. Assessment of antimycobacterial activity of a series of mainly marine derived natural products. Planta Med. 2000, 66, 337–342. [Google Scholar] [CrossRef]
- Fusetani, N.; Hirota, H.; Okino, T.; Tomono, Y.; Yoshimura, E. Antifouling activity of isocyanoterpenoids and related compounds isolated from a marine sponge and nudibranchs. J. Nat. Toxins 1996, 5, 249–259. [Google Scholar]
- Sim, D.C.-M.; Mudianta, I.W.; White, A.M.; Martiningsih, N.W.; Loh, J.J.M.; Cheney, K.L.; Garson, M.J. New sesquiterpenoid isonitriles from three species of phyllidid nudibranchs. Fitoterapia 2018, 126, 69–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwashima, M.; Terada, I.; Iguchi, K.; Yamori, T. New biologically active marine sesquiterpenoid and steroid from the Okinawan sponge of the genus Axinyssa. Chem. Pharm. Bull. 2002, 50, 1286–1289. [Google Scholar] [CrossRef] [Green Version]
- Gulavita, N.K.; De Silva, E.D.; Hagadone, M.R.; Karuso, P.; Scheuer, P.J.; van Duyne, G.D.; Clardy, J. Nitrogenous bisabolene sesquiterpenes from marine invertebrates. J. Org. Chem. 1986, 51, 5136–5139. [Google Scholar] [CrossRef]
- Kitano, Y.; Ito, T.; Suzuki, T.; Nogata, Y.; Shinshima, K.; Yoshimura, E.; Chiba, K.; Tada, M.; Sakaguchi, I. Synthesis and antifouling activity of 3-isocyanotheonellin and its analogues. J. Chem. Soc. Perkin Trans. 2002, 1, 2251–2255. [Google Scholar] [CrossRef]
- Yasman, Y.; Edrada, R.A.; Wray, V.; Proksch, P. New 9-thiocyanatopupukeanane sesquiterpenes from the nudibranch Phyllidia varicosa and its sponge-prey Axinyssa aculeata. J. Nat. Prod. 2003, 66, 1512–1514. [Google Scholar] [CrossRef]
- Jaisamut, S.; Prabpai, S.; Tancharoen, C.; Yuenyongsawad, S.; Hannongbua, S.; Kongsaeree, P.; Plubrukarn, A. Bridged tricyclic sesquiterpenes from the tubercle nudibranch Phyllidia coelestis Bergh. J. Nat. Prod. 2013, 76, 2158–2161. [Google Scholar] [CrossRef]
- Ritson-Williams, R.; Paul, V.J. Marine benthic invertebrates use multimodal cues for defense against reef fish. Mar. Ecol. Progr. Ser. 2007, 340, 29–39. [Google Scholar] [CrossRef]
- Tanaka, J.; Higa, T. Two new cytotoxic carbonimidic dichlorides from the nudibranch Reticulidia fungia. J. Nat. Prod. 1999, 62, 1339–1340. [Google Scholar] [CrossRef]
- Zhang, W.; Gavagnin, M.; Guo, Y.-W.; Mollo, E.; Ghiselin, M.T.; Cimino, G. Terpenoid metabolites of the nudibranch Hexabranchus sanguineus from the South China Sea. Tetrahedron 2007, 63, 4725–4729. [Google Scholar] [CrossRef]
- Hellou, J.; Andersen, R.J.; Thompson, J.E. Terpenoids from the dorid nudibranch Cadlina luteomarginata. Tetrahedron 1982, 38, 1875–1879. [Google Scholar] [CrossRef]
- Nakano, T.; Villamizar, J.; Maillo, M.A. A new entry to (+)-albicanol and (+)-bicyclofarnesol. J. Chem. Res. Synop. 1995, 1, 330–331. [Google Scholar]
- Barrero, A.F.; Manzaneda, E.A.; Altarejos, J.; Salido, S.; Ramos, J.M.; Simmonds, M.S.J.; Blaney, W.M. Synthesis of biologically active drimanes and homodrimanes from (−)-sclareol. Tetrahedron 1995, 51, 7435–7450. [Google Scholar] [CrossRef]
- Fontana, A.; Gavagnin, M.; Mollo, E.; Trivellone, E.; Ortea, J.; Cimino, G. Chemical studies of Cadlina molluscs from the Cantabrian sea (Atlantic Ocean). Comp. Biochem. Physiol. 1995, 111, 283–290. [Google Scholar] [CrossRef]
- Dumdei, E.J.; Kubanek, J.; Coleman, J.E.; Pika, J.; Andersen, R.J.; Steiner, J.R.; Clardy, J. New terpenoid metabolites from the skin extracts, an egg mass, and dietary sponges of the Northeastern Pacific dorid nudibranch Cadlina luteomarginata. Can. J. Chem. 1997, 75, 773–789. [Google Scholar] [CrossRef] [Green Version]
- Kubanek, J.; Graziani, E.I.; Andersen, R.J. Investigations of terpenoid biosynthesis by the dorid nudibranch Cadlina luteomarginata. J. Org. Chem. 1997, 62, 7239–7246. [Google Scholar] [CrossRef]
- Johnson, R.F.; Gosliner, T.M. Traditional taxonomic groupings mask evolutionary history: A molecular phylogeny and new classification of the chromodorid nudibranchs. PLoS ONE 2012, 7, e33479. [Google Scholar] [CrossRef] [Green Version]
- Gaspar, H.; Rodrigues, A.I.; Calado, G. Comparative study of chemical defences from two allopatric north atlantic subspecies of Hypselodoris picta (Mollusca: Opisthobranchia). Açoreana 2009, 6, 137–143. [Google Scholar]
- McPhail, K.L.; Davies-Coleman, M.T.; Coetzee, P. A new furanosesterterpene from the South African nudibranch Hypselodoris capensis and a Dictyoceratida sponge. J. Nat. Prod. 1998, 61, 961–964. [Google Scholar] [CrossRef]
- Karuso, P.; Scheuer, P.J. Natural products from three nudibranchs: Nembrotha kubaryana, Hypselodoris infucata and Chromodoris petechialis. Molecules 2002, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mollo, E.; Gavagnin, M.; Carbone, M.; Guo, Y.-W.; Cimino, G. Chemical studies on Indopacific Ceratosoma nudibranchs illuminate the protective role of their dorsal horn. Chemoecology 2005, 15, 31–36. [Google Scholar] [CrossRef]
- Winters, A.E.; White, A.M.; Dewi, A.S.; Mudianta, I.W.; Wilson, N.G.; Forster, L.C.; Garson, M.J.; Cheney, K.L. Distribution of defensive metabolites in nudibranch molluscs. J. Chem. Ecol. 2018, 44, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Cronin, G.; Hay, M.; Fenical, W.; Lindquist, N. Distribution, density, and sequestration of host chemical defenses by the specialist nudibranch Tritonia hamnerorum found at high densities on the sea fan Gorgonia ventalina. Mar. Ecol. Prog. Ser. 1995, 119, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.E.; Andersen, R.J. Terpenoid metabolites from skin extracts of the dendronotid nudibranch Tochuina tetraquetra. Can. J. Chem. 1987, 65, 2244–2247. [Google Scholar] [CrossRef]
- Pika, J.; Faulkner, D.J. Four sesquiterpenes from the South African nudibranch Leminda millecra. Tetrahedron 1994, 50, 3065–3070. [Google Scholar] [CrossRef]
- McPhail, K.L.; Davies-Coleman, M.T.; Starmer, J. Sequestered chemistry of the arminacean nudibranch Leminda millecra in Algoa Bay, South Africa. J. Nat. Prod. 2001, 64, 1183–1190. [Google Scholar] [CrossRef]
- Ishibashi, M.; Yamaguchi, Y.; Hirano, Y.J. Bioactive natural products from nudibranchs. In Biomaterials from Aquatic and Terrestrial Organisms; Fingerman, M., Nagabhushanam, R., Eds.; Science Publishers: Enfield, NH, USA, 2006; pp. 513–535. [Google Scholar]
- Affeld, S.; Kehraus, S.; Wägele, H.; König, G.M. Dietary derived sesquiterpenes from Phyllodesmium lizardensis. J. Nat. Prod. 2009, 72, 298–300. [Google Scholar] [CrossRef]
- Mao, S.C.; Gavagnin, M.; Mollo, E.; Guo, Y.-W. A new rare asteriscane sesquiterpene and other related derivatives from the Hainan aeolid nudibranch Phyllodesmium magnum. Biochem. System. Ecol. 2011, 39, 408–411. [Google Scholar] [CrossRef]
- Slattery, M.; Avila, C.; Starmer, J.; Paul, V.J. A sequestered soft coral diterpene in the aeolid nudibranch Phyllodesmium guamensis. J. Exp. Mar. Biol. Ecol. 1998, 226, 33–49. [Google Scholar] [CrossRef]
- Manzo, E.; Ciavatta, M.L.; Gavagnin, M.; Puliti, R.; Mollo, E.; Guo, Y.-W.; Mattia, C.A.; Mazzarella, L.; Cimino, G. Structure and absolute stereochemistry of novel C15-halogenated acetogenins from the anaspidean mollusc Aplysia dactylomela. Tetrahedron 2005, 61, 7456–7460. [Google Scholar] [CrossRef]
- Faulkner, D.J.; Stallard, M.O. 7-chloro-3,7-dimethyl-1,4,6-trobromo-1-octen-3-ol, a novel monoterpene alcohol from Aplysia californica. Tetrahedron Lett. 1973, 14, 1171–1174. [Google Scholar] [CrossRef]
- Faulkner, D.J.; Stallard, M.O.; Fayos, J.; Clardy, J. (3R,4S,7S)- trans, trans-3,7-dimethyl-1,8,8- tribromo-3,4,7-trichloro-1,5-octadiene, a novel monoterpene from the sea hare, Aplysia californica. J. Am. Chem. Soc. 1973, 95, 3413–3414. [Google Scholar] [CrossRef]
- Stallard, M.O.; Faulkner, D.J. Chemical constituents of the digestive gland of the sea hare Aplysia californica-II. Chemical transformations. Comp. Biochem. Physiol. B Biochem. Molec. Biol. 1974, 49, 37–41. [Google Scholar] [CrossRef]
- Ireland, C.; Stallard, M.O.; Faulkner, D.J. Some chemical constituents of the digestive gland of the sea hare Aplysia californica. J. Org. Chem. 1976, 41, 2461–2465. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, E.; Nappo, M.; Avila, C.; Vagias, C.; Roussis, V. Metabolites from the sea hare Aplysia fasciata. J. Nat. Prod. 2009, 72, 1716–1719. [Google Scholar] [CrossRef]
- Mudianta, W.I.; Martiningsih, N.W.; Dodik Prasetia, I.N.; Nursid, M. Bioactive terpenoid from the balinese nudibranch Hypselodoris infucata. Indonesian J. Pharm. 2016, 27, 104–110. [Google Scholar] [CrossRef]
- Fontana, A.; Ciavatta, M.L.; D’Souza, L.; Mollo, E.; Naik, C.G.; Parameswaran, P.S.; Wahidulla, S.; Cimino, G. Selected chemo-ecological studies of marine opisthobranchs from Indian coasts. J. Indian Instit. Sci. 2001, 81, 403–415. [Google Scholar]
- Pereira, F.R.; Berlinck, R.G.S.; Rodrigues Filho, E.; Veloso, K.; Ferreira, A.G.; Padula, V. Metabólitos Secundários dos nudibrânquios Tambja stegosauriformis, Hypselodoris lajensis e Okenia zoobotryon e dos briozoários Zoobotryon verticillatum e Bugula dentata da costa do Brasil. Quim. Nova 2012, 35, 2194–2201. [Google Scholar] [CrossRef] [Green Version]
- Yamamura, S.; Hirata, Y. Structures of aplysin and aplysinol, naturally occurring bromo-compounds. Tetrahedron 1963, 19, 1485–1496. [Google Scholar] [CrossRef]
- Díaz-Marrero, A.R.; De La Rosa, J.M.; Brito, I.; Darias, J.; Cueto, M. Dactylomelatriol, a biogenetically intriguing omphalane-derived marine sesquiterpene. J. Nat. Prod. 2012, 75, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Valdés, A. The ring doesn’t mean a thing: Molecular data suggest a new taxonomy for two Pacific species of sea hares (Mollusca: Opisthobranchia, Aplysiidae). Pacific Sci. 2013, 67, 283–294. [Google Scholar] [CrossRef]
- Suciati, S.; Lambert, L.K.; Garson, M.J. Structures and anatomical distribution of oxygenated diterpenes in the Australian nudibranch Chromodoris reticulata. Aust. J. Chem. 2011, 64, 757–765. [Google Scholar] [CrossRef]
- Gavagnin, M.; Spinella, A.; Crispino, A.; Epifanio, R.D.A.; Marn, A.; Cimino, G. Chemical-components of the Mediterranean ascoglossan Thuridilla hopei. Gazz. Chim. Ital. 1993, 123, 205–208. [Google Scholar]
- Carbone, M.; Ciavatta, M.L.; de Rinaldis, G.; Castelluccio, F.; Mollo, E.; Gavagnin, M. Identification of thuridillin-related aldehydes from Mediterranean sacoglossan mollusk Thuridilla hopei. Tetrahedron 2014, 70, 3770–3773. [Google Scholar] [CrossRef]
- Gavagnin, M.; Marín, A.; Mollo, E.; Crispino, A.; Villani, G.; Cimino, G. Secondary metabolites from Mediterranean Elysioidea: Origin and biological role. Comp. Biochem. Physiol. B 1994, 108, 107–115. [Google Scholar] [CrossRef]
- Somerville, M.J.; Katavic, P.L.; Lambert, L.K.; Pierens, G.K.; Blanchfield, J.T.; Cimino, G.; Mollo, E.; Gavagnin, M.; Banwell, M.G.; Garson, M.J. Isolation of thuridillins D-F, diterpene metabolites from the Australian sacoglossan mollusk Thuridilla splendens; relative configuration of the epoxylactone ring. J. Nat. Prod. 2012, 75, 1618–1624. [Google Scholar] [CrossRef]
- Paul, V.J.; Fenical, W. Bioactive terpenoids from Caribbean marine algae of the genera Penicillus and Udotea (Chlorophyta). Tetrahedron 1984, 40, 2913–2918. [Google Scholar] [CrossRef]
- Paul, V.J.; Sun, H.H.; Fenical, W. Udoteal, a linear diterpenoid feeding deterrent from the tropical green alga Udotea flabellum. Phytochemistry 1982, 21, 468–469. [Google Scholar] [CrossRef]
- Coll, J.; Bowden, B.; Tapiolas, D.; Willis, R.; Djura, P.; Streamer, M.; Trott, L. Studies of Australian soft corals—XXXV: The terpenoid chemistry of soft corals and its implications. Tetrahedron 1985, 41, 1085–1092. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; García-Matucheski, S.; Carbone, M.; Villani, G.; Nicotera, M.R.; Muniain, C.; Gavagnin, M. Chemistry of two distinct aeolid Spurilla species: Ecological implications. Chem. Biodivers. 2017, 14, e1700125. [Google Scholar] [CrossRef] [PubMed]
- Petraki, A.; Ioannou, E.; Papazafiri, P.; Roussis, V. Dactylomelane diterpenes from the sea hare Aplysia depilans. J. Nat. Prod. 2015, 78, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.J.; Fenical, W. Isolation of halimedatrial: Chemical defense adaptation in the calcareous reef-building alga Halimeda. Science 1983, 221, 747–749. [Google Scholar] [CrossRef] [PubMed]
- Paul, V.J.; van Alstyne, K.L. Use of ingested algal diterpenoids by Elysia halimedae Macnae (Opisthobranchia: Ascoglossa) as antipredator defenses. J. Exp. Mar. Biol. Ecol. 1988, 119, 15–29. [Google Scholar] [CrossRef]
- Paul, V.J.; van Alstyne, K.L. Activation of chemical defenses in the tropical green algae Halimeda spp. J. Exp. Mar. Biol. Ecol. 1992, 160, 191–203. [Google Scholar] [CrossRef]
- White, A.M.; Dao, K.; Vrubliauskas, D.; Könst, Z.A.; Pierens, G.K.; Mándi, A.; Andrews, K.T. Skinner-Adams, T.S.; Clarke, M.E.; Narbutas, P.T.; et al. Catalyst-controlled stereoselective synthesis secures the structure of the antimalarial isocyanoterpene pustulosaisonitrile-1. J. Org. Chem. 2017, 82, 13313–13323. [Google Scholar] [CrossRef]
- Ciavatta, M.L.; Villani, G.; Trivellone, E.; Cimino, G. Two new labdane aldehydes from the skin of the notaspidean Pleurobranchaea meckelii. Tetrahedron Lett. 1995, 36, 8673–8676. [Google Scholar] [CrossRef]
- Kladi, M.; Ntountaniotis, D.; Zervou, M.; Vagias, C.; Ioannou, E.; Roussis, V. Glandulaurencianols A-C, brominated diterpenes from the red alga, Laurencia glandulifera and the sea hare, Aplysia punctata. Tetrahedron Lett. 2014, 55, 2835–2837. [Google Scholar] [CrossRef]
- Findlay, J.A.; Li, G. Novel terpenoids from the sea hare Aplysia punctata. Can. J. Chem. 2002, 80, 1697–1707. [Google Scholar] [CrossRef]
- Caccamese, S.; Toscano, R.M.; Cerrini, S.; Gavuzzo, E. Laurencianol, a new halogenated diterpenoid from the marine alga Laurencia obtusa. Tetrahedron Lett. 1982, 23, 114–116. [Google Scholar] [CrossRef]
- Minale, L.; Riccio, R. Constituents of the digestive gland of the molluscs of the genus Aplysia—I. Novel diterpenes from Aplysia depilans. Tetrahedron Lett. 1976, 31, 2711–2714. [Google Scholar] [CrossRef]
- Di Prisco, C.L.; Dessi’ Fulgheri, F.; Tomasucci, M. Identification and biosynthesis of steroids in the marine mollusc Aplysia depilans. Comp. Biochem. Physiol. B 1973, 45, 303–310. [Google Scholar] [CrossRef]
- Jiménez, C.; Quiñoá, E.; Castedo, L.; Riguera, R. Epidioxy sterols from the tunicates Dendrodoa grossularia and Ascidiella aspersa and the Gastropoda Aplysia depilans and Aplysia punctata. J. Nat. Prod. 1986, 49, 905–909. [Google Scholar] [CrossRef]
- Yamamura, S.; Hirata, Y. A naturally-occurring bromo-compound, aplysin-20 from Aplysia kurodai. Bull. Chem. Soc. Jpn. 1971, 44, 2560–2562. [Google Scholar] [CrossRef] [Green Version]
- Yamamura, S.; Terada, Y. Isoaplysin-20, a natural bromine-containing diterpene, from Aplysia kurodai. Tetrahedron Lett. 1977, 25, 2171–2172. [Google Scholar] [CrossRef]
- Ojika, M.; Yoshida, Y.; Okumura, M.; Ieda, S.; Yamada, K. Aplysiadiol, a new brominated diterpene from the marine mollusc Aplysia kurodai. J. Nat. Prod. 1990, 53, 1619–1622. [Google Scholar] [CrossRef]
- Ojika, M.; Kigoshi, H.; Yoshikawa, K.; Nakayama, Y.; Yamada, K. A new bromo diterpene, epi-aplysin-20, and ent-isoconcinndiol from the marine mollusc Aplysia kurodai. Bull. Chem. Soc. Jpn. 1992, 65, 2300–2302. [Google Scholar] [CrossRef]
- Miyamoto, T.; Higuchi, R.; Komori, T. Isolation and structures of aplykurodins A and B, two new isoprenoids from the marine mollusk Aplysia kurodai. Tetrahedron Lett. 1986, 27, 1153–1156. [Google Scholar] [CrossRef]
- Machida, K.; Matsumoto, T.; Fusetani, N.; Nakao, Y. Dolabellol A, a new halogenated diterpene isolated from the Opisthobranch Dolabella auricularia. Chem. Lett. 2017, 46, 1676–1678. [Google Scholar] [CrossRef] [Green Version]
- Gray, C.A.; Davies-Coleman, M.T.; McQuaid, C. Labdane diterpenes from the South African marine pulmonate Trimusculus costatus. Nat. Prod. Lett. 1998, 12, 47–53. [Google Scholar] [CrossRef]
- Manker, D.C.; Faulkner, D.J. Investigation of the role of diterpenes produced by marine pulmonates Trimusculus reticulatus and T. conica. J. Chem. Ecol. 1996, 22, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Marrero, A.R.; Dorta, E.; Cueto, M.; Rovirosa, J.; San-Martín, A.; Loyola, A.; Darias, J. Labdane diterpenes with a new oxidation pattern from the marine pulmonate Trimusculus peruvianus. Tetrahedron 2003, 59, 4805–4809. [Google Scholar] [CrossRef]
- Rovirosa, J.; Quezada, E.; San-Martin, A. New diterpene from the mollusc Trimusculus peruvianus. Bol. Soc. Chil. Quim. 1992, 37, 143–145. [Google Scholar]
- Van Wyk, A.W.W.; Gray, C.A.; Whibley, C.E.; Osoniyi, O.; Hendricks, D.T.; Caira-Mino, R.; Davies-Coleman, M.T. Bioactive metabolites from the South African marine mollusk Trimusculus costatus. J. Nat. Prod. 2008, 71, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Gavagnin, M.; Ungur, N.; Castelluccio, F.; Muniain, C.; Cimino, G. New minor diterpenoid diacylglycerols from the skin of the nudibranch Anisodoris fontaini. J. Nat. Prod. 1999, 62, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Pika, J.; Faulkner, D.J. Unusual chlorinated homo-diterpenes from the South African nudibranch Chromodoris hamiltoni. Tetrahedron 1995, 51, 8189–8198. [Google Scholar] [CrossRef]
- Tanaka, J.; Higa, T. The absolute configuration of kurospongin a new furanoterpene from a marine sponge, Spongia sp. Tetrahedron 1988, 44, 2805–2810. [Google Scholar] [CrossRef]
- Kashman, Y.; Croweiss, A.; Shmueli, U. Latrunculin, a new 2-thiazolidinone macrolide from the marine sponge Latrunculia magnifica. Tetrahedron Lett. 1980, 21, 3629–3632. [Google Scholar] [CrossRef]
- Kashman, Y.; Croweiss, A.; Kidor, R.; Blasberger, D.; Carmely, S. Latrunculins: NMR study, two new toxins and a synthetic approach. Tetrahedron 1985, 41, 1905–1914. [Google Scholar] [CrossRef]
- Forster, L.C.; White, A.M.; Cheney, K.L.; Garson, M.J. Oxygenated terpenes from the Indo-Pacific nudibranchs Goniobranchus splendidus and Goniobranchus collingwoodi. Nat. Prod. Comm. 2018, 13, 299–302. [Google Scholar] [CrossRef] [Green Version]
- White, A.M.; Pierens, G.K.; Forster, L.C.; Winters, A.E.; Cheney, K.L.; Garson, M.J. Rearranged diterpenes and norditerpenes from three Australian Goniobranchus mollusks. J. Nat. Prod. 2016, 79, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Winters, A.E.; White, A.M.; Cheney, K.L.; Garson, M.J. Geographic variation in diterpene-based secondary metabolites and level of defence in an aposematic nudibranch, Goniobranchus splendidus. J. Moll. Stud. 2019, 85, 133–142. [Google Scholar] [CrossRef]
- Carbone, M.; Ciavatta, M.L.; Manzo, E.; Li, X.-L.; Mollo, E.; Mudianta, I.W.; Guo, Y.-W.; Gavagnin, M. Amphilectene diterpene isonitriles and formamido derivatives from the Hainan nudibranch Phyllidia coelestis. Mar. Drugs 2019, 17, 603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavagnin, M.; Ungur, N.; Castelluccio, F.; Cimino, G. Novel verrucosins from the skin of the Mediterranean nudibranch Doris verrucosa. Tetrahedron 1997, 53, 1491–1504. [Google Scholar] [CrossRef]
- Cimino, G.; Gavagnin, M.; Sodano, G.; Puliti, R.; Mattia, C.A.; Mazzarella, L. Verrucosin-A and-B, ichthyotoxic diterpenoic acid glycerides with a new carbon skeleton from the dorid nudibranch Doris verrucosa. Tetrahedron 1988, 44, 2301–2310. [Google Scholar] [CrossRef]
- Gavagnin, M.; Spinella, A.; Cimino, G.; Sodano, G. Stereochemistry of ichthyotoxic diacylglycerols from opisthobranch molluscs. Tetrahedron Lett. 1990, 31, 6093–6094. [Google Scholar] [CrossRef]
- Fontana, A.; Tramice, A.; Cutignano, A.; d’Ippolito, G.; Renzulli, L.; Cimino, G. Studies of the biogenesis of verrucosins, toxic diterpenoid glycerides of the Mediterranean mollusc Doris verrucosa. Eur. J. Org. Chem. 2003, 3104–3108. [Google Scholar] [CrossRef]
- Young, R.M.; Baker, B.J. The defensive chemistry of the Irish nudibranch Archidoris pseudoargus (Gastropoda, Opisthobranchia). Planta Med. 2015, 81, PQ26. [Google Scholar] [CrossRef]
- Gavagnin, M.; De Napoli, A.; Cimino, G.; Iken, K.; Avila, C.; Garcia, F.J. Absolute configuration of diterpenoid diacylglycerols from the Antarctic nudibranch Austrodoris kerguelenensis. Tetrahedron Asym. 1999, 10, 2647–2650. [Google Scholar] [CrossRef]
- Iken, K.; Avila, C.; Fontana, A.; Gavagnin, M. Chemical ecology and origin of defensive compounds in the Antarctic nudibranch Austrodoris kerguelenensis (Opisthobranchia: Gastropoda). Mar. Biol. 2002, 141, 101–109. [Google Scholar]
- Diyabalanage, T.; Iken, K.B.; McClintock, J.B.; Amsler, C.D.; Baker, B.J. Palmadorins A− C, diterpene glycerides from the Antarctic nudibranch Austrodoris kerguelenensis. J. Nat. Prod. 2010, 73, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Maschek, J.A.; Mevers, E.; Diyabalanage, T.; Chen, L.; Ren, Y.; McClintock, J.B.; Amsler, C.D.; Wu, J.; Baker, B.J. Palmadorin chemodiversity from the Antarctic nudibranch Austrodoris kerguelenensis and inhibition of Jak2/STAT5-dependent HEL leukemia cells. Tetrahedron 2012, 68, 9095–9104. [Google Scholar] [CrossRef]
- Cutignano, A.; Zhang, W.; Avila, C.; Cimino, G.; Fontana, A. Intrapopulation variability in the terpene metabolism of the Antarctic opisthobranch mollusc Austrodoris kerguelenensis. Eur. J. Org. Chem. 2011, 5383–5389. [Google Scholar] [CrossRef]
- Wilson, N.G.; Maschek, J.A.; Baker, B.J. A species flock driven by predation? Secondary metabolites support diversification of slugs in Antarctica. PLoS ONE 2013, 8, e80277. [Google Scholar] [CrossRef] [Green Version]
- Moles, J.; Wägele, H.; Cutignano, A.; Fontana, A.; Ballesteros, M.; Avila, C. Giant embryos and hatchlings of Antarctic nudibranchs (Mollusca: Gastropoda: Heterobranchia). Mar. Biol. 2017, 164, 114–127. [Google Scholar] [CrossRef]
- Bogdanov, A.; Kehraus, S.; Bleidissel, S.; Preisfeld, G.; Schillo, D.; Piel, J.; Brachmann, A.O.; Wägele, H.; König, G.M. Defense in the Aeolidoidean genus Phyllodesmium (Gastropoda). J. Chem. Ecol. 2014, 40, 1013–1024. [Google Scholar] [CrossRef]
- Bogdanov, A.; Hertzer, C.; Kehraus, S.; Nietzer, S.; Rohde, S.; Schupp, P.J.; Wägele, H.; König, G.M. Defensive diterpene from the Aeolidoidean Phyllodesmium longicirrum. J. Nat. Prod. 2016, 79, 611–615. [Google Scholar] [CrossRef]
- Bogdanov, A.; Hertzer, C.; Kehraus, S.; Nietzer, S.; Rohde, S.; Schupp, P.J.; Wägele, H.; König, G.M. Secondary metabolome and its defensive role in the aeolidoidean Phyllodesmium longicirrum (Gastropoda, Heterobranchia, Nudibranchia). Beilstein J. Org. Chem. 2017, 13, 502–519. [Google Scholar] [CrossRef] [Green Version]
- Ciavatta, M.L.; Manzo, E.; Mollo, E.; Mattia, C.A.; Tedesco, C.; Irace, C.; Guo, Y.-W.; Li, X.-B.; Cimino, G.; Gavagnin, M. Tritoniopsins A–D, cladiellane-based diterpenes from the South China Sea nudibranch Tritoniopsis elegans and its prey Cladiella krempfi. J. Nat. Prod. 2011, 74, 1902–1907. [Google Scholar] [CrossRef]
- Zhang, W.; Gavagnin, M.; Guo, Y.-W.; Mollo, E.; Cimino, G. Chemical studies on the South China Sea nudibranch Dermatobranchus ornatus and its suggested prey gorgonian Muricella sp. Chin. J. Org. Chem. 2006, 26, 1667–1672. [Google Scholar]
- Hochlowski, J.E.; Faulkner, D.J. A diterpene related to cladiellin from a Pacific soft coral. Tetrahedron Lett. 1980, 21, 4055–4056. [Google Scholar] [CrossRef]
- Guerriero, A.; D’Ambrosio, M.; Pietra, F. Verecynarmin A, a novel briarane diterpenoid isolated from both the Mediterranean nudibranch mollusc Armina maculata and its prey, the pennatulacean octocoral Veretillum cynomorium. Helv. Chim. Acta 1987, 70, 984–991. [Google Scholar] [CrossRef]
- Guerriero, A.; D’Ambrosio, M.; Pietra, F. Slowly interconverting conformers of the briarane diterpenoids verecynarmin B, C, and D, isolated from the nudibranch mollusc Armina maculata and the octocoral Veretillum cynomorium of the East Pyrenean waters. Helv. Chim. Acta 1988, 71, 472–485. [Google Scholar] [CrossRef]
- Guerriero, A.; D’Ambrosio, M.; Pietra, F. Isolation of the cembranoid preverecynarmin alongside some briaranes, the verecynarmins, from both the nudibranch mollusc Armina maculata and the octocoral Veretillum cynomorium of the East Pyrenean Mediterranean Sea. Helv. Chim. Acta 1990, 73, 277–283. [Google Scholar] [CrossRef]
- Fu, X.; Palomar, A.J.; Hong, E.P.; Schmitz, F.J.; Valeriote, F.A. Cytotoxic lissoclimide-type diterpenes from the molluscs Pleurobranchus albiguttatus and Pleurobranchus forskalii. J. Nat. Prod. 2004, 67, 1415–1418. [Google Scholar] [CrossRef] [PubMed]
- Bian, W.-T.; You, Z.-J.; Wang, C.-Y.; Shao, C.-L. Brominated pimare diterpenoids from the sea hare Aplysia pulmonica from the South China Sea. Chem. Nat. Comp. 2014, 50, 557–559. [Google Scholar] [CrossRef]
- McPhail, K.L.; Davies-Coleman, M.T. New spongiane diterpenes from the East African nudibranch Chromodoris hamiltoni. Tetrahedron 1997, 53, 4655–4660. [Google Scholar] [CrossRef]
- Miyamoto, T.; Sakamoto, K.; Arao, K.; Komori, T.; Higuchi, R.; Sasaki, T. Dorisenones, cytotoxic spongian diterpenoids, from the nudibranch Chromodoris obsoleta. Tetrahedron 1996, 52, 8187–8198. [Google Scholar] [CrossRef]
- Kohmoto, S.; McConnell, O.J.; Wrigth, A.; Cross, S. Isospongiadiol, a cytotoxic and antiviral diterpene from a Caribbean deep water marine sponge, Spongia sp. Chem. Lett. 1987, 16, 1687–1690. [Google Scholar] [CrossRef]
- Fontana, A.; Cavaliere, P.; Ungur, N.; D’Souza, L.; Parameswaram, P.S.; Cimino, G. New scalaranes from the nudibranch Glossodoris atromarginata and its sponge prey. J. Nat. Prod. 1999, 62, 1367–1370. [Google Scholar] [CrossRef]
- Fontana, A.; Mollo, E.; Ricciardi, D.; Fakhr, I.; Cimino, G. Chemical studies of Egyptian opisthobranchs: Spongian diterpenoids from Glossodoris atromarginata. J. Nat. Prod. 1997, 60, 444–448. [Google Scholar] [CrossRef]
- De Silva, E.D.; Scheuer, P.J. Furanoditerpenoids from the dorid nudibranch Casella atromarginata. Heterocycles 1982, 17, 167–170. [Google Scholar]
- Cambie, R.C.; Craw, P.A.; Stone, M.J.; Bergquist, P.R. Chemistry of sponges, IV. Spongian diterpenes from Hyatella intestinalis. J. Nat. Prod. 1988, 51, 293–297. [Google Scholar] [CrossRef]
- Agena, M.; Tanaka, C.; Hanif, N.; Yasumoto-Hirose, M.; Tanaka, J. New cytotoxic spongian diterpenes from the sponge Dysidea cf. arenaria. Tetrahedron 2009, 65, 1495–1499. [Google Scholar] [CrossRef]
- Midland, S.L.; Wing, R.M.; Sims, J.J. New crenulides from the sea hare Aplysia vaccaria. J. Org. Chem. 1983, 48, 1906–1909. [Google Scholar] [CrossRef]
- Sun, H.H.; McEnroe, F.J.; Fenical, W. Acetoxycrenulide, a new bicyclic cyclopropane-containing diterpenoid from the brown seaweed Dictyota crenulata. J. Org. Chem. 1983, 48, 1903–1906. [Google Scholar] [CrossRef]
- Cutignano, A.; Moles, J.; Avila, C.; Fontana, A. Granuloside, a unique linear homosesterterpene from the Antarctic nudibranch Charcotia granulosa. J. Nat. Prod. 2015, 78, 1761–1764. [Google Scholar] [CrossRef]
- Moles, J.; Wägele, H.; Cutignano, A.; Fontana, A.; Avila, C. Distribution of granuloside in the Antarctic nudibranch Charcotia granulosa (Gastropoda: Heterobranchia: Charcotiidae). Mar. Biol. 2016, 163, 54–65. [Google Scholar] [CrossRef]
- Daoust, J.; Fontana, A.; Merchant, C.E.; De Voogd, N.J.; Patrick, B.O.; Kieffer, T.J.; Andersen, R.J. Ansellone A, a sesterterpenoid isolated from the nudibranch Cadlina luteromarginata and the sponge Phorbas sp., activates the cAMP signaling pathway. Org. Lett. 2010, 12, 3208–3211. [Google Scholar] [CrossRef]
- El Sayed, K.A.; Bartyzel, P.; Shen, X.; Perry, T.L.; Zjawiony, J.K.; Hamann, M.T. Marine natural products as anti-tuberculosis agents. Tetrahedron 2000, 56, 949–953. [Google Scholar] [CrossRef]
- Jacobs, R.S.; Bober, M.A.; Pinto, I.; Williams, A.B.; Jacobson, P.B.; de Carvalho, M.S. Pharmacological studies of marine novel marine metabolites. In Advances in Marine Biotechnology; Attaway, D.H., Zaborsky, O.R., Eds.; Plenum: New York, NY, USA, 1993; Volume 1, pp. 77–99. [Google Scholar]
- Gavagnin, M.; Mollo, E.; Docimo, T.; Guo, Y.-W.; Cimino, G. Scalarane metabolites of the nudibranch Glossodoris rufomarginata and its dietary sponge from the South China Sea. J. Nat. Prod. 2004, 67, 2104–2107. [Google Scholar] [CrossRef] [PubMed]
- Manzo, E.; Gavagnin, M.; Somerville, M.J.; Mao, S.-C.; Ciavatta, M.L.; Mollo, E.; Schupp, P.J.; Garson, M.J.; Guo, V.; Cimino, G. Chemistry of Glossodoris nudibranchs: Specific occurrence of 12-keto scalaranes. J. Chem. Ecol. 2007, 33, 2325–2336. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.D.; Paul, V.J. Chemical defenses of three Glossodoris nudibranchs and their dietary Hyrtios sponges. Mar. Ecol. Progr. Ser. 1991, 77, 221–232. [Google Scholar] [CrossRef]
- Avila, C.; Paul, V.J. Chemical ecology of the nudibranch Glossodoris pallida: Is the location of diet-derived metabolites important for defense? Mar. Ecol. Progr. Ser. 1997, 150, 171–180. [Google Scholar] [CrossRef]
- Fontana, A.; Mollo, E.; Ortea, J.; Gavagnin, M.; Cimino, G. Scalarane and homoscalarane compounds from the nudibranchs Glossodoris sedna and Glossodoris dalli: Chemical and biological properties. J. Nat. Prod. 2000, 63, 527–530. [Google Scholar] [CrossRef]
- Miyamoto, T.; Sakamoto, K.; Amano, H.; Arakawa, Y.; Nagarekawa, Y.; Komori, T.; Higuchi, R.; Sasaki, T. New cytotoxic sesterterpenoids from the nudibranch Chromodoris inornata. Tetrahedron 1999, 55, 9133–9142. [Google Scholar] [CrossRef]
- Graziani, E.I.; Allen, T.M.; Andersen, R.J. Lovenone, a cytotoxic degraded triterpenoid isolated from skin extracts of the North Sea dorid nudibranch Adalaria loveni. Tetrahedron Lett. 1995, 36, 1763–1766. [Google Scholar] [CrossRef]
- Spinella, A.; Mollo, E.; Trivellone, E.; Cimino, G. Testudinariol A and B, two unusual triterpenoids from the skin and the mucus of the marine mollusc Pleurobranchus testudinarius. Tetrahedron 1997, 53, 16891–16896. [Google Scholar] [CrossRef]
- Pawlik, J.R.; Albizati, K.F.; Faulkner, D.J. Evidence of a defensive role for limatulone, a novel triterpene from the limpet Collisella limatula. Mar. Ecol. Progr. Ser. 1986, 30, 251–260. [Google Scholar] [CrossRef]
- Carbone, M.I. Molluschi Opistobranchi: Uno Straordinario Modello Per Selezionare Molecole Biologicamente Attive. Ph.D. Thesis, Università degli Studi di Salerno, Salerno, Italy, 2007; p. 217. [Google Scholar]
- Manzo, E.; Gavagnin, M.; Bifulco, G.; Cimino, P.; Di Micco, S.; Ciavatta, M.L.; Guo, Y.-W.; Cimino, G. Aplysiols A and B, squalene-derived polyethers from the mantle of the sea hare Aplysia dactylomela. Tetrahedron 2007, 63, 9970–9978. [Google Scholar] [CrossRef]
- Pennings, S.C.; Paul, V.J.; Dunbar, D.C.; Hamann, M.T.; Lumbang, W.; Novack, B.; Jacobs, R.S. Unpalatable compounds in the marine gastropod Dolabella auricularia: Distribution and effect of diet. J. Chem. Ecol. 1999, 25, 735–755. [Google Scholar] [CrossRef]
- Suenaga, K.; Shibata, T.; Takada, N.; Kigoshi, H.; Yamada, K. Aurilol, a cytotoxic bromotriterpene isolated from the sea hare Dolabella auricularia. J. Nat. Prod. 1998, 61, 515–518. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid Metabolism in Plants. Molec. Plant Metab. Synt. Biol. 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vershinin, A. Biological functions of carotenoids—Diversity and evolution. BioFactors 1999, 10, 99–104. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Lobo-da-Cunha, A.; Taveira, M.; Ferreira, M.; Valentão, P.; Andrade, P.B. Digestive gland from Aplysia depilans Gmelin: Leads for inflammation treatment. Molecules 2015, 20, 15766–15780. [Google Scholar] [CrossRef] [Green Version]
- Gavagnin, M.; Ungur, N.; Mollo, E.; Templado, J.; Cimino, G. Structure and synthesis of a progesterone homologue from the skin of the dorid nudibranch Aldisa smaragdina. Eur. J. Org. Chem. 2002, 9, 1500–1504. [Google Scholar] [CrossRef]
- Granato, A.C.; Berlinck, R.G.S.; Magalhaes, A.; Schefer, A.B.; Ferreira, A.G.; De Sanctis, B.; De Freitas, J.C.; Hajdu, E.; Migotto, A.E. Natural products from the marine sponges Aaptos sp. and Hymeniacidon aff. heliophila, and from the nudibranch Doris aff. verrucosa. Quim. Nov. 2000, 23, 594–599. [Google Scholar]
- Graziani, E.I.; Andersen, R.J.; Krug, P.J.; Faulkner, D.J. Stable isotope incorporation evidence for the de novo biosynthesis of terpenoic acid glycerides by dorid nudibranchs. Tetrahedron 1996, 52, 6869–6878. [Google Scholar] [CrossRef]
- Kubanek, J.; Andersen, R.J. Evidence for de novo biosynthesis of the polyketide fragment of diaulusterol A by the Northeastern Pacific dorid nudibranch Diaulula sandiegensis. J. Nat. Prod. 1999, 62, 777–779. [Google Scholar] [CrossRef]
- Cimino, G.; De Rosa, S.; De Stefano, S.; Sodano, G. Cholest-4-en-4,16β,18,22R-tetrol-3-one 16,18-diacetate a novel polyhydroxilated steroid from the hydroid Eudendrium sp. Tetrahedron Lett. 1980, 21, 3303–3304. [Google Scholar] [CrossRef]
- Spinella, A.; Gavagnin, M.; Crispino, A.; Cimino, G. 4-acetylaplykurodin B and aplykurodinone B, two ichthyotoxic degraded sterols from the Mediterranean mollusk Aplysia fasciata. J. Nat. Prod. 1992, 5, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.J.; Zubía, E.; Salvá, J. 3-epi-aplykurodinone B, a new degraded sterol from Aplysia fasciata. J. Nat. Prod. 1997, 60, 488–489. [Google Scholar] [CrossRef] [PubMed]
- Kita, M.; Kawamura, A.; Kigoshi, H. Aplysiasecosterols B and C: Two new 9,11-secosteroids with a cis-fused 1,4-quinone structure from the sea hare Aplysia kurodai. Tetrahedron Lett. 2016, 57, 858–860. [Google Scholar] [CrossRef] [Green Version]
- Gavagnin, M.; Carbone, M.; Nappo, M.; Mollo, E.; Roussis, V.; Cimino, G. First chemical study of anaspidean Syphonota geographica: Structure of degraded sterols aplykurodinone-1 and -2. Tetrahedron 2005, 61, 617–621. [Google Scholar] [CrossRef]
- Carbone, M.; Gavagnin, M.; Mollo, E.; Bidello, M.; Roussis, V.; Cimino, G. Further syphonosides from the sea hare Syphonota geographica and the sea-grass Halophila stipulacea. Tetrahedron 2008, 64, 191–196. [Google Scholar] [CrossRef]
- Fisch, K.M.; Hertzer, C.; Böhringer, N.; Wuisan, Z.G.; Schillo, D.; Bara, R.; Kaligis, F.; Wägele, H.; König, G.M.; Schäberle, T.F. The potential of Indonesian heterobranchs found around Bunaken island for the production of bioactive compounds. Mar. Drugs. 2017, 15, 384. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group (Species)/Compounds | Anaspidea (5) | Sacoglossa (11) |
|---|---|---|
| Linear Monoterpenoids | >10 1 | 5 |
| Monocyclic Monoterpenoids | >10 1 | 2 |
| Group (Species)/Compounds | Nudibranchia (51) | Anaspidea (7) | Sacoglossa (2) | Pulmonata (1) |
|---|---|---|---|---|
| Linear Sesquiterpenoids | 1 | 0 | 0 | 0 |
| Monocyclic Sesquiterpenoids | 5 | >5 1 | 3 | 2 |
| Bicyclic Sesquiterpenoids | >60 1 | >6 1 | 0 | 0 |
| Tricyclic Sesquiterpenoids | >5 1 | 3 | 0 | 0 |
| Group (Species)/Compounds | Nudibranchia (39) | Pleurobranchoidea (3) | Anaspidea (8) | Sacoglossa (5) | Pulmonata (3) |
|---|---|---|---|---|---|
| Linear Diterpenoids | 1 | 0 | 0 | 11 | 0 |
| Monocyclic Diterpenoids | >8 1 | 0 | 10 | 2 | 0 |
| Bicyclic Diterpenoids | 8 | 2 | >15 1 | 0 | >2 1 |
| Tricyclic Diterpenoids | >85 1 | 7 | 5 | 0 | 0 |
| Tetracyclic Diterpenoids | 25 | 0 | 4 | 0 | 0 |
| Group (Species)/Compounds | Nudibranchia (13) |
|---|---|
| Linear Sesterterpenoids | 1 |
| Bicyclic Sesterterpenoids | 1 |
| Tetracyclic Sesterterpenoids | >16 1 |
| Group (Species)/Compounds | Nudibranchia (1) | Pleurobranchoidea (2) | Anaspidea (2) |
|---|---|---|---|
| Cyclic Triterpenoids | 1 | 2 | 3 |
| Group (Species)/Compounds | Nudibranchia (8) | Anaspidea (5) |
|---|---|---|
| Carotenoids | 0 | 8 |
| Steroids | >10 1 | >8 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avila, C. Terpenoids in Marine Heterobranch Molluscs. Mar. Drugs 2020, 18, 162. https://doi.org/10.3390/md18030162
Avila C. Terpenoids in Marine Heterobranch Molluscs. Marine Drugs. 2020; 18(3):162. https://doi.org/10.3390/md18030162
Chicago/Turabian StyleAvila, Conxita. 2020. "Terpenoids in Marine Heterobranch Molluscs" Marine Drugs 18, no. 3: 162. https://doi.org/10.3390/md18030162
APA StyleAvila, C. (2020). Terpenoids in Marine Heterobranch Molluscs. Marine Drugs, 18(3), 162. https://doi.org/10.3390/md18030162