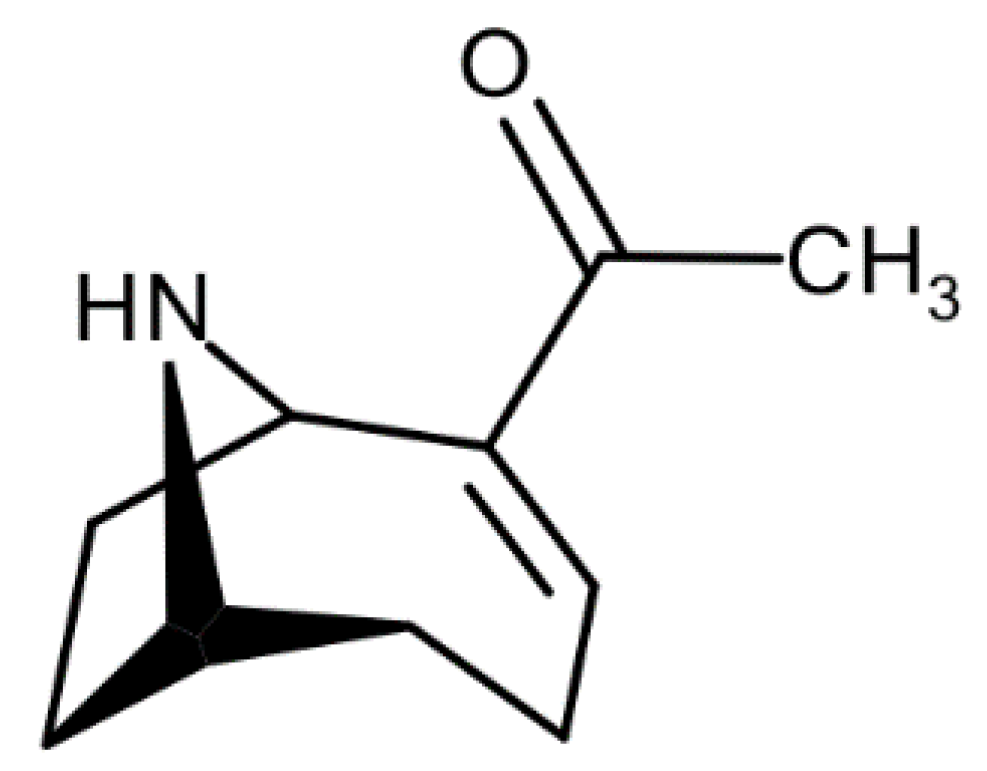
First Evidence of the Presence of Anatoxin-A in Sea Figs Associated with Human Food Poisonings in France

 , and
, and
Abstract
1. Introduction
2. Results
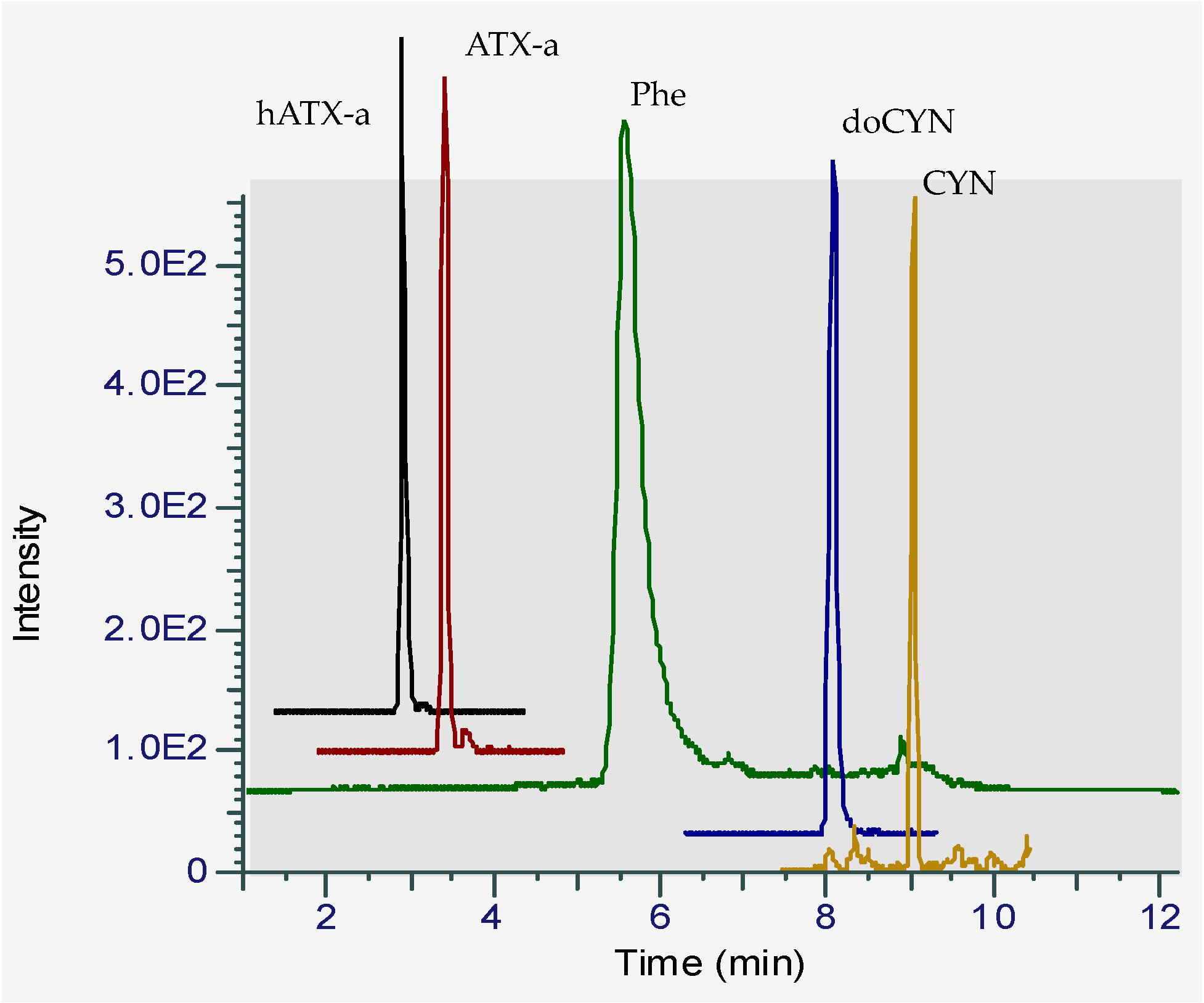
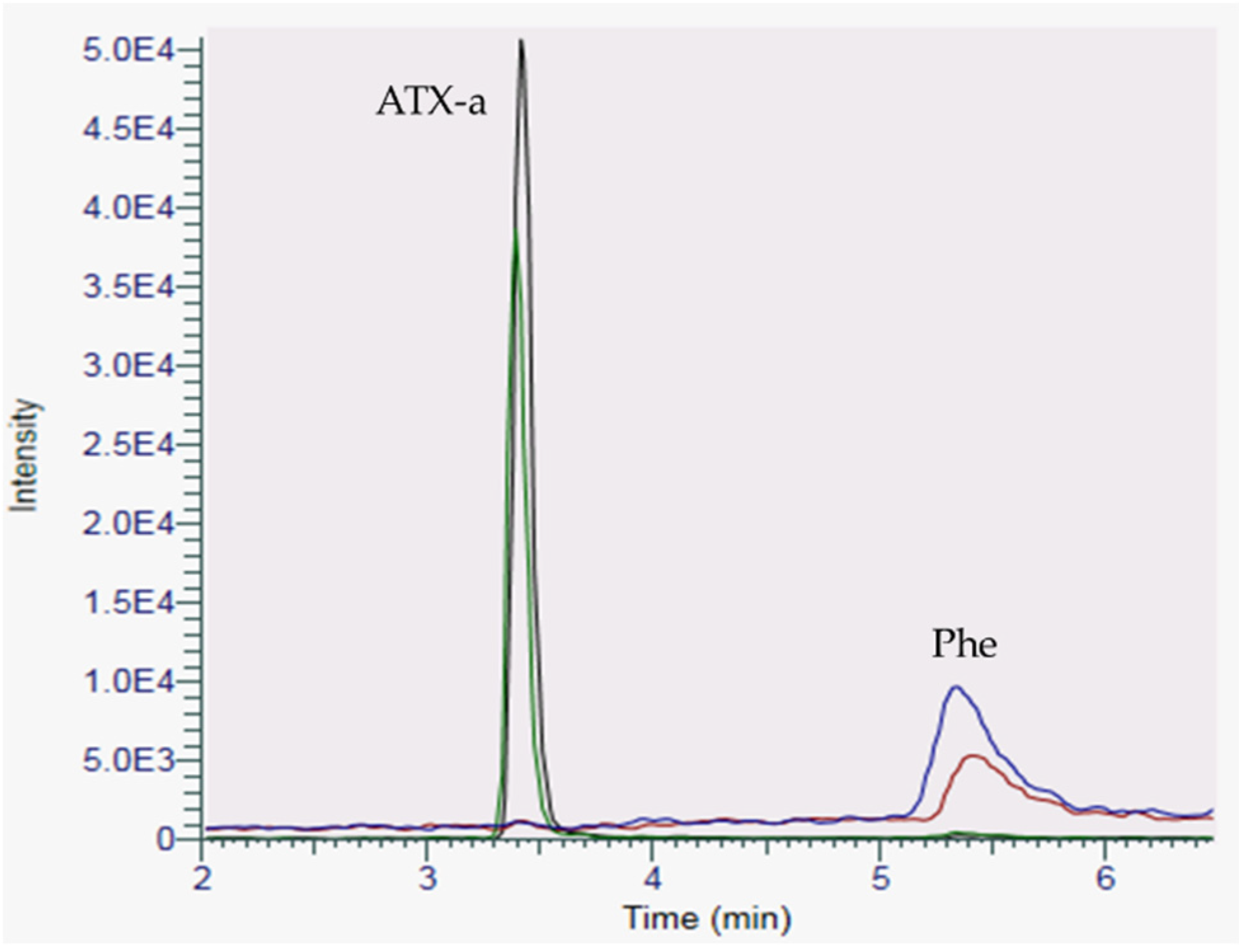
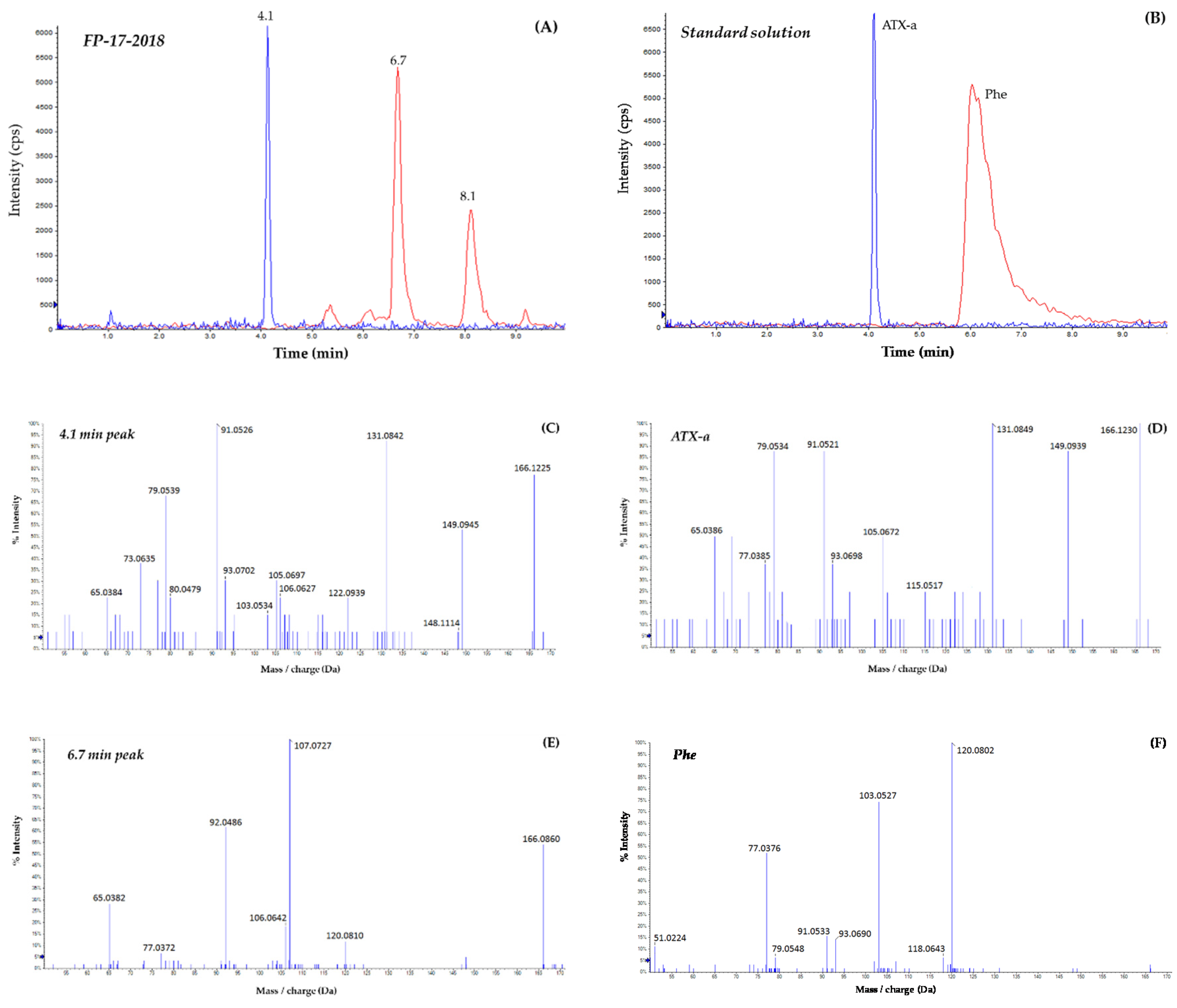
2.1. HILIC-MS Analysis in Low Resolution
2.2. Confirmatory Analysis Using HILIC-HRMS
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Sea Figs
4.3. Extraction
4.4. Cleanup
4.5. HILIC-MS Analysis
4.5.1. HILIC-LRMS Analysis
- (A)
- H2O + 0.015% FA + 0.015% NH3
- (B)
- 90% ACN + 0.01% FA
- (C)
- H2O + 0.5% FA
- (D)
- 100% MeOH
4.5.2. HILIC-HRMS Analysis
4.5.3. Determination of the Sea Figs Matrix Effects in HILIC-LRMS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Roje-Busatto, R.; Ujevic, I. PSP toxins profile in ascidian Microcosmus vulgaris (Heller, 1877) after human poisoning in Croatia (Adriatic Sea). Toxicon 2014, 79, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.C.; Ogata, T.; Veit, C.H.; Kodama, M. Occurrence of tetrodotoxin and paralytic shellfish toxins in Phallusia nigra (Tunicata, Ascidiacea) from the Brazilian coast. J. Venom. Anim. Toxins 1996, 2, 28–38. [Google Scholar] [CrossRef]
- Lopez-Rivera, A.; Pinto, M.; Insinilla, A.; Suarez Isla, B.; Uribe, E.; Alvarez, G.; Lehane, M.; Furey, A.; James, K.J. The occurrence of domoic acid linked to a toxic diatom bloom in a new potential vector: The tunicate Pyura chilensis (piure). Toxicon 2009, 54, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Gibble, C.M.; Peacock, M.B.; Kudela, R.M. Evidence of freshwater algal toxins in marine shellfish: Implications for human and aquatic health. Harmful Algae 2016, 59, 59–66. [Google Scholar] [CrossRef]
- Peacock, M.B.; Gibble, C.M.; Senn, D.B.; Cloern, J.E.; Kudela, R.M. Blurred lines: Multiple freshwater and marine algal toxins at the land-sea interface of San Francisco Bay, California. Harmful Algae 2018, 73, 138–147. [Google Scholar] [CrossRef]
- Laurent, D.; Kerbrat, A.S.; Darius, H.T.; Girard, E.; Golubic, S.; Benoit, E.; Sauviat, M.P.; Chinain, M.; Molgo, J.; Pauillac, S. Are cyanobacteria involved in Ciguatera Fish Poisoning-like outbreaks in New Caledonia? Harmful Algae 2008, 7, 827–838. [Google Scholar] [CrossRef]
- Mejean, A.; Peyraud-Thomas, C.; Kerbrat, A.S.; Golubic, S.; Pauillac, S.; Chinain, M.; Laurent, D. First identification of the neurotoxin homoanatoxin-a from mats of Hydrocoleum lyngbyaceum (marine cyanobacterium) possibly linked to giant clam poisoning in New Caledonia. Toxicon 2010, 56, 829–835. [Google Scholar] [CrossRef]
- Testai, E.; Scardala, S.; Vichi, S.; Buratti, F.M.; Funari, E. Risk to human health associated with the environmental occurrence of cyanobacterial neurotoxic alkaloids anatoxins and saxitoxins. Crit. Rev. Toxicol. 2016, 46, 385–419. [Google Scholar] [CrossRef]
- Harada, K.-I.; Kimura, Y.; Ogawa, K.; Suzuki, M.; Dahlem, A.M.; Beasley, V.R.; Carmichael, W.W. A new procedure for the analysis and purification of naturally occurring anatoxin-a from the blue-green alga Anabaena flos-aquae. Toxicon 1989, 27, 1289–1296. [Google Scholar] [CrossRef]
- Rellan, S.; Osswald, J.; Vasconcelos, V.; Gago-Martinez, A. Analysis of anatoxin-a in biological samples using liquid chromatography with fluorescence detection after solid phase extraction and solid phase microextraction. J. Chromatogr. A 2007, 1156, 134–140. [Google Scholar] [CrossRef]
- James, K.J.; Sherlock, I.R.; Stack, M.A. Anatoxin-a in Irish freshwater and cyanobacteria, determined using a new fluorimetric liquid chromatographic method. Toxicon 1997, 35, 963–971. [Google Scholar] [CrossRef]
- James, K.J.; Furey, A.; Sherlock, I.R.; Stack, M.A.; Twohig, M.; Caudwell, F.B.; Skulberg, O.M. Sensitive determination of anatoxin-a, homoanatoxin-a and their degradation products by liquid chromatography with fluorimetric detection. J. Chromatogr. A 1998, 798, 147–157. [Google Scholar] [CrossRef]
- Rellán, S.; Osswald, J.; Saker, M.; Gago-Martinez, A.; Vasconcelos, V. First detection of anatoxin-a in human and animal dietary supplements containing cyanobacteria. Food Chem. Toxicol. 2009, 47, 2189–2195. [Google Scholar] [CrossRef] [PubMed]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and Analysis of Occurrence, Exposure and Toxicity of Cyanobacteria Toxins in Food; EFSA: Parma, Italy, 2016; p. 309. [Google Scholar] [CrossRef]
- Furey, A.; Crowley, J.; Hamilton, B.; Lehane, M.; James, K.J. Strategies to avoid the mis-identification of anatoxin-a using mass spectrometry in the forensic investigation of acute neurotoxic poisoning. J. Chromatogr. A 2005, 1082, 91–97. [Google Scholar] [CrossRef]
- Dell’Aversano, C.; Eaglesham, G.K.; Quilliam, M.A. Analysis of cyanobacterial toxins by hydrophilic interaction liquid chromatography-mass spectrometry. J. Chromatogr. A 2004, 1028, 155–164. [Google Scholar] [CrossRef]
- Schmitt, C.; Torrents, R.; Domange, B.; Simon, N.; de Haro, L. Cerebellar syndrome associated with ingestion of Mediterranean Microcosmus: A French case series. Clin. Toxicol. (Phila) 2019, 57, 221–223. [Google Scholar] [CrossRef]
- Dom, I.; Bire, R.; Hort, V.; Lavison-Bompard, G.; Nicolas, M.; Guerin, T. Extended Targeted and Non-Targeted Strategies for the Analysis of Marine Toxins in Mussels and Oysters by (LC-HRMS). Toxins (Basel) 2018, 10, 375. [Google Scholar] [CrossRef]
- Dom, I. Analyse non Ciblée des Biotoxines Marines dans les Produits de la Pêche. Ph.D. Thesis, Institut des Sciences et Industries du Vivant et de l’Environnement (AgroParisTech), Paris, France, 2018. [Google Scholar]
- Turner, A.D.; McNabb, P.S.; Harwood, D.T.; Selwood, A.I.; Boundy, M.J. Single-Laboratory Validation of a Multitoxin Ultra-Performance LC-Hydrophilic Interaction LC-MS/MS Method for Quantitation of Paralytic Shellfish Toxins in Bivalve Shellfish. J. AOAC Int. 2015, 98, 609–621. [Google Scholar] [CrossRef]
- Preece, E.P.; Moore, B.C.; Hardy, F.J.; Deobald, L.A. First detection of microcystin in Puget Sound, Washington, mussels (Mytilus trossulus). Lake Reservoir. Manag. 2015, 31, 50–54. [Google Scholar] [CrossRef]
- Kust, A.; Méjean, A.; Ploux, O. Biosynthesis of Anatoxins in Cyanobacteria: Identification of the Carboxy-anatoxins as the Penultimate Biosynthetic Intermediates. J. Nat. Prod. 2020, 83, 142–151. [Google Scholar] [CrossRef]
- Backer, L.C.; Landsberg, J.H.; Miller, M.; Keel, K.; Taylor, T.K. Canine cyanotoxin poisonings in the United States (1920s-2012): Review of suspected and confirmed cases from three data sources. Toxins 2013, 5, 1597–1628. [Google Scholar] [CrossRef] [PubMed]
- Chatziefthimiou, A.D.; Richer, R.; Rowles, H.; Powell, J.T.; Metcalf, J.S. Cyanotoxins as a potential cause of dog poisonings in desert environments. Vet. Rec. 2014, 174, 484–485. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Harkema, L.; Begeman, L.; Lurling, M. First report of (homo)anatoxin-a and dog neurotoxicosis after ingestion of benthic cyanobacteria in The Netherlands. Toxicon 2012, 60, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Beulker, C.; Geiser, B.; Hoffmann, A.; Kröger, R.; Teske, K.; Hoppe, J.; Mundhenk, L.; Neurath, H.; Sagebiel, D.; et al. Fatal neurotoxicosis in dogs associated with tychoplanktic, anatoxin-a producing tychonema sp. In mesotrophic lake tegel, Berlin. Toxins 2018, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Gugger, M.; Lenoir, S.; Berger, C.; Ledreux, A.; Druart, J.C.; Humbert, J.F.; Guette, C.; Bernard, C. First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin-a associated with dog neurotoxicosis. Toxicon 2005, 45, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Puschner, B.; Hoff, B.; Tor, E.R. Diagnosis of anatoxin-a poisoning in dogs from North America. J. Vet. Diagn. Investig. 2008, 20, 89–92. [Google Scholar] [CrossRef]
- Puschner, B.; Pratt, C.; Tor, E.R. Treatment and diagnosis of a dog with fulminant neurological deterioration due to anatoxin-a intoxication. J. Vet. Emerg. Crit. Care 2010, 20, 518–522. [Google Scholar] [CrossRef]
- Wood, S.A.; Puddick, J.; Fleming, R.; Heussner, A.H. Detection of anatoxin-producing Phormidium in a New Zealand farm pond and an associated dog death. N. Z. J. Bot. 2017, 55, 36–46. [Google Scholar] [CrossRef]
- Wood, S.A.; Selwood, A.I.; Rueckert, A.; Holland, P.T.; Milne, J.R.; Smith, K.F.; Smits, B.; Watts, L.F.; Cary, C.S. First report of homoanatoxin-a and associated dog neurotoxicosis in New Zealand. Toxicon 2007, 50, 292–301. [Google Scholar] [CrossRef]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Pflugmacher, S. Contribution of hot spring cyanobacteria to the mysterious deaths of Lesser Flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Morrison, L.F.; Krienitz, L.; Ballot, A.; Krause, E.; Kotut, K.; Pütz, S.; Wiegand, C.; Pflugmacher, S.; Codd, G.A. Analysis of the cyanotoxins anatoxin-a and microcystins in Lesser Flamingo feathers. Toxicol. Environ. Chem. 2006, 88, 159–167. [Google Scholar] [CrossRef]
- Behm, B. Coroner cites algae in teen’s death. Milwaukee J. Sentin. 2003, 5, 3. [Google Scholar]
- Carmichael, W.W.; Yuan, M.; Friday, C.F. Human Mortality from Accidental Ingestion of Toxic Cyanobacteria—A Case re-Examined (Abstract). In Proceedings of the Sixth International Conference on Toxic Cyanobacteria, Bergen, Norway, 21–27 June 2004; pp. 61–62. [Google Scholar]
- Mons, M.P.; Van Egmond, H.P.; Speijers, G.J.A. Paralytic Shellfish Poisoning; A Review; RIVM Report 388802005; RIVM: Bilthoven, The Netherlands, 1998; p. 60. Available online: https://www.rivm.nl/bibliotheek/rapporten/388802005.pdf (accessed on 12 December 2019).
- Oregon Health Authority Frequently Asked Questions: Fishing and Blue-Green Algae Blooms. Available online: https://www.oregon.gov/oha/PH/HEALTHYENVIRONMENTS/RECREATION/HARMFULALGAEBLOOMS/Documents/fishermanAlgae_factsFINAL.pdf (accessed on 12 December 2019).
- Trainer, V.L.; Hardy, F.J. Integrative monitoring of marine and freshwater harmful algae in Washington State for public health protection. Toxins (Basel) 2015, 7, 1206–1234. [Google Scholar] [CrossRef] [PubMed]
- Alberta Health Services. Blue-Green Algae Health Advisories. Available online: https://www.albertahealthservices.ca/news/bga.aspx (accessed on 12 December 2019).
- Butler, N.; Linville, R. Toxicological Summary and Suggested Action Levels to Reduce Potential Adverse Health Effects of Six Cyanotoxins; Office of Environmental Health Hazard Assessment: Sacramento, CA, USA, 2012; p. 119. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ATX-a Concentration (µg/kg) |
|---|---|
| FP-1-2011 | 193.7 |
| FP-6-2012 | 1240.2 |
| FP-17-2018 | 1132.6 |
| Sea fig control | 22.5 |
| Mussel control | <LOD * |
| Toxin | Formula | Mass (Da) | Extraction Mass [M + H]+ (Da) |
|---|---|---|---|
| ATX-a | C10H15NO | 165.11536 | 166.12264 |
| hATX-a | C11H17NO | 179.13101 | 180.13829 |
| Carboxy ATX-a | C11H15NO3 | 209.10519 | 210.11247 |
| Carboxy hATX-a | C12H17NO3 | 223.12084 | 224.12812 |
| Carboxy dihydroATX-a | C11H17NO3 | 211.12084 | 212.12812 |
| N-methyl ATX a | C11H17NO | 179.13101 | 180.13829 |
| (10S)-ATX alcohol | C10H17NO | 167.13101 | 168.13829 |
| (10R)-ATX alcohol | C10H17NO | 167.13101 | 168.13829 |
| nor ATX-a | C9H13NO | 151.09971 | 152.10699 |
| Dihydro ATX-a | C10H17NO | 167.13101 | 168.13829 |
| Dihydro hATX-a | C11H19NO | 181.14666 | 182.15394 |
| Epoxy ATX-a | C10H15NO2 | 181.11028 | 182.11756 |
| Epoxy hATX-a | C11H17NO2 | 195.12593 | 196.13321 |
| ATX-(a)s | C7H17N4O4P | 252.09874 | 253.10602 |
| Phe | C9H11NO2 | 165.07898 | 166.08626 |
| Heading | Heading |
|---|---|
| Column | Acquity Glycan BEH Amide column 130 Å 1.7 µm, 2.1 × 150 mm |
| Pre-column | Acquity Glycan BEH Amide VanGuard Pre-column, 130 Å 1.7 µm, 2.1 × 5 mm |
| Column temperature | 70 °C |
| Injection volume (µL) | 2 µL |
| Toxin | Transition * | Precursor Ion (m/z) | Product Ions (m/z) | Ionization (+/-) | Collision Energy (V) | S-Lens (V) |
|---|---|---|---|---|---|---|
| ATX | Q | 166.1 | 131.1 | + | 14 | 53 |
| q | 166.1 | 105.1 | + | 16 | 53 | |
| hATX | Q | 180.1 | 145.1 | + | 14 | 56 |
| q | 180.1 | 117.1 | + | 20 | 56 | |
| CYN | Q | 416.2 | 336.1 | + | 20 | 79 |
| q | 416.2 | 194.1 | + | 34 | 79 | |
| doCYN | Q | 400.1 | 194.1 | + | 32 | 99 |
| q | 400.1 | 320.1 | + | 20 | 99 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biré, R.; Bertin, T.; Dom, I.; Hort, V.; Schmitt, C.; Diogène, J.; Lemée, R.; De Haro, L.; Nicolas, M. First Evidence of the Presence of Anatoxin-A in Sea Figs Associated with Human Food Poisonings in France. Mar. Drugs 2020, 18, 285. https://doi.org/10.3390/md18060285
Biré R, Bertin T, Dom I, Hort V, Schmitt C, Diogène J, Lemée R, De Haro L, Nicolas M. First Evidence of the Presence of Anatoxin-A in Sea Figs Associated with Human Food Poisonings in France. Marine Drugs. 2020; 18(6):285. https://doi.org/10.3390/md18060285
Chicago/Turabian StyleBiré, Ronel, Thomas Bertin, Inès Dom, Vincent Hort, Corinne Schmitt, Jorge Diogène, Rodolphe Lemée, Luc De Haro, and Marina Nicolas. 2020. "First Evidence of the Presence of Anatoxin-A in Sea Figs Associated with Human Food Poisonings in France" Marine Drugs 18, no. 6: 285. https://doi.org/10.3390/md18060285
APA StyleBiré, R., Bertin, T., Dom, I., Hort, V., Schmitt, C., Diogène, J., Lemée, R., De Haro, L., & Nicolas, M. (2020). First Evidence of the Presence of Anatoxin-A in Sea Figs Associated with Human Food Poisonings in France. Marine Drugs, 18(6), 285. https://doi.org/10.3390/md18060285