
Interactions of Nereistoxin and Its Analogs with Vertebrate Nicotinic Acetylcholine Receptors and Molluscan ACh Binding Proteins


Abstract
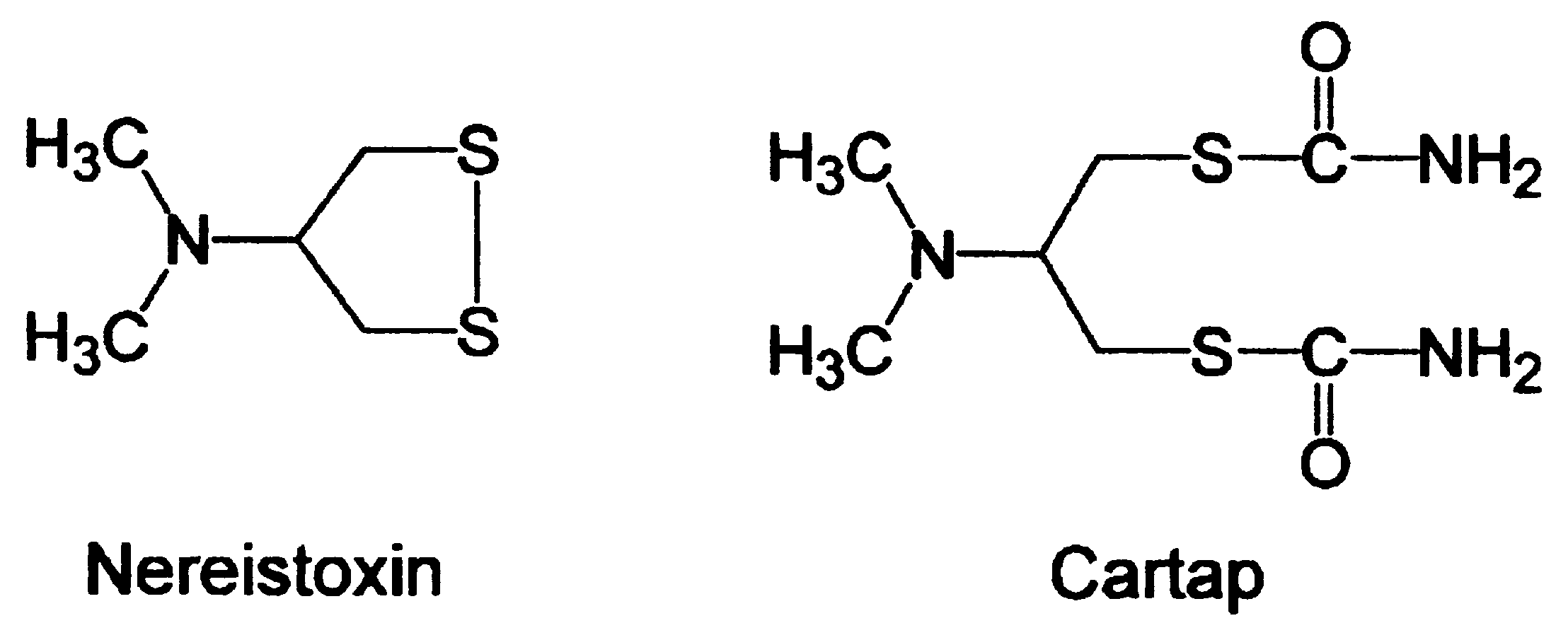
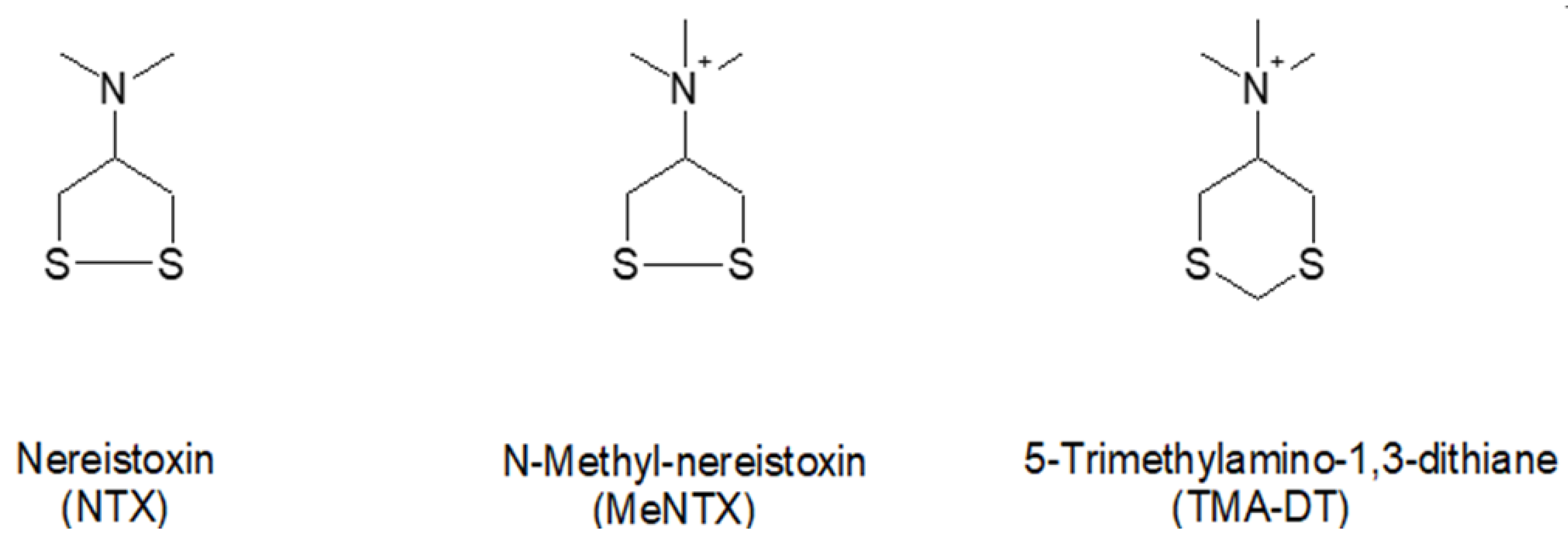
1. Introduction
2. Results
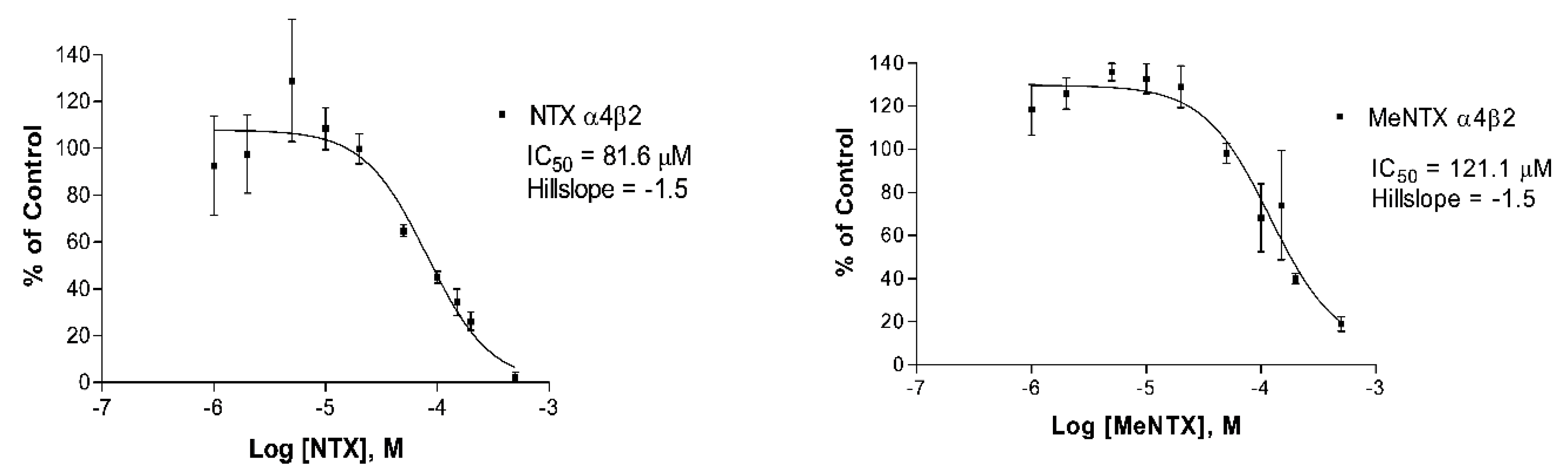
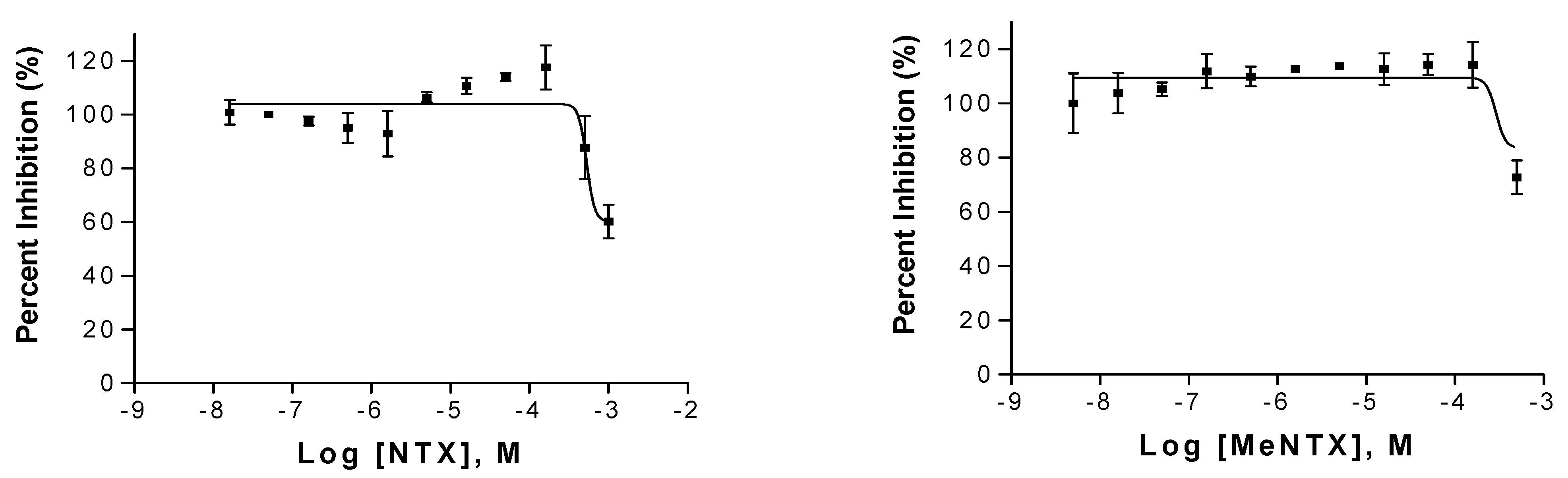
2.1. Effects of NTX on α4β2 Neuronal Receptors
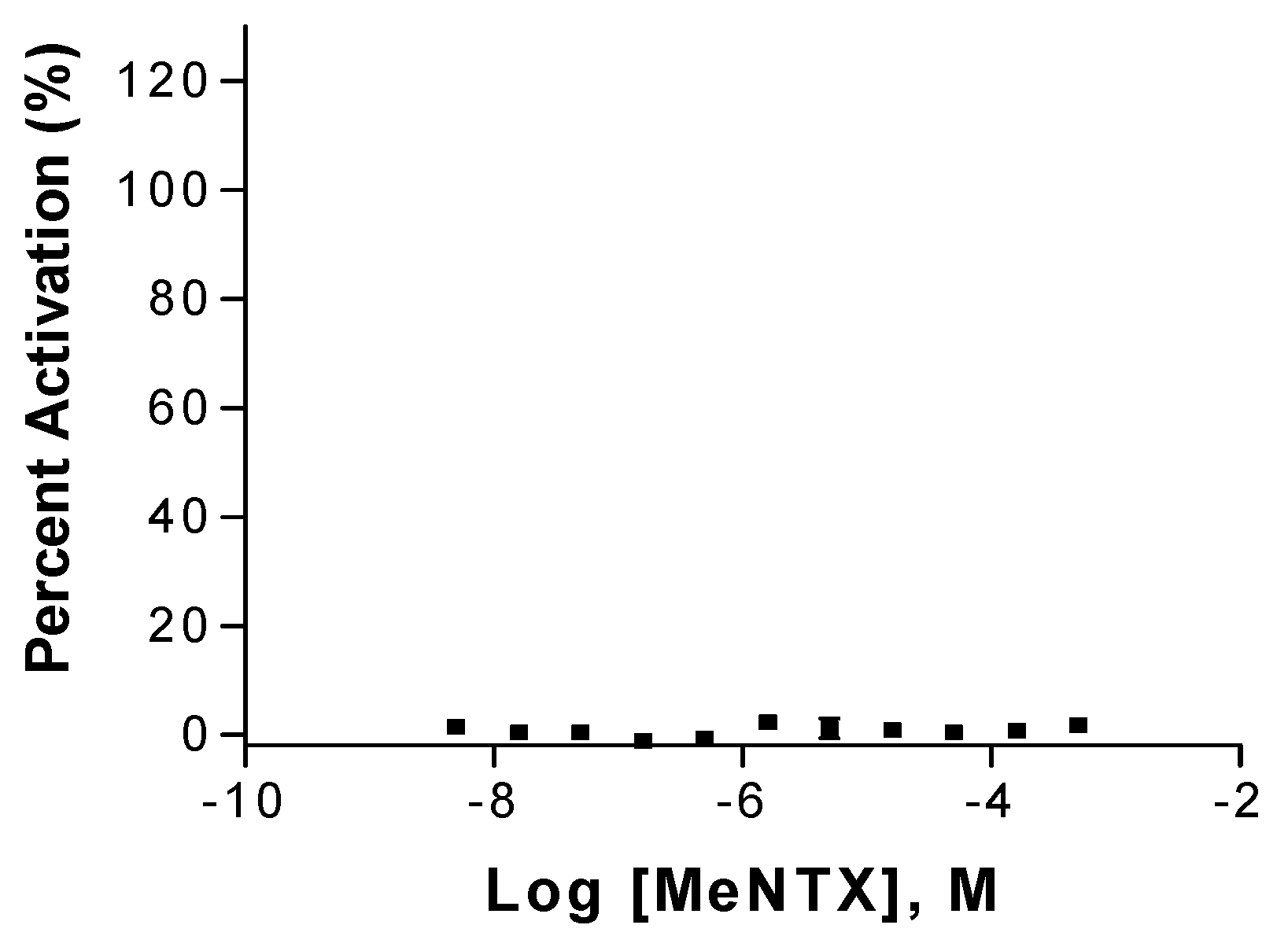
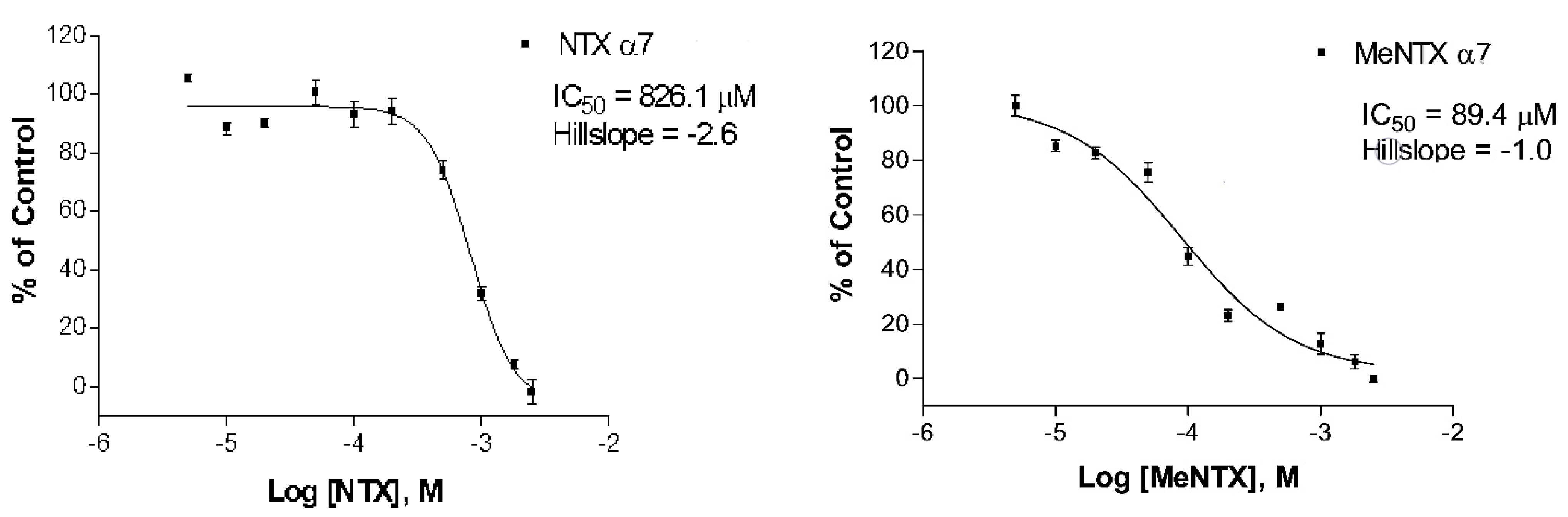
2.2. Effects on α7Neuronal Receptors
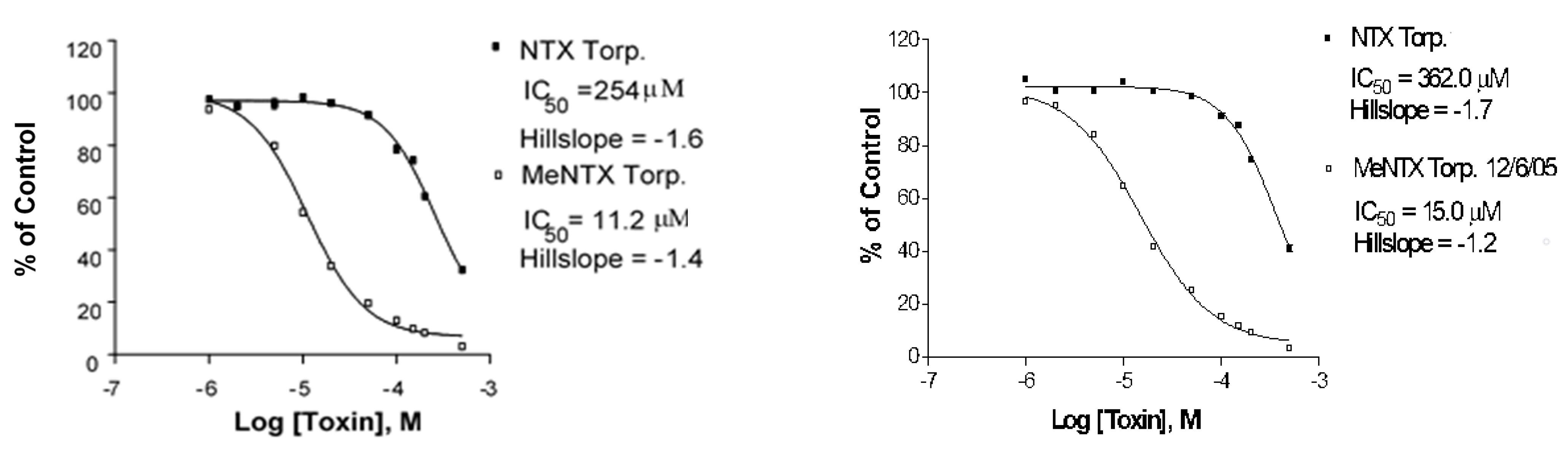
2.3. Torpedo Electric Organ nAChR Binding
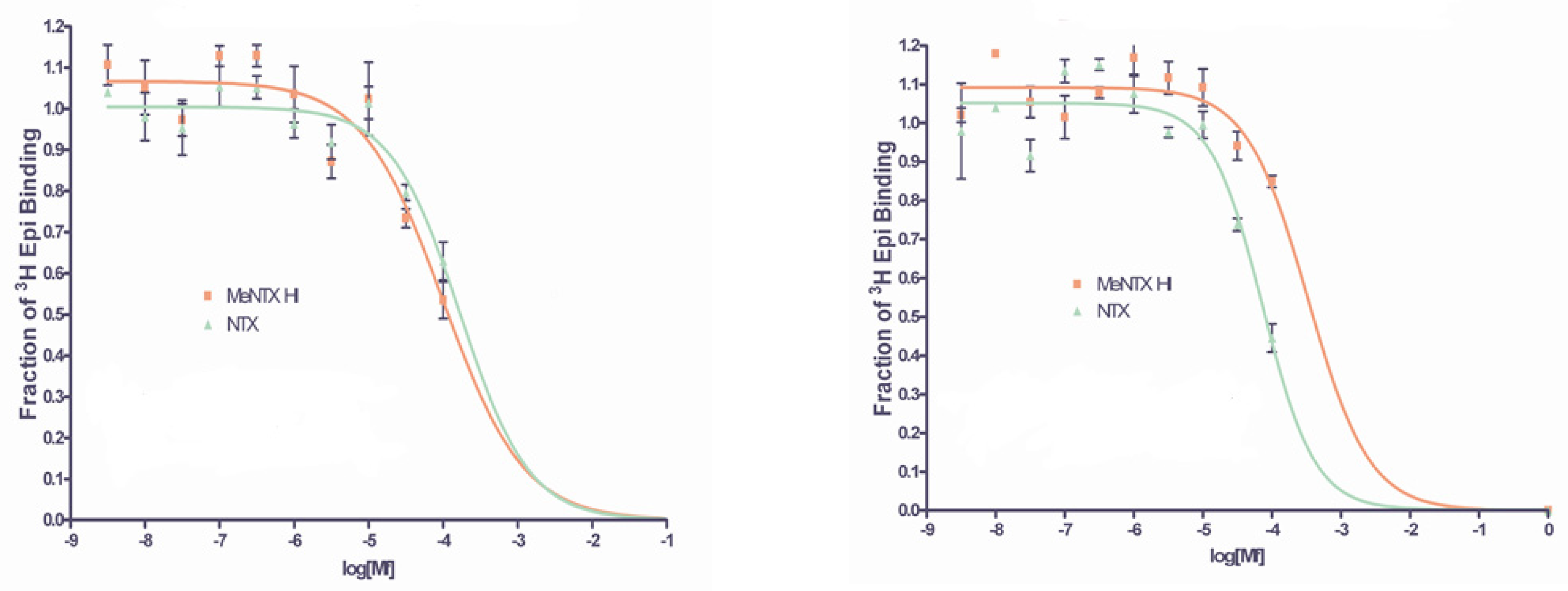
2.4. Binding of NTX and MeNTX to Molluscan ACh Binding Proteins
3. Discussion
4. Materials and Methods
4.1. NTX Compounds
4.2. Radioligand Binding Assays
4.3. Radioligand Binding Assay for AChBPs
4.4. Nicotinic Receptor FlexStation Functional Assays
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nitta, S. Uber Nereistoxin, einen giftigen Bestandteil von Lumbriconereis heteropoda Marenz (Eunicidae). Yakagaku Zasshi 1934, 54, 648. [Google Scholar] [CrossRef][Green Version]
- Okaichi, T.; Hashimoto, Y. The structure of nereistoxin. Agric. Biol. Chem. 1962, 26, 224–227. [Google Scholar] [CrossRef]
- Deguchi, T.; Narahashi, T.; Haas, H.G. Mode of action of nereistoxin on the neuromuscular transmission in the frog. Pest. Biochem. Physiol. 1971, 1, 196–204. [Google Scholar] [CrossRef]
- Eldefrawi, A.T.; Bakry, N.M.; Eldefrawi, M.E.; Tsai, M.-C.; Albuquerque, E.X. Nereistoxin interaction with the acetylcholine receptor-ionic channel complex. Mol. Pharmacol. 1980, 17, 172–179. [Google Scholar]
- Kem, W. Nereistoxin and Related Compounds. In Experimental and Clinical Neurotoxicology, 2nd ed.; Spencer, P.S., Schaumburg, H.H., Eds.; Oxford University Press: New York, NY, USA, 2000; pp. 858–859. [Google Scholar]
- Kao, P.N.; Karlin, A. Acetylcholine receptor binding site contains a disulfide crosslink between adjacent cysteinyl residues. J. Biol. Chem. 1986, 261, 8085–8088. [Google Scholar] [CrossRef]
- Nagawa, Y.; Saji, Y.; Chiba, S.; Yu, T. Neuromuscular blocking actions of nereistoxin and its derivatives and antagonism by sulfhydryl compounds. Jpn. J. Pharmacol. 1971, 21, 185–197. [Google Scholar] [CrossRef]
- Xie, Y.; Lane, W.V.; Loring, R.H. Nereistoxin: A naturally occurring toxin with redox effects on neuronal nicotinic acetylcholine receptors in chick retina. J. Pharm. Exp. Ther. 1993, 264, 689–694. [Google Scholar]
- Tomizawa, M.; Otsuka, H. Pharmacological effects of imidacloprid and its related compounds on the nicotinic acetylcholine receptor with its ion channel from the Torpedo electric organ. J. Pestic. Sci. 1995, 20, 49056. [Google Scholar]
- Sattelle, D.B.; Harrow, I.D.; David, J.A.; Pelhate, M.; Callec, J.J.; Gepner, J.I.; Hall, L.M. Nereistoxin: Actions on a CNS acetylcholine receptor/ion channel in the cockroach Periplaneta americana. J. Exp. Biol. 1985, 118, 37–52. [Google Scholar] [CrossRef]
- Lee, S.; Tomizawa, M.; Casida, J.E. Nereistoxin and cartap neurotoxicity attributable to direct block of the insect nicotinic receptor/channel. Agric. Food Chem. 2003, 51, 2646–2652. [Google Scholar] [CrossRef]
- Xing, H.; Andrud, K.W.; Soti, F.; Rouchaud, A.; Jahn, S.C.; Lu, Z.; Cho, Y.H.; Habibi, S.; Corsino, P.; Slavov, S.; et al. A Methyl Scan of the Pyrrolidinium Ring of Nicotine Reveals Significant Differences in Its Interactions with α7 and α4β2 Nicotinic Acetylcholine Receptors. Mol. Pharmacol. 2020, 98, 168–180. [Google Scholar] [CrossRef]
- Delpech, V.R.; Ihara, M.; Coddou, C.; Matsuda, K.; Sattelle, D.B. Action of nereistoxin on recombinant neuronal nicotinic acetylcholine receptors expressed in Xenopus laevis oocytes. Invert. Neurosci. 2003, 5, 29–35. [Google Scholar] [CrossRef]
- Konishi, K. Studies of Organic Insecticides. Agric. Biol. Chem. 1968, 32, 1199–1204. [Google Scholar]
- Barlow, R.B.; Hamilton, J.T. Effects of pH on the activity of nicotine and nicotine monomethiodide on the rat diaphragm preparation. Br. J. Pharmacol. Chemother. 1962, 18, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.T. The influence of pH on the activity of nicotine at the neuromuscular junction. Can. J. Biochem. Physiol. 1963, 41, 283–289. [Google Scholar] [CrossRef]
- Bartels, E.; Podleski, T.R. Action of nicotine on the electroplax and difference of potency between ionized and unionized forms. Biochim. Biophys. Acta 1964, 79, 511–520. [Google Scholar] [CrossRef]
- Mosckovitz, R.; Gershoni, J.M. Three possible disulfides in the acetylcholine receptor α-subunit. J. Biol. Chem. 1988, 263, 1017–1022. [Google Scholar] [CrossRef]
- Blum, A.P.; Gleitsman, K.R.; Lester, H.A.; Dougherty, D.A. Evidence for an extended hydrogen bond network in the binding site of the nicotinic receptor: Role of the vicinal disulfide of the α1 subunit. J. Biol. Chem. 2011, 286, 32251–32258. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.S.; Chiara, D.C.; Cohen, J.B. Mapping the structural requirements for nicotinic acetylcholine receptor activation by using tethered alkyltrimethylammonium agonists and antagonists. Biochemistry 2006, 45, 10641–10653. [Google Scholar] [CrossRef] [PubMed]
- Celie, P.H.; van Rossum-Fikkert, S.E.; van Dijk, W.J.; Brejc, K.; Smit, A.B.; Sixma, T.K. Nicotine and carbamylcholine binding to nicotinic acetylcholine receptors as studied in AChBP crystal structures. Neuron 2004, 41, 907–914. [Google Scholar] [CrossRef]
- Hibbs, R.E.; Sulzenbacher, G.; Shi, J.; Talley, T.T.; Kem, W.R.; Marchot, P.; Taylor, P.; Bourne, Y. Structural determinants for interaction of partial agonists with acetylcholine binding protein and neuronal α7 nicotinic acetylcholine receptor. EMBO J. 2009, 28, 3040–3051. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, E.X.; Tsai, M.-C.; Aronstam, R.S.; Witkop, B.; Eldefrawi, S.T.; Elefrawi, M.E. Phencyclidine interactions with the ionic channel of the acetylcholine receptor and electrogenic membrane. Proc. Natl. Acad. Sci. USA 1980, 77, 1224–1228. [Google Scholar] [CrossRef]
- Aronstam, R.S.; King, C., Jr.; Albuquerque, E.X.; Daly, J.W.; Feigl, D.M. Binding of [3H]phencyclidine to the nicotinic receptor-ion channel complex of Torpedo electroplax. Biochem. Pharmacol. 1985, 34, 3037–3304. [Google Scholar] [CrossRef]
- Howard, E.G.; Lindsey, R.V., Jr. The chemistry of some 5-m-dithianones and dithiacycloalkanones. J. Am. Chem. Soc. 1960, 82, 158–164. [Google Scholar] [CrossRef]
- Kem, W.R.; Mahnir, V.M.; Papke, R.L.; Lingle, C.J. Anabaseine is a potent agonist on muscle and neuronal alpha-bungarotoxin-sensitive nicotinic receptors. J. Pharm. Exp. Ther. 1997, 283, 979–992. [Google Scholar]
- Kem, W.; Soti, F.; Wildeboer, K.; LeFrancois, S.; MacDougall, K.; Wei, D.-Q.; Chou, K.-C.; Arias, H. The nemertine toxin anabaseine and its derivative DMXBA (GTS-21): Chemical and pharmacological properties. Mar. Drugs 2006, 4, 255–273. [Google Scholar] [CrossRef]
- Andrud, K.; Xing, H.; Gabrielsen, B.; Bloom, L.; Mahnir, V.; Lee, S.; Green, B.T.; Lindstrom, J.; Kem, W.R. Investigation of the possible pharmacologically active forms of the nicotinic acetylcholine receptor agonist anabaseine. Mar. Drugs 2019, 16, 614. [Google Scholar] [CrossRef] [PubMed]
- Kem, W.R.; Mahnir, V.M.; Prokai, L.; Papke, R.L.; Cao, X.; LeFrancois, S.; Wildeboer, K.; Prokai-Tatrai, K.; Porter-Papke, J.; Soti, F. Hydroxy metabolites of the Alzheimer’s drug candidate 3-[(2,4-dimethoxy)benzylidene]-anabaseine dihydrochloride (GTS-21): Their molecular properties, interactions with brain nicotinic receptors, and brain penetration. Mol. Pharmacol. 2004, 6, 56–67. [Google Scholar] [CrossRef]
- Hansen, S.B.; Radic, Z.; Talley, T.T.; Molles, B.E.; Deerinck, T.; Tsigelny, I.; Taylor, P. Tryptophan fluorescence reveals conformational changes in the acetylcholine binding protein. J. Biol. Chem. 2002, 277, 41299–41302. [Google Scholar] [CrossRef]
- Hibbs, R.E.; Johnson, D.A.; Shi, J.; Hansen, S.B.; Taylor, P. Structural dynamics of the alpha-neurotoxin-acetylcholine-binding protein complex: Hydrodynamic and fluorescence anisotropy decay analyses. Biochemistry 2005, 44, 16602–16611. [Google Scholar] [CrossRef]
- Talley, T.T.; Yalda, S.; Ho, K.-Y.; Tor, Y.; Soti, F.; Kem, W.R.; Taylor, P. Spectroscopic analysis of benzylidene anabaseine complexes with acetylcholine binding proteins as models for ligand-nicotinic receptor interactions. Biochemistry 2006, 45, 8894–8902. [Google Scholar] [PubMed]
- Fitch, R.W.; Xiao, Y.; Kellar, K.J.; Daly, J.W. Membrane potential fluorescence: A rapid and highly sensitive assay for nicotinic receptor channel function. Proc. Natl. Acad. Sci. USA 2003, 100, 4909–4914. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rat Brain | Rat Brain | Torpedo | |
|---|---|---|---|
| Compound | α4β2 1 | α7 2 | Electric Organ 2 |
| IC50 ± SEM (µM) | |||
| NTX | 60 ± 22 (6) | 390 ± 8.6 (4) | 230 ± 71 (3) |
| MeNTX | 120 ± 13 (3) | 45 ± 2.3 (2) | 9.3 ± 4.9 (4) |
| DMA-DT | 160 ± 96 (3) | 640 ± 220 (2) | >500 (1) |
| TMA-DT | 290 ± 60 (4) | 90 ± 4.3 (2) | 150 (1) |
| Compound | Ls BP | Ac BP | Ac BP Y55W |
|---|---|---|---|
| IC50 ± SEM (µM) | |||
| NTX | 140 ± 24 (2) | 71 ±1.9 (3) | 110 ± 25 (3) |
| MeNTX | 163 ± 8.7 (3) | 260 ± 71 (3) | 74 ± 5.2 (3) |
| DMA-DT | 22 ± 2.4 (3) | 28 ± 0.27 (3) | 41.4 ± 4.4 (3) |
| TMA-DT | 3.7 ± 0.39 (3) | 20 ± 2.0 (3) | 18.0 ± 2.8 (3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kem, W.R.; Andrud, K.; Bruno, G.; Xing, H.; Soti, F.; Talley, T.T.; Taylor, P. Interactions of Nereistoxin and Its Analogs with Vertebrate Nicotinic Acetylcholine Receptors and Molluscan ACh Binding Proteins. Mar. Drugs 2022, 20, 49. https://doi.org/10.3390/md20010049
Kem WR, Andrud K, Bruno G, Xing H, Soti F, Talley TT, Taylor P. Interactions of Nereistoxin and Its Analogs with Vertebrate Nicotinic Acetylcholine Receptors and Molluscan ACh Binding Proteins. Marine Drugs. 2022; 20(1):49. https://doi.org/10.3390/md20010049
Chicago/Turabian StyleKem, William R., Kristin Andrud, Galen Bruno, Hong Xing, Ferenc Soti, Todd T. Talley, and Palmer Taylor. 2022. "Interactions of Nereistoxin and Its Analogs with Vertebrate Nicotinic Acetylcholine Receptors and Molluscan ACh Binding Proteins" Marine Drugs 20, no. 1: 49. https://doi.org/10.3390/md20010049
APA StyleKem, W. R., Andrud, K., Bruno, G., Xing, H., Soti, F., Talley, T. T., & Taylor, P. (2022). Interactions of Nereistoxin and Its Analogs with Vertebrate Nicotinic Acetylcholine Receptors and Molluscan ACh Binding Proteins. Marine Drugs, 20(1), 49. https://doi.org/10.3390/md20010049