
Polyene Macrolactams from Marine and Terrestrial Sources: Structure, Production Strategies, Biosynthesis and Bioactivities





Abstract
:
1. Introduction
2. PMLs Structure

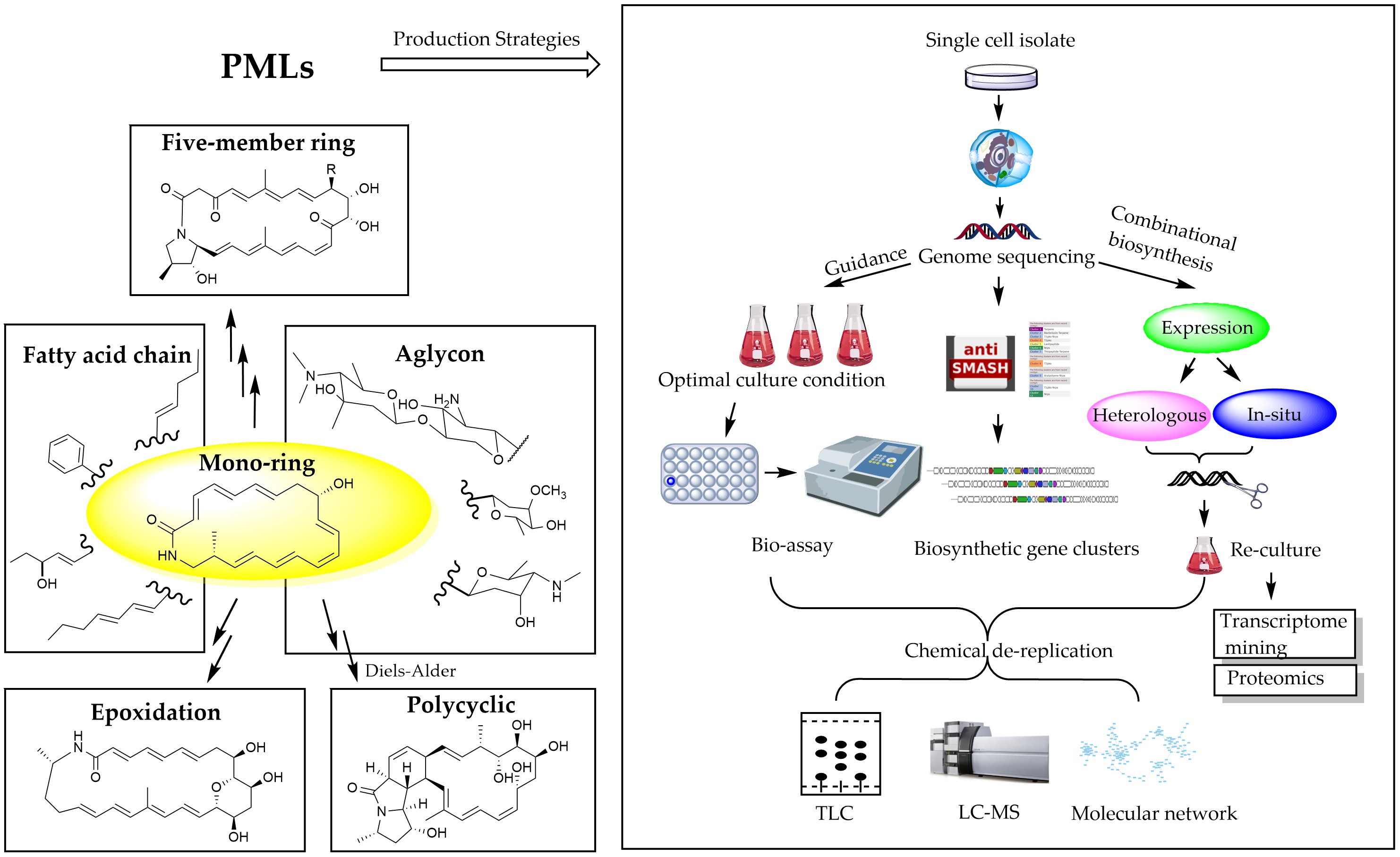{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO | Compound | Character | Member | Strain | Resource | Biological Activity | Access Strategy | Ref. |
|---|---|---|---|---|---|---|---|---|
| 1 | cyclamenol A | Mono-ring | 20 | Streptomyces sp. MHW 846 | Default | Cytotoxicity, Anti-inflammatory | Natural screening and isolation | [13] |
| 2–6 | kenalactams A–E | Mono-ring | 22–32 | Nocardiopsis sp. CG3(DSM 106572) | Saltpan | C-E Cytotoxicity antibacterial; anti-viral (HCV); C-antifungal, | Bioassay-guided metabolomic analyses | [20] |
| 7 | micromonosporin A | Mono-ring | 24 | Micromonospora sp. | Peat swamp | Unable to be evaluated | Natural screening and isolation | [21] |
| 8–9 | agA and agB | Mono-ring | 24 | Streptomyces sp. CS149 mutants | Insects | Cytotoxicity | Combinatorial Biosynthesis and Mutasynthesis | [22,23] |
| 10 | FW05328-1 | Mono-ring | 26 | Micromonospora sp. | Marine | Cytotoxicity | Natural screening and isolation | [24] |
| 11 | bombyxamycin A | Mono-ring | 26 | Streptomyces sp. SD53 | Insects | Antibacterial and Cytotoxicity | Insect-bacterial mutualisms | [4] |
| 12 | bombyxamycin C | Mono-ring | 26 | Streptomyces sp. | Insects | Antibacterial | Insect-bacterial mutualisms | [25] |
| 13–15 | lobosamide A/B/C | Mono-ring | 26 | Micromonospora sp. RL09-050-HVF-A | Marine | Anti-trypanosomal | Genomic mining | [10] |
| 16 | micromonolactam | Mono-ring | 26 | Micromonospora sp. CMS I2-32 | Marine | No activity to bacterial and fungal | Chemical screening (LC-DAD-ESIMS) | [26] |
| 17 | sceliphrolactam | Mono-ring | 26 | Streptomyces caementarium | Mud | Anti-fungal | Insect-bacterial mutualisms | [27,28] |
| 18–19 | mirilactams A/B | Mono-ring | 26 | Micromonospora sp. RL09-050-HVF,Actinosynnema mirum | Marine | Lack of anti-trypanosomal activity | Natural screening and isolation | [10,29] |
| 20 | salinilactam A | Mono-ring | 26 | Salinispora tropica | Marine | Default | Genome-Guided Discovery | [30] |
| 21 | streptolactam A | Mono-ring | 26 | Streptomyces sp. OUCMDZ-3159 | Marine | Antifungal | Natural screening and isolation | [31] |
| 22 | BE-14106 (GT32-A) | Mono-ring/fatty acidchain | 20 | Streptomyces sp | Sea island soil | Antibacterial, Cytotoxicity | Natural screening and isolation | [5,32] |
| 23 | GT-32 B | Mono-ring/fatty acidchain | 20 | Streptomyces sp | Soil | Weak antibacterial, Cytotoxicity | Natural screening and isolation | [33] |
| 24 | JBIR-150 | Mono-ring/fatty acidchain | 20 | Streptomyces sp. | Marine | Cytotoxicity | Bioassay-guided and chemical screening (LC-DAD-ESIMS) | [34] |
| 25 | heronamides C | Mono-ring/fatty acidchain | 20 | Streptomyces sp. | Marine | Non-cytotoxic, Effect on cell morphology | Chemical screening (LC-DAD-ESIMS) | [35] |
| 26 | 8-deoxyheronamide C | Mono-ring/fatty acidchain | 20 | Streptomyces sp. | Marine | Non-cytotoxic, target membrane | Bioassay-guided | [36] |
| 27 | heronamides F | Mono-ring/fatty acid chain | 20 | Streptomyces sp. SCSIO 03032 | Marine | Cytotoxicity | Chemical screening (LC-DAD-ESIMS) | [37] |
| 28 | ML449 | Mono-ring/fatty acidchain | 20 | Streptomyces MP39-85 | Marine | Antibacterial, Cytotoxicity | Bioassay-guided | [6] |
| 29 | aureoverticillactam | Mono-ring/fatty acidchain | 22 | Streptomyces aureoverticillatus (NPS001583) | Marine | Antibacterial, Cytotoxicity | Bioassay-guided | [38] |
| 30–31 | macrotermycins A/C | Mono-ring/Glycosylation | 20 | Amycolatopsis sp. M39 | Insects | Anti-bacterial and antifungal parasite | Bioassay-guided metabolomic analyses | [39] |
| 32–33 | vicenistatin/vicenistatin M | Mono-ring/Glycosylation | 20 | Streptomyces halstedii HC34 | Sea island soil | Antitumor or Against xenografted models | Natural screening and isolation | [8] |
| 34 | sannastatin | Mono-ring/Glycosylation | 20 | Streptomyces sannanensis | Animal waste | Cytotoxicity | Natural screening and isolation | [40] |
| 35 | incednine | Mono-ring/Glycosylation | 24 | Streptomyces sp. ML694-90F3 | Default | Inhibit anti-apoptotic | Natural screening and isolation | [12] |
| 36 | silvalactam | Mono-ring/Glycosylation | 24 | Streptomyces sp. | Plant | Anti-G+ bacteria, Cytotoxic activity | Natural screening and isolation | [41] |
| 37 | auroramycin | Mono-ring/Glycosylation | 24 | Streptomyces roseoporous NRRL 15998 | Default | Anti-fungal, anti-MASA | CRISPR-Cas mediated genome editing | [7] |
| 38–39 | sipanmycins A and B | Mono-ring/Glycosylation | 24 | Streptomyces sp. CS149 | Insects | Cytotoxicity | Combinatorial biosynthesis and mutasynthesis | [23] |
| 40 | mirilactam E | Mono-ring/epoxidation | 26 | Actinosynnnema mirum NBRC 14,064 | Plant | Not antibacterial activity and cytotoxicity | Co-culture | [29] |
| 41 | mirilactam D | Mono-ring/epoxidation | 26 | Actinosynnnema mirum NBRC 14,064 | Plant | No antibacterial activity and cytotoxicity | Co-culture | [29] |
| 42 | bombyxamycin B | Mono-ring/epoxidation | 26 | Streptomyces sp. SD53 | Insects | Antibacterial, cytotoxicity | Insect-bacterial mutualisms | [4] |
| 43 | dracolactam B | Mono-ring/epoxidation | 26 | Micromonospora wenchangensis HEK-797 | Marine | Default | Co-culture | [42] |
| 44 | pretilactam | Mono-ring/epoxidation | 26 | Actinosynnema pretiosum ATCC 31565 | Default | No anti-bacterial (Bs and Ca) | Genome-Guided Discovery | [43] |
| 45 | streptolactam C | Mono-ring/epoxidation | 26 | Streptomyces sp. OUCMDZ-3159 | Marine | Antifungal (Ca) | Natural screening and isolation | |
| 46–47 | cyclamenol E-F | Mono-ring/A five-member ring | 20 | Streptomyces sp. OUCMDZ-4348 | Sand, the Antarctic | E antitumor, no cytotoxicity | Bioassay-guided | [44] |
| 48 | leinamycin | Mono-ring/A five-member ring | 20 | Streptomycetes sp. S-140, Streptomyces atroolivaceus | Soil | Antibacterial, antitumor | Genomic mining | [9,45,46] |
| 49 | hitachimycin/stubomycin. | Mono-ring/A five-member ring | 22 | Actinomycete strain No. KM-4927/Streptomyces strain No. KG-2245 | Soil | Antiprotozoal/antifungal | Natural screening and isolation | [18] |
| 50–51 | niizalactam A and B | Mono-ring/A five-member ring | 26 | Streptomyces sp. NZ-6 | Terrestrial | No antimicrobial activity and cytotoxicity | Co-culture | [47] |
| 52 | piceamycin | Mono-ring/A five-member ring | 26 | Streptomyces sp. SD53 | Insects | Antibacterial, Cytotoxicity | Insect-bacterial mutualisms | [25] |
| 53 | viridenomycin | Mono-ring/A five-member ring | 26 | Streptomyces gannmycicus | Soil | Cytotoxicity | Natural screening and isolation | [48,49] |
| 54–56 | cyclamenol B–D | Polycyclic | 20–22 | Streptomyces sp. OUCMDZ-4348 | Sand, Antarctic | Only B selective cytotoxicity | Bioassay-guided | [13] |
| 57 | dracolactam A | Polycyclic | 26 | Micromonospora wenchangensis HEK-797 | Marine | Default | Co-culture | [42] |
| 58 | mirilactam C | Polycyclic | 26 | Actinosynnnema mirum NBRC 14,064 | Plant | No antibacterial activity and cytotoxicity | Co-culture | [29] |
| 59 | verticlactam B | Polycyclic | 24 | Streptomyces avermitilis SUKA17/Streptomyces spiroverticillatus JC-8444 | Default | Anti-parasitic | Metabonomics LC/MS | [50] |
| 60 | streptolactam B | Polycyclic | 26 | Streptomyces sp. OUCMDZ-3159 | Marine | No prominent cytotoxic activity | Natural screening and isolation | [31] |
| 61 | tripartilactam | Polycyclic | 26 | Streptomyces strain SNA112 | Insects | Na+/K+ATPase inhibitor | Insect-bacterial mutual and Chemical screening (LC-DAD-UV) | [51] |
| 61 | niizalactam C | Polycyclic | 26 | Streptomyces sp. NZ-6 | Terrestrial | No antimicrobial activity and cytotoxicity | Co-culture | [47] |
| 62–63 | heronamides A/B | Polycyclic/fatty acid chain | 28 | Streptomyces sp. CMB-M0406 | Marine | No antibacterial activity and cytotoxicity | Chemical screening (LC-DAD-ESIMS) | [35] |
| 64–65 | heronamides D–E | Polycyclic/fatty acid chain | 28 | Streptomyces sp. SCSIO 03032 | Marine | No antibacterial activity and cytotoxicity | OSMC (alternative medium) | [37] |
| 67–72 | heronamides G–L | Polycyclic/fatty acid chain | 28 | Streptomyces niveus | Forest soil | No antibacterial activity and cytotoxicity | Bioassay-guided | [52] |
| 73 | sagamilactam | Polycyclic/fatty acid chain | 34 | Actinomadura sp. K13-0306 | Sea island soil | Anti-trypanosomal | Chemical screening (LC-DAD-ESIMS) | [11] |
| 74–75 | ciromicins A–B | Polycyclic/glycosylation | 22 | Nocardiopsis sp. FU40 | Default | Cytotoxicity | Co-culture and metabolomic | [53] |
| 76 | macrotermycin D | Polycyclic/glycosylation | 20 | Amycolatopsis sp. M39 | Insects | No antibacterial and antifungal activity | Bioassay-guided metabolomic | [39] |
| 77 | verticilactam C | Polycyclic/epoxidation | 24 | Streptomyces avermitilis SUKA17 | Default | Anti- malaria parasite | Heterologous expression | [50] |
| 78 | verticilactam | Polycyclic/epoxidation | 24 | Streptomyces spiroverticillatus JC-8444 | Default | No biological effects | Chemical screening (LC-DAD-ESIMS) | [54] |
| 79 | macrotermycins B | Polycyclic/glycosylation and Epoxidation | 20 | Amycolatopsis sp. M39 | Insects | No antibacterial and antifungal activity | Bioassay-guided metabolomic analyses | [39] |
| 80 | cremimycin | Polycyclic/fatty acid chain/glycosylation | 22 | Streptomyces sp. MJ635-86F5 | Soil | Anti-G + bacterial, Cytotoxicity | Natural screening and isolation | [55] |
| 81 | cylindramide | Polycyclic/PTM | 26 | Halichondria cylindrata | Marine | Cytotoxicity (B16) | Natural screening and isolation | [56] |
| 82 | discodermide | Polycyclic/PTM | 26 | Discodermia dissoluta | Marine | Cytotoxicity and antifungal | Natural screening and isolation | [57] |
| 83 | HSAF | Polycyclic/PTM | 26 | Lysobacter enzymogenes, Streptomyces sp. SR107 | Soil | Anti-fungal | Bioassay-guided | [58,59] |
| 84 | 3-deOH-HSAF | Polycyclic/PTM | 26 | Lysobacter enzymogenes | Soil | Lost antifungal activity | Gene knockout | [60] |
| 85 | geodin A | Polycyclic/PTM | 26 | Geodia | Marine | Cytotoxicity | Bioassaydirected | [61] |
| 86 | clifednamide A | Polycyclic/PTM | 26 | Streptomyces sp. strain JV178 | Garden soil | Default | Combinatorial biosynthesis | [62] |
| 87–88 | frontalamides A–B | Polycyclic/PTM | 26 | Streptomyces sp. SPB78 | Marine | Antifungal | OSMAC + chemical screening | [63] |
| 89–95 | pactamide A--G | Polycyclic/PTM | 26 | Streptomyces pactum SCSIO 02999 | Default | Cytotoxicity | Promoter engineering and heterologous expression | [64] |
| 96 | butremycin | Polycyclic/PTM | 26 | Micromonospora sp. K310 | Mangrove | Weak antibacterial | LC- MS | [65] |
| 97 | ikarugamycin | Polycyclic/PTM | 26 | Streptomyces sp. NO 8603 | Soil | Antiprotozoal, anti-G+ bacterial | Natural screening and isolation | [66] |
| 98–99 | lysobacteramide A and B | Polycyclic/PTM | 26 | Lysobacter enzymogenes C3 | Default | Cytotoxicity; B- Anti-fungal | OSMC | [67] |
| 100 | alteramide A | Polycyclic/PTM | 26 | Alteromonas sp. | Marine | Cytotoxicity | Natural screening and isolation | [68] |
| 101 | aburatubolactam A | Polycyclic/PTM | 26 | Streptomyces sp. SCRC A-20 | Marine | Inhibit superoxide anion generation | Default | [69] |
| 102 | alteramide B | Polycyclic/PTM | 26 | Lysobacter enzymogenes C3 | Default | Anti-fungal(yeast and Ca) | Natural screening and isolation | [70] |
| 103–108 | combamides A−F | Polycyclic/PTM | 26 | Streptomyces sp. S10 | Garden soil | F weakly inhibited SPI-1 | Combinatorial Biosynthesis | [71] |
3. PMLs Production Strategies
3.1. Culture Strategies for PMLs Production
3.1.1. Elicitors Supplementary
3.1.2. Bacterial Co-Cultivation
3.2. Chemical Analytical De-Replication
3.3. Genetic Tools for PMLs Biosynthesis
3.3.1. Genomic Mining
3.3.2. Heterologous or In Situ Expression and Synthetic Biology
3.3.3. Chemical Synthesis under the Guidance of Bioinformatics
4. PMLs Biosynthetic Pathways
4.1. PMLs Initial β-Amino Acid Units
4.2. PMLs Extender Unit Incorporation and Cyclization
4.3. Epoxy Groups and Intramolecular Polycyclic Systems
4.4. Glycosylated Groups
5. Mechanism of Action
6. The Challenges and Opportunities of Current Research on PMLs
6.1. Bottleneck Limitation for PMLs Isolation and Storage
6.2. PMLs Poor Solubility Blocking Bioactivities Assessment
6.3. Multidisciplinary Effort toward Further PMLs Discovery
6.3.1. Innovation of Strain Source
6.3.2. Establishment of Chemical Tool Enzymology Library
6.3.3. Pharmacology and Mechanism of Bioactivity
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Bergeijk, D.A.; Terlouw, B.R.; Medema, M.H.; van Wezel, G.P. Ecology and genomics of Actinobacteria: New concepts for natural product discovery. Nat. Rev. Microbiol. 2020, 18, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Gui, M.; Li, H.; Yu, C.; Li, H.; Zeng, Z.; Sun, P. Secondary Metabolites from Marine Micromonospora: Chemistry and Bioactivities. Chem. Biodiver. 2020, 17, e2000024. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, Y.H.; Beom, J.Y.; Chung, B.; Shin, Y.; Byun, W.S.; Moon, K.; Bae, M.; Lee, S.K.; Oh, K.B.; Shin, J.; et al. Bombyxamycins A and B, Cytotoxic Macrocyclic Lactams from an Intestinal Bacterium of the Silkworm Bombyx mori. Org. Lett. 2019, 21, 1804–1808. [Google Scholar] [CrossRef] [PubMed]
- Kojiri, K.; Nakajima, S.; Suzuki, H.; Kondo, H.; Suda, H. A new macrocyclic lactam antibiotic, BE-14106. I. Taxonomy, isolation, biological activity and structural elucidation. J. Antibiot. 1992, 45, 868–874. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, H.; Degnes, K.F.; Dikiy, A.; Fjaervik, E.; Klinkenberg, G.; Zotchev, S.B. Insights into the evolution of macrolactam biosynthesis through cloning and comparative analysis of the biosynthetic gene cluster for a novel macrocyclic lactam, ML-449. Appl. Environ. Microbiol. 2010, 76, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.H.; Wong, F.T.; Yeo, W.L.; Ching, K.C.; Lim, Y.W.; Heng, E.; Chen, S.; Tsai, D.J.; Lauderdale, T.L.; Shia, K.S.; et al. Auroramycin: A Potent Antibiotic from Streptomyces roseosporus by CRISPR-Cas9 Activation. Chembiochem 2018, 19, 1716–1719. [Google Scholar] [CrossRef]
- Shindo, K.; Kamishohara, M.; Odagawa, A.; Matsuoka, M.; Kawai, H. Vicenistatin, a novel 20-membered macrocyclic lactam antitumor antibiotic. J. Antibiot. 1993, 46, 1076–1081. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Asano, K.; Kawamoto, I.; Takiguchi, T.; Katsumata, S.; Takahashi, K.; Nakano, H. Leinamycin, a new antitumor antibiotic from Streptomyces: Producing organism, fermentation and isolation. J. Antibiot. 1989, 42, 1768–1774. [Google Scholar]
- Schulze, C.J.; Donia, M.S.; Siqueira-Neto, J.L.; Ray, D.; Raskatov, J.A.; Green, R.E.; McKerrow, J.H.; Fischbach, M.A.; Linington, R.G. Genome-Directed Lead Discovery: Biosynthesis, Structure Elucidation, and Biological Evaluation of Two Families of Polyene Macrolactams against Trypanosoma brucei. ACS Chem. Biol. 2015, 10, 2373–2381. [Google Scholar] [CrossRef]
- Kimura, T.; Iwatsuki, M.; Asami, Y.; Ishiyama, A.; Hokari, R.; Otoguro, K.; Matsumoto, A.; Sato, N.; Shiomi, K.; Takahashi, Y.; et al. Anti-trypanosomal compound, sagamilactam, a new polyene macrocyclic lactam from Actinomadura sp. K13-0306. J. Antibiot. 2016, 69, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Futamura, Y.; Sawa, R.; Umezawa, Y.; Igarashi, M.; Nakamura, H.; Hasegawa, K.; Yamasaki, M.; Tashiro, E.; Takahashi, Y.; Akamatsu, Y.; et al. Discovery of incednine as a potent modulator of the anti-apoptotic function of Bcl-xL from microbial origin. J. Am. Chem. Soc. 2008, 130, 1822–1823. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Fan, Y.; Zhu, G.; Chen, H.; Zhu, W.; Fu, P. Polycyclic Macrolactams Generated via Intramolecular Diels-Alder Reactions from an Antarctic Streptomyces Species. Org. Lett. 2019, 21, 4816–4820. [Google Scholar] [CrossRef] [PubMed]
- Jeanvoine, A.; Rocchi, S.; Bellanger, A.P.; Reboux, G.; Millon, L. Azole-resistant Aspergillus fumigatus: A global phenomenon originating in the environment? Med. Mal. Infect. 2020, 50, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Meis, J.F.; Chowdhary, A.; Rhodes, J.L.; Fisher, M.C.; Verweij, P.E. Clinical implications of globally emerging azole resistance in Aspergillus fumigatus. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef]
- Kaur, L.; Abhijeet; Jain, S.K. Safe and Effective Delivery of Amphotericin B: A Survey of Patents. Recent Pat. Nanotechnol. 2017, 11, 214–234. [Google Scholar] [CrossRef]
- Ottilie, S.; Goldgof, G.M.; Cheung, A.L.; Walker, J.L.; Vigil, E.; Allen, K.E.; Antonova-Koch, Y.; Slayman, C.W.; Suzuki, Y.; Durrant, J.D. Two inhibitors of yeast plasma membrane ATPase 1 (ScPma1p): Toward the development of novel antifungal therapies. J. Cheminform. 2018, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, R.; de Lera, A.R. Natural polyenic macrolactams and polycyclic derivatives generated by transannular pericyclic reactions: Optimized biogenesis challenging chemical synthesis. Nat. Prod. Rep. 2021, 38, 1136–1220. [Google Scholar] [CrossRef]
- Messaoudi, O.; Sudarman, E.; Bendahou, M.; Jansen, R.; Stadler, M.; Wink, J. Kenalactams A-E, Polyene Macrolactams Isolated from Nocardiopsis sp. CG3. J. Nat. Prod. 2019, 82, 1081–1088. [Google Scholar] [CrossRef]
- Thawai, C.; Kittakoop, P.; Tanasupawat, S.; Suwanborirux, K.; Thebtaranonth, Y. Micromonosporin A, a novel 24-membered polyene lactam macrolide from Micromonospora sp. isolated from peat swamp forest. Chem. Biodiver. 2010, 1, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Malmierca, M.G.; Pérez-Victoria, I.; Serrano, J.M.; Reyes, F.; Olano, C. New Sipanmycin Analogues Generated by Combinatorial Biosynthesis and Mutasynthesis Approaches Relying on the Substrate Flexibility of Key Enzymes in the Biosynthetic Pathway. Appl. Env. Microb. 2019, 86, e02453-19. [Google Scholar] [CrossRef] [PubMed]
- Malmierca, M.G.; Pérez-Victoria, I.; Martín, J.; Reyes, F.; Méndez, C.; Olano, C.; Salas, J.A. Cooperative involvement of glycosyltransferases in the transfer of aminosugars in the 1 biosynthesis of the macrolactam sipanmycin by Streptomyces sp. CS149. Appl. Environ. Microbiol. 2018, 84, e01462-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Y.L.; Wu, Y.D.; Wang, C.X.; Lin, R.; Xie, Y.; Fang, D.S.; Jiang, H.; Lian, Y.Y. Structure elucidation and antitumour activity of a new macrolactam produced by marine-derived actinomycete Micromonospora sp. FIM05328. Nat. Prod. Res. 2018, 32, 2133–2138. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.H.; Kang, S.; Byun, W.S.; Jeon, C.W.; Oh, D.C. Absolute Configuration and Antibiotic Activity of Piceamycin. J. Nat. Prod. 2020, 83, 277–285. [Google Scholar] [CrossRef]
- Skellam, E.J.; Stewart, A.K.; Strangman, W.K.; Wright, J.L. Identification of micromonolactam, a new polyene macrocyclic lactam from two marine Micromonospora strains using chemical and molecular methods: Clarification of the biosynthetic pathway from a glutamate starter unit. J. Antibiot. 2013, 66, 431–441. [Google Scholar] [CrossRef]
- Oh, D.C.; Poulsen, M.; Currie, C.R.; Clardy, J. Sceliphrolactam, a Polyene Macrocyclic Lactam from a Wasp-Associated Streptomyces sp. Org. Lett. 2011, 13, 752–755. [Google Scholar] [CrossRef]
- Low, Z.J.; Pang, L.M.; Ding, Y.; Cheang, Q.W.; Le Mai Hoang, K.; Thi Tran, H.; Li, J.; Liu, X.W.; Kanagasundaram, Y.; Yang, L.; et al. Identification of a biosynthetic gene cluster for the polyene macrolactam sceliphrolactam in a Streptomyces strain isolated from mangrove sediment. Sci. Rep. 2018, 8, 1594. [Google Scholar] [CrossRef]
- Hoshino, S.; Ozeki, M.; Wong, C.P.; Zhang, H.; Hayashi, F.; Awakawa, T.; Morita, H.; Onaka, H.; Abe, I. Mirilactams C-E, Novel Polycyclic Macrolactams Isolated from Combined-Culture of Actinosynnema mirum NBRC 14064 and Mycolic Acid-Containing Bacterium. Chem. Pharm. Bull. 2018, 66, 660–667. [Google Scholar] [CrossRef] [Green Version]
- Udwary, D.W.; Zeigler, L.; Asolkar, R.N.; Singan, V.; Lapidus, A.; Fenical, W.; Jensen, P.R.; Moore, B.S. Genome sequencing reveals complex secondary metabolome in the marine actinomycete Salinispora tropica. Proc. Natl. Acad. Sci. USA 2007, 104, 10376–10381. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Wang, D.; Zhang, R.; Wang, Y.; Kong, F.; Fu, P.A.-O.; Zhu, W.A.-O. Novel Macrolactams from a Deep-Sea-Derived Streptomyces Species. Mar. Drugs 2021, 19, 13. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, H.; Degnes, K.F.; Sletta, H.; Fjaervik, E.; Dikiy, A.; Herfindal, L.; Bruheim, P.; Klinkenberg, G.; Bredholt, H.; Nygård, G.; et al. Biosynthesis of macrolactam BE-14106 involves two distinct PKS systems and amino acid processing enzymes for generation of the aminoacyl starter unit. Chem. Biol. 2009, 16, 1109–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, I.; Oda, Y.; Nishiie, Y.; Ochiai, K.; Mizukami, T. GT32-B new 20-membered macrocyclic lactam antibiotic. J. Antibiot. 1997, 50, 186–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahara, T.; Fujiwara, T.; Kagaya, N.; Shin-Ya, K. JBIR-150, a novel 20-membered polyene macrolactam from marine-derived Streptomyces sp. OPMA00071. J. Antibiot. 2018, 71, 390–392. [Google Scholar] [CrossRef] [PubMed]
- Raju, R.; Piggott, A.M.; Conte, M.M.; Capon, R.J. Heronamides A-C, new polyketide macrolactams from an Australian marine-derived Streptomyces sp. A biosynthetic case for synchronized tandem electrocyclization. Org. Biomol. Chem. 2010, 8, 4682–4689. [Google Scholar] [CrossRef]
- Sugiyama, R.; Nishimura, S.; Matsumori, N.; Tsunematsu, Y.; Hattori, A.; Kakeya, H. Structure and biological activity of 8-deoxyheronamide C from a marine-derived Streptomyces sp.: Heronamides target saturated hydrocarbon chains in lipid membranes. J. Am. Chem. Soc. 2014, 136, 5209–5212. [Google Scholar] [CrossRef]
- Zhang, W.; Li, S.; Zhu, Y.; Chen, Y.; Chen, Y.; Zhang, H.; Zhang, G.; Tian, X.; Pan, Y.; Zhang, S.; et al. Heronamides D-F, polyketide macrolactams from the deep-sea-derived Streptomyces sp. SCSIO 03032. J. Nat. Prod. 2014, 77, 388–391. [Google Scholar] [CrossRef]
- Mitchell, S.S.; Nicholson, B.; Teisan, S.; Lam, K.S.; Potts, B. Aureoverticillactam, a Novel 22-Atom Macrocyclic Lactam from the Marine Actinomycete Streptomyces aureoverticillatus. J. Nat. Prod. 2004, 67, 1400–1402. [Google Scholar] [CrossRef]
- Beemelmanns, C.; Ramadhar, T.R.; Kim, K.H.; Klassen, J.L.; Cao, S.; Wyche, T.P.; Hou, Y.; Poulsen, M.; Bugni, T.S.; Currie, C.R.; et al. Macrotermycins A–D, Glycosylated Macrolactams from a Termite-Associated Amycolatopsis sp. M39. Org. Lett. 2017, 19, 1000–1003. [Google Scholar] [CrossRef]
- Yang, S.X.; Gao, J.M.; Zhang, A.L.; Laatsch, H. RETRACTED: Sannastatin, a novel toxic macrolactam polyketide glycoside produced by actinomycete Streptomyces sannanensis. Bioorganic. Med. Chem. Lett. 2011, 21, 3905–3908. [Google Scholar] [CrossRef]
- Schulz, D.; Nachtigall, J.; Geisen, U.; Kalthoff, H.; Imhoff, J.F.; Fiedler, H.P.; Süssmuth, R.D. Silvalactam, a 24-membered macrolactam antibiotic produced by Streptomyces sp. Tü 6392. J. Antibiot. 2012, 65, 369–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, S.; Okada, M.; Awakawa, T.; Asamizu, S.; Onaka, H.; Abe, I. Mycolic Acid Containing Bacterium Stimulates Tandem Cyclization of Polyene Macrolactam in a Lake Sediment Derived Rare Actinomycete. Org. Lett. 2017, 19, 4992–4995. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, X.; Sun, G.; Li, L.; Wu, L. Genome-Guided Discovery of Pretilactam from Actinosynnema pretiosum ATCC 31565. Molecules 2019, 24, 2281. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Wang, J.; Chen, H.; Wang, Y.; Fu, P. Cyclamenols E and F, two diastereoisomeric bicyclic macrolactams with cyclopentane moiety from an Antarctic Streptomyces species. Org. Chem. Front. 2019, 7, 310–317. [Google Scholar] [CrossRef]
- Cheng, Y.Q.; Tang, G.L.; Shen, B. Identification and Localization of the Gene Cluster Encoding Biosynthesis of the Antitumor Macrolactam Leinamycin in Streptomyces atroolivaceus S-140. J. Bacteriol. 2002, 184, 7013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.L.; Cheng, Y.Q.; Shen, B. Leinamycin Biosynthesis Revealing Unprecedented Architectural Complexity for a Hybrid Polyketide Synthase and Nonribosomal Peptide Synthetase. Chem. Biodiver. 2004, 11, 33–45. [Google Scholar] [CrossRef]
- Hoshino, S.; Okada, M.; Wakimoto, T.; Zhang, H.; Hayashi, F.; Onaka, H.; Abe, I. Niizalactams A-C, Multicyclic Macrolactams Isolated from Combined Culture of Streptomyces with Mycolic Acid-Containing Bacterium. J. Nat. Prod. 2015, 78, 3011–3017. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Furihata, K.; Hayakawa, Y.; Seto, H. Structure of viridenomycin. Tetrahedron Lett. 1991, 32, 659–662. [Google Scholar] [CrossRef]
- Mulholland, N.P.; Pattenden, G. A silicon-tethered tandem radical cyclisation–trapping strategy to the fully substituted cyclopentene ring in viridenomycin. Tetrahedron 2008, 64, 7400–7406. [Google Scholar] [CrossRef]
- Nogawa, T.A.-O.; Terai, A.; Amagai, K.A.-O.; Hashimoto, J.; Futamura, Y.; Okano, A.; Fujie, M.; Satoh, N.; Ikeda, H.A.-O.; Shin-Ya, K.A.-O.; et al. Heterologous Expression of the Biosynthetic Gene Cluster for Verticilactam and Identification of Analogues. J. Nat. Prod. 2020, 83, 3598–3605. [Google Scholar] [CrossRef]
- Park, S.H.; Moon, K.; Bang, H.S.; Kim, S.H.; Kim, D.G.; Oh, K.B.; Shin, J.; Oh, D.C. Tripartilactam, a Cyclobutane-Bearing Tricyclic Lactam from a Streptomyces sp. in a Dung Beetle’s Brood Ball. Org. Lett. 2012, 14, 1258–1261. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Han, L.; Jiang, Y.; Li, G.; Zheng, Z.; Cao, B.; Guan, P.; Mu, Y.; Lin, B.; Huang, X. Heronamides G–L, polyene macrolactams from Streptomyces niveus. RSC Adv. 2018, 8, 17121–17131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derewacz, D.K.; Covington, B.C.; McLean, J.A.; Bachmann, B.O. Mapping Microbial Response Metabolomes for Induced Natural Product Discovery. ACS Chem. Biol. 2015, 10, 1998–2006. [Google Scholar] [CrossRef] [Green Version]
- Nogawa, T.; Okano, A.; Takahashi, S.; Uramoto, M.; Konno, H.; Saito, T.; Osada, H. Verticilactam, a New Macrolactam Isolated from a Microbial Metabolite Fraction Library. Org. Lett. 2011, 12, 4564–4567. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, M.; Tsuchida, T.; Kinoshita, N.; Kamijima, M.; Sawa, R.; Sawa, T.; Naganawa, H.; Hamada, M.; Takeuchi, T.; Yamazaki, K.; et al. Cremimycin, a novel 19-membered macrocyclic lactam antibiotic, from Streptomyces sp. J. Antibiot. 1998, 51, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanazawa, S.; Fusetani, N.; Matsunaga, S. Cylindramide: Cytotoxic tetramic acid lactam from the marine sponge Halichondria cylindrata Tanita & Hoshino. Tetrahedron Lett. 1993, 34, 1065–1068. [Google Scholar]
- Gunasekera, S.P.; Gunasekera, M.; McCarthy, P. Discodermide: A new bioactive macrocyclic lactam from the marine sponge Discodermia dissoluta. J. Org. Chem. 1991, 56, 4830–4833. [Google Scholar] [CrossRef]
- Lou, L.; Qian, G.; Xie, Y.; Hang, J.; Chen, H.; Zaleta-Rivera, K.; Li, Y.; Shen, Y.; Dussault, P.H.; Liu, F. Biosynthesis of HSAF, a Tetramic Acid-containing Macrolactam from Lysobacter enzymogenes. J. Am. Chem. Soc. 2011, 133, 643–645. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chen, H.; Ding, Y.; Xie, Y.; Wang, H.; Cerny, R.L.; Shen, Y.; Du, L. Iterative Assembly of Two Separate Polyketide Chains by the Same Single-Module Bacterial Polyketide Synthase in the Biosynthesis of HSAF. Angew. Chem. Int. Ed. 2014, 53, 7654–7660. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Huffman, J.; Li, Y.; Du, L.; Shen, Y. 3-Hydroxylation of the polycyclic tetramate macrolactam in the biosynthesis of antifungal HSAF from Lysobacter enzymogenes C3. MedChemComm 2012, 3, 982–986. [Google Scholar] [CrossRef]
- Capon, R.J.; Skene, C.; Lacey, E.; Gill, J.H.; Wadsworth, D.; Friedel, T. Geodin A magnesium salt: A novel nematocide from a southern australian marine sponge, Geodia. J. Nat. Prod. 1999, 62, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Ding, E.; Blodgett, J. Native and Engineered Clifednamide Biosynthesis in Multiple Streptomyces spp. ACS Synth. Biol. 2018, 7, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Blodgett, J.; Oh, D.C.; Ca, O.S.; Currie, C.R.; Kolter, R.; Clardy, J. Common biosynthetic origins for polycyclic tetramate macrolactams from phylogenetically diverse bacteria. Proc. Natl. Acad. Sci. USA 2010, 107, 11692–11697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Zhang, W.; Zhang, G.; Zhu, Y.; Chen, Y.; Liu, W.; Yuan, C.; Zhang, Q.; Zhang, H.; Zhang, L.; et al. Activation and characterization of a cryptic gene cluster reveals a cyclization cascade for polycyclic tetramate macrolactams. Chem. Sci. 2017, 8, 1607–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyeremeh, K.; Acquah, K.S.; Sazak, A.; Houssen, W.; Tabudravu, J.; Deng, H.; Jaspars, M. Butremycin, the 3-hydroxyl derivative of ikarugamycin and a protonated aromatic tautomer of 5′-methylthioinosine from a Ghanaian Micromonospora sp. K310. Mar. Drugs 2014, 12, 999–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jomon, K.; Kuroda, Y.; Ajisaka, M.; Sakai, H. A new antibiotic, ikarugamycin. J. Antibiot. 1972, 25, 271–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Wu, P.; Wright, S.J.; Du, L.; Wei, X. Bioactive Polycyclic Tetramate Macrolactams from Lysobacter enzymogenes and Their Absolute Configurations by Theoretical ECD Calculations. J. Nat. Prod. 2015, 78, 1841–1847. [Google Scholar] [CrossRef]
- Shigemori, H.; Bae, M.A.; Yazawa, K.; Sasaki, T.; Kobayashi, J. Alteramide A, a new tetracyclic alkaloid from a bacterium Alteromonas sp. associated with the marine sponge Halichondria okadai. J. Org. Chem. 1992, 57, 4317–4320. [Google Scholar] [CrossRef]
- Bae, M.-A.; Yamada, K.; Ijuin, Y.; Tsuji, T.; Yazawa, K.; Uemura, D. Aburatubolactam A, a novel inhibitor of superoxide anion generation from a marine microorganism. Heterocycl. Commun. 1996, 2, 315–318. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Y.; Li, Z.; Zhang, J.; Lu, C.; Wang, H.; Shen, Y.; Du, L. Alteramide B is a microtubule antagonist of inhibiting Candida albicans. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 2097–2106. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, H.; Song, R.; Chen, J.; Li, T.; Li, Y.; Du, L.; Shen, Y. Targeted Discovery and Combinatorial Biosynthesis of Polycyclic Tetramate Macrolactam Combamides A–E. Org. Lett. 2018, 20, 3504–3508. [Google Scholar] [CrossRef] [PubMed]
- Azizah, M.; Pripdeevech, P.; Thongkongkaew, T.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. UHPLC-ESI-QTOF-MS/MS-Based Molecular Networking Guided Isolation and Dereplication of Antibacterial and Antifungal Constituents of Ventilago denticulata. Antibiotics 2020, 9, 606. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Pan, C.; Cen, S.; Fu, L.; Cao, X.; Wang, H.; Wang, K.; Wu, B. Comparative metabolomics reveals defence-related modification of citrinin by Penicillium citrinum within a synthetic Penicillium-Pseudomonas community. Environ. Microbiol. 2019, 21, 496–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Felicio, R.; Ballone, P.; Bazzano, C.F.; Alves, L.F.G.; Sigrist, R.; Infante, G.P.; Niero, H.; Rodrigues-Costa, F.; Fernandes, A.Z.N.; Tonon, L.A.C.; et al. Chemical elicitors induce rare bioactive secondary metabolites in deep-sea bacteria under laboratory conditions. Metabolites 2021, 11, 107. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Waters, A.L.; Sims, J.W.; Fullmer, A.; Ellison, S.; Hamann, M.T. Complex marine natural products as potential epigenetic and production regulators of antibiotics from a marine Pseudomonas aeruginosa. Microb. Ecol. 2013, 65, 1068–1075. [Google Scholar] [CrossRef] [Green Version]
- Biarnes-Carrera, M.; Breitling, R.; Takano, E. Butyrolactone signalling circuits for synthetic biology. Curr. Opin. Chem. Biol. 2015, 28, 91–98. [Google Scholar] [CrossRef]
- Barreales, E.G.; Payero, T.D.; Jambrina, E.; Aparicio, J.F. The Streptomyces filipinensis Gamma-Butyrolactone System Reveals Novel Clues for Understanding the Control of Secondary Metabolism. Appl. Environ. Microbiol. 2020, 86, e00443-20. [Google Scholar] [CrossRef]
- Liu, L.; Ji, M.; Wang, F.; Tian, Z.; Wang, S. N-acyl-L-homoserine lactones release and microbial community changes in response to operation temperature in an anammox biofilm reactor. Chemosphere 2020, 262, 127602. [Google Scholar] [CrossRef]
- Readel, E.; Portillo, A.; Talebi, M.; Armstrong, D.W. Enantiomeric separation of quorum sensing autoinducer homoserine lactones using GC-MS and LC-MS. Anal. Bioanal. Chem. 2020, 412, 2927–2937. [Google Scholar] [CrossRef]
- Luo, H.; Liu, H.; Cao, Y.; Xu, D.; Mao, Z.; Mou, Y.; Meng, J.; Lai, D.; Liu, Y.; Zhou, L. Enhanced production of botrallin and TMC-264 with in situ macroporous resin adsorption in mycelial liquid culture of the endophytic fungus Hyalodendriella sp. Ponipodef. Molecules 2014, 19, 14221–14234. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Escribano, J.P.; Bibb, M.J. Engineering Streptomyces coelicolor for heterologous expression of secondary metabolite gene clusters. Microb. Biotechnol. 2011, 4, 207–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Kakeya, H. Cryptic Chemical Communication: Secondary Metabolic Responses Revealed by Microbial Co-culture. Chem. Asian J. 2020, 15, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cai, K.; Yao, R. A new macrolactam derivative from the marine actinomycete HF-11225. J. Antibiot. 2018, 71, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.A.; Tanaka, M.; Tanouchi, K.; Ishikawa, N.; Ahmed, F.; Sadhu, S.K.; Ishibashi, M. Hes1-Binding Compounds Isolated by Target Protein Oriented Natural Products Isolation (TPO-NAPI). J. Nat. Prod. 2017, 80, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.L.; Wang, Y.J.; Chung, H.H.; Chen, K.C.; Shen, T.L.; Hsu, C.C. Molecular networking as a dereplication strategy for monitoring metabolites of natural product treated cancer cells. Rapid Commun. Mass Spectrom. 2020, 34, e8549. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; de Felicio, R.; Fenner, A.; et al. Molecular networking as a dereplication strategy. J. Nat. Prod. 2013, 76, 1686–1699. [Google Scholar] [CrossRef] [Green Version]
- Saha, S.; Esposito, G.; Urajová, P.; Mareš, J.; Ewe, D.; Caso, A.; Macho, M.; Delawská, K.; Kust, A.; Hrouzek, P.; et al. Discovery of Unusual Cyanobacterial Tryptophan-Containing Anabaenopeptins by MS/MS-Based Molecular Networking. Molecules 2020, 25, 3786. [Google Scholar] [CrossRef]
- Medema, M.H.; Paalvast, Y.; Nguyen, D.D.; Melnik, A.; Dorrestein, P.C.; Takano, E.; Breitling, R. Pep2Path: Automated mass spectrometry-guided genome mining of peptidic natural products. PLoS Comput. Biol. 2014, 10, e1003822. [Google Scholar] [CrossRef]
- Ding, C.Y.G.; Pang, L.M.; Liang, Z.X.; Goh, K.K.K.; Glukhov, E.; Gerwick, W.H.; Tan, L.T. MS/MS-Based Molecular Networking Approach for the Detection of Aplysiatoxin-Related Compounds in Environmental Marine Cyanobacteria. Mar. Drugs 2018, 16, 505. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Lu, Q.; Sun, C.; Lukianov, D.; Osterman, I.A.; Sergiev, P.V.; Dontsova, O.A.; Hu, X.; You, X.; Liu, S.; et al. Hetiamacin E and F, New Amicoumacin Antibiotics from Bacillus subtilis PJS Using MS/MS-Based Molecular Networking. Molecules 2020, 25, 4446. [Google Scholar] [CrossRef]
- Howarth, A.; Ermanis, K.; Goodman, J.M. DP4-AI automated NMR data analysis: Straight from spectrometer to structure. Chem. Sci. 2020, 11, 4351–4359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandão, P.; Torimura, M.; Kurane, R.; Bull, A. Dereplication for biotechnology screening: PyMS analysis and PCR-RFLP-SSCP (PRS) profiling of 16S rRNA genes of marine and terrestrial actinomycetes. Appl. Microbiol. Biotechnol. 2002, 58, 77–83. [Google Scholar] [PubMed]
- Carro, L.; Nouioui, I. Taxonomy and systematics of plant probiotic bacteria in the genomic era. AIMS Microbiol. 2017, 3, 383–412. [Google Scholar] [CrossRef]
- Pan, G.; Xu, Z.; Guo, Z.; Hindra Ma, M.; Yang, D.; Zhou, H.; Gansemans, Y.; Zhu, X.; Huang, Y.; Zhao, L.X. Discovery of the leinamycin family of natural products by mining actinobacterial genomes. Proc. Natl. Acad. Sci. USA 2017, 114, E11131–E11140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flinspach, K.; Westrich, L.; Kaysser, L.; Siebenberg, S.; Gomez-Escribano, J.P.; Bibb, M.; Gust, B.; Heide, L. Heterologous expression of the biosynthetic gene clusters of coumermycin A(1), clorobiocin and caprazamycins in genetically modified Streptomyces coelicolor strains. Biopolymers 2010, 93, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Linares-Otoya, L.; Linares-Otoya, V.; Armas-Mantilla, L.; Blanco-Olano, C.; Crüsemann, M.; Ganoza-Yupanqui, M.L.; Campos-Florian, J.; König, G.M.; Schäberle, T.F. Identification and heterologous expression of the kocurin biosynthetic gene cluster. Microbiology 2017, 163, 1409–1414. [Google Scholar] [CrossRef]
- Novakova, R.; Núñez, L.E.; Homerova, D.; Knirschova, R.; Feckova, L.; Rezuchova, B.; Sevcikova, B.; Menéndez, N.; Morís, F.; Cortés, J.; et al. Increased heterologous production of the antitumoral polyketide mithramycin A by engineered Streptomyces lividans TK24 strains. Appl. Microbiol. Biotechnol. 2018, 102, 857–869. [Google Scholar] [CrossRef]
- Sekurova, O.N.; Schneider, O.; Zotchev, S.B. Novel bioactive natural products from bacteria via bioprospecting, genome mining and metabolic engineering. Microb. Biotechnol. 2019, 12, 828–844. [Google Scholar] [CrossRef] [Green Version]
- Becerril, A.; Pérez-Victoria, I.; Ye, S.; Braña, A.F.; Martín, J.; Reyes, F.; Salas, J.A.; Méndez, C. Discovery of Cryptic Largimycins in Streptomyces Reveals Novel Biosynthetic Avenues Enriching the Structural Diversity of the Leinamycin Family. ACS Chem. Biol. 2020, 15, 1541–1553. [Google Scholar] [CrossRef]
- Li, L.; Koirala, B.; Hernandez, Y.; MacIntyre, L.W.; Ternei, M.A.; Russo, R.; Brady, S.F. Identification of structurally diverse menaquinone-binding antibiotics with in vivo activity against multidrug-resistant pathogens. Nat. Microbiol. 2022, 7, 120–131. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, W.; Saha, S.; Zhang, C. Recent Advances in Discovery, Biosynthesis and Genome Mining of Medicinally Relevant Polycyclic Tetramate Macrolactams. Curr. Top. Med. Chem. 2016, 16, 1727–1739. [Google Scholar] [CrossRef] [PubMed]
- Morinaka, B.I.; Lakis, E.; Verest, M.; Helf, M.J.; Scalvenzi, T.; Vagstad, A.L.; Sims, J.; Sunagawa, S.; Gugger, M.; Piel, J. Natural noncanonical protein splicing yields products with diverse β-amino acid residues. Science 2018, 359, 779–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyanaga, A.; Kudo, F.; Eguchi, T. Mechanisms of β-amino acid incorporation in polyketide macrolactam biosynthesis. Curr. Opin. Chem. Biol. 2016, 35, 58–64. [Google Scholar] [CrossRef]
- Kudo, F.; Miyanaga, A.; Eguchi, T. Biosynthesis of natural products containing β-amino acids. Nat. Prod. Rep. 2014, 31, 1056–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogasawara, Y.; Kakinuma, K.; Eguchi, T. Involvement of glutamate mutase in the biosynthesis of the unique starter unit of the macrolactam polyketide antibiotic vicenistatin. J. Antibiot. 2005, 58, 468–472. [Google Scholar] [CrossRef]
- Miyanaga, A.; Iwasawa, S.; Shinohara, Y.; Kudo, F.; Eguchi, T. Structure-based analysis of the molecular interactions between acyltransferase and acyl carrier protein in vicenistatin biosynthesis. Proc. Natl. Acad. Sci. USA 2016, 113, 1802–1807. [Google Scholar] [CrossRef] [Green Version]
- Minami, A.; Eguchi, T. Substrate flexibility of vicenisaminyltransferase VinC involved in the biosynthesis of vicenistatin. J. Am. Chem. Soc. 2007, 129, 5102–5107. [Google Scholar] [CrossRef]
- Takaishi, M.; Kudo, F.; Eguchi, T. A unique pathway for the 3-aminobutyrate starter unit from L-glutamate through β-glutamate during biosynthesis of the 24-membered macrolactam antibiotic, incednine. Org. Lett. 2012, 14, 4591–4593. [Google Scholar] [CrossRef]
- Takaishi, M.; Kudo, F.; Eguchi, T. Identification of the incednine biosynthetic gene cluster: Characterization of novel β-glutamate-β-decarboxylase IdnL3. J. Antibiot. 2013, 66, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Cieślak, J.; Miyanaga, A.; Takaku, R.; Takaishi, M.; Amagai, K.; Kudo, F.; Eguchi, T. Biochemical characterization and structural insight into aliphatic β-amino acid adenylation enzymes IdnL1 and CmiS6. Proteins 2017, 85, 1238–1247. [Google Scholar] [CrossRef]
- Cieślak, J.; Miyanaga, A.; Takaishi, M.; Kudo, F.; Eguchi, T. Functional and structural characterization of IdnL7, an adenylation enzyme involved in incednine biosynthesis. Acta Crystallogr. F Struct. Biol. Commun. 2019, 75 Pt 4, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Kudo, F.; Kawamura, K.; Uchino, A.; Miyanaga, A.; Eguchi, T. Genome Mining of the Hitachimycin Biosynthetic Gene Cluster: Involvement of a Phenylalanine-2,3-aminomutase in Biosynthesis. ChemBioChem 2015, 16, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Batsanov, A.S.; Knowles, J.P.; Lightfoot, A.P.; Maw, G.; Thirs, C.E.; Twiddle, S.; Whiting, A. A novel, efficient, diastereo- and enantioselective Mukaiyama aldol-based synthesis of a vinyl cyclopentanone core derivative of viridenomycin. Organic. Lett. 2007, 9, 5565–5568. [Google Scholar] [CrossRef] [PubMed]
- Constandinos, B.L. A concise synthesis of the functionalised cyclopentane unit in the antitumoural antibiotic viridenomycin. Tetrahedron Lett. 2005, 46, 1913–1915. [Google Scholar]
- Mulholland, N.P.; Pattenden, G. A route to the fully substituted cyclopentane unit of viridenomycin using a tandem radical cyclisation–trapping strategy. Tetrahedron Lett. 2005, 46, 937–939. [Google Scholar] [CrossRef]
- Amagai, K.; Kudo, F.; Eguchi, T. Biosynthetic pathway of macrolactam polyketide antibiotic cremimycin. Tetrahedron 2011, 67, 8559–8563. [Google Scholar] [CrossRef]
- Amagai, K.; Takaku, R.; Kudo, F.; Eguchi, T. A unique amino transfer mechanism for constructing the β-amino fatty acid starter unit in the biosynthesis of the macrolactam antibiotic cremimycin. ChemBioChem 2013, 14, 1998–2006. [Google Scholar] [CrossRef]
- Kawasaki, D.; Chisuga, T.; Miyanaga, A.; Kudo, F.; Eguchi, T. Structural Analysis of the Glycine Oxidase Homolog CmiS2 Reveals a Unique Substrate Recognition Mechanism for Formation of a β-Amino Acid Starter Unit in Cremimycin Biosynthesis. Biochemistry 2019, 58, 2706–2709. [Google Scholar] [CrossRef]
- Tang, G.L.; Cheng, Y.Q.; Shen, B. Chain initiation in the leinamycin-producing hybrid nonribosomal peptide/polyketide synthetase from Streptomyces atroolivaceus S-140. Discrete, monofunctional adenylation enzyme and peptidyl carrier protein that directly load D-alanine. J. Biol. Chem 2007, 282, 20273–20282. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, D.; Sohng, J.K.; Pandey, R.P. Engineering actinomycetes for biosynthesis of macrolactone polyketides. Microb. Cell Factories 2019, 18, 137. [Google Scholar] [CrossRef]
- Chen, H.; Bian, Z.; Ravichandran, V.; Li, R.; Zhang, Y. Biosynthesis of polyketides by trans-AT polyketide synthases in Burkholderiales. Crit. Rev. Microbiol. 2019, 45, 162–181. [Google Scholar] [CrossRef] [PubMed]
- Miyanaga, A. Michael additions in polyketide biosynthesis. Nat. Prod. Rep. 2019, 36, 531–547. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, D.; Miyanaga, A.; Chisuga, T.; Kudo, F.; Eguchi, T. Functional and Structural Analyses of the Split-DH Domain in the Biosynthesis of Macrolactam Polyketide Cremimycin. Biochemistry 2019, 58, 4799–4803. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, X.; Chen, Y.; He, Z.; Yu, P.; Liang, Y. Dynamical Trajectory Study of the Transannular [6+4] and Ambimodal Cycloaddition in the Biosynthesis of Heronamides. J. Org. Chem. 2020, 85, 9440–9445. [Google Scholar] [CrossRef]
- Yu, P.; Patel, A.; Houk, K.N. Transannular [6 + 4] and Ambimodal Cycloaddition in the Biosynthesis of Heronamide A. J. Am. Chem. Soc. 2015, 137, 13518–13523. [Google Scholar] [CrossRef]
- Booth, T.J.; Alt, S.; Capon, R.J.; Wilkinson, B. Synchronous intramolecular cycloadditions of the polyene macrolactam polyketide heronamide C. Chem. Commun. 2016, 52, 6383–6386. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, H.; Liu, Y.; Jiao, Y.; Li, S.; Shen, Y.; Du, L. Biosynthesis of the Polycyclic System in the Antifungal HSAF and Analogues from Lysobacter enzymogenes. Angew. Chem. Int. Ed. Engl. 2018, 57, 6221–6225. [Google Scholar] [CrossRef]
- Klas, K.; Tsukamoto, S.; Sherman, D.H.; Williams, R.M. Natural Diels-Alderases: Elusive and Irresistable. J. Org. Chem. 2015, 80, 11672–11685. [Google Scholar] [CrossRef] [Green Version]
- Yeo, W.L.; Heng, E.; Tan, L.L.; Lim, Y.W.; Ching, K.C.; Tsai, D.J.; Jhang, Y.W.; Lauderdale, T.L.; Shia, K.S.; Zhao, H.; et al. Biosynthetic engineering of the antifungal, anti-MRSA auroramycin. Microb. Cell Fact. 2020, 19, 3. [Google Scholar] [CrossRef] [Green Version]
- Elshahawi, S.; Shaaban, K.A.; Kharel, M.K.; Thorson, J.S. A comprehensive review of glycosylated bacterial natural products. Chem. Soc. Rev. 2015, 44, 7591–7697. [Google Scholar]
- Genilloud, O. Actinomycetes: Still a source of novel antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.X.; Yun, B.S.; Ma, M.; Basu, H.S.; Church, D.R.; Ingenhorst, G.; Huang, Y.; Yang, D.; Lohman, J.R.; Tang, G.L.; et al. Leinamycin E1 acting as an anticancer prodrug activated by reactive oxygen species. Proc. Natl. Acad. Sci. USA 2015, 112, 8278–8283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imani Nejad, M.; Yang, D.; Shen, B.; Gates, K.S. Oxidative activation of leinamycin E1 triggers alkylation of guanine residues in double-stranded DNA. Chem. Commun. 2018, 54, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Alfatah, M.; Kong, K.W.; Hoon, S.; Yeo, W.L.; Ching, K.C.; Jie Hui Goh, C.; Zhang, M.M.; Lim, Y.H.; Wong, F.T.; et al. Chemogenomic profiling in yeast reveals antifungal mode-of-action of polyene macrolactam auroramycin. PLoS ONE 2019, 14, e0218189. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Sugiyama, R.; Nishimura, S.; Ishikawa, N.; Arai, M.A.; Ishibashi, M.; Kakeya, H. Stereochemical Assignment and Biological Evaluation of BE-14106 Unveils the Importance of One Acetate Unit for the Antifungal Activity of Polyene Macrolactams. J. Nat. Prod. 2016, 79, 1877–1880. [Google Scholar] [CrossRef]
- Kanoh, N.; Itoh, S.; Fujita, K.; Sakanishi, K.; Sugiyama, R.; Terajima, Y.; Iwabuchi, Y.; Nishimura, S.; Kakeya, H. Asymmetric Total Synthesis of Heronamides A-C: Stereochemical Confirmation and Impact of Long-Range Stereochemical Communication on the Biological Activity. Chemistry 2016, 22, 8586–8595. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Z.; Li, Y.; Lu, C.; Wang, H.; Shen, Y.; Du, L. HSAF-induced antifungal effects in Candida albicans through ROS-mediated apoptosis. RSC Adv. 2016, 6, 30895–30904. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.S.; Zarins-Tutt, J.S.; Agbo, M.; Gao, H.; Diego-Taboada, A.; Gan, M.L.; Hamed, R.B.; Abraham, E.R.; Mackenzie, G.; Evans, P.A.; et al. A natural solution to photoprotection and isolation of the potent polyene antibiotic, marinomycin A. Chem. Sci. 2019, 10, 7549–7553. [Google Scholar] [CrossRef] [Green Version]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Jiang, H.; Liu, X.-W.; Zhou, J.; Wu, B. Polyene Macrolactams from Marine and Terrestrial Sources: Structure, Production Strategies, Biosynthesis and Bioactivities. Mar. Drugs 2022, 20, 360. https://doi.org/10.3390/md20060360
Zhao W, Jiang H, Liu X-W, Zhou J, Wu B. Polyene Macrolactams from Marine and Terrestrial Sources: Structure, Production Strategies, Biosynthesis and Bioactivities. Marine Drugs. 2022; 20(6):360. https://doi.org/10.3390/md20060360
Chicago/Turabian StyleZhao, Wei, Hong Jiang, Xiao-Wan Liu, Jian Zhou, and Bin Wu. 2022. "Polyene Macrolactams from Marine and Terrestrial Sources: Structure, Production Strategies, Biosynthesis and Bioactivities" Marine Drugs 20, no. 6: 360. https://doi.org/10.3390/md20060360