
Expression Plasticity of Peroxisomal Acyl-Coenzyme A Oxidase Genes Implies Their Involvement in Redox Regulation in Scallops Exposed to PST-Producing Alexandrium

Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of ACOXs in Scallop Genome
2.2. Conserved Structures of ACOX Genes in C. farreri and P. yessoensis
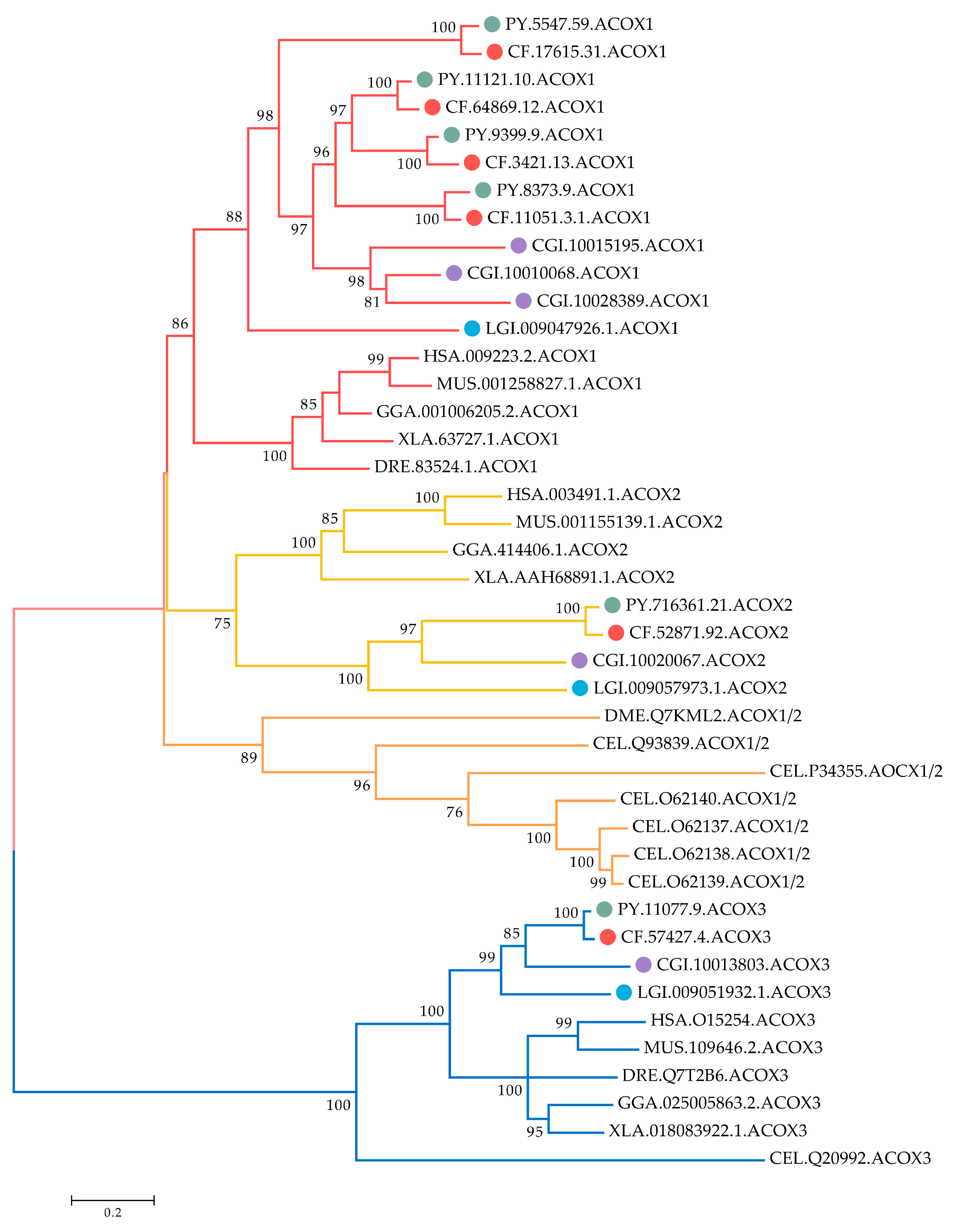
2.3. Phylogenetic Relationship of ACOXs between Bivalves and Other Organisms
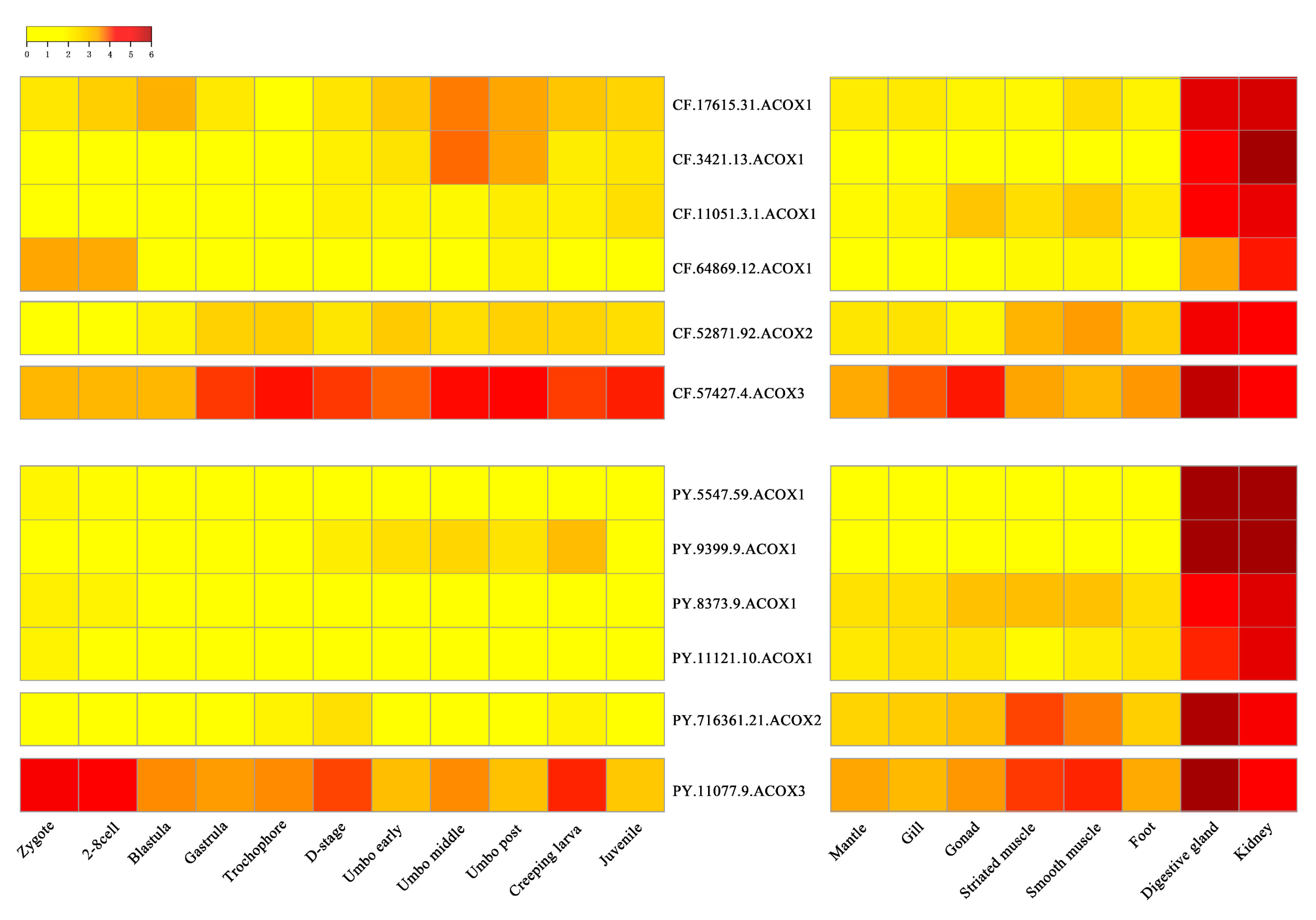
2.4. Expression Profiles of Scallops ACOXs in Developmental Stages and Adult Organs/Tissues
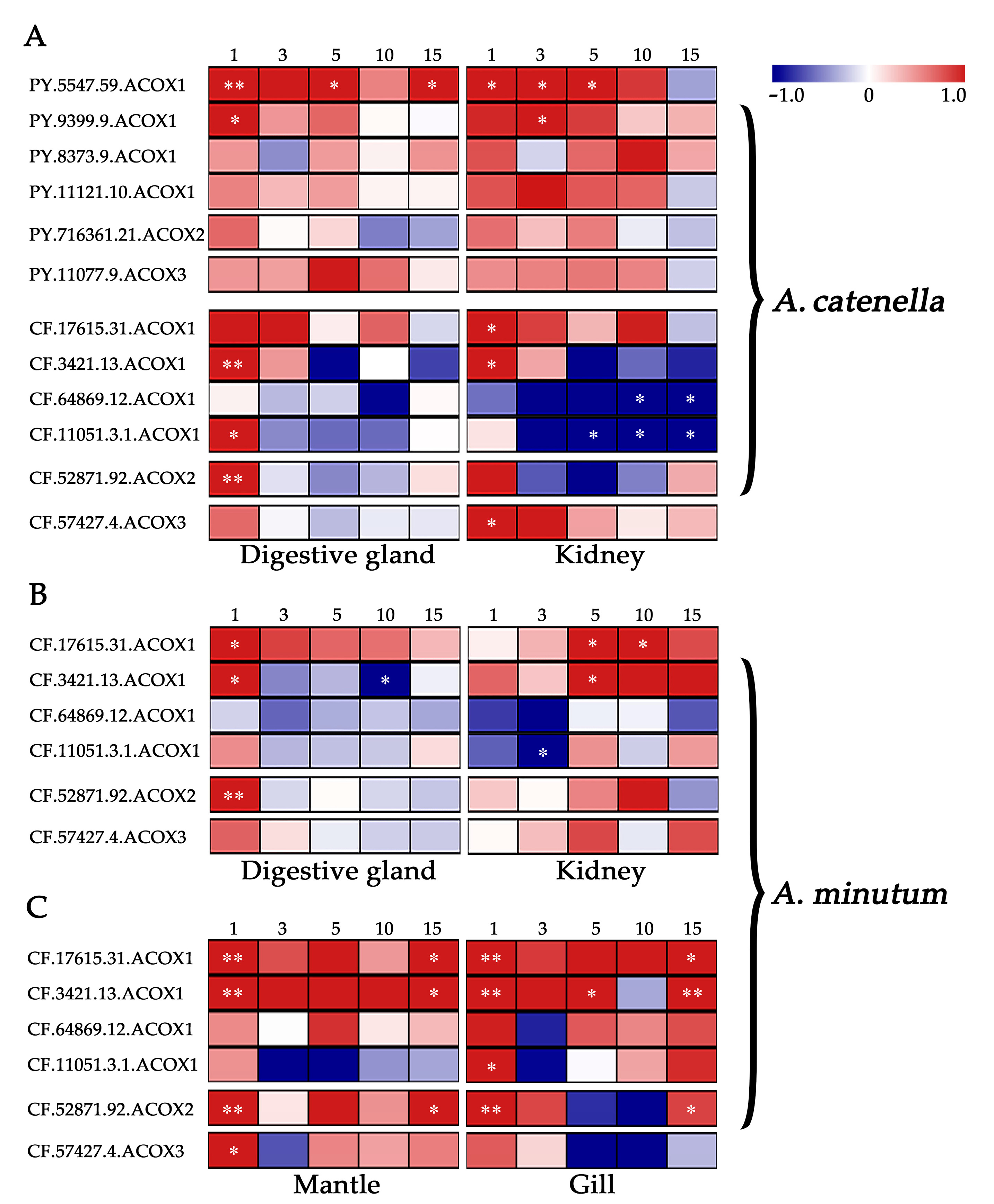
2.5. Expression Regulation of Scallops ACOXs after Toxic Dinoflagellates Exposure
3. Materials and Methods
3.1. Identification and Sequence Analysis of ACOX Genes in C. farreri and P. yessoensis
3.2. Multiple Alignment and Phylogenetic Analysis
3.3. Expression Analysis of ACOX Genes during Developmental Stages and in Adult Organs/Tissues
3.4. Expression Analysis of ACOX Genes Exposed to Toxic Dinoflagellates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, G.; Li, L.; Meng, J.; Qi, H.; Qu, T.; Xu, F.; Zhang, L. Molecular basis for adaptation of oysters to stressful marine intertidal environments. Annu. Rev. Anim. Biosci. 2016, 4, 357–381. [Google Scholar] [CrossRef] [PubMed]
- Repnik, U.; Turk, B. Lysosomal-mitochondrial cross-talk during cell death. Mitochondrion 2010, 10, 662–669. [Google Scholar] [CrossRef]
- Kurz, T.; Eaton, J.W.; Brunk, U.T. Redox activity within the lysosomal compartment: Implications for aging and apoptosis. Antioxid. Redox Signal. 2010, 13, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Nordgren, M.; Fransen, M. Peroxisomal metabolism and oxidative stress. Biochimie 2014, 98, 56–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, J.K.; Hashimoto, T. Peroxisomal beta-oxidation and peroxisome proliferator-activated receptor alpha: An adaptive metabolic system. Annu. Rev. Nutr. 2001, 21, 193–230. [Google Scholar] [CrossRef]
- Camões, F.; Islinger, M.; Guimarães, S.C.; Kilaru, S.; Schuster, M.; Godinho, L.F.; Steinberg, G.; Schrader, M. New insights into the peroxisomal protein inventory: Acyl-CoA oxidases and -dehydrogenases are an ancient feature of peroxisomes. Biochim. Biophys. Acta 2015, 1853, 111–125. [Google Scholar] [CrossRef] [Green Version]
- Van Veldhoven, P.P. Biochemistry and genetics of inherited disorders of peroxisomal fatty acid metabolism. J. Lipid Res. 2010, 51, 2863–2895. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Zhang, Y.; Chai, L.; Wang, H. Histological changes, lipid metabolism and oxidative stress in the liver of Bufo gargarizans exposed to cadmium concentrations. Chemosphere 2017, 179, 337–346. [Google Scholar] [CrossRef]
- Mi, R.; Sun, Y.; Li, J.; Ma, S.; Wen, Z.; Li, X.; Meng, N.; Li, Y.; Du, X.; Li, S. Immune-related proteins detected through iTRAQ-based proteomics analysis of intestines from Apostichopus japonicus in response to tussah immunoreactive substances. Fish Shellfish Immunol. 2018, 74, 436–443. [Google Scholar] [CrossRef]
- Gu, Z.; Jia, R.; He, Q.; Cao, L.; Du, J.; Feng, W.; Jeney, G.; Xu, P.; Yin, G. Alteration of lipid metabolism, autophagy, apoptosis and immune response in the liver of common carp (Cyprinus carpio) after long-term exposure to bisphenol A. Ecotoxicol. Environ. Saf. 2021, 211, 111923. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, S.; Zhang, H.; Lin, S. Omics study of harmful algal blooms in China: Current status, challenges, and future perspectives. Harmful Algae 2021, 107, 102079. [Google Scholar] [CrossRef] [PubMed]
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bricelj, V.M.; Connell, L.; Konoki, K.; Macquarrie, S.P.; Scheuer, T.; Catterall, W.A.; Trainer, V.L. Sodium channel mutation leading to saxitoxin resistance in clams increases risk of PSP. Nature 2005, 434, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef]
- Li, Y.; Sun, X.; Hu, X.; Xun, X.; Zhang, J.; Guo, X.; Jiao, W.; Zhang, L.; Liu, W.; Wang, J.; et al. Scallop genome reveals molecular adaptations to semi-sessile life and neurotoxins. Nat. Commun. 2017, 8, 1721. [Google Scholar] [CrossRef]
- Fabioux, C.; Sulistiyani, Y.; Haberkorn, H.; Hégaret, H.; Amzil, Z.; Soudant, P. Exposure to toxic Alexandrium minutum activates the detoxifying and antioxidant systems in gills of the oyster Crassostrea gigas. Harmful Algae 2015, 48, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Ma, F.; Fan, H.; Li, A. Effects of feeding Alexandrium tamarense, a paralytic shellfish toxin producer, on antioxidant enzymes in scallops (Patinopecten yessoensis) and mussels (Mytilus galloprovincialis). Aquaculture 2013, 396, 76–81. [Google Scholar] [CrossRef]
- Cao, R.; Wang, D.; Wei, Q.; Wang, Q.; Yang, D.; Liu, H.; Dong, Z.; Zhang, X.; Zhang, Q.; Zhao, J. Integrative biomarker assessment of the influence of saxitoxin on marine bivalves: A comparative study of the two bivalve species oysters, Crassostrea gigas, and scallops, Chlamys farreri. Front. Physiol. 2018, 9, 1173. [Google Scholar] [CrossRef] [Green Version]
- Lian, S.; Zhao, L.; Xun, X.; Lou, J.; Li, M.; Li, X.; Wang, S.; Zhang, L.; Hu, X.; Bao, Z. Genome-wide identification and characterization of SODs in Zhikong scallop reveals gene expansion and regulation divergence after toxic dinoflagellate exposure. Mar. Drugs 2019, 17, 700. [Google Scholar] [CrossRef] [Green Version]
- Hlaing, S.M.M.; Lou, J.; Cheng, J.; Xun, X.; Li, M.; Lu, W.; Hu, X.; Bao, Z. Tissue-biased and species-specific regulation of glutathione peroxidase (GPx) genes in scallops exposed to toxic dinoflagellates. Toxins 2020, 13, 21. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.; Cheng, J.; Xun, X.; Li, X.; Li, M.; Zhang, X.; Li, T.; Bao, Z.; Hu, X. Glutathione S-transferase genes in scallops and their diverse expression patterns after exposure to PST-producing dinoflagellates. Mar. Life Sci. Technol. 2020, 2, 252–261. [Google Scholar] [CrossRef]
- Tan, K.S.; Ransangan, J. Factors influencing the toxicity, detoxification and biotransformation of paralytic shellfish toxins. Rev. Environ. Contam. Toxicol. 2015, 235, 1–25. [Google Scholar] [PubMed]
- Wang, S.; Zhang, J.; Jiao, W.; Li, J.; Xun, X.; Sun, Y.; Guo, X.; Huan, P.; Dong, B.; Zhang, L.; et al. Scallop genome provides insights into evolution of bilaterian karyotype and development. Nat. Ecol. Evol. 2017, 1, 120. [Google Scholar] [CrossRef]
- Kim, S.; Kim, K.J. Structural insight into the substrate specificity of acyl-CoA oxidase1 from Yarrowia lipolytica for short-chain dicarboxylyl-CoAs. Biochem. Biophys. Res. Commun. 2018, 495, 1628–1634. [Google Scholar] [CrossRef]
- Tokuoka, K.; Nakajima, Y.; Hirotsu, K.; Miyahara, I.; Nishina, Y.; Shiga, K.; Tamaoki, H.; Setoyama, C.; Tojo, H.; Miura, R. Three-dimensional structure of rat-liver acyl-CoA oxidase in complex with a fatty acid: Insights into substrate-recognition and reactivity toward molecular oxygen. J. Biochem. 2006, 139, 789–795. [Google Scholar] [CrossRef]
- Brocard, C.; Hartig, A. Peroxisome targeting signal 1: Is it really a simple tripeptide? Biochim. Biophys. Acta 2006, 1763, 1565–1573. [Google Scholar] [CrossRef] [Green Version]
- Lespinet, O.; Wolf, Y.I.; Koonin, E.V.; Aravind, L. The role of lineage-specific gene family expansion in the evolution of eukaryotes. Genome Res. 2002, 12, 1048–1059. [Google Scholar] [CrossRef] [Green Version]
- He, A.Y.; Liu, C.Z.; Chen, L.Q.; Ning, L.J.; Zhang, M.L.; Li, E.C.; Du, Z.Y. Identification, characterization and nutritional regulation of two isoforms of acyl-coenzyme A oxidase 1 gene in Nile tilapia (Oreochromis niloticus). Gene 2014, 545, 30–35. [Google Scholar] [CrossRef]
- Chen, Y.; Yan, T.; Tan, Z.J.; Wang, L.P.; Zhou, M.J. Toxicity of dinoflagellate Alexandrium species. Oceanol. Limnol. Sinica 2007, 38, 55–56. [Google Scholar]
- Hu, B.; Li, M.; Yu, X.; Xun, X.; Lu, W.; Li, X.; Li, Y.; Lou, J.; Wang, S.; Zhang, L.; et al. Diverse expression regulation of Hsp70 genes in scallops after exposure to toxic Alexandrium dinoflagellates. Chemosphere 2019, 234, 62–69. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira David, J.A.; Salaroli, R.B.; Fontanetti, C.S. Fine structure of Mytella falcata (Bivalvia) gill filaments. Micron 2008, 39, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Zhang, W.; Zhang, X.; Tian, Y.; Wang, X.; Hao, Z.; Chang, Y. Transcriptional changes in the Japanese scallop (Mizuhopecten yessoensis) shellinfested by Polydora provide insights into the molecular mechanism of shell formation and immunomodulation. Sci. Rep. 2018, 8, 17664. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Kong, F.Z.; Xun, X.G.; Dai, L.; Geng, H.X.; Hu, X.L.; Yu, R.C.; Bao, Z.M.; Zhou, M.J. Biokinetics and biotransformation of paralytic shellfish toxins in different tissues of Yesso scallops, Patinopecten yessoensis. Chemosphere 2020, 261, 128063. [Google Scholar] [CrossRef] [PubMed]
- Simakov, O.; Marletaz, F.; Cho, S.J.; Edsinger-Gonzales, E.; Havlak, P.; Hellsten, U.; Kuo, D.H.; Larsson, T.; Lv, J.; Arendt, D.; et al. Insights into bilaterian evolution from three spiralian genomes. Nature 2013, 493, 526–531. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Navarro, J.M.; Muñoz, M.G.; Contreras, A.M. Temperature as a factor regulating growth and toxin content in the dinoflagellate Alexandrium catenella. Harmful Algae 2006, 5, 762–769. [Google Scholar] [CrossRef]
- Hwang, D.F.; Lu, Y.H. Influence of environmental and nutritional factors on growth, toxicity, and toxin profile of dinoflagellate Alexandrium minutum. Toxicon 2000, 38, 1491–1503. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.; Xun, X.; Li, M.; Lou, J.; Zhang, Y.; Shi, J.; Hu, J.; Bao, Z.; Hu, X. Toxin- and species-dependent regulation of ATP-binding cassette (ABC) transporters in scallops after exposure to paralytic shellfish toxin-producing dinoflagellates. Aquat. Toxicol. 2021, 230, 105697. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Bao, Z.; Hu, J.; Shao, M.; Zhang, L.; Bi, K.; Zhan, A.; Huang, X. Cloning and characterization of tryptophan 2,3-dioxygenase gene of Zhikong scallop Chlamys farreri (Jones and Preston 1904). Aquac. Res. 2006, 37, 1187–1194. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Chromosome No. | Scaffold ID | CDS (bp) | Amino Acid (aa) | ACOX Pfam Position | Isoelectric Point (pI) | Molecular Weight (kDa) |
|---|---|---|---|---|---|---|---|
| CF.17615.31.ACOX1 | Chr10 | 17615 | 1989 | 662 | 477–658 | 8.42 | 74.92 |
| CF.3421.13.ACOX1 | Chr2 | 3421 | 2013 | 670 | 479–660 | 8.89 | 75.31 |
| CF.11051.3.1.ACOX1 | Chr10 | 11051 | 2022 | 673 | 481–662 | 8.75 | 75.11 |
| CF.64869.12.ACOX1 | Chr10 | 64869 | 1896 | 631 | 437–619 | 8.40 | 70.84 |
| CF.52871.92.ACOX2 | Chr7 | 52871 | 2019 | 672 | 484–665 | 7.82 | 75.26 |
| CF.57427.4.ACOX3 | Chr9 | 57427 | 2187 | 728 | 536–722 | 5.74 | 81.47 |
| PY.5547.59.ACOX1 | Chr18 | 5547 | 1989 | 662 | 477–658 | 8.86 | 74.70 |
| PY.9399.9.ACOX1 | Chr18 | 9399 | 2013 | 670 | 479–660 | 8.64 | 74.51 |
| PY.8373.9.ACOX1 | Chr18 | 8373 | 2022 | 673 | 481–662 | 8.65 | 75.36 |
| PY.11121.10.ACOX1 | Chr18 | 11121 | 2010 | 669 | 473–655 | 7.26 | 75.20 |
| PY.716361.21.ACOX2 | Chr6 | 716361 | 2019 | 672 | 484–665 | 7.55 | 75.71 |
| PY.11077.9.ACOX3 | Chr10 | 11077 | 2187 | 728 | 536–722 | 5.81 | 81.36 |
| Species | ACOX1 | ACOX2 | ACOX3 | Total |
|---|---|---|---|---|
| Homo sapiens | 1 | 1 | 1 | 3 |
| Mus musculus | 1 | 1 | 1 | 3 |
| Gallus gallus | 1 | 1 | 1 | 3 |
| Xenopus laevis | 1 | 1 | 1 | 3 |
| Danio rerio | 1 | 0 | 1 | 2 |
| Lottia gigantean | 1 | 1 | 1 | 3 |
| Crassostrea gigas | 3 | 1 | 1 | 5 |
| Patinopecten yessoensis | 4 | 1 | 1 | 6 |
| Chlamys farreri | 4 | 1 | 1 | 6 |
| Drosophila melanogaster | 1 (ACOX1/2) | 0 | 1 | |
| Caenorhabditis elegans | 6 (ACOX1/2) | 1 | 7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Wang, Y.; Tang, Z.; Wang, H.; Hu, J.; Bao, Z.; Hu, X. Expression Plasticity of Peroxisomal Acyl-Coenzyme A Oxidase Genes Implies Their Involvement in Redox Regulation in Scallops Exposed to PST-Producing Alexandrium. Mar. Drugs 2022, 20, 472. https://doi.org/10.3390/md20080472
Li M, Wang Y, Tang Z, Wang H, Hu J, Bao Z, Hu X. Expression Plasticity of Peroxisomal Acyl-Coenzyme A Oxidase Genes Implies Their Involvement in Redox Regulation in Scallops Exposed to PST-Producing Alexandrium. Marine Drugs. 2022; 20(8):472. https://doi.org/10.3390/md20080472
Chicago/Turabian StyleLi, Moli, Yangrui Wang, Zhihong Tang, Huizhen Wang, Jingjie Hu, Zhenmin Bao, and Xiaoli Hu. 2022. "Expression Plasticity of Peroxisomal Acyl-Coenzyme A Oxidase Genes Implies Their Involvement in Redox Regulation in Scallops Exposed to PST-Producing Alexandrium" Marine Drugs 20, no. 8: 472. https://doi.org/10.3390/md20080472
APA StyleLi, M., Wang, Y., Tang, Z., Wang, H., Hu, J., Bao, Z., & Hu, X. (2022). Expression Plasticity of Peroxisomal Acyl-Coenzyme A Oxidase Genes Implies Their Involvement in Redox Regulation in Scallops Exposed to PST-Producing Alexandrium. Marine Drugs, 20(8), 472. https://doi.org/10.3390/md20080472