
Ribosomal DNA Sequence-Based Taxonomy and Antimicrobial Activity of Prorocentrum spp. (Dinophyceae) from Mauritius Coastal Waters, South-West Indian Ocean



Abstract
:1. Introduction
2. Results
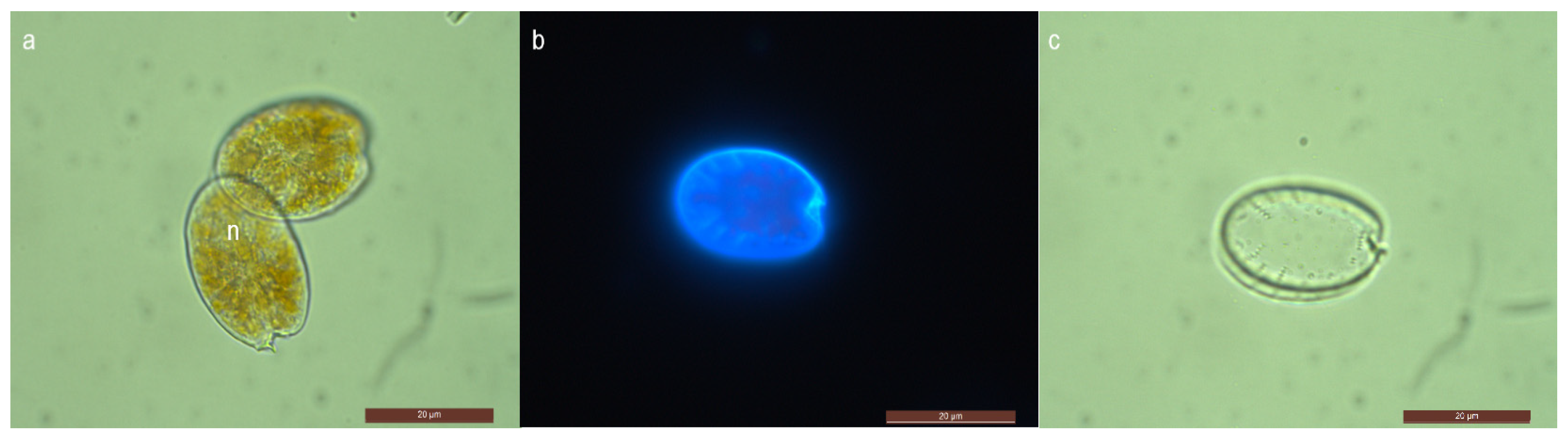
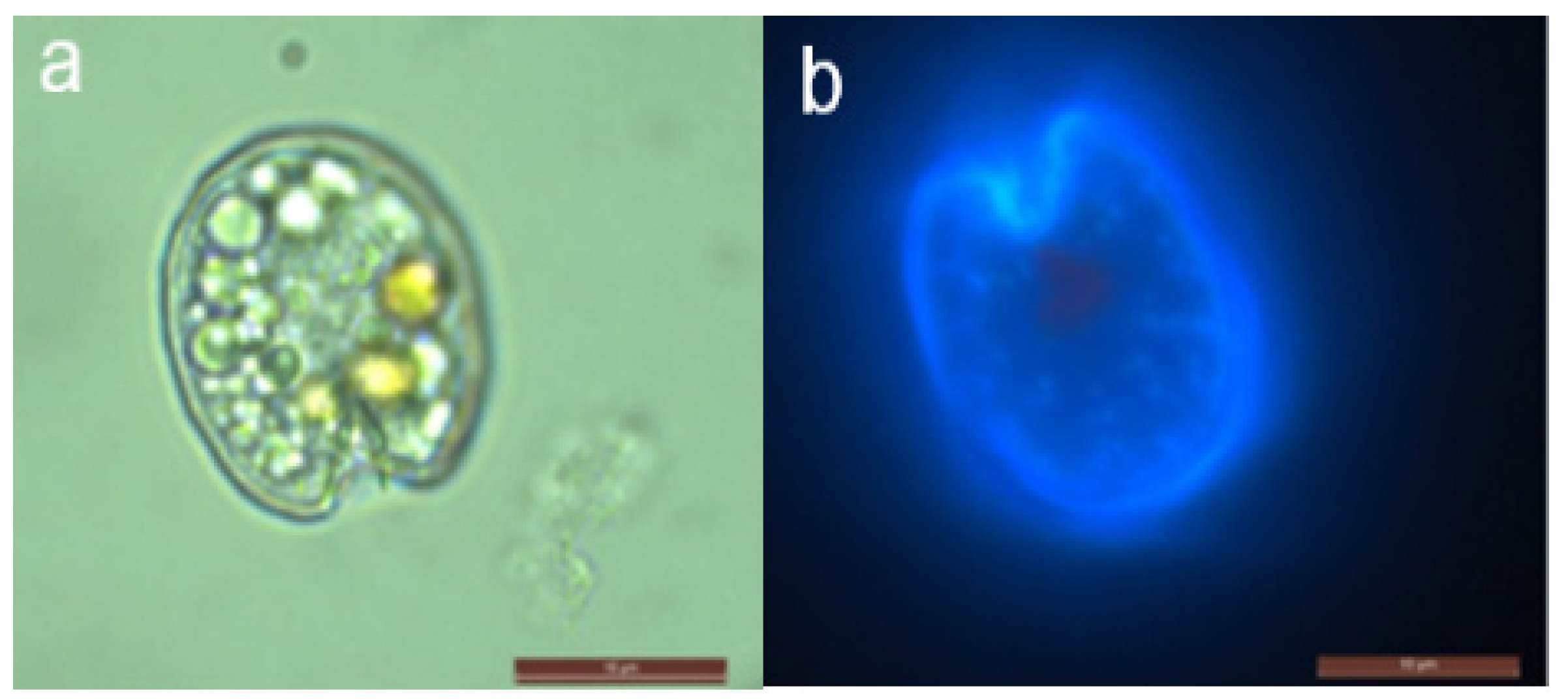
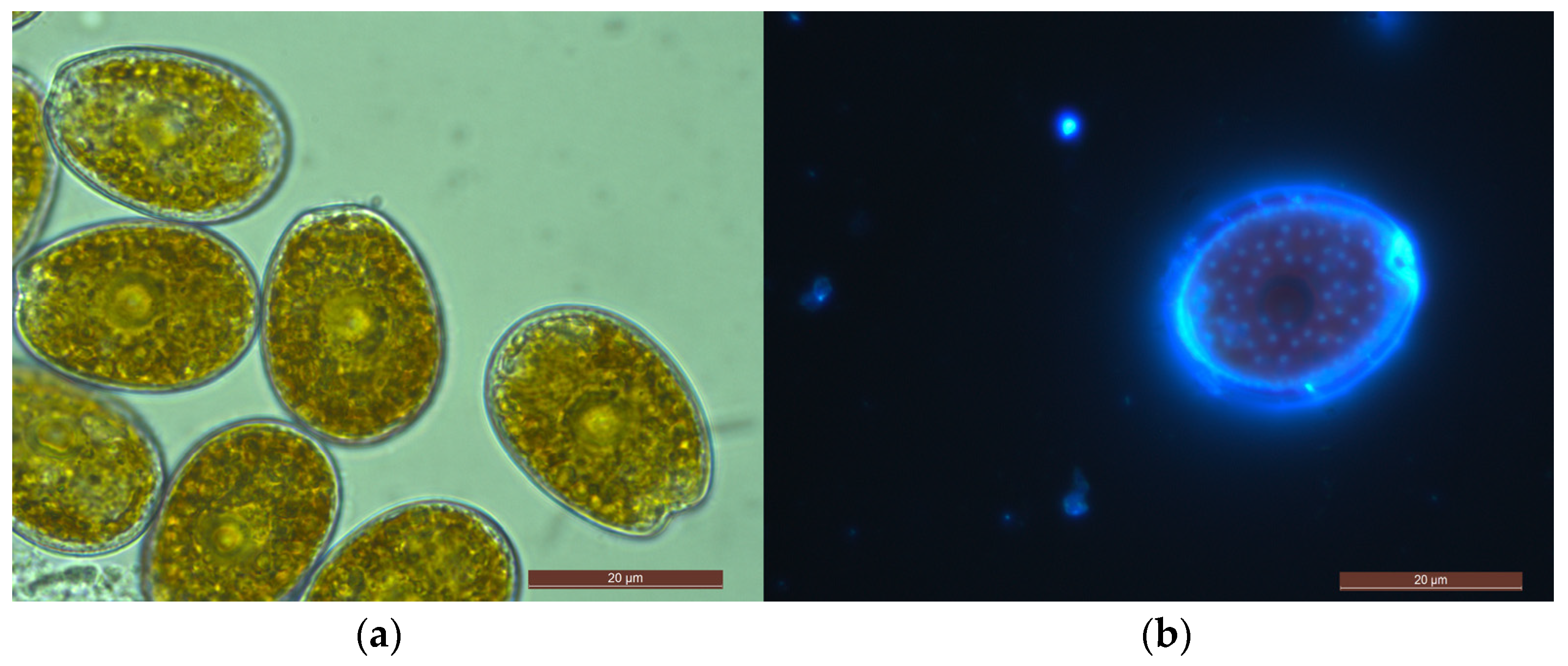
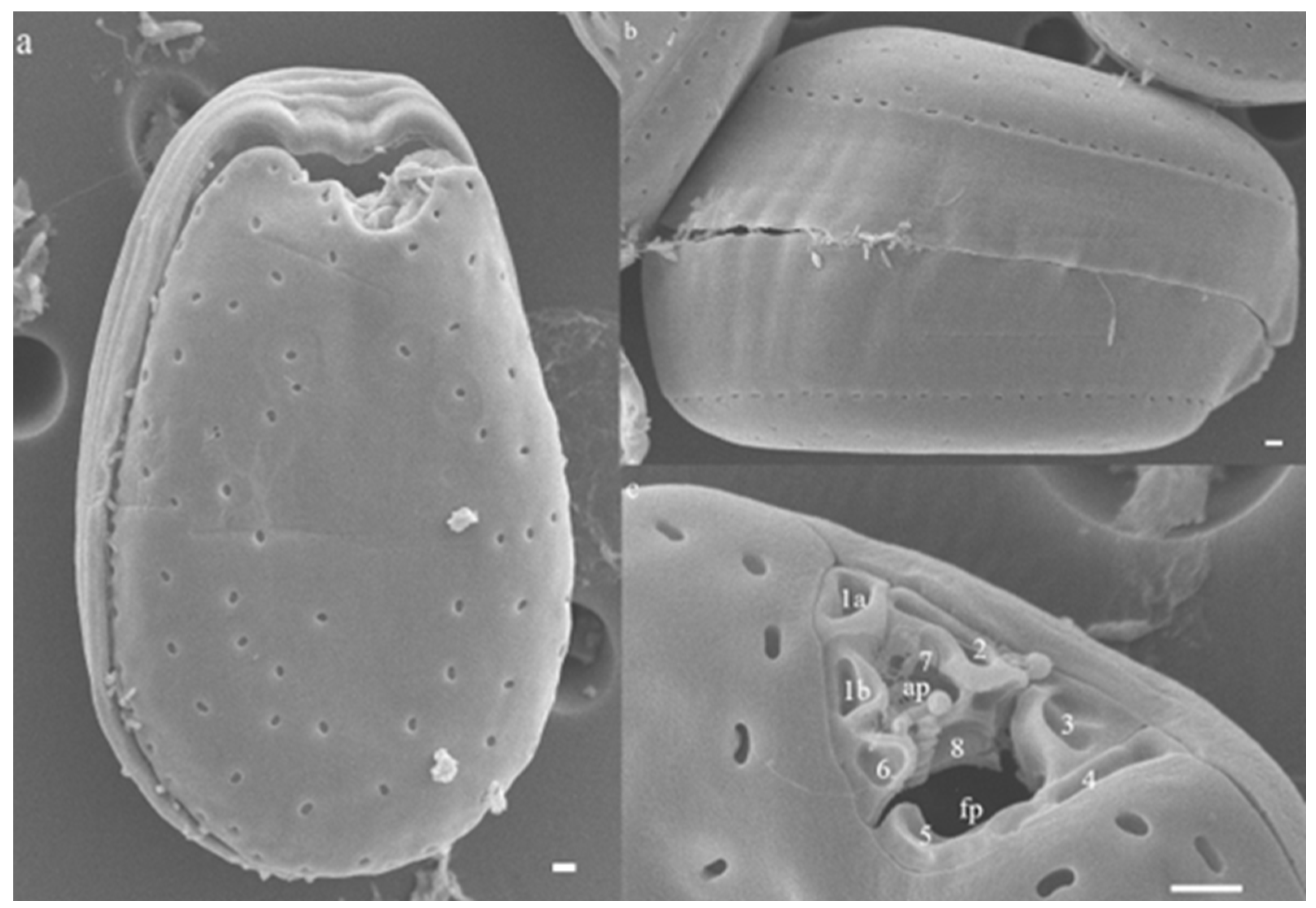
2.1. Morphological Features of Prorocentrum Species Isolated
2.1.1. Prorocentrum rhathymum
2.1.2. Prorocentrum fukuyoi Complex
2.1.3. Prorocentrum lima Complex
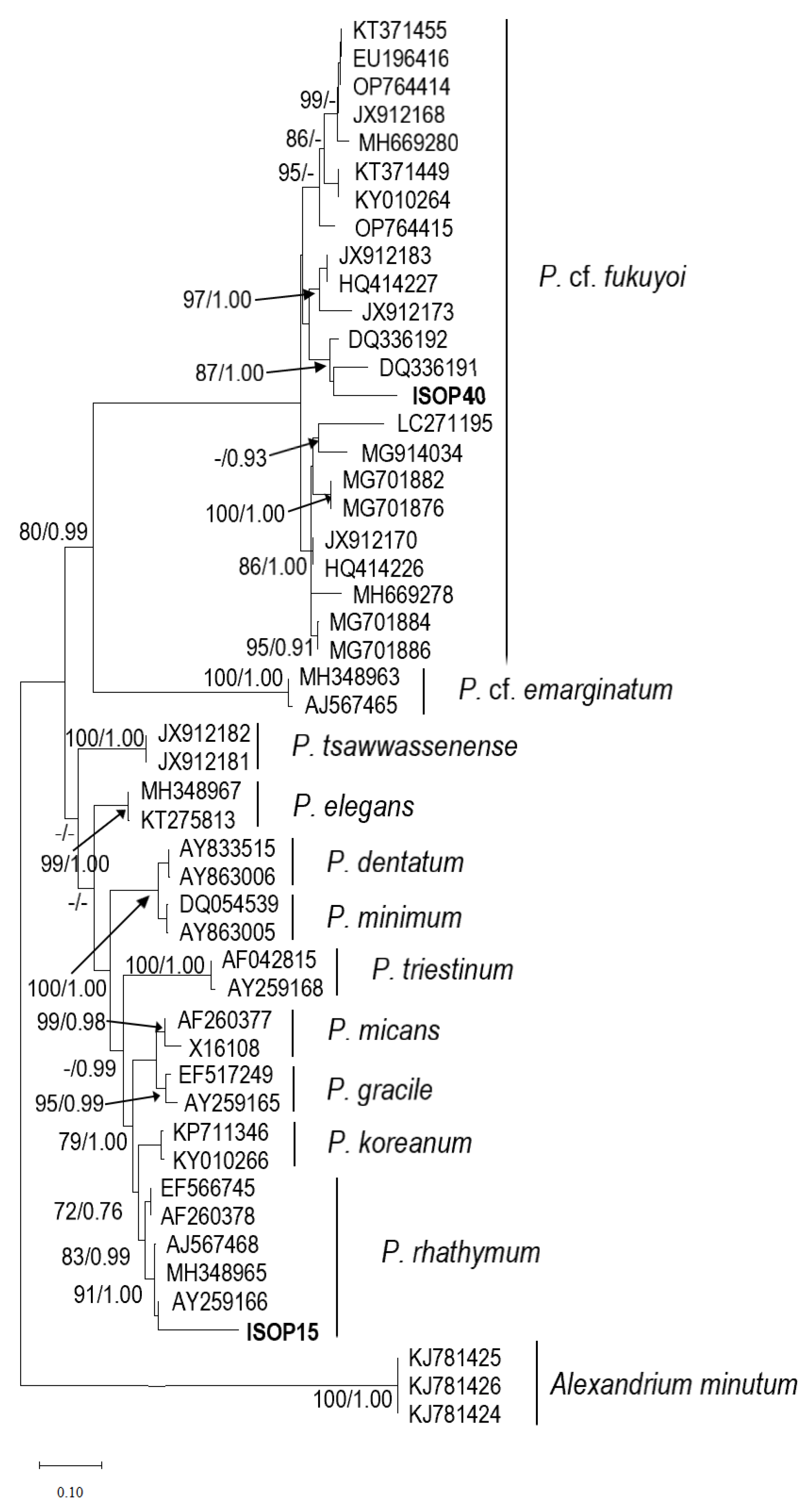
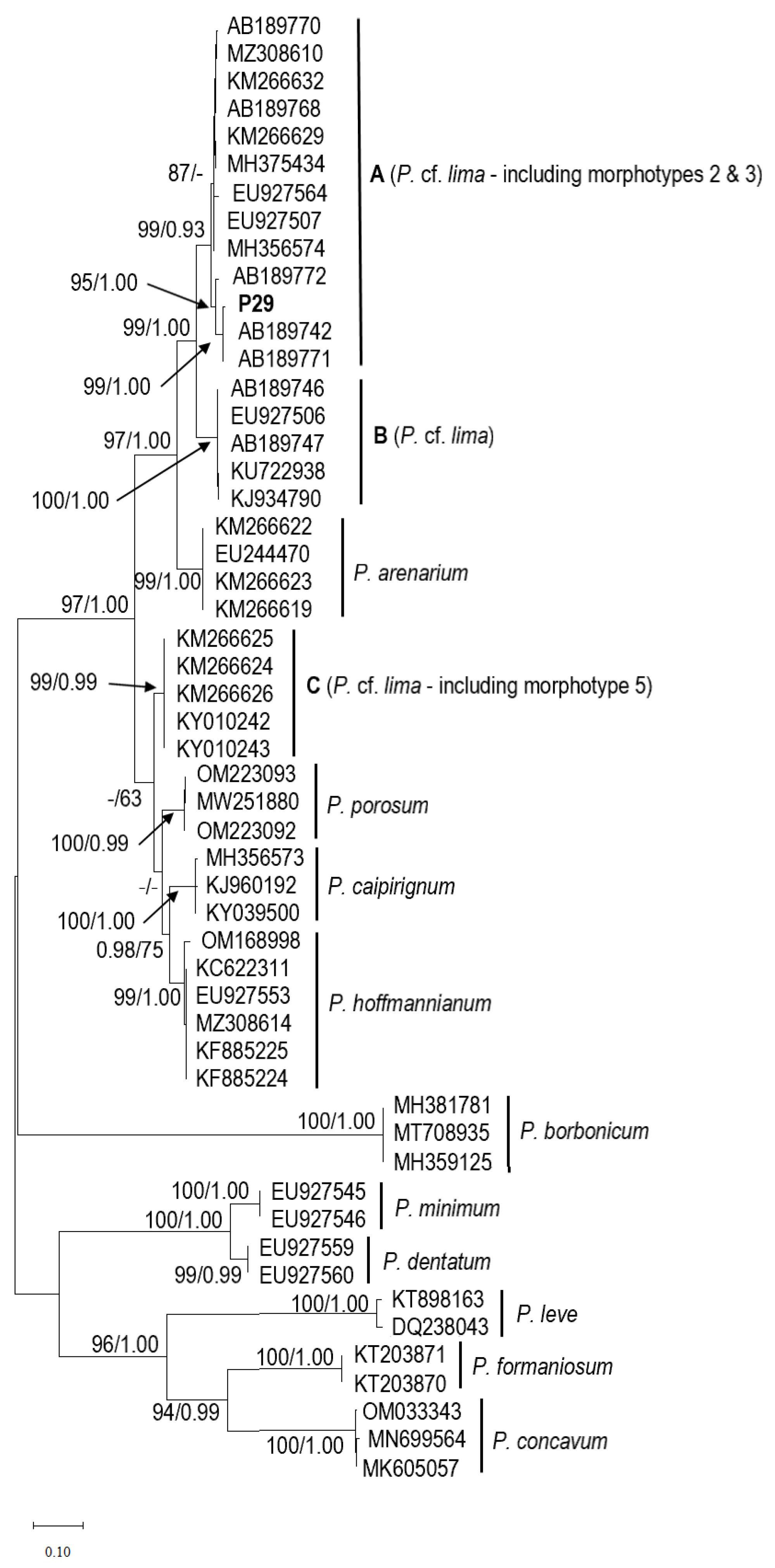
2.2. Phylogenetics of Prorocentrum Species Isolated
2.2.1. Prorocentrum rhathymum
2.2.2. Prorocentrum fukuyoi Complex
2.2.3. Prorocentrum lima Complex
2.3. Antimicrobial Activities of Extracts
3. Discussion
4. Materials and Methods
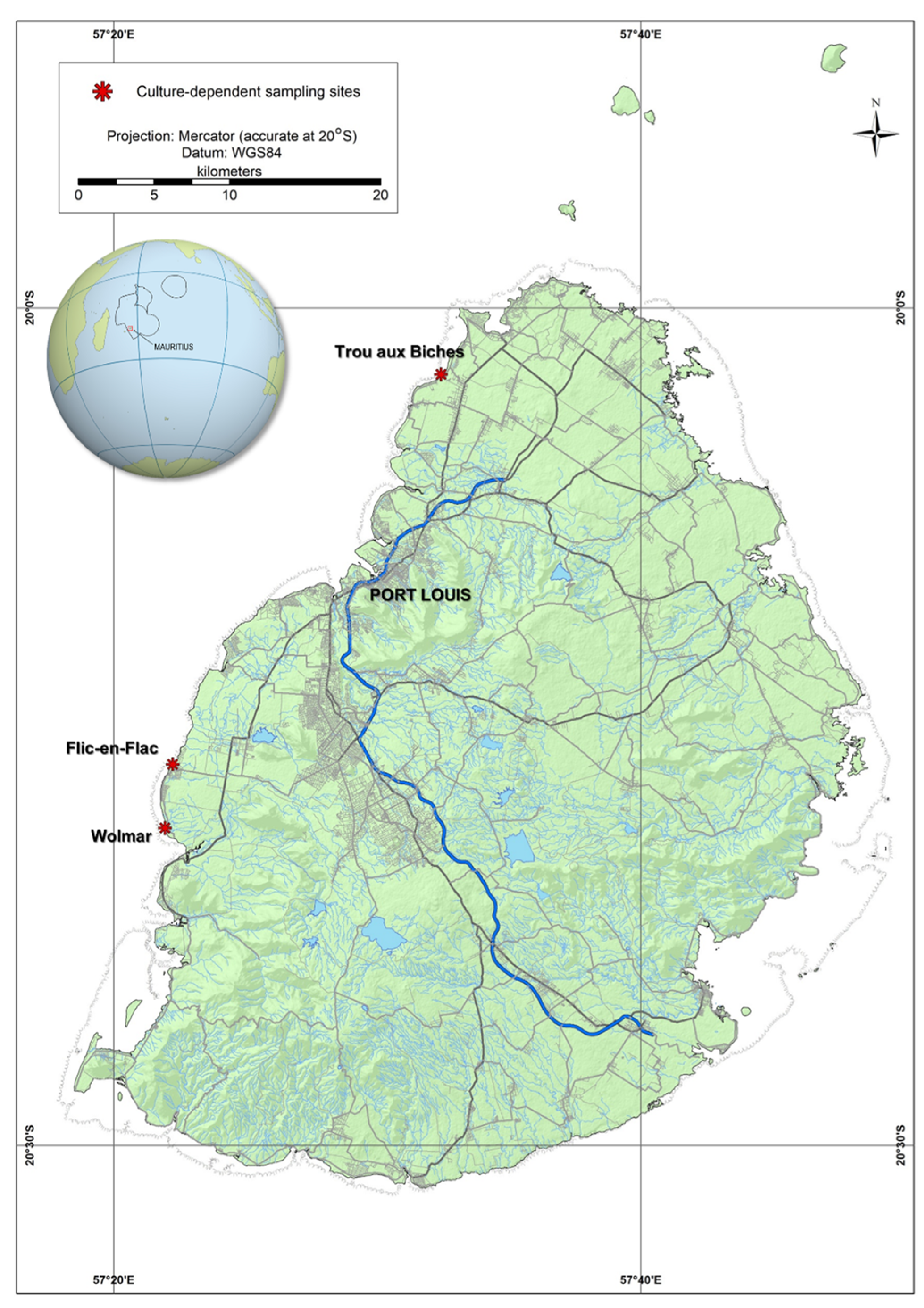
4.1. Study Area and Sample Collections
4.2. Light Microscopy (LM) and Scanning Electron Microscopy (SEM) Observations
4.3. DNA Extraction and PCR Amplification
4.4. Phylogenetic Analyses
4.5. Antimicrobial Potential
4.5.1. Intracellular Extract
4.5.2. Extracellular Proteins and Polysaccharides Extracts
4.5.3. Bacterial Pathogens
4.6. Antimicrobial Assay
4.6.1. Antimicrobial Susceptibility Test
4.6.2. Determination of Minimum Inhibitory Concentration
4.6.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nethravathy, M.U.; Mehar, J.G.; Mudliar, S.N.; Shekh, A.Y. Recent Advances in Microalgal Bioactives for Food, Feed, and Healthcare Products: Commercial Potential, Market Space, and Sustainability. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1882–1897. [Google Scholar]
- Karthikeyan, A.; Joseph, A.; Nair, B.G. Promising bioactive compounds from the marine environment and their potential effects on various diseases. J. Genet. Eng. Biotechnol. 2022, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Berdalet, E.; Bravo, I.; Evans, J.; Fraga, S.; Kibler, S.; Kudela, M.; Larsen, J.; Litaker, W.; Penna, A.; Tester, P.A. Global Ecology and Oceanography of Harmful Algal Blooms, GEOHAB Core Research Project: HABs in Benthic Systems. 2012. Available online: https://www.researchgate.net/publication/235759642_Global_ecology_and_oceanography_of_harmful_algal_blooms_GEOHAB_Core_Research_Project_HABs_in_benthic_systems (accessed on 4 September 2017).
- Hallegraeff, G.; Enevoldsen, H.; Zingone, A. Global harmful algal bloom status reporting. Harmful Algae 2021, 102, 101992. [Google Scholar] [CrossRef] [PubMed]
- Soliño, L.; Costa, P.R. Global impact of ciguatoxins and ciguatera fish poisoning on fish, fisheries and consumers. Environ. Res. 2020, 182, 109111. [Google Scholar] [CrossRef] [PubMed]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estévez, P.; Leao, J.M.; Gago-Martinez, A. Chapter 7—Marine biotoxins as natural contaminants in seafood: European perspective. In Present Knowledge in Food Safety; Knowles, M.E., Anelich, L.E., Boobis, A.R., Popping, B., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 115–127. [Google Scholar]
- Ferrara, L. Dinoflagellates Important Marine Producers of Natural Bio-Compounds with High Biotechnological and Pharmacological Potential. J. Food Chem. Nanotechnol. 2020, 6, 138–149. [Google Scholar] [CrossRef]
- Shenoy, B.D.; Jeewon, R.; Wang, H.; Kaur, A.; Ho, W.H.; Bhat, D.J.; Crous, P.W.; Hyde, K.D. Sequence data reveals phylogenetic affinities of fungal anamorphs Bahusutrabeeja, Diplococcium, Natarajania, Paliphora, Polyschema, Rattania and Spadicoides. Fungal Divers. 2010, 44, 161–169. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Phukhamsakda, C.; Jayawardena, R.S.; Jeewon, R.; Promputtha, I.; Hyde, K.D. Investigating species boundaries in Colletotrichum. Fungal Divers. 2021, 107, 107–127. [Google Scholar] [CrossRef]
- Luo, Z.-L.; Bhat, D.J.; Jeewon, R.; Boonmee, S.; Bao, D.-F.; Zhao, Y.-C.; Chai, H.-M.; Su, H.-Y.; Su, X.-J.; Hyde, K.D. Molecular phylogeny and morphological characterization of asexual fungi (Tubeufiaceae) from freshwater habitats in Yunnan, China. Cryptogam. Mycol. 2017, 38, 27–53. [Google Scholar] [CrossRef]
- Zhang, Y.; Jeewon, R.; Fournier, J.; Hyde, K.D. Multi-gene phylogeny and morphotaxonomy of Amniculicola lignicola: Novel freshwater fungus from France and its relationships to the Pleosporales. Fungal Biol. 2008, 112, 1186–1194. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Salgueiro, F.; Menezes, M.; Oliveira, F.d.A.; Magalhães, V.C.P.; De Paula, J.C.; Morris, S. Prorocentrum lima from the South Atlantic: Morphological, molecular and toxicological characterization. Harmful Algae 2016, 57, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Faust, M.A.; Larsen, J.; Moestrup, J. ICES Identification Leaflets for Plankton. Potentially Toxic Phytoplankton. Genus Prorocentrum (Dinophyceae); International Council for the Exploration of the Sea: Copenhagen, Denmark, 1999; Volume 184, pp. 1–24. [Google Scholar]
- Shah, M.M.R.; Samarakoon, K.W.; Ko, J.-Y.; Lakmal, H.H.C.; Lee, J.-H.; An, S.-J.; Jeon, Y.-J.; Lee, J.-B. Potentiality of benthic dinoflagellate cultures and screening of their bioactivities in Jeju Island, Korea. Afr. J. Biotechnol. 2014, 13, 792–805. [Google Scholar]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- De Vera, C.R.; Díaz Crespín, G.; Hernández Daranas, A.; Montalvão Looga, S.; Lillsunde, K.-E.; Tammela, P.; Perälä, M.; Hongisto, V.; Virtanen, J.; Rischer, H. Marine microalgae: Promising source for new bioactive compounds. Mar. Drugs 2018, 16, 317. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Muñoz, D.; Praptiwi, R.A.; Lawton, L.A.; Edwards, C. High value phycotoxins from the dinoflagellate Prorocentrum. Front. Mar. Sci. 2021, 8, 638739. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2023. Available online: https://www.algaebase.org (accessed on 25 January 2023).
- Hoppenrath, M.; Chomérat, N.; Horiguchi, T.; Schweikert, M.; Nagahama, Y.; Murray, S. Taxonomy and phylogeny of the benthic Prorocentrum species (Dinophyceae)—A proposal and review. Harmful Algae 2013, 27, 1–28. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Cen, J.; Wang, H.; Cui, L.; Dong, Y.; Lu, S. Morphotypes of Prorocentrum lima (Dinophyceae) from Hainan Island, South China Sea: Morphological and molecular characterization. Phycologia 2015, 54, 503–516. [Google Scholar] [CrossRef]
- Nascimento, S.M.; Mendes, M.C.Q.; Menezes, M.; Rodríguez, F.; Alves-de-Souza, C.; Branco, S.; Riobó, P.; Franco, J.; Nunes, J.M.C.; Huk, M.; et al. Morphology and phylogeny of Prorocentrum caipirignum sp. nov. (Dinophyceae), a new tropical toxic benthic dinoflagellate. Harmful Algae 2017, 70, 73–89. [Google Scholar] [CrossRef]
- Verma, A.; Kazandjian, A.; Sarowar, C.; Harwood, D.T.; Murray, J.S.; Pargmann, I.; Hoppenrath, M.; Murray, S.A. Morphology and Phylogenetics of Benthic Prorocentrum Species (Dinophyceae) from Tropical Northwestern Australia. Toxins 2019, 11, 571. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Li, Q.; Liu, H.; Liu, Y.; Huang, L.; Wu, H.; Qiu, J.; Zhang, H.; Lü, S. Taxonomy and toxin profile of harmful benthic Prorocentrum (Dinophyceae) species from the Xisha Islands, South China Sea. J. Oceanol. Limnol. 2022, 40, 1171–1190. [Google Scholar] [CrossRef]
- Hurbungs, M.D.; Jayabalan, N.; Chineah, V. Seasonal Distribution of Potentially Toxic Dinoflagellates in the Lagoon of Trou Aux Biches, Mauritius; Food and Agriculture Research Council: Reduit, Mauritius, 2001; pp. 211–217. [Google Scholar]
- Neermul, P.; Dhunnoo, G.; Emrith, V.; Fakoo, M.; Jeenally, N.; Bhantooa, R.; Elaheebocus, K. Spatial and Seasonal Variations of Harmful Benthic Dinoflagellates in the Coastal Waters of Mauritius. In Proceedings of the Fifth International Conference in Ocean Engineering, Singapore, 10 November 2021; Sundar, V., Sannasiraj, S.A., Sriram, V., Nowbuth, M.D., Eds.; Springer: Singapore, 2021; pp. 269–279. [Google Scholar]
- Hansen, G.; Turquet, J.; Quod, J.P.; Ten-Hage, L.; Lugomela, C.; Kyewalyanga, M.; Hurbungs, M.D.; Wawiye, P.; Ogongo, B.; Tunje, S. Potentially Harmful Microalgae of the Western Indian Ocean (a Guide Based on a Preliminary Survey); UNESCO: Paris, France, 2001; Volume 41, p. 105. [Google Scholar]
- Nagahama, Y.; Murray, S.; Tomaru, A.; Fukuyo, Y. Species boundaries in the toxic dinoflagellate Prorocentrum lima (Dinophyceae, Prorocentrales), based on morphological and phylogenetics characters. J. Phycol. 2011, 47, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Sepúlveda, A.; Medlin, L.K.; Murugan, G.; Sierra-Beltrán, A.P.; Cruz-Villacorta, A.A.; Hernández-Saavedra, N.Y. Are Prorocentrum hoffmannianum and Prorocentrum belizeanum (Dinophyceae, Prorocentrales), the same species? An integration of morphological and molecular data. J. Phycol. 2015, 51, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Loeblich III, A.R.; Sherley, J.L.; Schmidt, R.J. The correct position of flagellar insertion in Prorocentrum and description of Prorocentrum rhathymum sp. nov.(Pyrrhophyta). J. Plankton Res. 1979, 1, 113–120. [Google Scholar] [CrossRef]
- Fukuyo, Y. Taxonomical Study on Benthic Dinoflagellates Collected in Coral Reefs. Bull. Jpn. Soc. Sci. Fish. 1981, 47, 967–978. [Google Scholar] [CrossRef] [Green Version]
- Mohammad-Noor, N.; Daugbjerg, N.; Moestrup, Ø.; Anton, A. Marine epibenthic dinoflagellates from Malaysia—A study of live cultures and preserved samples based on light and scanning electron microscopy. Nord. J. Bot. 2007, 24, 629–690. [Google Scholar] [CrossRef]
- Aligizaki, K.; Nikolaidis, G.; Katikou, P.; Baxevanis, A.D.; Abatzopoulos, T.J. Potentially toxic epiphytic Prorocentrum (Dinophyceae) species in Greek coastal waters. Harmful Algae 2009, 8, 299–311. [Google Scholar] [CrossRef]
- Lim, Z.F.; Luo, Z.; Lee, L.K.; Hii, K.S.; Teng, S.T.; Chan, L.L.; Chomérat, N.; Krock, B.; Gu, H.; Lim, P.T.; et al. Taxonomy and toxicity of Prorocentrum from Perhentian Islands (Malaysia), with a description of a non-toxigenic species Prorocentrum malayense sp. nov. (Dinophyceae). Harmful Algae 2019, 83, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Gómez, F.; Qiu, D.; Lin, S. The Synonymy of the Toxic Dinoflagellates Prorocentrum mexicanum and P. rhathymum and the Description of P. steidingerae sp. nov. (Prorocentrales, Dinophyceae). J. Eukaryot. Microbiol. 2017, 64, 668–677. [Google Scholar] [CrossRef]
- Faust, M.A. Morphologic details of six benthic species of Prorocentrum (Pyrrophyta) from a Mangrove Island, Twin Cays, Belize, including two new species. J. Phycol. 1990, 26, 548–558. [Google Scholar] [CrossRef]
- Chomérat, N.; Bilien, G.; Zentz, F. A taxonomical study of benthic Prorocentrum species (Prorocentrales, Dinophyceae) from Anse Dufour (Martinique Island, eastern Caribbean Sea). Mar. Biodivers. 2019, 49, 1299–1319. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Zhang, H.; Krock, B.; Lu, S.; Yang, W.; Gu, H. Morphology, molecular phylogeny and okadaic acid production of epibenthic Prorocentrum (Dinophyceae) species from the northern South China Sea. Algal Res. 2017, 22, 14–30. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Huang, S.; Krock, B.; Leaw, C.P.; Teng, S.T.; Piumsomboon, A.; Punnarak, P.; Roeroe, K.A.; Wang, N.; Gu, H. Cryptic speciation of benthic Prorocentrum (Dinophyceae) species and their potential as ecological indicators. J. Sea Res. 2022, 190, 102304. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Chézeau, A.; Turquet, J.; Quod, J.P.; Puiseux-Dao, S. Morphological and toxicological variability of Prorocentrum lima clones isolated from four locations in the south-west Indian Ocean. Toxicon 2001, 39, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.S. Morphological variation of dinoflagellate Prorocentrum lima (Ehrenberg) Dodge with geographical difference. J. Environ. Biol. 2004, 25, 51–58. [Google Scholar] [PubMed]
- Nascimento, S.M.; Purdie, D.A.; Morris, S. Morphology, toxin composition and pigment content of Prorocentrum lima strains isolated from a coastal lagoon in southern UK. Toxicon 2005, 45, 633–649. [Google Scholar] [CrossRef]
- Moreira-González, A.R.; Fernandes, L.F.; Uchida, H.; Uesugi, A.; Suzuki, T.; Chomérat, N.; Bilien, G.; Mafra, L.L. Variations in morphology, growth, and toxicity among strains of the Prorocentrum lima species complex isolated from Cuba and Brazil. J. Appl. Phycol. 2019, 31, 519–532. [Google Scholar] [CrossRef]
- Oyeku, O.G.; Mandal, S.K. Taxonomy and okadaic acid production of a strain of Prorocentrum lima (Dinophyceae) isolated from the Bay of Bengal, North Indian Ocean. Toxicon 2021, 196, 32–43. [Google Scholar] [CrossRef]
- Widiarti, R.; Zamani, N.P.; Bengen, D.G.; Madduppa, H. Morphotype diversity of Prorocentrum lima in the western part of Indonesian waters. Biodiversitas J. Biol. Divers. 2021, 22, 607–614. [Google Scholar] [CrossRef]
- Faust, M.A. Morhoplogy of ciguatera-causing Prorocentrum lima (Pyrrophyta) from widely differing sites. J. Phycol. 1991, 27, 642–648. [Google Scholar] [CrossRef]
- Arteaga-Sogamoso, E.; Rodríguez, F.; Amato, A.; Ben-Gigirey, B.; Fraga, S.; Mafra, L.L.; Fernandes, L.F.; de Azevedo Tibiriçá, C.E.J.; Chomérat, N.; Nishimura, T.; et al. Morphology and phylogeny of Prorocentrum porosum sp. nov. (Dinophyceae): A new benthic toxic dinoflagellate from the Atlantic and Pacific Oceans. Harmful Algae 2023, 121, 102356. [Google Scholar] [CrossRef]
- Nishimura, T.; Uchida, H.; Noguchi, R.; Oikawa, H.; Suzuki, T.; Funaki, H.; Ihara, C.; Hagino, K.; Arimitsu, S.; Tanii, Y.; et al. Abundance of the benthic dinoflagellate Prorocentrum and the diversity, distribution, and diarrhetic shellfish toxin production of Prorocentrum lima complex and P. caipirignum in Japan. Harmful Algae 2020, 96, 101687. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.C.; Gião, M.S.; Seabra, R.; Ferreira, A.S.; Tamagnini, P.; Moradas-Ferreira, P.; Malcata, F.X. Evaluation of the antioxidant activity of cell extracts from microalgae. Mar. Drugs 2013, 11, 1256–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarakoon, K.W.; Ko, J.-Y.; Rahman, S.M.; Lee, J.-H.; Kang, M.-C.; Kwon, O.-N.; Lee, J.-B.; Jeon, Y.-J. In vitro studies of anti-inflammatory and anticancer activities of organic solvent extracts from cultured marine microalgae. Algae 2013, 28, 111–119. [Google Scholar] [CrossRef]
- Andersen, R.; Kawachi, M. Traditional microalgae isolation techniques. Algal Cult. Tec. Cap 2005, 6, 83–100. [Google Scholar]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms: I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Fritz, L.; Triemer, R.E. A rapid simple technique utilizing calcofluor white M2R for the visualization of dinoflagellate thecal plates. J. Phycol. 1985, 21, 662–664. [Google Scholar] [CrossRef]
- Mussai, P.; Larsen, J.; Jeewon, R. Molecular characterization and morpho-taxonomy of Gambierdiscus caribaeus Vandersea, Litaker, M.A.Faust, Kibler, W.C.Holland & P.A.Tester (Dinophyceae) from Mauritius Island, South-West Indian Ocean. Cryptogam. Algol. 2023, 44, 1–11. [Google Scholar]
- Richlen, M.L.; Barber, P.H. A technique for the rapid extraction of microalgal DNA from single live and preserved cells. Mol. Ecol. Notes 2005, 5, 688–691. [Google Scholar] [CrossRef]
- Scholin, C.A.; Herzog, M.; Sogin, M.; Anderson, D.M. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae) II. Sequence analysis of a fragment of the LSU rRNA gene. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, A. Antibiotic susceptibility testing by a standardized single diffusion method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Intitute, (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; Approved Standard: Ninth Edition; M07-A9; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Yasumoto, T.; Battocchi, C.; Giacobbe, M.; Amorim, A.; et al. Investigation of toxin profile of Mediterranean and Atlantic strains of Ostreopsis cf. siamensis (Dinophyceae) by liquid chromatography–high resolution mass spectrometry. Harmful Algae 2013, 23, 19–27. [Google Scholar] [CrossRef]
- Sánchez, B.; Chaignepain, S.; Schmitter, J.M.; Urdaci, M.C. A method for the identification of proteins secreted by lactic acid bacteria grown in complex media. FEMS Microbiol. Lett. 2009, 295, 226–229. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Zone of Inhibition | ||||||
|---|---|---|---|---|---|---|---|
| Gram-Positive Strains a | Gram-Negative Strains b | PC | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| Intracellular Extract | |||||||
| ISOP15 | 12 ± 0.2 | 20 ± 0.2 | 20 ± 0.25 | 30 ± 0.0 | 20 ± 0.25 | 10 ± 0.42 | 20 ± 0.3 |
| P29 | 13 ± 1.1 | 25 ± 0.4 | 23. ± 0.4 | 24 ± 0.8 | 15 ± 0.4 | 27 ± 0.0 | 24 ± 1.5 |
| ISOP40 | 15 ± 0.4 | 19 ± 0.4 | 15 ± 0.4 | 21 ± 0.4 | 21 ± 0.4 | 15 ± 0.4 | 14 ± 1.1 |
| Extracellular Protein | |||||||
| ISOP15 | 11 ± 0.2 | 7 ± 0.5 | 14 ± 0.2 | 27 ± 0.5 | 14 ± 0.2 | 15 ± 0.4 | 23 ± 1.0 |
| P29 | 15 ± 0.4 | 11 ± 0.0 | 7 ± 0.5 | 18 ± 0.5 | 16 ± 0.3 | 7 ± 0.4 | 13 ± 1.1 |
| ISOP40 | 12 ± 0.5 | 16 ± 1.2 | 9 ± 0.4 | 20 ± 0.4 | 17 ± 0.5 | 7 ± 0.5 | 18 ± 0.5 |
| Extracellular Polysaccharides | |||||||
| ISOP15 | 14 ± 0.2 | 20 ± 0.3 | 16 ± 1.1 | 28 ± 0.2 | 1 4 ± 1.1 | 15 ± 0.4 | 19 ± 0.2 |
| P29 | 14 ± 0.7 | 13 ± 0.1 | 12 ± 0.5 | 21 ± 0.6 | 15 ± 0.4 | 14 ± 1.5 | 24 ± 1.1 |
| ISOP40 | 24 ± 0.4 | 15 ± 0.4 | 15 ± 0.4 | 15 ± 0.4 | 15 ± 0.4 | 11 ± 0.2 | 20 ± 1.5 |
| Isolates | Minimum Inhibitory Concentration (μg/mL) | |||||
|---|---|---|---|---|---|---|
| Gram-Positive Strains a | Gram-Negative Strains b | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | |
| Intracellular Extract | ||||||
| ISOP15 | 5 | 2.5 | 1.25 | 2.5 | 2.5 | 5 |
| P29 | 5 | 2.5 | 0.625 | 1.25 | 5 | 1.25 |
| ISOP40 | 1.25 | 1.25 | 1.25 | 0.625 | 5 | 2.5 |
| Extracellular Protein | ||||||
| ISOP15 | 10 | 10 | 5 | 0.625 | 2.5 | 5 |
| P29 | 5 | 5 | 10 | 5 | 5 | 10 |
| ISOP40 | 2.5 | 5 | 5 | 2.5 | 2.5 | 10 |
| Extracellular Polysaccharides | ||||||
| ISOP15 | 1.25 | 5 | 2.5 | 0.625 | 5 | 5 |
| P29 | 5 | 5 | 5 | 2.5 | 5 | 2.5 |
| ISOP40 | 0.625 | 2.5 | 5 | 5 | 5 | 2.5 |
| Positive Control | ||||||
| 10 µg/mL streptomycin | 0.625 | 1.25 | 1.25 | 0.625 | 0.625 | 0.625 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mussai, P.; Larsen, J.; Alrefaei, A.F.; Jeewon, R. Ribosomal DNA Sequence-Based Taxonomy and Antimicrobial Activity of Prorocentrum spp. (Dinophyceae) from Mauritius Coastal Waters, South-West Indian Ocean. Mar. Drugs 2023, 21, 216. https://doi.org/10.3390/md21040216
Mussai P, Larsen J, Alrefaei AF, Jeewon R. Ribosomal DNA Sequence-Based Taxonomy and Antimicrobial Activity of Prorocentrum spp. (Dinophyceae) from Mauritius Coastal Waters, South-West Indian Ocean. Marine Drugs. 2023; 21(4):216. https://doi.org/10.3390/md21040216
Chicago/Turabian StyleMussai, Prakash, Jacob Larsen, Abdulwahed Fahad Alrefaei, and Rajesh Jeewon. 2023. "Ribosomal DNA Sequence-Based Taxonomy and Antimicrobial Activity of Prorocentrum spp. (Dinophyceae) from Mauritius Coastal Waters, South-West Indian Ocean" Marine Drugs 21, no. 4: 216. https://doi.org/10.3390/md21040216
APA StyleMussai, P., Larsen, J., Alrefaei, A. F., & Jeewon, R. (2023). Ribosomal DNA Sequence-Based Taxonomy and Antimicrobial Activity of Prorocentrum spp. (Dinophyceae) from Mauritius Coastal Waters, South-West Indian Ocean. Marine Drugs, 21(4), 216. https://doi.org/10.3390/md21040216