
Tisochrysis lutea F&M-M36 Mitigates Risk Factors of Metabolic Syndrome and Promotes Visceral Fat Browning through β3-Adrenergic Receptor/UCP1 Signaling
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Effects of T. lutea F&M-M36 vs. Fenofibrate on Body and Organs Weight and on Fat Mass
2.2. Effects of T. lutea F&M-M36 and Fenofibrate on Metabolic Profile, Adiponectin, and Blood Pressure
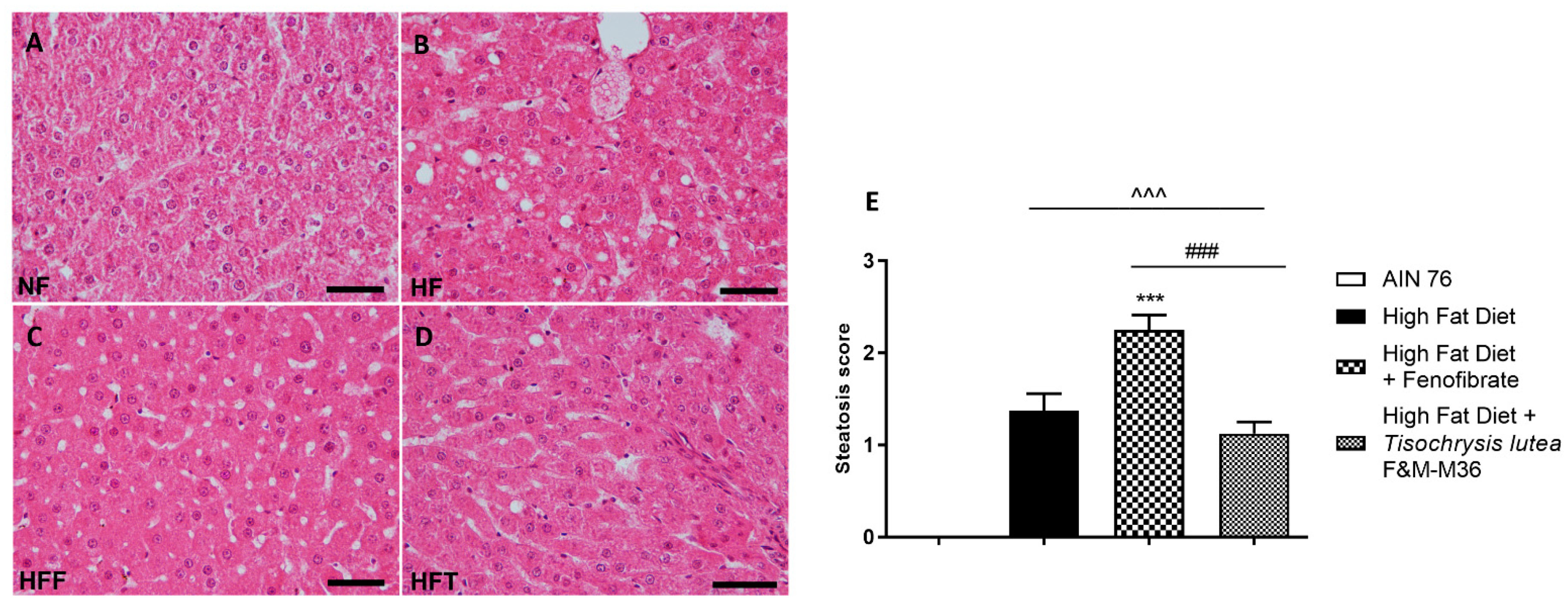
2.3. Effects of T. lutea F&M-M36 and Fenofibrate on Hepatic Steatosis
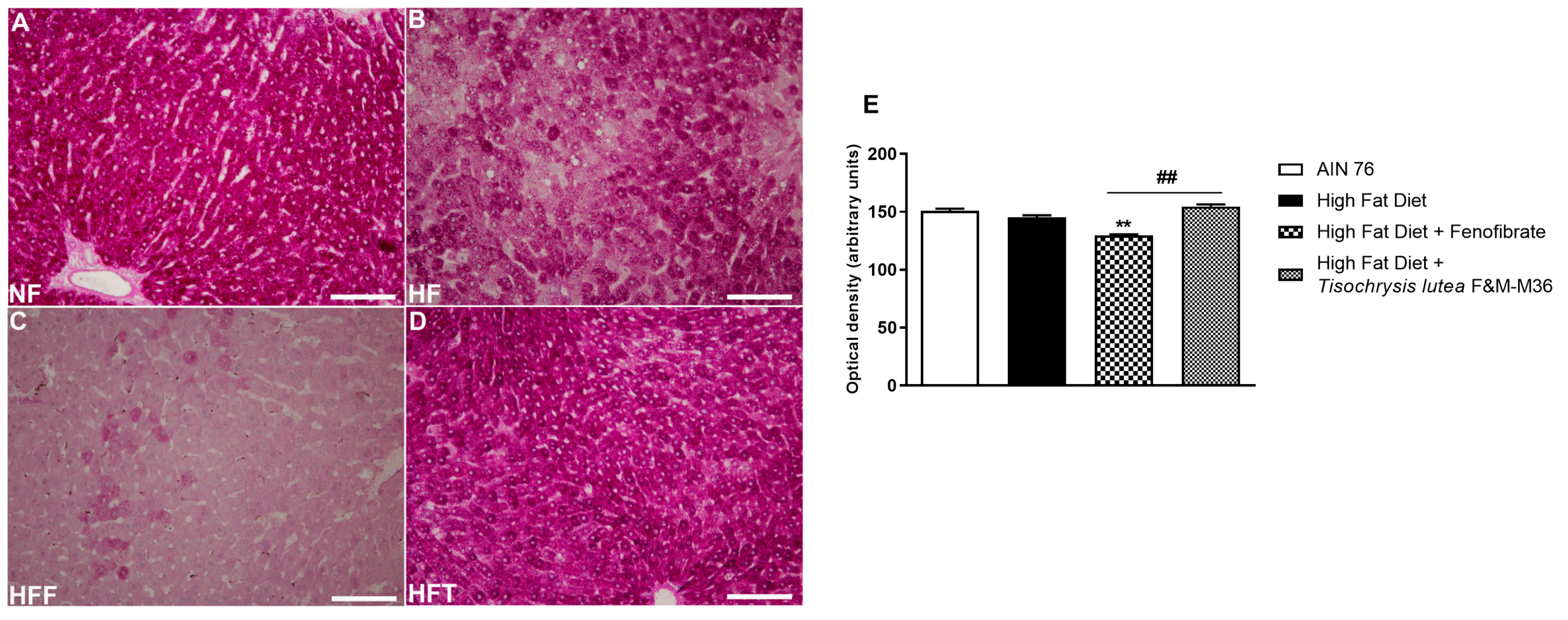
2.4. Effects of T. lutea F&M-M36 and Fenofibrate on Glycogen Storage in the Liver
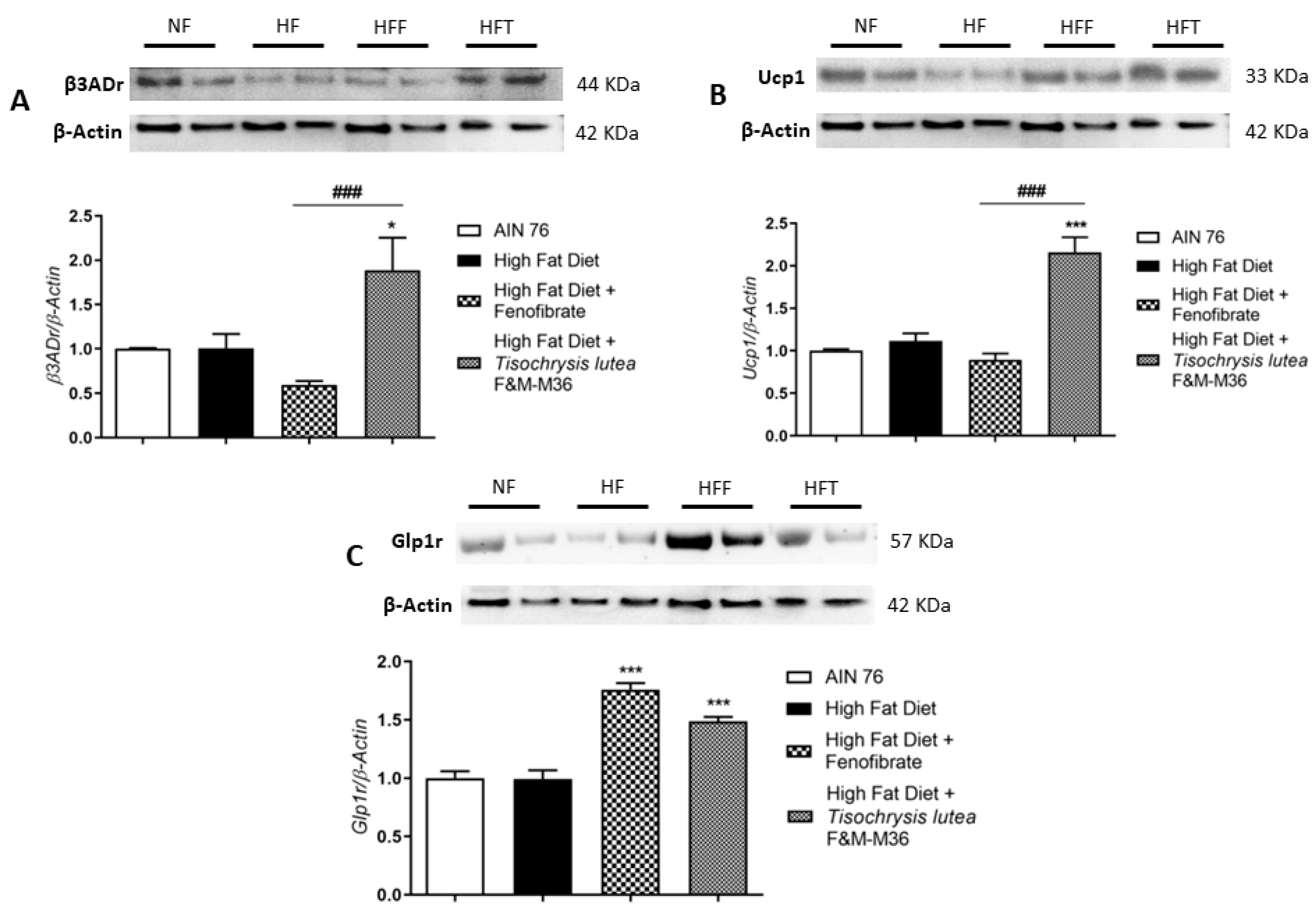
2.5. Effects of T. lutea F&M-M36 and Fenofibrate on β3ADr, Ucp1, and Glp1r Protein Expression in Visceral Adipose Tissue
2.6. Effects of T. lutea F&M-M36 and Fenofibrate on Pro-Inflammatory Cytokines mRNA Expression in Visceral Adipose Tissue
2.7. Effect of T. lutea F&M-M36 on Whole-Gene Expression Profiles in Visceral Adipose Tissue
3. Discussion
4. Materials and Methods
4.1. Microalgae Cultivation and Production
4.2. Animals and Treatment Design
4.3. Blood Pressure Measurement
4.4. Food and Water Consumption
4.5. Macroscopic Examinations and Histological Analyses
4.6. Blood Biochemistry
4.7. Fecal Lipid Content
4.8. Periodic Acid–Schiff (PAS) Staining
4.9. Western Blot
4.10. Total RNA Extraction and Real-Time PCR
4.11. Gene Expression Profiling
4.12. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M. Drug therapy of the metabolic syndrome: Minimizing the emerging crisis in polypharmacy. Nat. Rev. Drug Discov. 2006, 5, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, K.; Oku, M.; Hayashi, S.; Inujima, A.; Shibahara, N.; Chen, L.; Igarashi, Y.; Tobe, K.; Saito, S.; Kadowaki, M.; et al. Identifying pre-disease signals before metabolic syndrome in mice by dynamical network biomarkers. Sci. Rep. 2019, 9, 8767. [Google Scholar] [CrossRef] [PubMed]
- Bigagli, E.; Cinci, L.; Niccolai, A.; Biondi, N.; Rodolfi, L.; D’Ottavio, M.; D’Ambrosio, M.; Lodovici, M.; Tredici, M.R.; Luceri, C. Preliminary data on the dietary safety, tolerability and effects on lipid metabolism of the marine microalga Tisochrysis lutea. Algal. Res. 2018, 34, 244–249. [Google Scholar] [CrossRef]
- Bigagli, E.; D’Ambrosio, M.; Cinci, L.; Niccolai, A.; Biondi, N.; Rodolfi, L.; Dos Santos Nascimiento, L.B.; Tredici, M.R.; Luceri, C. A Comparative In Vitro Evaluation of the Anti-Inflammatory Effects of a Tisochrysis lutea Extract and Fucoxanthin. Mar. Drugs 2021, 19, 334. [Google Scholar] [CrossRef]
- Niccolai, A.; Chini Zittelli, G.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Microalgae of interest as food source: Biochemical composition and digestibility. Algal Res. 2019, 42, 101617. [Google Scholar] [CrossRef]
- Mayer, C.; Richard, L.; Côme, M.; Ulmann, L.; Nazih, H.; Chénais, B.; Ouguerram, K.; Mimouni, V. The Marine Microalga, Tisochrysis lutea, Protects against Metabolic Disorders Associated with Metabolic Syndrome and Obesity. Nutrients 2021, 13, 430. [Google Scholar] [CrossRef]
- Custódio, L.; Soares, F.; Pereira, H.; Barreira, L.; Vizetto-Duarte, C.; Rodrigues, M.J.; Rauter, A.P.; Alberício, F.; Varela, J. Fatty acid composition and biological activities of Isochrysis galbana T-ISO, Tetraselmis sp. and Scenedesmus sp.: Possible application in the pharmaceutical and functional food industries. J. Appl. Phycol. 2014, 26, 151–161. [Google Scholar] [CrossRef]
- Tenenbaum, A.; Fisman, E.Z. Fibrates are an essential part of modern anti-dyslipidemic arsenal: Spotlight on atherogenic dyslipidemia and residual risk reduction. Cardiovasc. Diabetol. 2012, 11, 125. [Google Scholar] [CrossRef]
- Duan, Y.; Zeng, L.; Zheng, C.; Song, B.; Li, F.; Kong, X.; Xu, K. Inflammatory Links Between High Fat Diets and Diseases. Front. Immunol. 2018, 9, 2649. [Google Scholar] [CrossRef] [PubMed]
- Vidigal Fde, C.; Ribeiro, A.Q.; Babio, N.; Salas-Salvadó, J.; Bressan, J. Prevalence of metabolic syndrome and pre-metabolic syndrome in health professionals: Latinmets brazil study. Diabetol. Metab. Syndr. 2015, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- de las Fuentes, L.; Brown, A.L.; Mathews, S.J.; Waggoner, A.D.; Soto, P.F.; Gropler, R.J.; Dávila-Román, V.G. Metabolic syndrome is associated with abnormal left ventricular diastolic function independent of left ventricular mass. Eur. Heart J. 2007, 28, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Gesteiro, E.; Megía, A.; Guadalupe-Grau, A.; Fernandez-Veledo, S.; Vendrell, J.; González-Gross, M. Early identification of metabolic syndrome risk: A review of reviews and proposal for defining pre-metabolic syndrome status. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2557–2574. [Google Scholar] [CrossRef]
- Ferreira, A.V.; Parreira, G.G.; Green, A.; Botion, L.M. Effects of fenofibrate on lipid metabolism in adipose tissue of rats. Metabolism 2006, 55, 731–735. [Google Scholar] [CrossRef]
- Zhang, Y.; Jia, X.B.; Liu, Y.C.; Yu, W.Q.; Si, Y.H.; Guo, S.D. Fenofibrate enhances lipid deposition via modulating PPARγ, SREBP-1c, and gut microbiota in ob/ob mice fed a high-fat diet. Front. Nutr. 2022, 9, 971581. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Li, X.; Feng, J.; Chen, X. Association of metabolic syndrome with atherogenic index of plasma in an urban Chinese population: A 15-year prospective study. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 1214–1219. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, Q.; Wei, Z.; Wei, J.; Cui, M. Atherogenic Index of Plasma and Coronary Artery Disease in the Adult Population: A Meta-Analysis. Front. Cardiovasc. Med. 2021, 8, 817441. [Google Scholar] [CrossRef]
- Rosenson, R.S. Effect of fenofibrate on adiponectin and inflammatory biomarkers in metabolic syndrome patients. Obes. Silver Spring 2009, 17, 504–509. [Google Scholar] [CrossRef]
- Lodovici, M.; Bigagli, E.; Tarantini, F.; Di Serio, C.; Raimondi, L. Losartan reduces oxidative damage to renal DNA and conserves plasma antioxidant capacity in diabetic rats. Exp. Biol. Med. 2015, 240, 1500–1504. [Google Scholar] [CrossRef]
- Manno, C.; Campobasso, N.; Nardecchia, A.; Triggiani, V.; Zupo, R.; Gesualdo, L.; Silvestris, F.; De Pergola, G. Relationship of para- and perirenal fat and epicardial fat with metabolic parameters in overweight and obese subjects. Eat Weight Disord. 2019, 24, 67–72. [Google Scholar] [CrossRef] [PubMed]
- D’Marco, L.; Salazar, J.; Cortez, M.; Salazar, M.; Wettel, M.; Lima-Martínez, M.; Rojas, E.; Roque, W.; Bermúdez, V. Perirenal fat thickness is associated with metabolic risk factors in patients with chronic kidney disease. Kidney Res. Clin. Pract. 2019, 38, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Tu, M.; Chen, Y.; Wang, W. Perirenal Fat Thickness: A Surrogate Marker for Metabolic Syndrome in Chinese Newly Diagnosed Type 2 Diabetes. Front. Endocrinol. 2022, 13, 850334. [Google Scholar] [CrossRef] [PubMed]
- De Pergola, G.; Campobasso, N.; Nardecchia, A.; Triggiani, V.; Caccavo, D.; Gesualdo, L.; Silvestris, F.; Manno, C. Para- and perirenal ultrasonographic fat thickness is associated with 24-h mean diastolic blood pressure levels in overweight and obese subjects. BMC Cardiovasc. Disord. 2015, 15, 108. [Google Scholar] [CrossRef] [PubMed]
- El Hadi, H.; Di Vincenzo, A.; Vettor, R.; Rossato, M. Food Ingredients Involved in White-to-Brown Adipose Tissue Conversion and in Calorie Burning. Front. Physiol. 2019, 9, 1954. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, C.; Sul, H.S. Signaling Pathways Regulating Thermogenesis. Front. Endocrinol. 2021, 12, 595020. [Google Scholar] [CrossRef]
- Ricquier, D. Uncoupling protein 1 of brown adipocytes, the only uncoupler: A historical perspective. Front. Endocrinol. 2011, 2, 85. [Google Scholar] [CrossRef]
- Park, Y.; Harris, W.S. Dose-dependent effects of n-3 polyunsaturated fatty acids on platelet activation in mildly hypertriglyceridemic subjects. J. Med. Food 2009, 12, 809–813. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochem. Biophys. Res. Commun. 2005, 332, 392–397. [Google Scholar] [CrossRef]
- Wu, M.T.; Chou, H.N.; Huang, C.J. Dietary fucoxanthin increases metabolic rate and upregulated mRNA expressions of the PGC-1alpha network, mitochondrial biogenesis and fusion genes in white adipose tissues of mice. Mar. Drugs 2014, 12, 964–982. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Z.; Tan, B.K.; Christian, M. Dietary polyphenols turn fat “brown”: A narrative review of the possible mechanisms. Trends Food Sci. Technol. 2020, 97, 221–232. [Google Scholar] [CrossRef]
- Kang, N.H.; Mukherjee, S.; Yun, J.W. Trans-Cinnamic Acid Stimulates White Fat Browning and Activates Brown Adipocytes. Nutrients 2019, 11, 577. [Google Scholar] [CrossRef]
- Valasek, M.A.; Clarke, S.L.; Repa, J.J. Fenofibrate reduces intestinal cholesterol absorption via PPARalpha-dependent modulation of NPC1L1 expression in mouse. J. Lipid Res. 2007, 48, 2725–2735. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Schwartz, S.M. Pharmacologic and Pharmacodynamic Equivalence of 2 Formulations of Orlistat. Clin. Pharmacol. Drug Dev. 2018, 7, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Chen, G.; Ma, M.; Qiu, N.; Zhu, L.; Li, J. Fatty acids modulate the expression levels of key proteins for cholesterol absorption in Caco-2 monolayer. Lipids Health Dis. 2018, 17, 32. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Hosokawa, M.; Matsukawa, N.; Hagio, M.; Shinoki, A.; Nishimukai, M.; Miyashita, K.; Yajima, T.; Hara, H. Suppressive effects of the marine carotenoids, fucoxanthin and fucoxanthinol on triglyceride absorption in lymph duct-cannulated rats. Eur. J. Nutr. 2010, 49, 243–249. [Google Scholar] [CrossRef]
- Ha, A.W.; Kim, W.K. The effect of fucoxanthin rich power on the lipid metabolism in rats with a high fat diet. Nutr. Res. Pract. 2013, 7, 287–293. [Google Scholar] [CrossRef]
- Wei, X.H.; Guo, X.; Pan, C.S.; Li, H.; Cui, Y.C.; Yan, L.; Fan, J.Y.; Deng, J.N.; Hu, B.H.; Chang, X.; et al. Quantitative Proteomics Reveal That Metabolic Improvement Contributes to the Cardioprotective Effect of T89 on Isoproterenol-Induced Cardiac Injury. Front. Physiol. 2021, 12, 653349. [Google Scholar] [CrossRef]
- Suárez, J.; Rivera, P.; Arrabal, S.; Crespillo, A.; Serrano, A.; Baixeras, E.; Pavón, F.J.; Cifuentes, M.; Nogueiras, R.; Ballesteros, J.; et al. Oleoylethanolamide enhances β-adrenergic-mediated thermogenesis and white-to-brown adipocyte phenotype in epididymal white adipose tissue in rat. Dis. Model Mech. 2014, 7, 129–141. [Google Scholar] [CrossRef]
- Dou, H.X.; Wang, T.; Su, H.X.; Gao, D.D.; Xu, Y.C.; Li, Y.X.; Wang, H.Y. Exogenous FABP4 interferes with differentiation, promotes lipolysis and inflammation in adipocytes. Endocrine 2020, 67, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Mita, T.; Furuhashi, M.; Hiramitsu, S.; Ishii, J.; Hoshina, K.; Ishimura, S.; Fuseya, T.; Watanabe, Y.; Tanaka, M.; Ohno, K.; et al. FABP4 is secreted from adipocytes by adenyl cyclase-PKA- and guanylyl cyclase-PKG-dependent lipolytic mechanisms. Obes. Silver Spring 2015, 23, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Senga, S.; Kobayashi, N.; Kawaguchi, K.; Ando, A.; Fujii, H. Fatty acid-binding protein 5 (FABP5) promotes lipolysis of lipid droplets, de novo fatty acid (FA) synthesis and activation of nuclear factor-kappa B (NF-κB) signaling in cancer cells. Biochim. Biophys. Acta Mol. Cell Biol. Lipids. 2018, 1863, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.S.; Hong, J.H.; Cho, K.H.; Jung, H.K. Grape skin extract reduces adipogenesis- and lipogenesis-related gene expression in 3T3-L1 adipocytes through the peroxisome proliferator-activated receptor-γ signaling pathway. Nutr. Res. 2012, 32, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, L.; Yang, W.; Wang, C.; Hu, G.; Mo, Z. Profiling of differentially expressed genes in adipose tissues of multiple symmetric lipomatosis. Mol. Med. Rep. 2017, 16, 6570–6579. [Google Scholar] [CrossRef]
- Ferhat, M.; Funai, K.; Boudina, S. Autophagy in Adipose Tissue Physiology and Pathophysiology. Antioxid. Redox Signal. 2019, 3, 487–501. [Google Scholar] [CrossRef]
- Lee, Y.S.; Park, M.S.; Choung, J.S.; Kim, S.S.; Oh, H.H.; Choi, C.S.; Ha, S.Y.; Kang, Y.; Kim, Y.; Jun, H.S. Glucagon-like peptide-1 inhibits adipose tissue macrophage infiltration and inflammation in an obese mouse model of diabetes. Diabetologia 2012, 55, 2456–2468. [Google Scholar] [CrossRef]
- Izaguirre, M.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Becerril, S.; Valentí, V.; Moncada, R.; Unamuno, X.; Silva, C.; de la Higuera, M.; et al. GLP-1 Limits Adipocyte Inflammation and Its Low Circulating Pre-Operative Concentrations Predict Worse Type 2 Diabetes Remission after Bariatric Surgery in Obese Patients. J. Clin. Med. 2019, 8, 479. [Google Scholar] [CrossRef]
- Tredici, M.R.; Rodolfi, L.; Biondi, N.; Bassi, N.; Sampietro, G. Techno-economic analysis of microalgal biomass production in a 1-ha Green Wall Panel (GWP®) plant. Algal Res. 2016, 19, 253–263. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt and Detonula confervacea Cleve. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Whitman, M.; Jenkins, C. Rate pressure product, age predicted maximum heart rate or heart rate reserve. Which one better predicts cardiovascular events following exercise stress echocardiography? Am. J. Cardiovasc. Dis. 2021, 11, 450–457. [Google Scholar] [PubMed]
- Takahashi, Y.; Fukusato, T. Histopathology of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. World J. Gastroenterol. 2014, 20, 15539–15548. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.W.; Lu, Y.; Li, F.; Yang, C.J.; Feng, Y.B.; Li, H.W.; Yao, W.F.; Shen, Z.H. Atherogenic index of plasma is an effective index for estimating abdominal obesity. Lipids Health Dis. 2018, 17, 11. [Google Scholar] [CrossRef]
- Kraus, D.; Yang, Q.; Kahn, B.B. Lipid Extraction from Mouse Feces. Bio-Protoc. 2015, 5, e1375. [Google Scholar] [CrossRef] [PubMed]
- Diaz Marin, R.; Crespo-Garcia, S.; Wilson, A.M.; Sapieha, P. RELi protocol: Optimization for protein extraction from white, brown and beige adipose tissues. MethodsX 2019, 6, 918–928. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NF | HF | HFF | HFT | |
|---|---|---|---|---|
| Weight gain (g) | 213.5 ± 7.7 | 252.5 ± 20.1 | 208.5 ± 11.6 | 281.3 ± 11.6 |
| Food intake (g) | 51.66 ± 0.3 | 27.51 ± 0.5 ^^^ | 31.55 ± 0.5 ^^^ | 27.51 ± 0.5 ^^^ |
| Calorie intake (kcal)/24 h | 195.8 ± 1.3 | 146.6 ± 2.64 ^^^ | 168.2 ± 2.4 ^^^ | 146.3 ± 2.63 ^^^ |
| Water intake (mL) | 52.47 ± 2 | 50.27 ± 1.8 | 58.27 ± 1.7 | 52.80 ± 1.2 |
| Fecal lipids excretion (µg/g 24 h dw) | 1.4 ± 0.2 | 2.0 ± 0.2 | 5.4 ± 1.2 ^^** | 4.2 ± 0.6 ^* |
| Liver (w/bw × 10−3) | 38.86 ± 0.80 | 36.85 ± 1.38 | 65.44 ± 0.95 ***^^^ | 32.63 ± 0.29 |
| Kidney (w/bw × 10−3) | 3.23 ± 0.07 | 3.18 ± 0.10 | 4.21 ± 0.13 ***^^^ | 3.04 ± 0.10 |
| Heart (w/bw × 10−3) | 3.53 ± 0.12 | 3.38 ± 0.06 | 3.41 ± 0.09 | 3.70 ± 0.09 |
| Visceral fat (w/bw × 10−3) | 1.38 ± 0.14 | 2.30 ± 0.24 | 2.04 ± 0.39 | 1.83 ± 0.22 |
| Epididymal fat (w/bw × 10−3) | 7.64 ± 0.73 | 12.37 ± 0.67 ^ | 10.24 ± 1.61 | 10.93 ± 1.20 |
| Renal fat (w/bw × 10−3) | 8.56 ± 1.40 | 15.89 ± 1.49 ^^ | 12.46 ± 1.40 | 10.26 ± 0.55 * |
| NF | HF | HFF | HFT | |
|---|---|---|---|---|
| TG (mg/dL) | 183.7 ± 15.1 | 254.0 ± 21.6 ^^ | 90.5 ± 3.2 ***^^^ | 141.0 ± 7.9 ** |
| TC (mg/dL) | 128 ± 5.1 | 134 ± 7.0 | 130 ± 8.3 | 139 ± 22.2 |
| HDL (mg/dL) | 89 ± 5.64 | 68 ± 7.8 | 78 ± 6.5 | 80 ± 10.2 |
| AIP | 2.2 ± 0.36 | 3.5 ± 0.5 | 1.2 ± 0.1 ** | 2.0 ± 0.36 * |
| Glucose (mg/dL) | 168.2 ± 9.1 | 212.1 ± 11.1 ^ | 137.5 ± 9.4 *** | 150.9 ± 9.1 ** |
| Adiponectin (ng/mL) | 37.9 ± 3.5 | 28.9 ± 1.8 | 80.5 ± 5.9 ***^^^ | 57.9 ± 3.4 ***^^ |
| Urinary uric acid (mg/dL) | 21.07 ± 4.6 | 25.05 ± 7.5 | 14.80 ± 6.1 | 13 ± 3.9 |
| SBP (mm Hg) | 156.2 ± 5.2 | 159.7 ± 3.6 | 164.6 ± 2.8 | 147.9 ± 1.9 |
| DBP (mm Hg) | 94.9 ± 6.4 | 111.7 ± 4.4 | 105.9 ± 8.1 | 80.56 ± 4.4 ** |
| MAP (mm Hg) | 115.3 ± 5.6 | 127.7 ± 4.1 | 125.4 ± 5.5 | 107.1 ± 4.9 * |
| RPP (mm Hg bpm) | 61,342 ± 2650 | 68,703 ± 3012 | 67,409 ± 3093 | 58,338 ± 1555 * |
| Up-Regulated | |||
|---|---|---|---|
| Gene-Set Name | Percent Changed | PermuteP | Gene Symbols |
| Notch signaling pathway:KEGG-rno04330 | 50.0 | 0.002 | Dll3|Dtx1|Jag2|Ncstn|Notch3|Notch4|Ptcra|Rbpj |
| Protein export:KEGG-rno03060 | 44.4 | 0.04 | LOC100361694|Oxa1l|Sec11c|Spcs2 |
| Mismatch repair:KEGG-rno03430 | 44.4 | 0.05 | Exo1|Lig1|Mlh1|Pold3 |
| Galactose metabolism:KEGG-rno00052 | 41.7 | 0.03 | Gaa|Galk1|Hk2|Hk3|Pfkl |
| Endocrine and other factor-regulated calcium reabsorption:KEGG-rno04961 | 33.3 | 0.02 | Ap2b1|Ap2m1|Clta|Cltb|Cltc|Dnm2|Plcb3|Plcb4|Prkcg |
| PPAR signaling pathway:KEGG-rno03320 | 33.3 | 0.01 | Acox1|Acsbg1|Dbi|Fabp4|Fabp5|LOC681458|Mmp1|Nr1h3|Pparg|Scd1|Slc27a1|Ubc |
| Huntington’s disease:KEGG-rno05016 | 31.9 | 0.00 | Ap2b1|Ap2m1|Atp5g1|Atp5g2|Atp5g3|Atp5hl1|Atp5o|Clta|Cltb|Cltc|Cox4i2|Cox5b|Dnali1|Grm5|LOC688963|Mt-co1|Ndufa10|Ndufa6|Ndufb10|Ndufb11|Ndufb2|Ndufs7|Ndufv1|Plcb3|Plcb4|Pparg|Sod1|Tp53|Uqcrh|Vdac3 |
| Gap junction:KEGG-rno04540 | 31.3 | 0.02 | Adcy5|Gnai2|Grm5|Htr2a|Plcb3|Plcb4|Prkcg|Tuba3a|Tubb4b|Tubb5 |
| Bacterial invasion of epithelial cells:KEGG-rno05100 | 31.3 | 0.02 | Arpc1a|Cdc42|Clta|Cltb|Cltc|Ctnnb1|Cttn|Dnm2|Pik3r2|Pxn |
| Pancreatic secretion:KEGG-rno04972 | 30.8 | 0.03 | Adcy5|Atp2a3|Cela2a|Clca1|Cpa1|Ctrb1|Pla2g1b|Plcb3|Plcb4|Prkcg|Rap1b|Slc4a2 |
| Antigen processing and presentation:KEGG-rno04612 | 28.9 | 0.04 | Calr|Ctsb|Hsp90aa1|LOC680121|LOC688090|Psme2|RT1-CE3|RT1-Da|RT1-M1-4|RT1-M6-2|RT1-T18 |
| Parkinson’s disease:KEGG-rno05012 | 28.4 | 0.004 | Atp5g1|Atp5g2|Atp5g3|Atp5hl1|Atp5o|Cox4i2|Cox5b|Gp1bb|LOC688963|Mt-co1|Ndufa10|Ndufa6|Ndufb10|Ndufb11|Ndufb2|Ndufs7|Ndufv1|Th|Uba1|Ubc|Ube2l3|Uqcrh|Vdac3 |
| Alzheimer’s disease:KEGG-rno05010 | 25.0 | 0.03 | Atp2a3|Atp5g1|Atp5g2|Atp5g3|Atp5hl1|Atp5o|Calm1|Cox4i2|Cox5b|Grin2c|LOC688963|Mt-co1|Ncstn|Ndufa10|Ndufa6|Ndufb10|Ndufb11|Ndufb2|Ndufs7|Ndufv1|Plcb3|Plcb4|Uqcrh |
| Down-regulated | |||
| Gene-Set Name | Percent Changed | PermuteP | gene symbols |
| Cytokine-cytokine receptor interaction:KEGG-rno04060 | 26.0 | 0.0085 | Ccl12|Ccr6|Cd40|Csf2rb|Cxcl11|Cxcl13|Cxcr4|Egf|Ifna2|Ifnar2|Il11ra1|Il12b|Il22|LOC100910178|Lta|Osmr|Pdgfrb|RGD1561246|Tnfrsf12a|Tnfrsf21|Tnfrsf8 |
| Regulation of autophagy:KEGG-rno04140 | 40 | 0.049 | Becn1|Ifna2|Prkaa2|RGD1561246 |
| AIN-76 Diet (NF) | High-Fat Diet (HF) | T. lutea F&M-M36 Enriched Diet (HFT) | |
|---|---|---|---|
| Lyophilized algal biomass | 5 | ||
| Corn oil | 5 | 3 | 2 |
| Lard | - | 30 | 30 |
| Sucrose | 50 | 34 | 33.4 |
| Starch | 15 | ||
| Casein | 20 | 24.6 | 22.5 |
| Cellulose | 5 | 2 | 1.1 |
| Mineral Mix AIN 76 | 3.5 | 4.1 | 4.1 |
| Vitamin Mix AIN 76 | 1 | 1.3 | 1 |
| Coline | 0.2 | 0.26 | 0.26 |
| DL Methionine | 0.3 | 0.4 | 0.4 |
| Antibody | Dilution | Supplier |
|---|---|---|
| β3ADr | 1:500 | Santa Cruz Biotechnology Inc. Dallas, TX, USA (SC-515763) |
| UCP-1 | 1:500 | Santa Cruz Biotechnology Inc. Dallas, TX, USA (SC-2934184) |
| GLP1r | 1:200 | Santa Cruz Biotechnology Inc. Dallas, TX, USA (sc-390774) |
| β-Actin | 1:1000 | Bioss Antibodies Woburn, MS, USA (bs-0061R) |
| Gene | Primer Forward | Primer Reverse |
|---|---|---|
| β-Actin | TACAGCTTCACCACCACAGC | TGGCCATCTCTTGCTCGAAG |
| IL-1β | GACTTCACCATGGAACCCGT | GGAGACTGCCCATTCTCGAC |
| IL-6 | GTGGCTAAGGACCAAGACCA | TAGCACACTAGGTTTGCCGAG |
| TNFα | AACACACGAGACGCTGAAGT | TCCAGTGAGTTCCGAAAGCC |
| ADRB3 | ACTCACCGCTCAACAGGTTT | TTCTGGAGAGTTGCGGTTCC |
| UCP1 | CCGAAACTGTACAGCGGTCT | CAGGAGTGTGGTGCAAAACC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Ambrosio, M.; Bigagli, E.; Cinci, L.; Gencarelli, M.; Chioccioli, S.; Biondi, N.; Rodolfi, L.; Niccolai, A.; Zambelli, F.; Laurino, A.; et al. Tisochrysis lutea F&M-M36 Mitigates Risk Factors of Metabolic Syndrome and Promotes Visceral Fat Browning through β3-Adrenergic Receptor/UCP1 Signaling. Mar. Drugs 2023, 21, 303. https://doi.org/10.3390/md21050303
D’Ambrosio M, Bigagli E, Cinci L, Gencarelli M, Chioccioli S, Biondi N, Rodolfi L, Niccolai A, Zambelli F, Laurino A, et al. Tisochrysis lutea F&M-M36 Mitigates Risk Factors of Metabolic Syndrome and Promotes Visceral Fat Browning through β3-Adrenergic Receptor/UCP1 Signaling. Marine Drugs. 2023; 21(5):303. https://doi.org/10.3390/md21050303
Chicago/Turabian StyleD’Ambrosio, Mario, Elisabetta Bigagli, Lorenzo Cinci, Manuela Gencarelli, Sofia Chioccioli, Natascia Biondi, Liliana Rodolfi, Alberto Niccolai, Francesca Zambelli, Annunziatina Laurino, and et al. 2023. "Tisochrysis lutea F&M-M36 Mitigates Risk Factors of Metabolic Syndrome and Promotes Visceral Fat Browning through β3-Adrenergic Receptor/UCP1 Signaling" Marine Drugs 21, no. 5: 303. https://doi.org/10.3390/md21050303