
Investigation of Physical Characteristics and In Vitro Anti-Inflammatory Effects of Fucoidan from Padina arborescens: A Comprehensive Assessment against Lipopolysaccharide-Induced Inflammation
 ,
,  ,
, 
Abstract
1. Introduction
2. Results
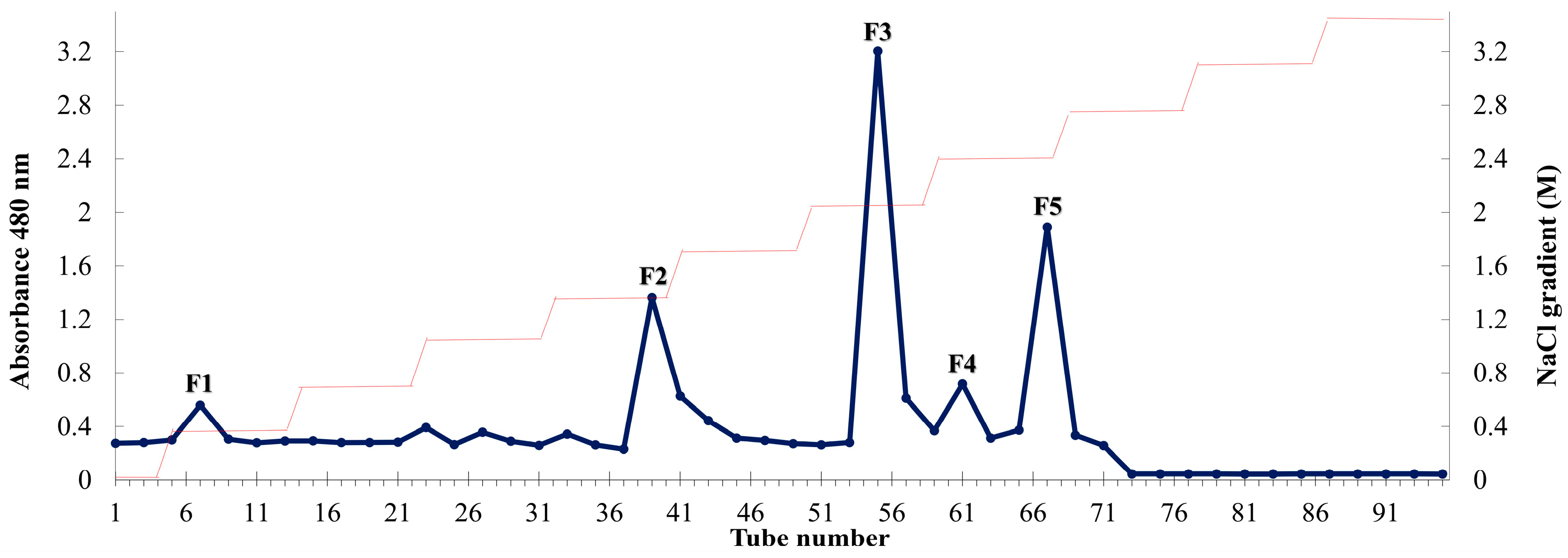
2.1. Composition of P. arborescens Extracts
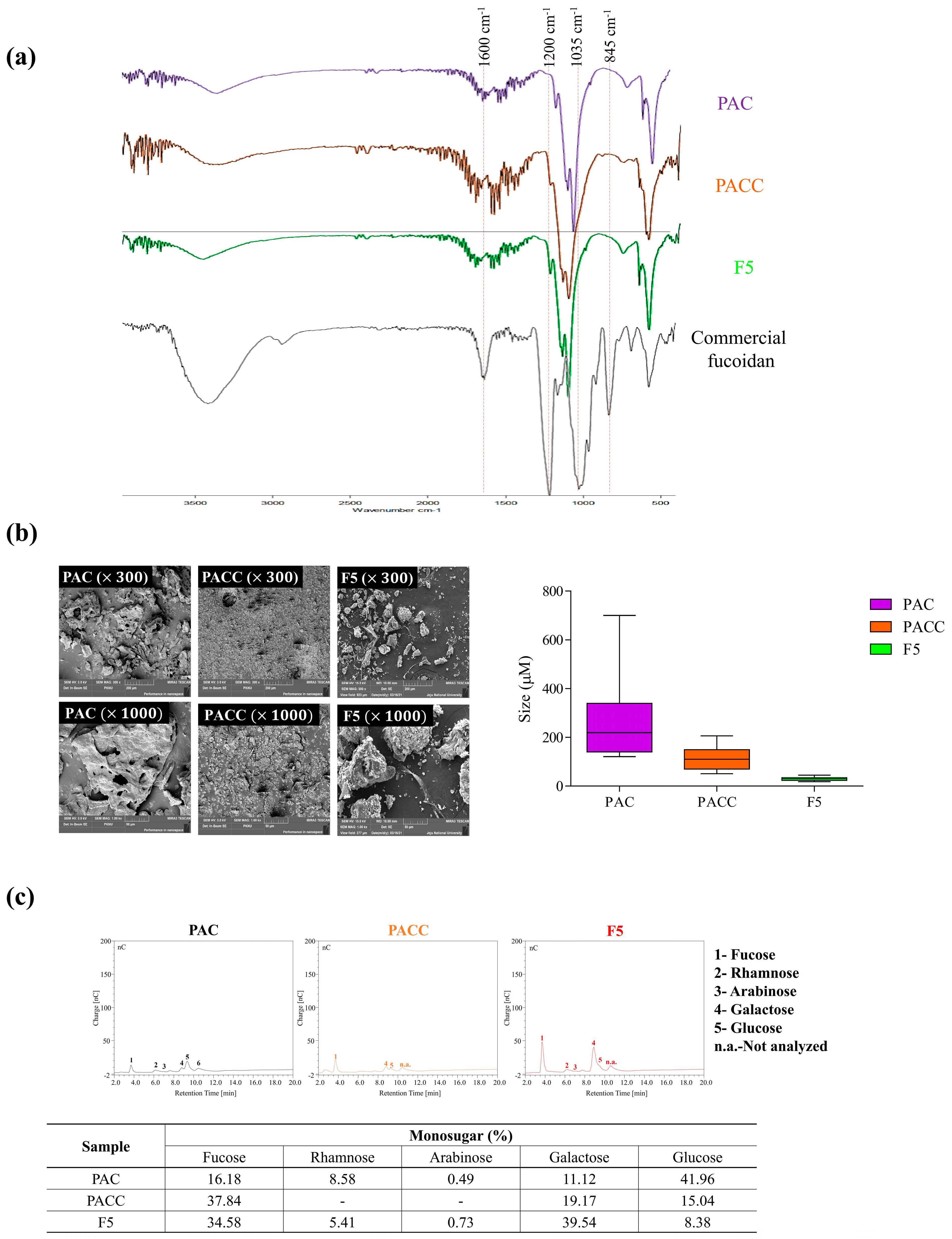
2.2. Polysaccharide Characterization through FTIR and Monosaccharide Profiling
2.3. Morphology and Size of F5
2.4. Monosaccharide Composition
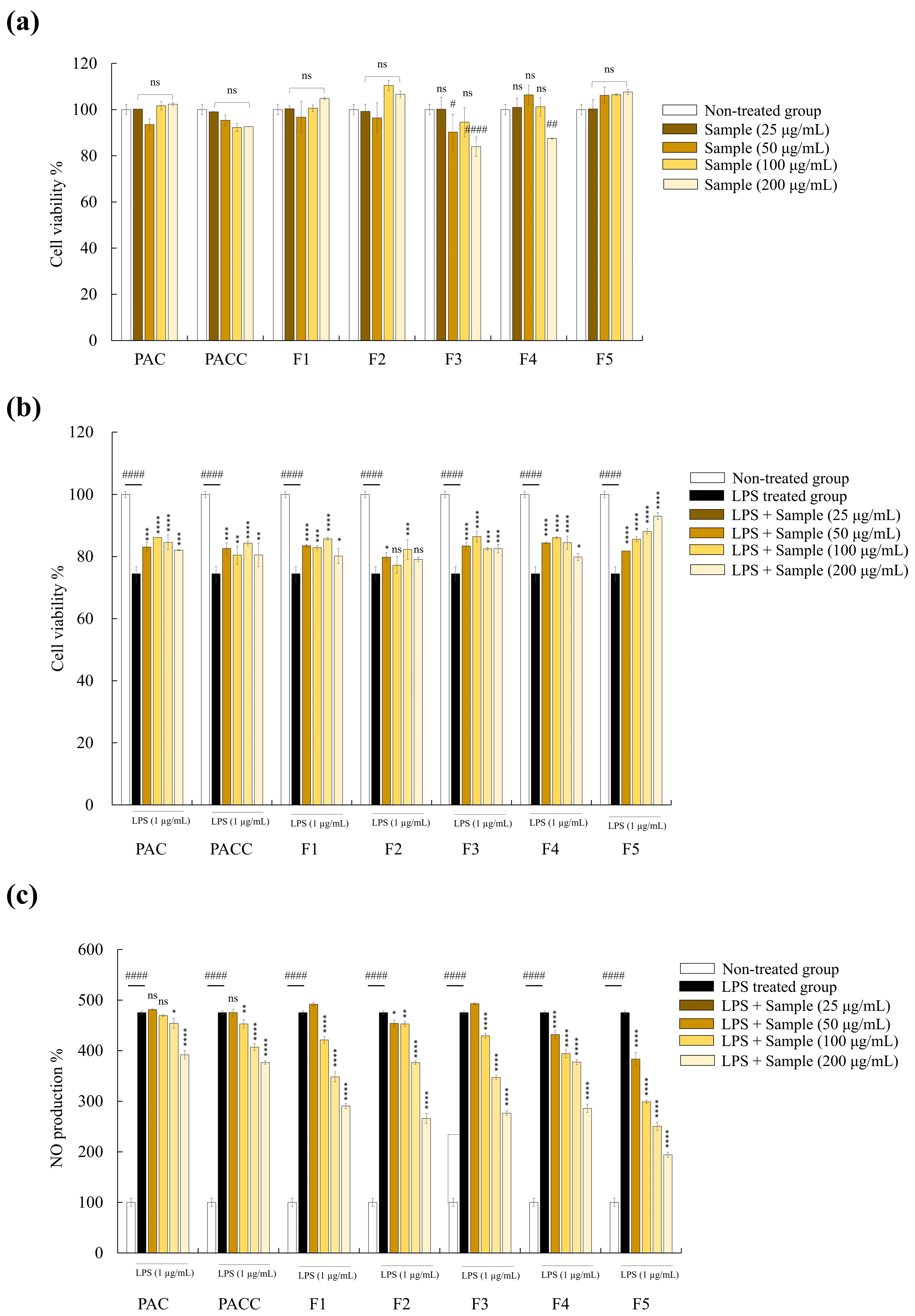
2.5. F5 Showed the Highest Protective Activity among Five Fucoidan Fractions
2.6. F5 Inhibits the Release of PGE2 and Inflammatory Cytokines from LPS-stimulated RAW 264.7 Cells
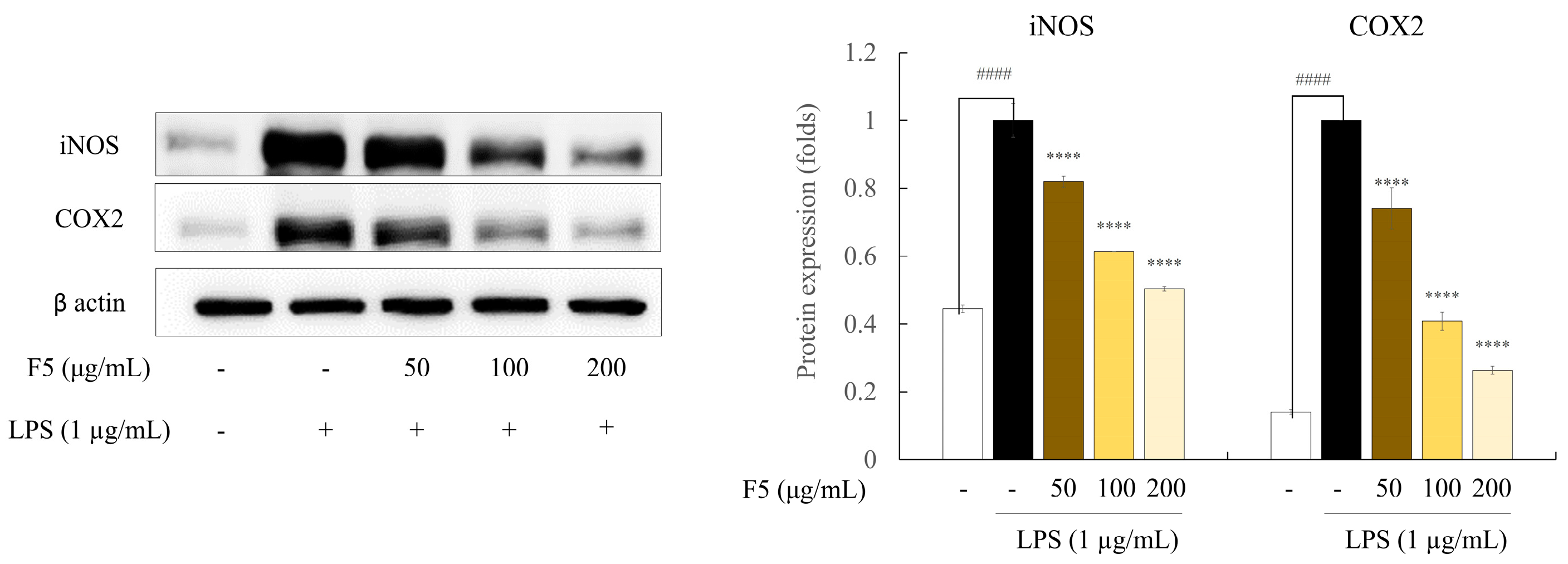
2.7. F5 Inhibits the Production of iNOS and COX-2 in LPS-induced RAW 264.7 Cells
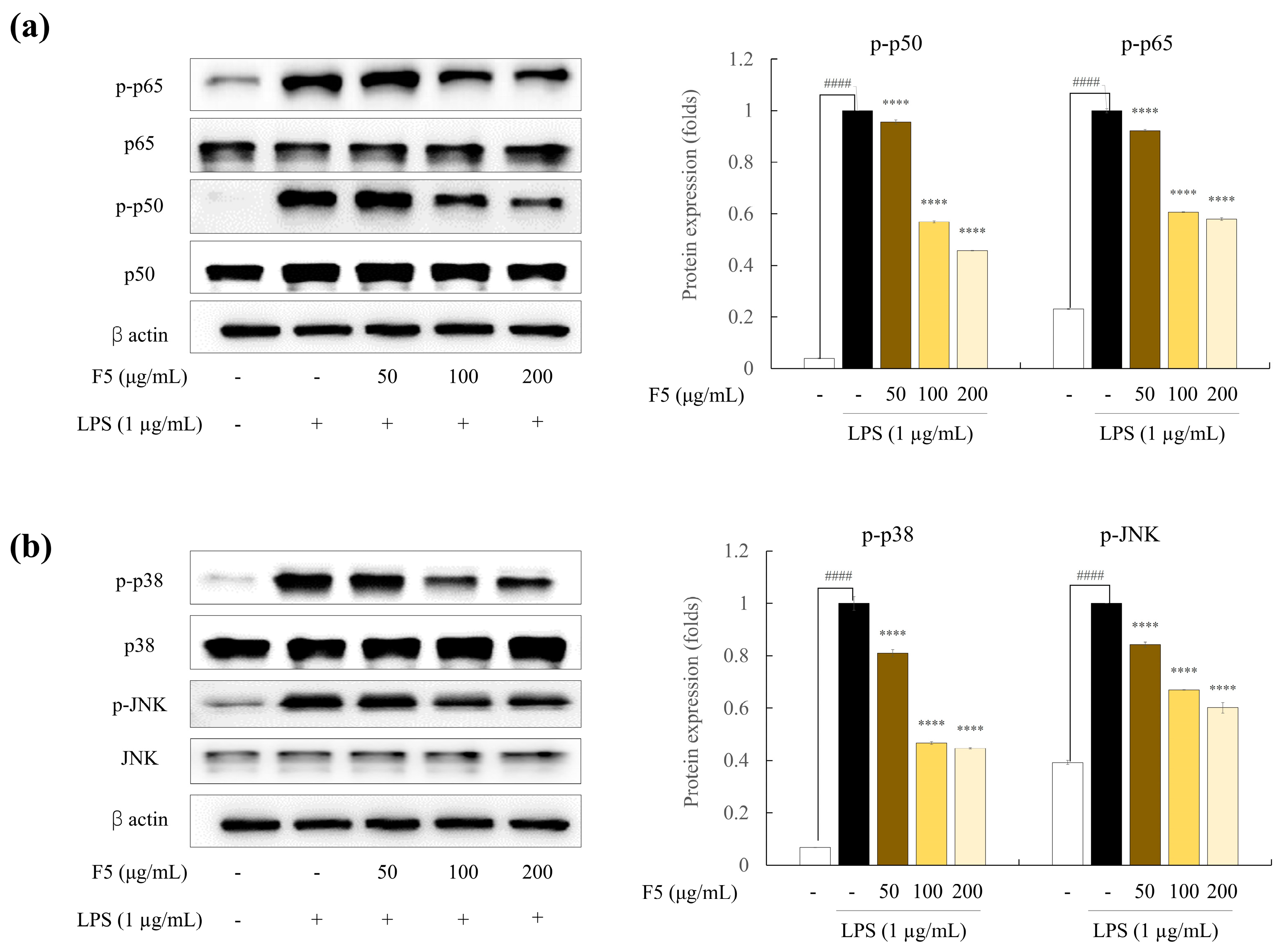
2.8. F5 downregulates the Over-Expression of NF-κB and MAPK Pathway Proteins
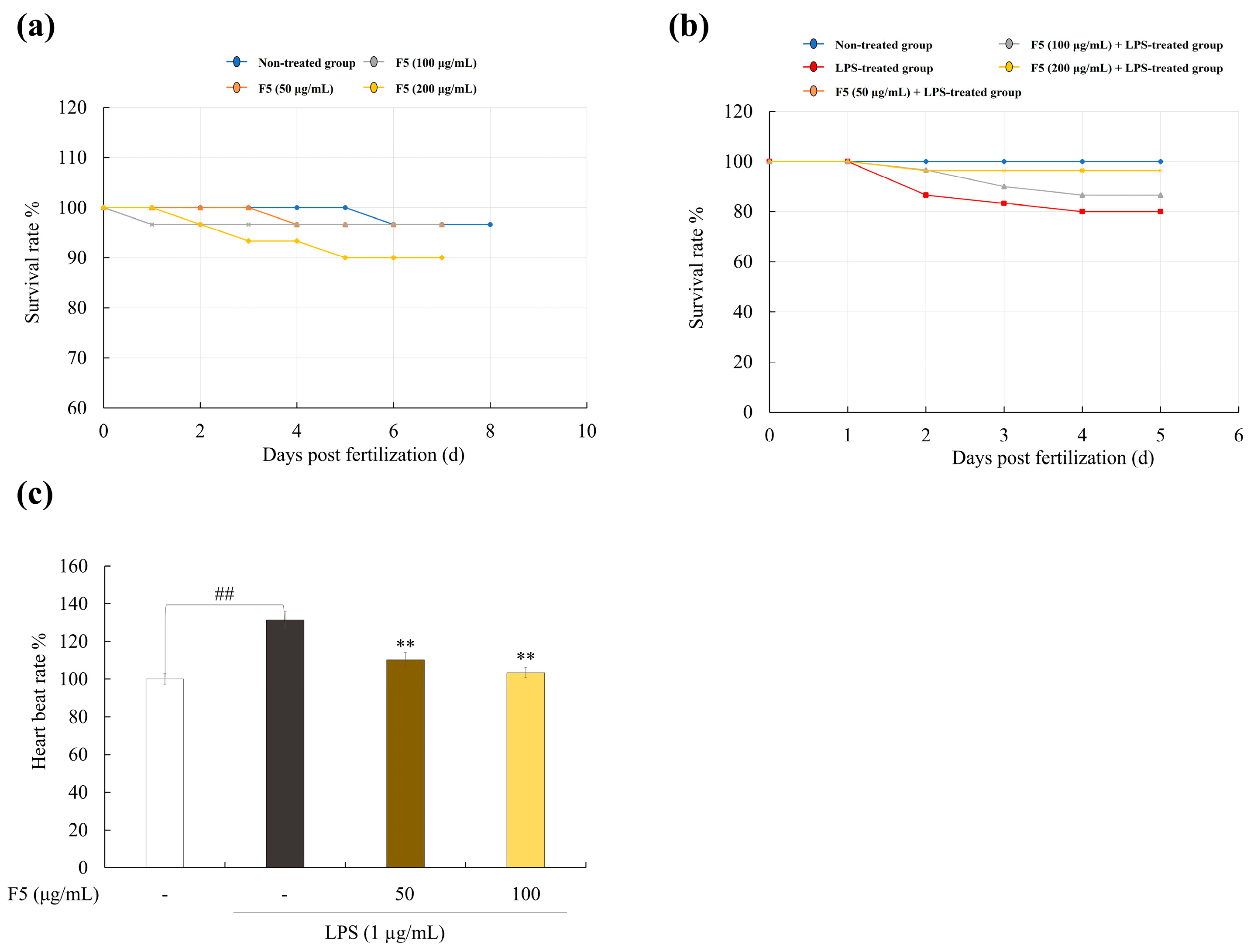
2.9. F5 Protects Zebrafish Larvae from LPS-Induced Toxicity
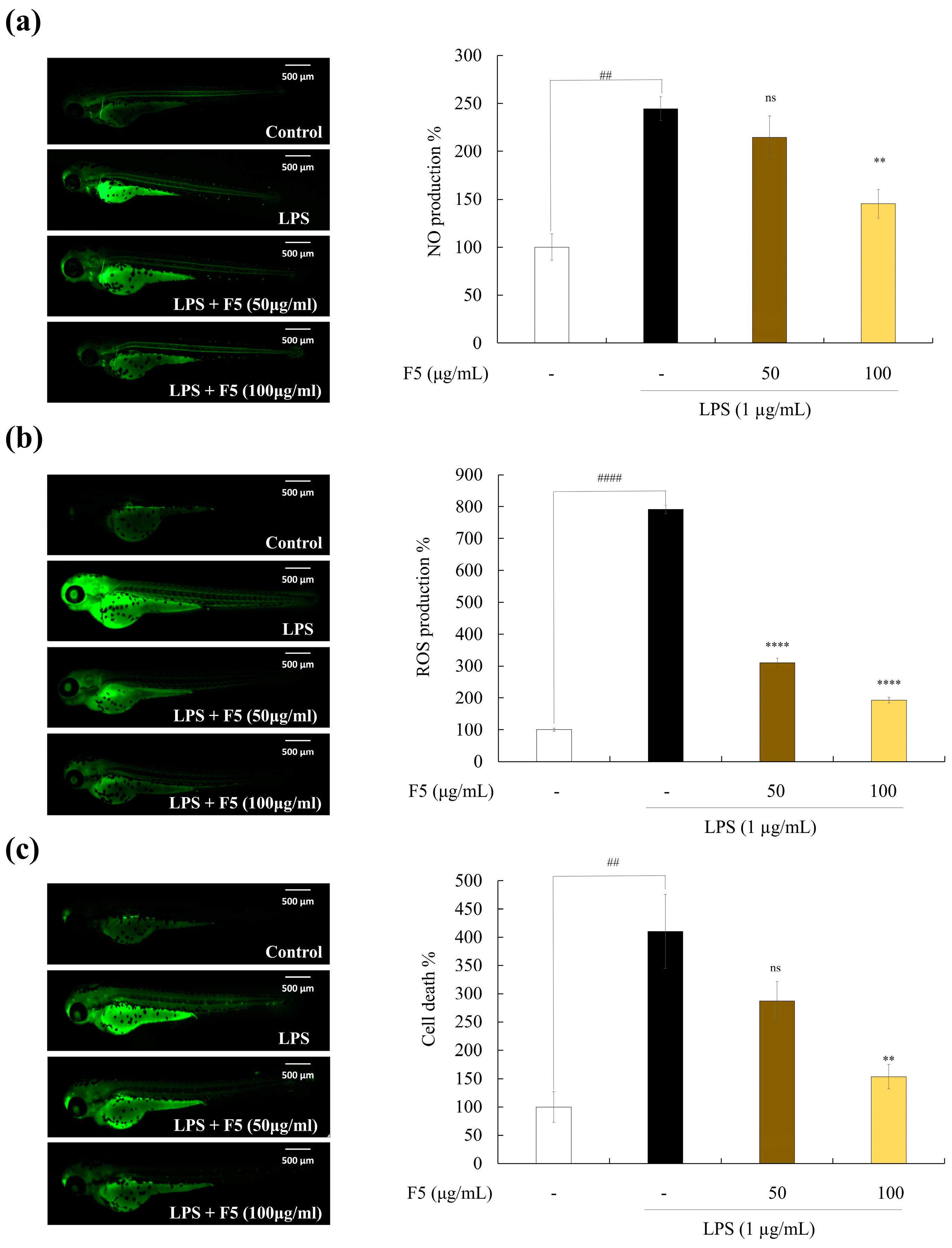
2.10. F5 Protects Zebrafish against LPS-induced Cell Death, NO, and ROS Production
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Sample Collection, Enzyme-Assisted Extraction, and Fucoidan Purification
4.3. Chemical Composition Analysis of Purified Fucoidan Fractions
4.4. Selection of Fucoidan Fraction
4.5. Monosaccharide Compositions Analysis
4.6. Evaluation of Surface Morphology
4.7. Fourier Transform Infrared (FTIR) Analysis
4.8. Determination of PGE2 and Pro-Inflammatory Cytokine Production
4.9. Western Blot Analysis
4.10. In Vivo Evaluation of F5 Anti-Inflammatory Activity in a Zebrafish Model
4.10.1. Zebrafish Maintenance
4.10.2. Survival Rate and Heart Rate Analysis in LPS-induced Zebrafish
4.10.3. Cell Death, ROS, and NO Production Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, H.-G.; Je, J.-G.; Hwang, J.; Jayawardena, T.U.; Nagahawatta, D.; Lu, Y.A.; Kim, H.-S.; Kang, M.-C.; Lee, D.-S.; Jeon, Y.-J. Comparision of antioxidant and anti-inflammatory activities of enzyme assisted hydrolysate from Ecklonia maxima blades and stipe. Fish. Aquat. Sci. 2021, 24, 197–206. [Google Scholar] [CrossRef]
- Taniguchi, K.; Karin, M. NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef]
- Hansson, G.K. Inflammation, atherosclerosis, and coronary artery disease. N. Engl. J. Med. 2005, 352, 1685–1695. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N. Technology, Bioactive potential and possible health effects of edible brown seaweeds. Trends Food Sci. Technol. 2011, 22, 315–326. [Google Scholar] [CrossRef]
- Ghosh, S.; Sarkar, T.; Pati, S.; Kari, Z.A.; Edinur, H.A.; Chakraborty, R. Novel bioactive compounds from marine sources as a tool for functional food development. Front. Mar. Sci. 2022, 9, 832957. [Google Scholar] [CrossRef]
- Liyanage, N.; Nagahawatta, D.; Jayawardena, T.U.; Jeon, Y.-J. The Role of Seaweed Polysaccharides in Gastrointestinal Health: Protective Effect against Inflammatory Bowel Disease. Life 2023, 13, 1026. [Google Scholar] [CrossRef] [PubMed]
- Nayak, G.; Bhuyan, R.; Sahu, A. Review on biomedical applications of marine algae-derived biomaterials. Univers. J. Public Health 2022, 10, 15–24. [Google Scholar] [CrossRef]
- Jayawardhana, H.; Lee, H.-G.; Liyanage, N.; Nagahawatta, D.; Ryu, B.; Jeon, Y.-J. Structural characterization and anti-inflammatory potential of sulfated polysaccharides from Scytosiphon lomentaria; attenuate inflammatory signaling pathways. J. Funct. Foods 2023, 102, 105446. [Google Scholar] [CrossRef]
- Liyanage, N.M.; Kim, Y.-S.; Nagahawatta, D.P.; Jin, H.; Yang, H.-W.; Jayawardhana, H.H.A.C.K.; Jayawardena, T.U.; Jeon, Y.-J. Sargassum horneri as a Prebiotic Dietary Supplement for Immunity Development in Streptococcus parauberis Infected Zebrafish Model. Front. Mar. Sci. 2022, 9, 901676. [Google Scholar] [CrossRef]
- Nagahawatta, D.; Liyanage, N.; Jayawardhana, H.; Lee, H.-G.; Jayawardena, T.U.; Jeon, Y.-J. Anti-Fine Dust Effect of Fucoidan Extracted from Ecklonia maxima Leaves in Macrophages via Inhibiting Inflammatory Signaling Pathways. Mar. Drugs 2022, 20, 413. [Google Scholar] [CrossRef]
- Nagahawatta, D.; Liyanage, N.; Je, J.-G.; Jayawardhana, H.; Jayawardena, T.U.; Jeong, S.-H.; Kwon, H.-J.; Choi, C.S.; Jeon, Y.-J. Polyphenolic compounds isolated from marine algae attenuate the replication of SARS-CoV-2 in the host cell through a multi-target approach of 3CLpro and PLpro. Mar. Drugs 2022, 20, 786. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of novel extraction technologies for bioactives from marine algae. J. Agric. Food Chem. 2013, 61, 4667–4675. [Google Scholar] [CrossRef]
- Park, M.H.; Han, J.-S. Padina arborescens extract protects high glucose-induced apoptosis in pancreatic β cells by reducing oxidative stress. Nutr. Res. Pract. 2014, 8, 494–500. [Google Scholar] [CrossRef]
- Park, M.H.; Han, J.S. Hypoglycemic effect of Padina arborescens extract in streptozotocin-induced diabetic mice. Prev. Nutr. Food Sci. 2012, 17, 239. [Google Scholar] [CrossRef]
- Reza, A.A.; Rashid, M.M.; Islam, M.S.; Hossen, M.A.; Ahmed, A.A.; Haque, M.A.; Nasrin, M.S.; Uddin, M.N.; Khan, J.; Rahman, M.A. Polyphenolics of purple devil fruits potentiate anti-inflammatory action by regulating the expression of inflammatory cytokines and apoptotic signaling molecules evident in extensive and combined experimental models. J. Funct. Foods 2023, 106, 105610. [Google Scholar] [CrossRef]
- Schulze-Osthoff, K.; Ferrari, D.; Riehemann, K.; Wesselborg, S. Regulation of NF-κB activation by MAP kinase cascades. Immunobiology 1997, 198, 35–49. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive proteins, peptides, and amino acids from macroalgae 1. J. Phycol. 2011, 47, 218–232. [Google Scholar] [CrossRef] [PubMed]
- del Pilar Sánchez-Camargo, A.; Montero, L.; Stiger-Pouvreau, V.; Tanniou, A.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Considerations on the use of enzyme-assisted extraction in combination with pressurized liquids to recover bioactive compounds from algae. Food Chem. 2016, 192, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.U.; Fernando, I.S.; Lee, W.W.; Sanjeewa, K.A.; Kim, H.-S.; Lee, D.-S.; Jeon, Y.-J. Isolation and purification of fucoidan fraction in Turbinaria ornata from the Maldives; Inflammation inhibitory potential under LPS stimulated conditions in in-vitro and in-vivo models. Int. J. Biol. Macromol. 2019, 131, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Liyanage, N.; Lee, H.-G.; Nagahawatta, D.; Jayawardhana, H.; Ryu, B.; Jeon, Y.-J. Characterization and therapeutic effect of Sargassum coreanum fucoidan that inhibits lipopolysaccharide-induced inflammation in RAW 264.7 macrophages by blocking NF-κB signaling. Int. J. Biol. Macromol. 2022, 223, 500–510. [Google Scholar] [CrossRef]
- Jayawardena, T.U.; Sanjeewa, K.A.; Nagahawatta, D.; Lee, H.-G.; Lu, Y.-A.; Vaas, A.; Abeytunga, D.; Nanayakkara, C.; Lee, D.-S.; Jeon, Y.-J. Anti-inflammatory effects of sulfated polysaccharide from Sargassum swartzii in macrophages via blocking TLR/NF-Κb signal transduction. Int. J. Biol. Macromol. 2020, 18, 601. [Google Scholar] [CrossRef]
- Wang, S.-H.; Huang, C.-Y.; Chen, C.-Y.; Chang, C.-C.; Huang, C.-Y.; Dong, C.-D.; Chang, J.-S. Structure and biological activity analysis of fucoidan isolated from Sargassum siliquosum. ACS Omega 2020, 5, 32447–32455. [Google Scholar] [CrossRef]
- Cheng, J.-J.; Chao, C.-H.; Chang, P.-C.; Lu, M.-K. Studies on anti-inflammatory activity of sulfated polysaccharides from cultivated fungi Antrodia cinnamomea. Food Hydrocoll. 2016, 53, 37–45. [Google Scholar] [CrossRef]
- Jiang, Z.; Okimura, T.; Yokose, T.; Yamasaki, Y.; Yamaguchi, K.; Oda, T. Effects of sulfated fucan, ascophyllan, from the brown Alga Ascophyllum nodosum on various cell lines: A comparative study on ascophyllan and fucoidan. J. Biosci. Bioeng. 2010, 110, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-G.; Jayawardena, T.U.; Song, K.-M.; Choi, Y.-S.; Jeon, Y.-J.; Kang, M.-C. Dietary fucoidan from a brown marine algae (Ecklonia cava) attenuates lipid accumulation in differentiated 3T3-L1 cells and alleviates high-fat diet-induced obesity in mice. Food Chem. Toxicol. 2022, 162, 112862. [Google Scholar] [CrossRef] [PubMed]
- Nagahawatta, D.; Lee, H.-G.; Liyanage, N.; Jayawardhana, H.; Wang, L.; Kim, H.-S.; Jeon, Y.-J. Alginic acid, a functional dietary ingredient derived from Ecklonia maxima stipe, attenuates the pro-inflammatory responses on particulate matter-induced lung macrophages. J. Funct. Foods 2023, 104, 105539. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Hwang, J.; Lee, S.G.; Jo, H.Y.; Oh, M.J.; Liyanage, N.; Je, J.-G.; An, H.J.; Jeon, Y.-J. Function, Structural characteristics of sulfated polysaccharides from Sargassum horneri and immune-enhancing activity of polysaccharides combined with lactic acid bacteria. Food Funct. 2022, 13, 8214–8227. [Google Scholar] [CrossRef]
- Liyanage, N.; Nagahawatta, D.; Jayawardhana, H.; Jayawardena, T.U.; Kim, Y.-S.; Lee, H.-G.; Park, Y.-J.; Jeon, Y.-J. Therapeutic effect of Sargassum swartzii against urban particulate matter–induced lung inflammation via toll-like receptor-mediated NF-κB pathway inhibition. Algal Res. 2023, 71, 103045. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Fernando, I.P.S.; Kim, E.-A.; Ahn, G.; Jee, Y.; Jeon, Y.-J. Anti-inflammatory activity of a sulfated polysaccharide isolated from an enzymatic digest of brown seaweed Sargassum horneri in RAW 264.7 cells. Nutr. Res. Pract. 2017, 11, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Clancy, R.M.; Abramson, S.B. Medicine, Nitric oxide: A novel mediator of inflammation. Exp. Biol. Med. 1995, 210, 93–101. [Google Scholar] [CrossRef]
- Lin, D.; Xu, X.; Chen, L.; Chen, L.; Deng, M.; Chen, J.; Ren, Z.; Lei, L.; Wang, J.; Deng, J.; et al. Supramolecular nanofiber of indomethacin derivative confers highly cyclooxygenase-2 (COX-2) selectivity and boosts anti-inflammatory efficacy. J. Control. Release 2023, 364, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.; Jayawardena, T.U.; Kim, H.-S.; Kim, S.-Y.; Ahn, G.; Kim, H.-J.; Fu, X.; Jee, Y.; Jeon, Y.-J. Ethanol extract separated from Sargassum horneri (Turner) abate LPS-induced inflammation in RAW 264.7 macrophages. Fish. Aquat. Sci. 2019, 22, 1–10. [Google Scholar] [CrossRef]
- Sangiuliano, B.; Pérez, N.M.; Moreira, D.F.; Belizário, J.E. Cell death-associated molecular-pattern molecules: Inflammatory signaling and control. Mediat. Inflamm. 2014, 2014. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of the Association of Official Analytical Chemists International; Association of Official Analytical Chemists: Washington, DC, USA, 1995; Volume 11, pp. 6–7. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Waterborg, J.H. The Lowry method for protein quantitation. In The Protein Protocols Handbook; Humana Press: Totowa, NJ, USA, 2009; pp. 7–10. [Google Scholar]
- Wang, L.; Je, J.-G.; Huang, C.; Oh, J.-Y.; Fu, X.; Wang, K.; Ahn, G.; Xu, J.; Gao, X.; Jeon, Y.-J. Anti-inflammatory effect of sulfated polysaccharides isolated from Codium fragile in vitro in RAW 264.7 macrophages and in vivo in zebrafish. Mar. Drugs 2022, 20, 391. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximate Compositions (%) | ||||
|---|---|---|---|---|
| Samples | Polysaccharide | Protein | Polyphenol | Sulfate |
| PAC | 38.93 ± 0.31 | 13.33 ± 1.62 | 11.50 ± 0.61 | 5.44 ± 0.54 |
| PACC | 74.80 ± 0.43 | 10.51 ± 1.22 | 7.71 ± 0.04 | 3.31 ±1.14 |
| F1 | 74.55 ± 1.01 | 4.01 ± 0.04 | 3.33 ± 1.82 | 11.39 ± 1.00 |
| F2 | 71.66 ± 0.56 | 3.70 ± 0.62 | 2,51 ± 0.55 | 16.77 ± 1.81 |
| F3 | 65.71 ± 0.94 | 2.04 ± 0.48 | 2.01 ±1.00 | 23.44 ± 0.95 |
| F4 | 58.44 ± 1.21 | 1.98 ± 0.82 | 2.51 ± 1.11 | 29.48 ± 1.12 |
| F5 | 51.54 ± 0.33 | 1.44 ± 0.96 | 1.92 ± 1.03 | 38.74 ± 1.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-G.; Liyanage, N.M.; Yang, F.; Kim, Y.-S.; Lee, S.-H.; Ko, S.-C.; Yang, H.-W.; Jeon, Y.-J. Investigation of Physical Characteristics and In Vitro Anti-Inflammatory Effects of Fucoidan from Padina arborescens: A Comprehensive Assessment against Lipopolysaccharide-Induced Inflammation. Mar. Drugs 2024, 22, 109. https://doi.org/10.3390/md22030109
Lee H-G, Liyanage NM, Yang F, Kim Y-S, Lee S-H, Ko S-C, Yang H-W, Jeon Y-J. Investigation of Physical Characteristics and In Vitro Anti-Inflammatory Effects of Fucoidan from Padina arborescens: A Comprehensive Assessment against Lipopolysaccharide-Induced Inflammation. Marine Drugs. 2024; 22(3):109. https://doi.org/10.3390/md22030109
Chicago/Turabian StyleLee, Hyo-Geun, N. M. Liyanage, Fengqi Yang, Young-Sang Kim, Seung-Hong Lee, Seok-Chun Ko, Hye-Won Yang, and You-Jin Jeon. 2024. "Investigation of Physical Characteristics and In Vitro Anti-Inflammatory Effects of Fucoidan from Padina arborescens: A Comprehensive Assessment against Lipopolysaccharide-Induced Inflammation" Marine Drugs 22, no. 3: 109. https://doi.org/10.3390/md22030109
APA StyleLee, H.-G., Liyanage, N. M., Yang, F., Kim, Y.-S., Lee, S.-H., Ko, S.-C., Yang, H.-W., & Jeon, Y.-J. (2024). Investigation of Physical Characteristics and In Vitro Anti-Inflammatory Effects of Fucoidan from Padina arborescens: A Comprehensive Assessment against Lipopolysaccharide-Induced Inflammation. Marine Drugs, 22(3), 109. https://doi.org/10.3390/md22030109