
Sea Urchin Food Waste into Bioactives: Collagen and Polyhydroxynaphtoquinones from P. lividus and S. granularis

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. PHNQs
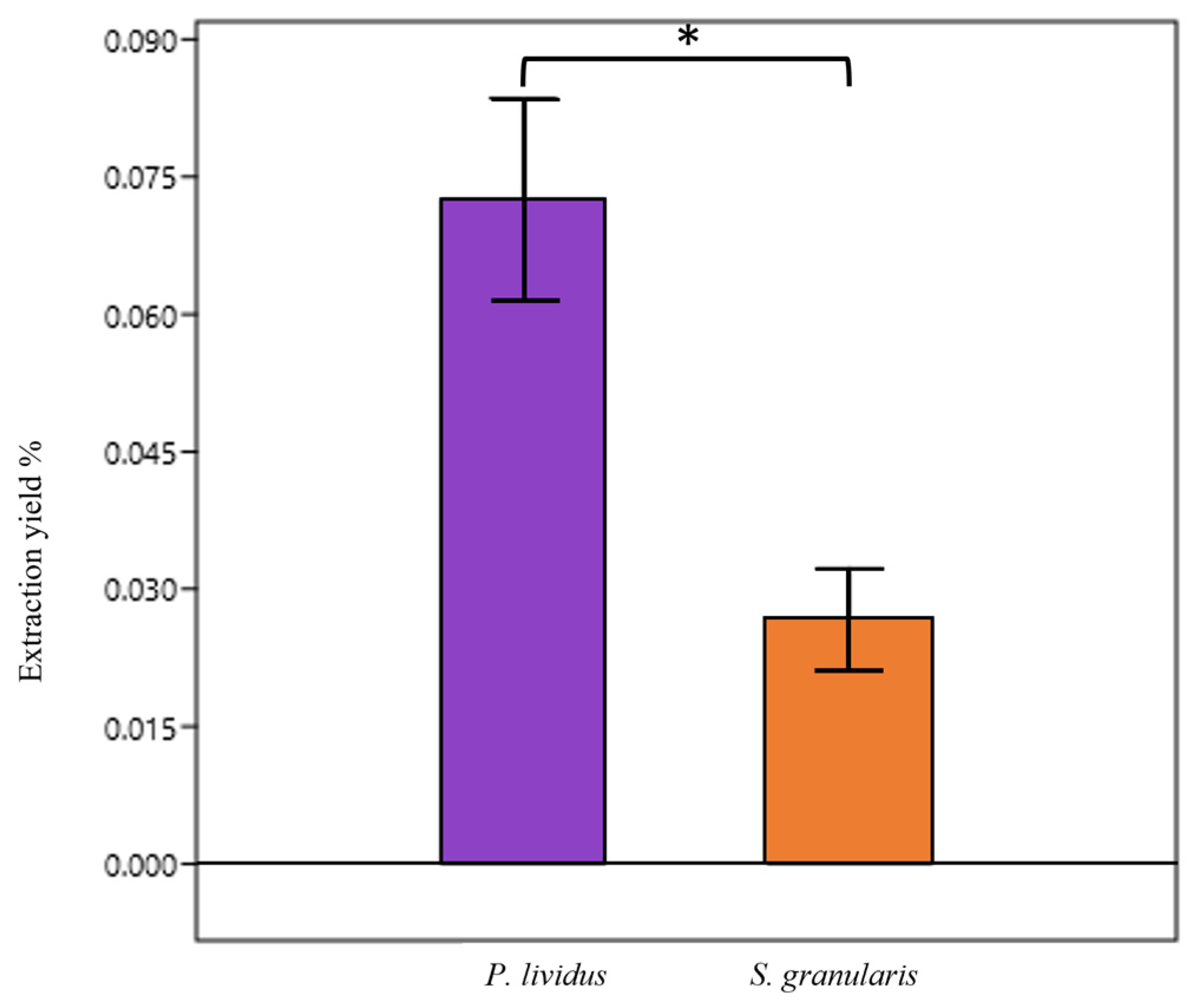
2.1.1. PHNQ Extraction
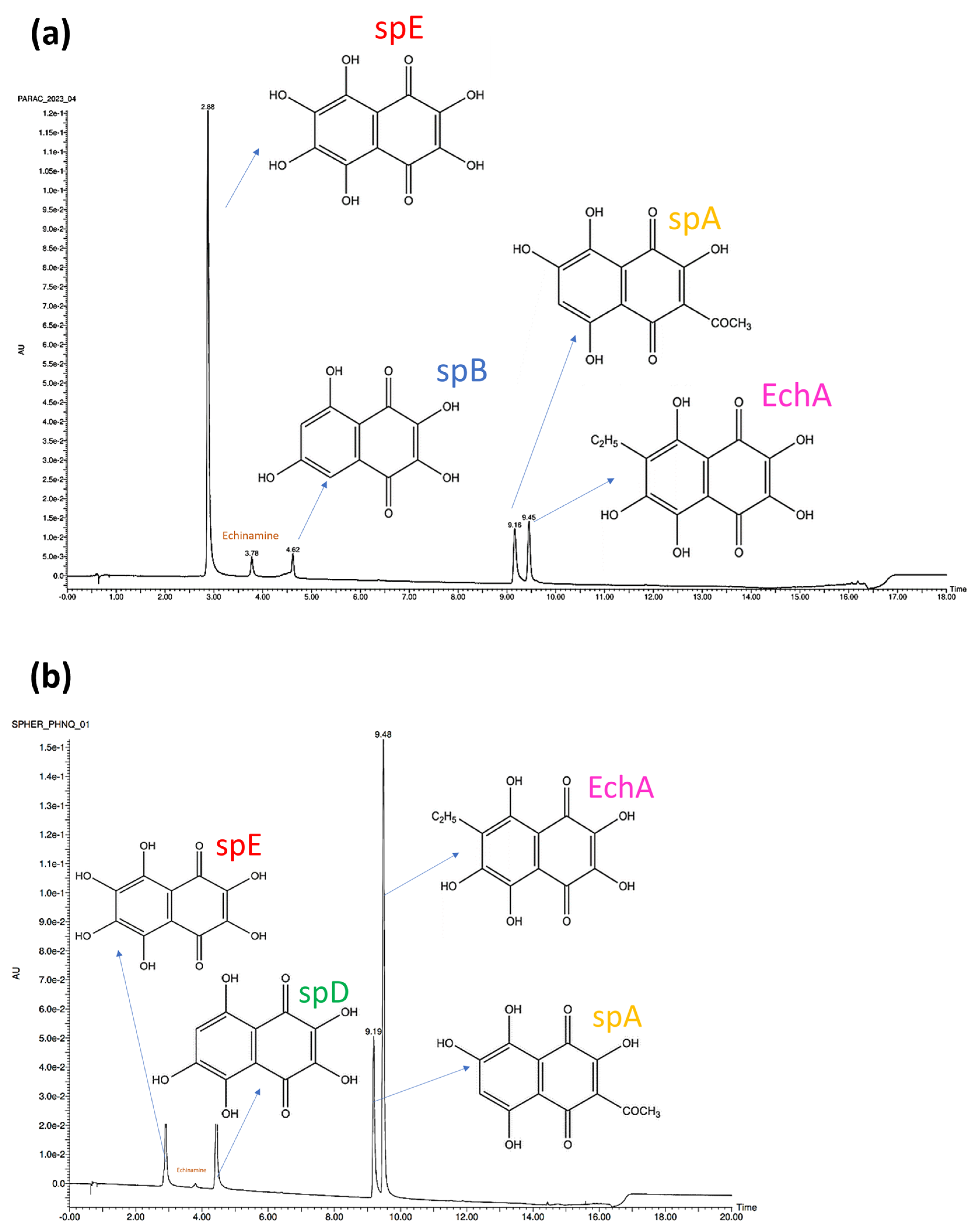
2.1.2. PHNQ Identification and Quantification
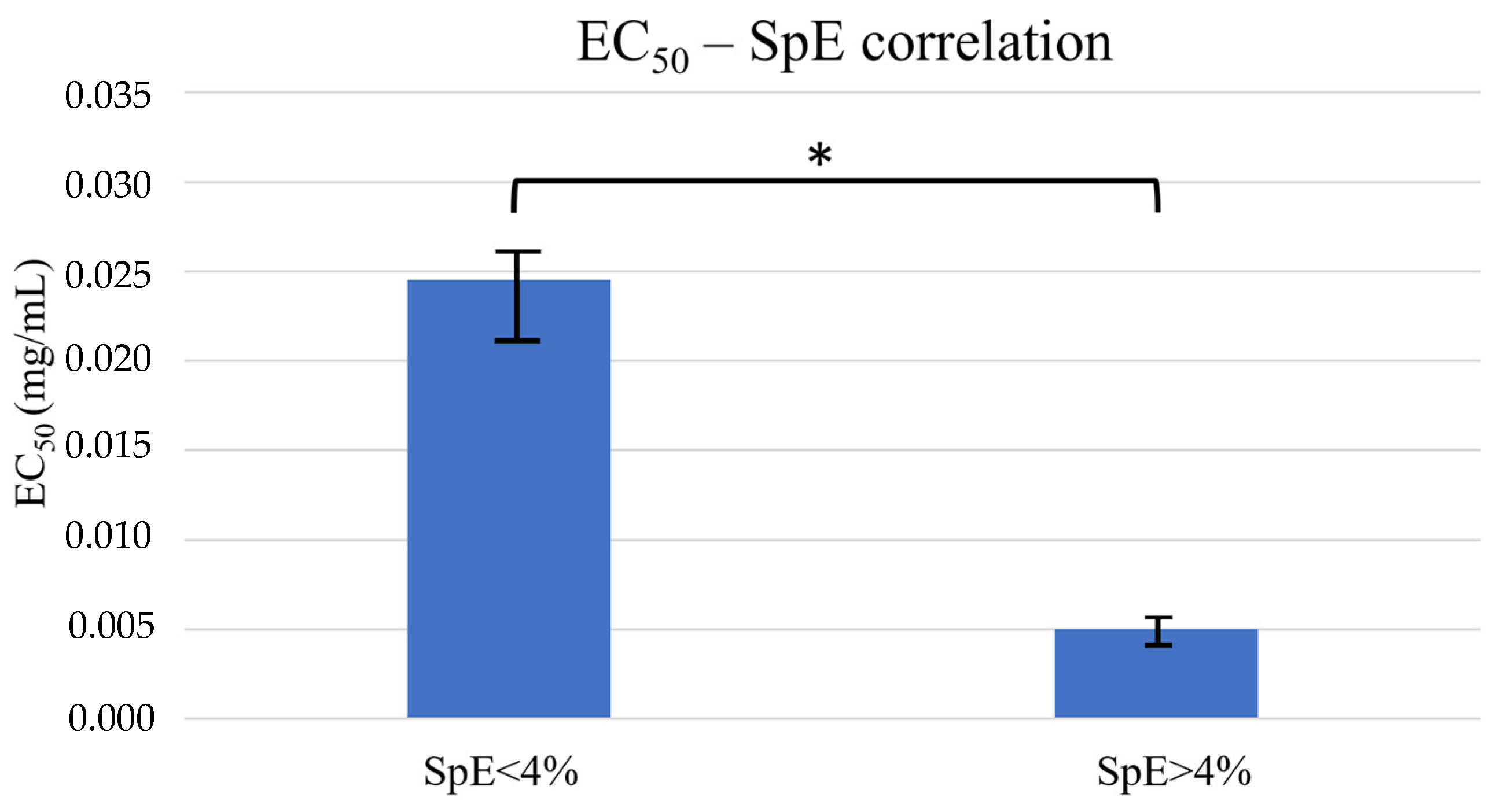
2.1.3. ABTS∙+ Assay
2.2. Collagen
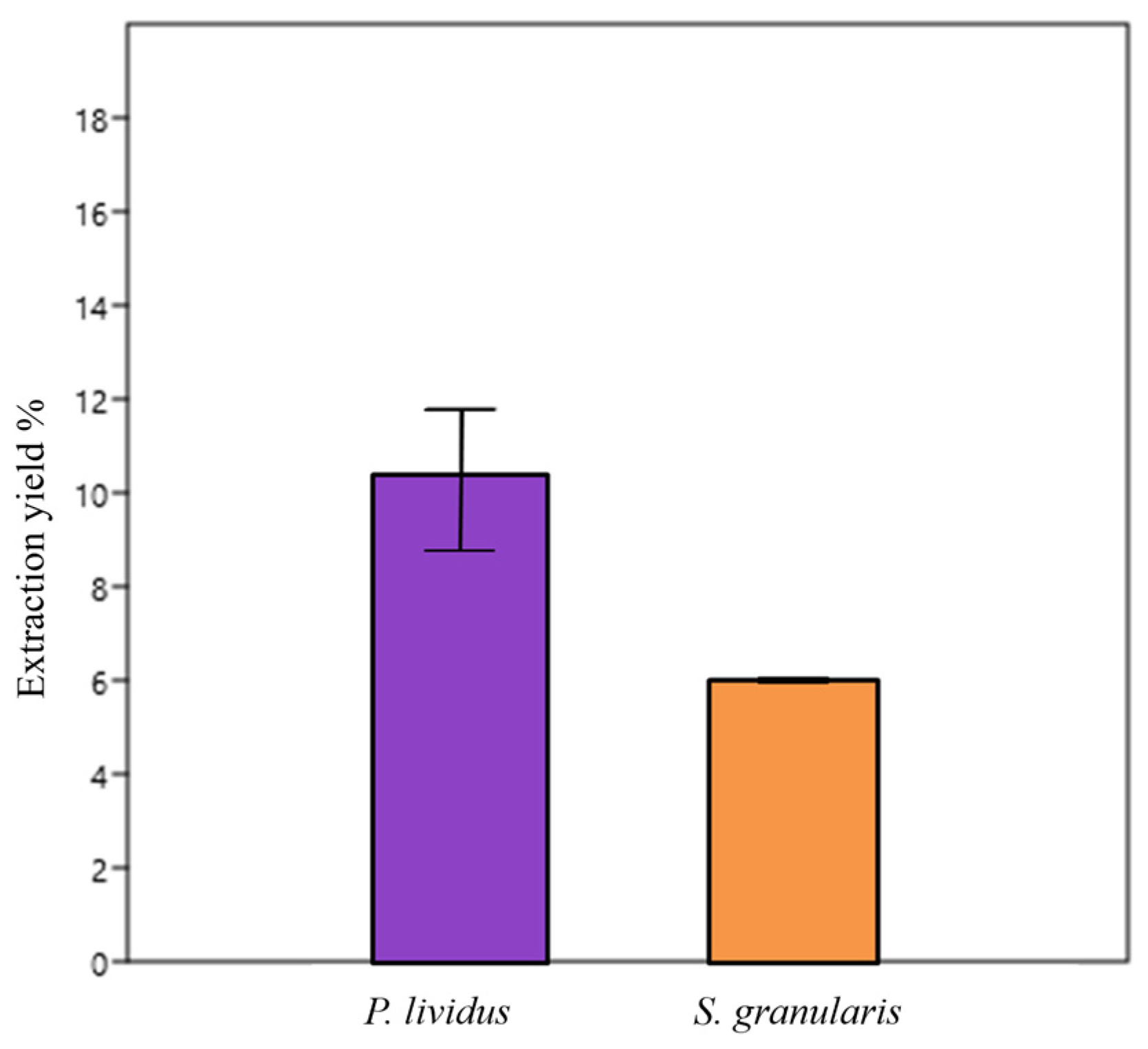
2.2.1. Collagen Extraction
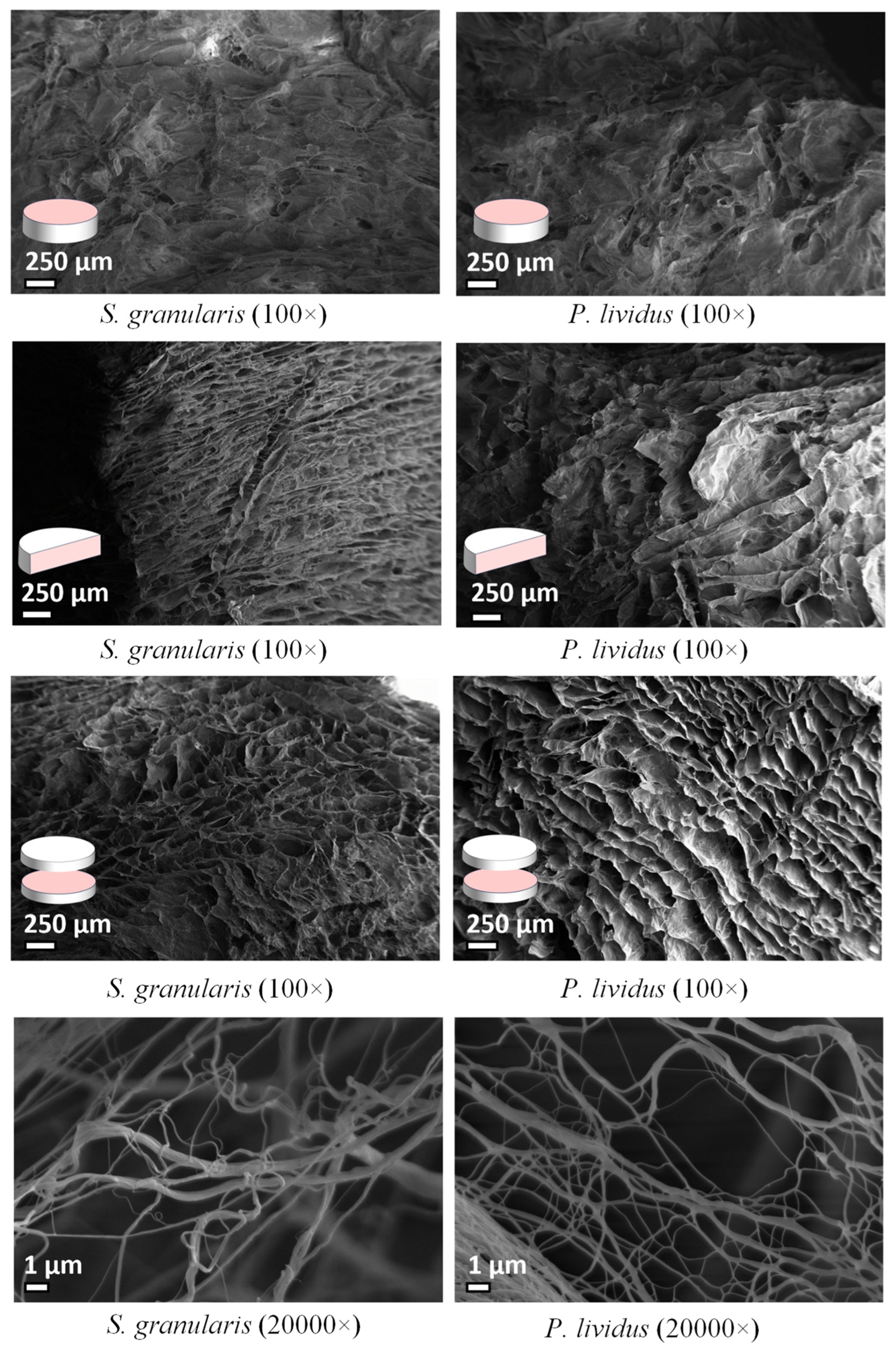
2.2.2. Ultrastructural Analysis
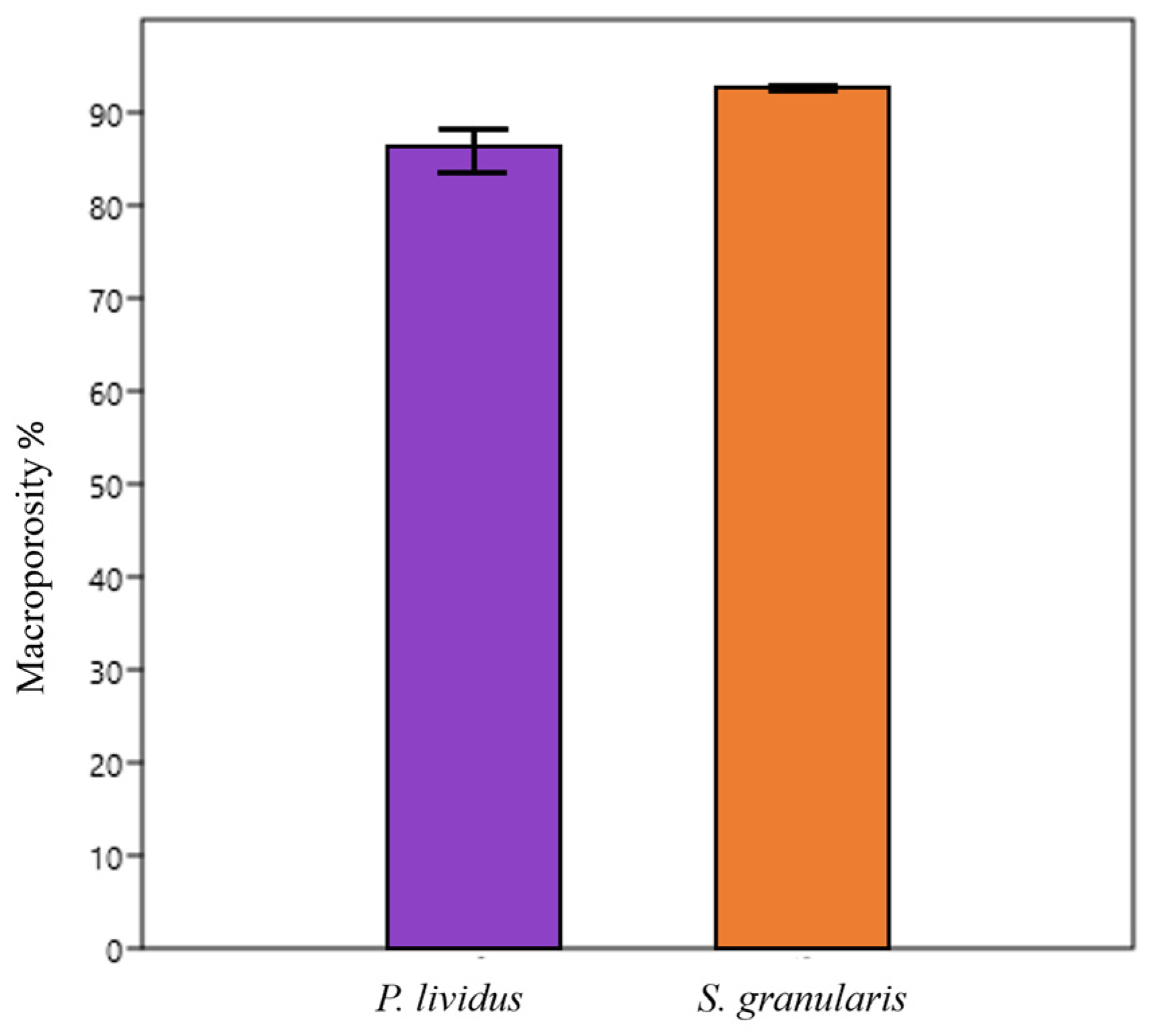
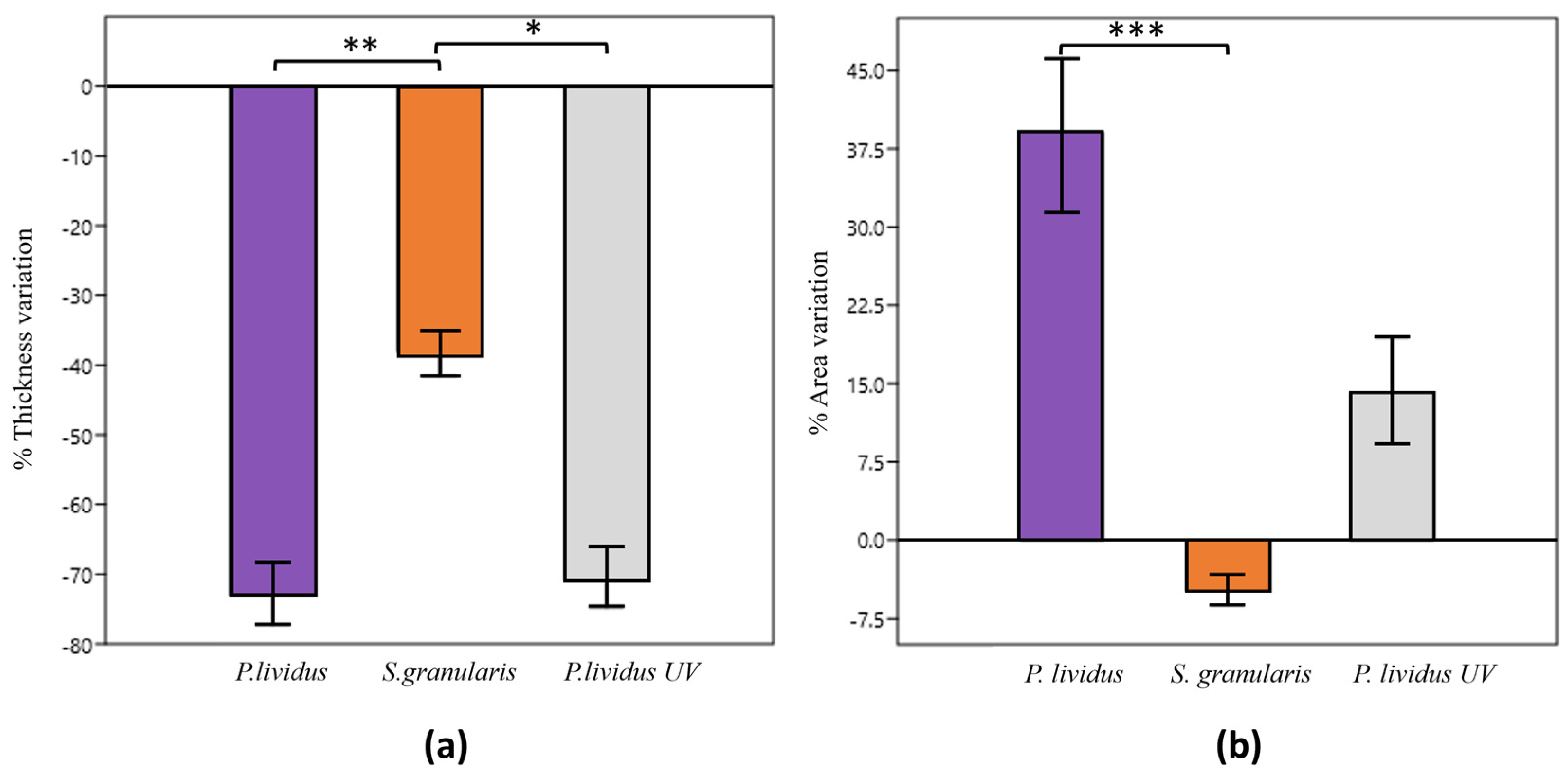
2.2.3. Macroporosity
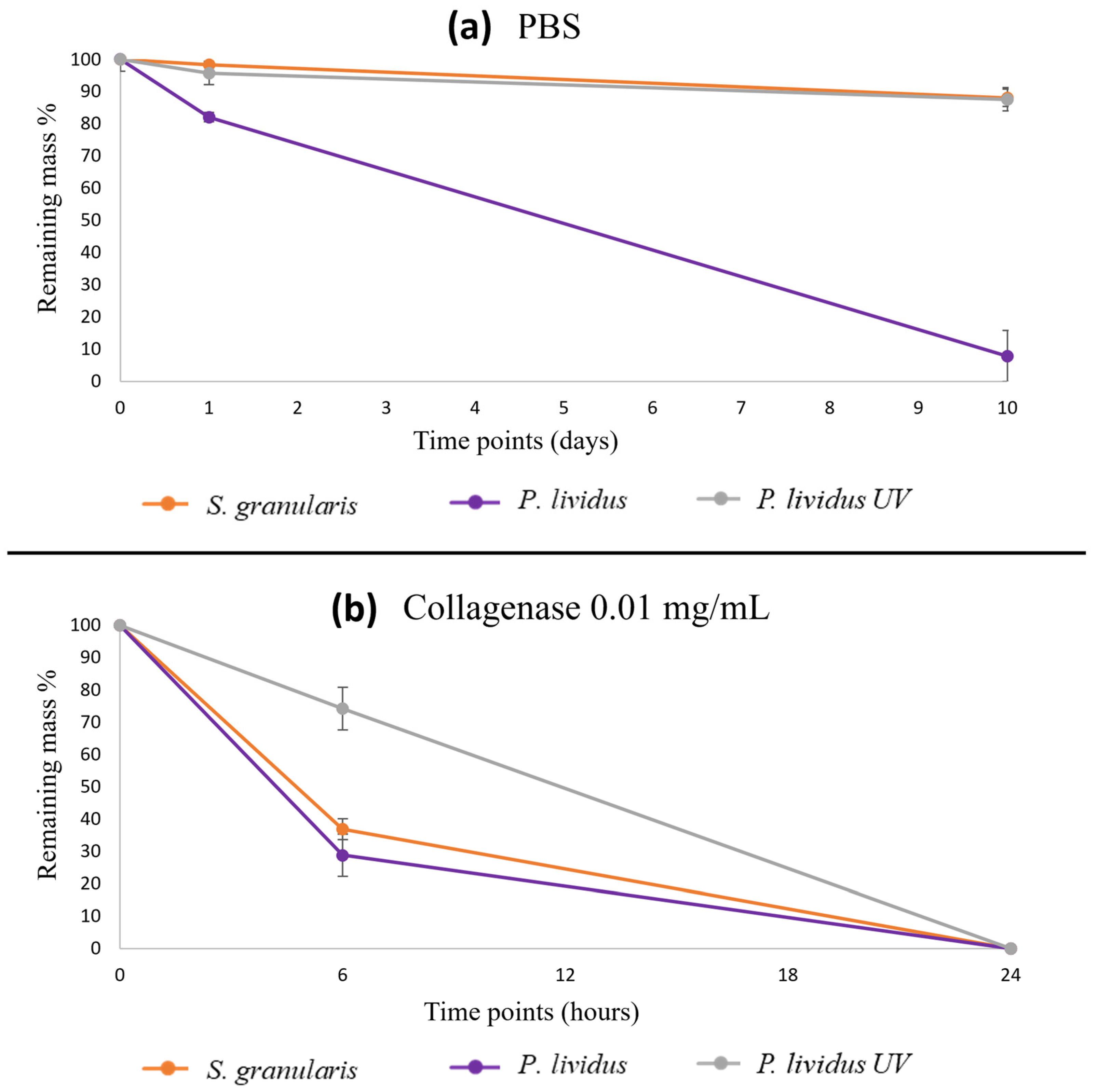
2.2.4. Degradation Tests
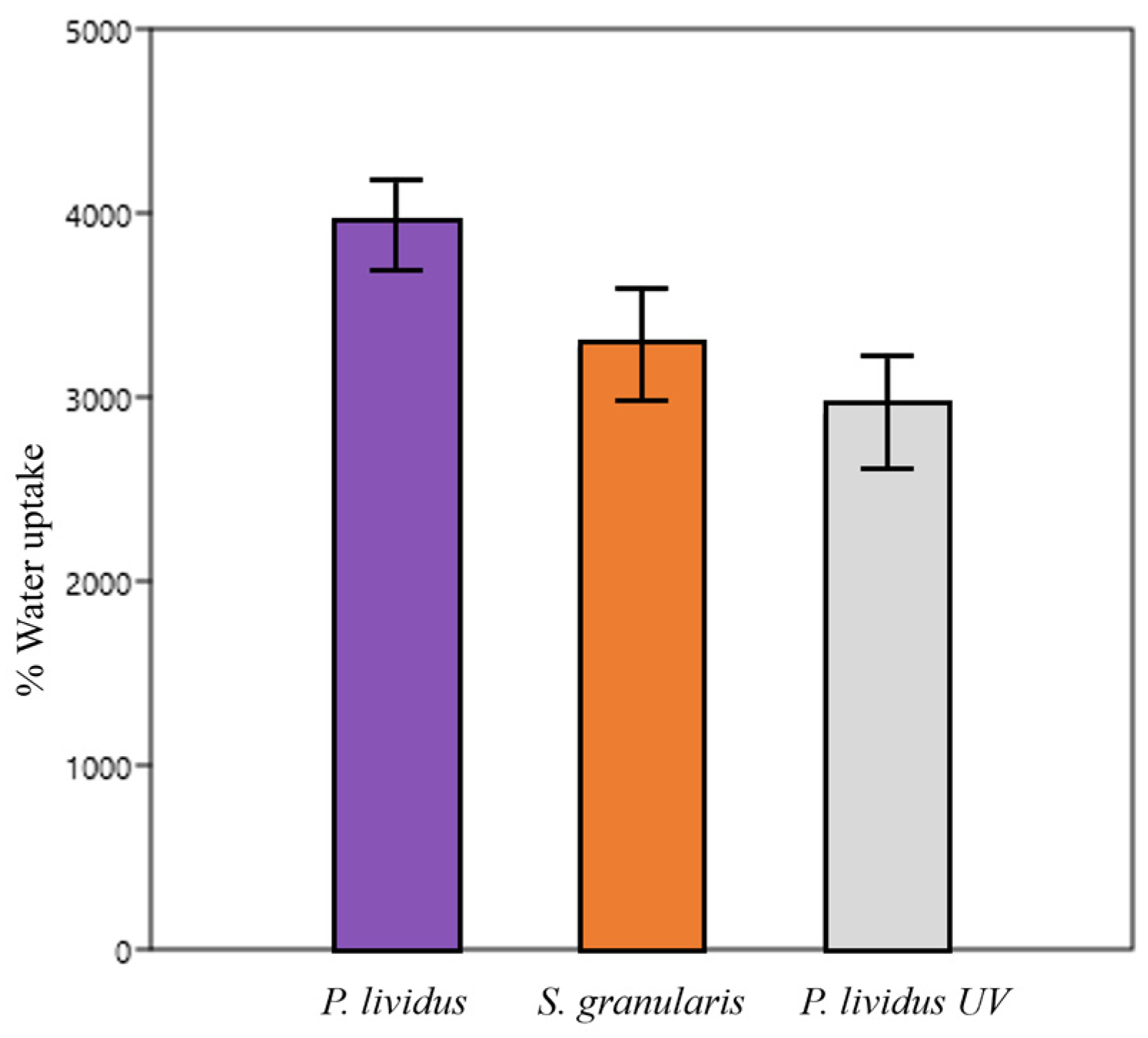
2.2.5. Hydrations Tests/Swelling Properties
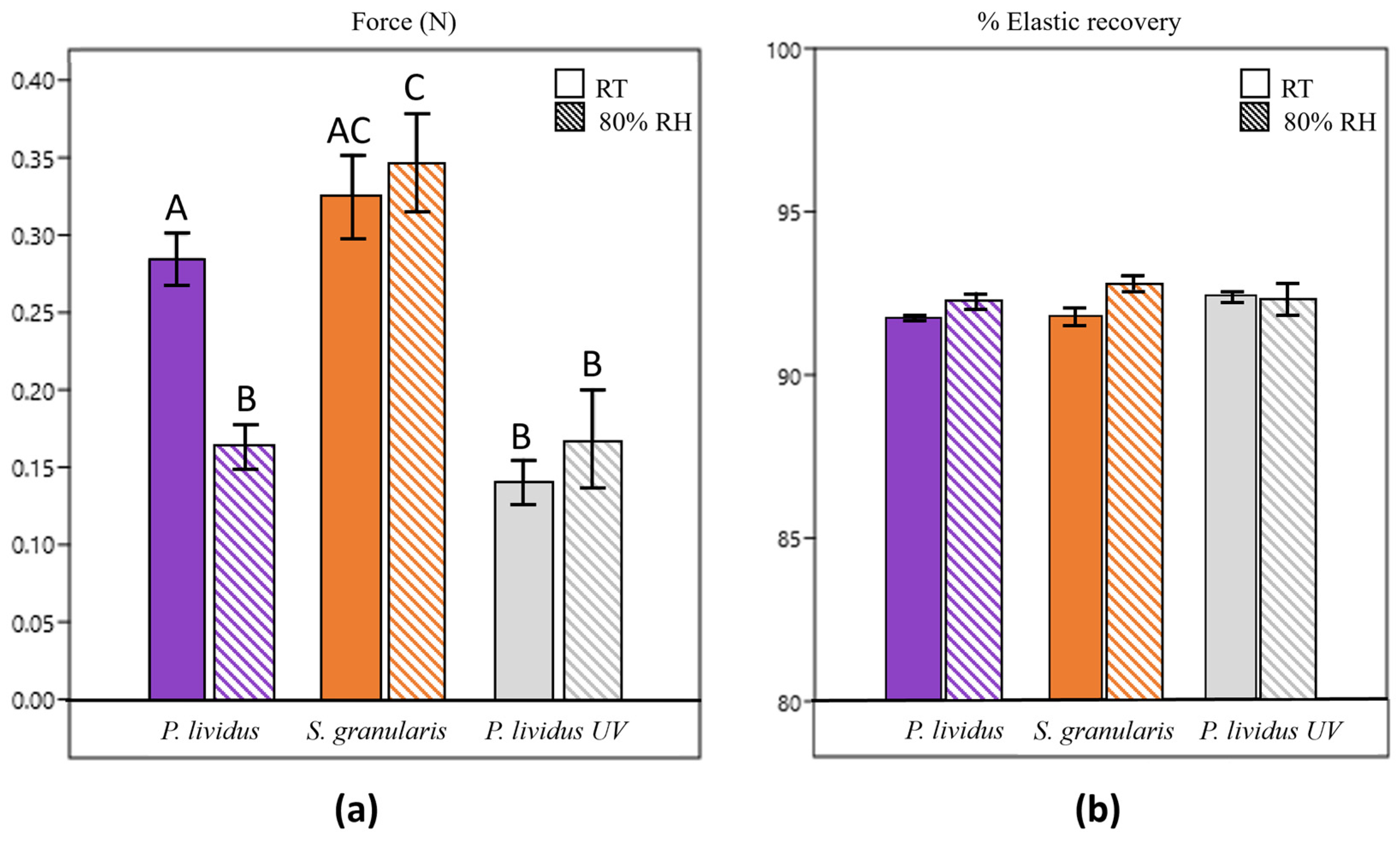
2.2.6. Mechanical Tests
3. Discussion
3.1. PHNQs
3.2. Collagen
4. Materials and Methods
4.1. Chemicals
4.2. PHNQs Extraction
4.3. Ultra-High-Pressure Liquid Chromatography–Mass Spectrometry
4.4. ABTS Assay
4.5. Collagen Extraction
4.6. Production of 3D Scaffolds
4.7. Ultrastructural Analysis: Scanning Electron Microscopy (SEM)
4.8. Macroporosity
4.9. Degradation Tests
4.10. Hydrations Tests
4.11. Mechanical Tests
4.12. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carboni, S.; Kelly, M.; Hughes, A.; Vignier, J.; Atack, T.; Migaud, H. Evaluation of Flow through Culture Technique for Commercial Production of Sea Urchin (Paracentrotus Lividus) Larvae. Aquac. Res. 2012, 45, 768–772. [Google Scholar] [CrossRef]
- Zilia, F.; Bacenetti, J.; Sugni, M.; Matarazzo, A.; Orsi, L. From Waste to Product: Circular Economy Applications from Sea Urchin. Sustainability 2021, 13, 5427. [Google Scholar] [CrossRef]
- Stefansson, G.; Kristinsson, H.; Ziemer, N.; Hannon, C.; James, P. Markets for Sea Urchins: A Review of Global Supply and Markets. Skýrsla Matís 2017, 45, 10–17. [Google Scholar]
- Melotti, L.; Venerando, A.; Zivelonghi, G.; Carolo, A.; Marzorati, S.; Martinelli, G.; Sugni, M.; Maccatrozzo, L.; Patruno, M. A Second Life for Seafood Waste: Therapeutical Promises of Polyhydroxynapthoquinones Extracted from Sea Urchin by-Products. Antioxidants 2023, 12, 1730. [Google Scholar] [CrossRef]
- Marzorati, S.; Martinelli, G.; Sugni, M.; Verotta, L. Green Extraction Strategies for Sea Urchin Waste Valorization. Front. Nutr. 2021, 8, 730747. [Google Scholar] [CrossRef]
- Hou, Y.A.; Vasileva, E.; Carne, A.; McConnell, M.; Bekhit, A.E.-D.A.; Mishchenko, N.P. Naphthoquinones of the Spinochrome Class: Occurrence, Isolation, Biosynthesis and Biomedical Applications. RSC Adv. 2018, 8, 32637–32650. [Google Scholar] [CrossRef]
- Vasileva, E.A.; Mishchenko, N.P.; Fedoreyev, S.A. Diversity of Polyhydroxynaphthoquinone Pigments in North Pacific Sea Urchins. Chem. Biodivers. 2017, 14, e1700182. [Google Scholar] [CrossRef]
- Brasseur, L.; Hennebert, E.; Fiévez, L.; Caulier, G.; Bureau, F.; Tafforeau, L.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. The Roles of Spinochromes in Four Shallow Water Tropical Sea Urchins and Their Potential as Bioactive Pharmacological Agents. Mar. Drugs 2017, 15, 179. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, S.; Yousefzadi, M.; Moein, S.; Rezadoost, H.; Bioki, N.A. Identification and Antioxidant of Polyhydroxylated Naphthoquinone Pigments from Sea Urchin Pigments of Echinometra Mathaei. Med. Chem. Res. 2016, 25, 1476–1483. [Google Scholar] [CrossRef]
- Powell, C.; Hughes, A.D.; Kelly, M.S.; Conner, S.; McDougall, G.J. Extraction and Identification of Antioxidant Polyhydroxynaphthoquinone Pigments from the Sea Urchin, Psammechinus Miliaris. LWT Food Sci. Technol. 2014, 59, 455–460. [Google Scholar] [CrossRef]
- Li, D.-M.; Zhou, D.-Y.; Zhu, B.-W.; Miao, L.; Qin, L.; Dong, X.-P.; Wang, X.-D.; Murata, Y. Extraction, Structural Characterization and Antioxidant Activity of Polyhydroxylated 1,4-Naphthoquinone Pigments from Spines of Sea Urchin Glyptocidaris Crenularis and Strongylocentrotus Intermedius. Eur. Food Res. Technol. 2013, 237, 331–339. [Google Scholar] [CrossRef]
- Zhou, D.-Y.; Qin, L.; Zhu, B.-W.; Wang, X.-D.; Tan, H.; Yang, J.-F.; Li, D.-M.; Dong, X.-P.; Wu, H.-T.; Sun, L.-M.; et al. Extraction and Antioxidant Property of Polyhydroxylated Naphthoquinone Pigments from Spines of Purple Sea Urchin Strongylocentrotus Nudus. Food Chem. 2011, 129, 1591–1597. [Google Scholar] [CrossRef]
- Kuwahara, R.; Hatate, H.; Yuki, T.; Murata, H.; Tanaka, R.; Hama, Y. Antioxidant Property of Polyhydroxylated Naphthoquinone Pigments from Shells of Purple Sea Urchin Anthocidaris Crassispina. LWT Food Sci. Technol. 2009, 42, 1296–1300. [Google Scholar] [CrossRef]
- Anderson, H.A.; Mathieson, J.W.; Thomson, R.H. Distribution of Spinochrome Pigments in Echinoids. Comp. Biochem. Physiol. 1969, 28, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Shikov, A.; Pozharitskaya, O.; Krishtopina, A.; Makarov, V. Naphthoquinone Pigments from Sea Urchins: Chemistry and Pharmacology. Phytochem. Rev. 2018, 17, 509–534. [Google Scholar] [CrossRef]
- Mischenko, N.P.; Fedoreyev, S.A.; Pokhilo, N.D.; Anufriev, V.P.; Denisenko, V.A.; Glazunov, V.P. Echinamines A and B, First Aminated Hydroxynaphthazarins from the Sea Urchin Scaphechinus Mirabilis. J. Nat. Prod. 2005, 68, 1390–1393. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, A.V.; Ivanova, M.V.; Levitsky, D.O. Echinochrome, a Naturally Occurring Iron Chelator and Free Radical Scavenger in Artificial and Natural Membrane Systems. Life Sci. 2005, 76, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Pozharitskaya, O.N.; Shikov, A.N.; Makarova, M.N.; Ivanova, S.A.; Kosman, V.M.; Makarov, V.G.; Bazgier, V.; Berka, K.; Otyepka, M.; Ulrichová, J. Antiallergic Effects of Pigments Isolated from Green Sea Urchin (Strongylocentrotus Droebachiensis) Shells. Planta Med. 2013, 79, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Katelnikova, A.E.; Kryshen, K.L.; Makarova, M.N.; Makarov, V.G.; Shikov, A.N. Mechanisms of Anti-Inflammatory Effect of Glycosylated Polypeptide Complex Extracted from Sea Urchin Strongylocentrotus Droebachiensis. Russ. J. Immunol. 2018, 21, 73–79. [Google Scholar] [CrossRef]
- Rubilar, T.; Barbieri, E.S.; Gazquez, A.; Avaro, M. Sea Urchin Pigments: Echinochrome A and Its Potential Implication in the Cytokine Storm Syndrome. Mar. Drugs 2021, 19, 267. [Google Scholar] [CrossRef]
- Shikov, A.N.; Ossipov, V.I.; Martiskainen, O.; Pozharitskaya, O.N.; Ivanova, S.A.; Makarov, V.G. The Offline Combination of Thin-Layer Chromatography and High-Performance Liquid Chromatography with Diode Array Detection and micrOTOF-Q Mass Spectrometry for the Separation and Identification of Spinochromes from Sea Urchin (Strongylocentrotus Droebachiensis) Shells. J. Chromatogr. A 2011, 1218, 9111–9114. [Google Scholar] [CrossRef] [PubMed]
- Di Benedetto, C.; Barbaglio, A.; Martinello, T.; Alongi, V.; Fassini, D.; Cullorà, E.; Patruno, M.; Bonasoro, F.; Barbosa, M.; Candia, D.; et al. Production, Characterization and Biocompatibility of Marine Collagen Matrices from an Alternative and Sustainable Source: The Sea Urchin Paracentrotus Lividus. Mar. Drugs 2014, 12, 4912–4933. [Google Scholar] [CrossRef]
- Ferrario, C.; Leggio, L.; Leone, R.; Di Benedetto, C.; Guidetti, L.; Coccè, V.; Ascagni, M.; Bonasoro, F.; La Porta, C.A.M.; Candia Carnevali, M.D.; et al. Marine-Derived Collagen Biomaterials from Echinoderm Connective Tissues. Mar. Environ. Res. 2017, 128, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.; Rusconi, F.; Pulaj, A.; Macchi, R.; Landini, P.; Paroni, M.; Colombo, G.; Martinello, T.; Melotti, L.; Gomiero, C.; et al. From Food Waste to Innovative Biomaterial: Sea Urchin-Derived Collagen for Applications in Skin Regenerative Medicine. Mar. Drugs 2020, 18, 414. [Google Scholar] [CrossRef] [PubMed]
- Carolo, A.; Melotti, L.; Zivelonghi, G.; Sacchetto, R.; Akyürek, E.E.; Martinello, T.; Venerando, A.; Iacopetti, I.; Sugni, M.; Martinelli, G.; et al. Mutable Collagenous Tissue Isolated from Echinoderms Leads to the Production of a Dermal Template That Is Biocompatible and Effective for Wound Healing in Rats. Mar. Drugs 2023, 21, 506. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Yamamoto, K.; Ikeda, T.; Yanagiguchi, K.; Hayashi, Y. Potency of Fish Collagen as a Scaffold for Regenerative Medicine. BioMed Res. Int. 2014, 2014, e302932. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Lv, Y. Application of Collagen Scaffold in Tissue Engineering: Recent Advances and New Perspectives. Polymers 2016, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Amirthalingam, S.; Hwang, N.S. 21—Collagen-Based Biomaterials for Tissue Engineering Applications. In Natural Biopolymers in Drug Delivery and Tissue Engineering; Jayakumar, R., Murali, V.P., Eds.; Woodhead Publishing Series in Biomaterials; Woodhead Publishing: Cambridge, UK, 2023; pp. 541–571. ISBN 978-0-323-98827-8. [Google Scholar]
- Rezvani Ghomi, E.; Nourbakhsh, N.; Akbari Kenari, M.; Zare, M.; Ramakrishna, S. Collagen-Based Biomaterials for Biomedical Applications. J. Biomed. Mater. Res. B Appl. Biomater. 2021, 109, 1986–1999. [Google Scholar] [CrossRef]
- Lin, Z.; Solomon, K.L.; Zhang, X.; Pavlos, N.J.; Abel, T.; Willers, C.; Dai, K.; Xu, J.; Zheng, Q.; Zheng, M. In Vitro Evaluation of Natural Marine Sponge Collagen as a Scaffold for Bone Tissue Engineering. Int. J. Biol. Sci. 2011, 7, 968–977. [Google Scholar] [CrossRef]
- Nagai, T.; Izumi, M.; Ishii, M. Fish Scale Collagen. Preparation and Partial Characterization. Int. J. Food Sci. Technol. 2004, 39, 239–244. [Google Scholar] [CrossRef]
- Song, E.; Yeon Kim, S.; Chun, T.; Byun, H.-J.; Lee, Y.M. Collagen Scaffolds Derived from a Marine Source and Their Biocompatibility. Biomaterials 2006, 27, 2951–2961. [Google Scholar] [CrossRef]
- Hoyer, B.; Bernhardt, A.; Lode, A.; Heinemann, S.; Sewing, J.; Klinger, M.; Notbohm, H.; Gelinsky, M. Jellyfish Collagen Scaffolds for Cartilage Tissue Engineering. Acta Biomater. 2014, 10, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-R.; Shiau, C.-Y.; Chen, H.-H.; Huang, B.-C. Isolation and Characterization of Acid and Pepsin-Solubilized Collagens from the Skin of Balloon Fish (Diodon Holocanthus). Food Hydrocoll. 2011, 25, 1507–1513. [Google Scholar] [CrossRef]
- Singh, P.; Benjakul, S.; Maqsood, S.; Kishimura, H. Isolation and Characterisation of Collagen Extracted from the Skin of Striped Catfish (Pangasianodon Hypophthalmus). Food Chem. 2011, 124, 97–105. [Google Scholar] [CrossRef]
- Heinemann, S.; Ehrlich, H.; Douglas, T.; Heinemann, C.; Worch, H.; Schatton, W.; Hanke, T. Ultrastructural Studies on the Collagen of the Marine Sponge Chondrosia Reniformis Nardo. Biomacromolecules 2007, 8, 3452–3457. [Google Scholar] [CrossRef] [PubMed]
- Addad, S.; Exposito, J.-Y.; Faye, C.; Ricard-Blum, S.; Lethias, C. Isolation, Characterization and Biological Evaluation of Jellyfish Collagen for Use in Biomedical Applications. Mar. Drugs 2011, 9, 967–983. [Google Scholar] [CrossRef]
- Wilkie, I.C.; Sugni, M.; Gupta, H.S.; Carnevali, M.D.C.; Elphick, M.R. The Mutable Collagenous Tissue of Echinoderms: From Biology to Biomedical Applications; Royal Society of Chemistry: London, UK, 2021. [Google Scholar] [CrossRef]
- Carnevali, M.D.; Bonasoro, F.; Wilkie, I.; Andrietti, F.; Melone, G. Functional Morphology of the Peristomial Membrane of Regular Sea-Urchins: Structural Organization and Mechanical Properties in Paracentrotus Lividus; CRC Press: Boca Raton, FL, USA, 1990; pp. 207–216. ISBN 978-1-00-307895-1. [Google Scholar]
- Candia Carnevali, M.D.; Sugni, M.; Bonasoro, F.; Wilkie, I.C. Mutable Collagenous Tissue: A Concept Generator for Biomimetic Materials and Devices. Mar. Drugs 2024, 22, 37. [Google Scholar] [CrossRef]
- Sun, J.; Chiang, F.-S. Use and Exploitation of Sea Urchins. In Echinoderm Aquaculture; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 25–45. ISBN 978-1-119-00581-0. [Google Scholar]
- Cirino, P.; Ciaravolo, M.; Paglialonga, A.; Toscano, A. Long-Term Maintenance of the Sea Urchin Paracentrotus Lividus in Culture. Aquac. Rep. 2017, 7, 27–33. [Google Scholar] [CrossRef]
- Guillou, M.; Michel, C. Reproduction and Growth of Sphaerechinus Granularis (Echinodermata: Echinoidea) in Southern Brittany. J. Mar. Biol. Assoc. United Kingd. 1993, 73, 179–192. [Google Scholar] [CrossRef]
- Boudouresque, C.; Verlaque, M. Ecology of Paracentrotus Lividus. Dev. Aquac. Fish. Sci. 2001, 32, 177–216. [Google Scholar] [CrossRef]
- Jordana, E.; Guillou, M.; Lumingas, L.J.L. Age and Growth of the Sea Urchin Sphaerechinus Granularis in Southern Brittany. J. Mar. Biol. Assoc. United Kingd. 1997, 77, 1199–1212. [Google Scholar] [CrossRef]
- Addis, P.; Moccia, D.; Secci, M. Effect of Two Different Habitats on Spine and Gonad Colour in the Purple Sea Urchin Paracentrotus Lividus. Mar. Ecol. 2014, 36, 178–184. [Google Scholar] [CrossRef]
- Ribeiro, A.R.; Barbaglio, A.; Oliveira, M.J.; Santos, R.; Coelho, A.V.; Ribeiro, C.C.; Wilkie, I.C.; Carnevali, M.D.C.; Barbosa, M.A. Correlations between the Biochemistry and Mechanical States of a Sea-Urchin Ligament: A Mutable Collagenous Structure. Biointerphases 2012, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Nhu Hieu, V.M.; Thanh Van, T.T.; Hang, C.T.T.; Mischenko, N.P.; Sergey, A.F.; Truong, H.B. Polyhydroxynaphthoquinone Pigment from Vietnam Sea Urchins as a Potential Bioactive Ingredient in Cosmeceuticals. Nat. Prod. Commun. 2020, 15, 1–8. [Google Scholar] [CrossRef]
- Lim, Y.-S.; Ok, Y.-J.; Hwang, S.-Y.; Kwak, J.-Y.; Yoon, S. Marine Collagen as A Promising Biomaterial for Biomedical Applications. Mar. Drugs 2019, 17, 467. [Google Scholar] [CrossRef]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens—Structure, Function, and Biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef] [PubMed]
- Chandika, P.; Ko, S.-C.; Oh, G.-W.; Heo, S.-Y.; Nguyen, V.-T.; Jeon, Y.-J.; Lee, B.; Jang, C.H.; Kim, G.; Park, W.S.; et al. Fish Collagen/Alginate/Chitooligosaccharides Integrated Scaffold for Skin Tissue Regeneration Application. Int. J. Biol. Macromol. 2015, 81, 504–513. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, E.; Pierfelice, T.V.; Lepore, S.; Iezzi, G.; D’Arcangelo, C.; Piattelli, A.; Covani, U.; Petrini, M. Hemostatic Collagen Sponge with High Porosity Promotes the Proliferation and Adhesion of Fibroblasts and Osteoblasts. Int. J. Mol. Sci. 2023, 24, 7749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhou, L.; Zhang, W. Control of Scaffold Degradation in Tissue Engineering: A Review. Tissue Eng. Part B Rev. 2014, 20, 492–502. [Google Scholar] [CrossRef]
- Bozkurt, A.; Apel, C.; Sellhaus, B.; van Neerven, S.; Wessing, B.; Hilgers, R.-D.; Pallua, N. Differences in Degradation Behavior of Two Non-Cross-Linked Collagen Barrier Membranes: An in Vitro and in Vivo Study. Clin. Oral Implant. Res. 2014, 25, 1403–1411. [Google Scholar] [CrossRef]
- Shui, H.; Shi, Q.; Pugno, N.M.; Chen, Q.; Li, Z. Effect of Mechanical Stimulation on the Degradation of Poly(Lactic Acid) Scaffolds with Different Designed Structures. J. Mech. Behav. Biomed. Mater. 2019, 96, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Wissing, T.B.; Bonito, V.; Bouten, C.V.C.; Smits, A.I.P.M. Biomaterial-Driven in Situ Cardiovascular Tissue Engineering—A Multi-Disciplinary Perspective. npj Regen. Med. 2017, 2, 18. [Google Scholar] [CrossRef] [PubMed]
- Davidenko, N.; Bax, D.V.; Schuster, C.F.; Farndale, R.W.; Hamaia, S.W.; Best, S.M.; Cameron, R.E. Optimisation of UV Irradiation as a Binding Site Conserving Method for Crosslinking Collagen-Based Scaffolds. J. Mater. Sci. Mater. Med. 2015, 27, 14. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Park, J.C.; Hwang, Y.S.; Kim, J.K.; Kim, J.G.; Suh, H. Characterization of UV-Irradiated Dense/Porous Collagen Membranes: Morphology, Enzymatic Degradation, and Mechanical Properties. Yonsei Med. J. 2001, 42, 172. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Ding, J. Effects of Porosity and Pore Size Onin Vitro Degradation of Three-Dimensional Porous Poly(D,L-Lactide-Co-Glycolide) Scaffolds for Tissue Engineering. J. Biomed. Mater. Res. 2005, 75A, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, L.; Calò, E.; Bonfrate, V.; Pedone, D.; Gallo, N.; Natali, M.L.; Sannino, A.; Madaghiele, M. Exploring the Effects of the Crosslink Density on the Physicochemical Properties of Collagen-Based Scaffolds. Polym. Test. 2021, 93, 106966. [Google Scholar] [CrossRef]
- Tziveleka, L.-A.; Sapalidis, A.; Kikionis, S.; Aggelidou, E.; Demiri, E.; Kritis, A.; Ioannou, E.; Roussis, V. Hybrid Sponge-Like Scaffolds Based on Ulvan and Gelatin: Design, Characterization and Evaluation of Their Potential Use in Bone Tissue Engineering. Materials 2020, 13, 1763. [Google Scholar] [CrossRef] [PubMed]
- Raftery, R.M.; Woods, B.; Marques, A.L.P.; Moreira-Silva, J.; Silva, T.H.; Cryan, S.-A.; Reis, R.L.; O’Brien, F.J. Multifunctional Biomaterials from the Sea: Assessing the Effects of Chitosan Incorporation into Collagen Scaffolds on Mechanical and Biological Functionality. Acta Biomater. 2016, 43, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ahn, S.-H.; Kim, G.H. Three-Dimensional Collagen/Alginate Hybrid Scaffolds Functionalized with a Drug Delivery System (DDS) for Bone Tissue Regeneration. Chem. Mater. 2012, 24, 881–891. [Google Scholar] [CrossRef]
- Albu, M.G.; Ferdes, M.; Kaya, D.A.; Ghica, M.V.; Titorencu, I.; Popa, L.; Albu, L. Collagen Wound Dressings with Anti-Inflammatory Activity. Mol. Cryst. Liq. Cryst. 2012, 555, 271–279. [Google Scholar] [CrossRef]
- Graham, H.K.; McConnell, J.C.; Limbert, G.; Sherratt, M.J. How Stiff Is Skin? Exp. Dermatol. 2019, 28, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Geerligs, M.; van Breemen, L.; Peters, G.; Ackermans, P.; Baaijens, F.; Oomens, C. In Vitro Indentation to Determine the Mechanical Properties of Epidermis. J. Biomech. 2011, 44, 1176–1181. [Google Scholar] [CrossRef] [PubMed]
- Ottenio, M.; Tran, D.; Ní Annaidh, A.; Gilchrist, M.D.; Bruyère, K. Strain Rate and Anisotropy Effects on the Tensile Failure Characteristics of Human Skin. J. Mech. Behav. Biomed. Mater. 2015, 41, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Ní Annaidh, A.; Bruyère, K.; Destrade, M.; Gilchrist, M.D.; Otténio, M. Characterization of the Anisotropic Mechanical Properties of Excised Human Skin. J. Mech. Behav. Biomed. Mater. 2012, 5, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska-Gralak, M.; Sadło, J.; Głuszewski, W.; Łyczko, K.; Przybytniak, G.; Lewandowska, H. The Combined Effect of Humidity and Electron Beam Irradiation on Collagen Type I—Implications for Collagen-Based Devices. Mater. Today Commun. 2022, 31, 103255. [Google Scholar] [CrossRef]
- Liang, W.-H.; Kienitz, B.L.; Penick, K.J.; Welter, J.F.; Zawodzinski, T.A.; Baskaran, H. Concentrated Collagen-Chondroitin Sulfate Scaffolds for Tissue Engineering Applications. J. Biomed. Mater. Res. Part A 2010, 94A, 1050–1060. [Google Scholar] [CrossRef] [PubMed]
- Gaharwar, A.K.; Cross, L.M.; Peak, C.W.; Gold, K.; Carrow, J.K.; Brokesh, A.; Singh, K.A. 2D Nanoclay for Biomedical Applications: Regenerative Medicine, Therapeutic Delivery, and Additive Manufacturing. Adv. Mater. 2019, 31, 1900332. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Yan, F.; Xiang, X.; Tang, Y.; Zhang, L.; Liu, J.; Qiu, L. Determination of Normal Skin Elasticity by Using Real-Time Shear Wave Elastography. J. Ultrasound Med. 2018, 37, 2507–2516. [Google Scholar] [CrossRef] [PubMed]
- Tajvar, S.; Hadjizadeh, A.; Samandari, S.S. Scaffold Degradation in Bone Tissue Engineering: An Overview. Int. Biodeterior. Biodegrad. 2023, 180, 105599. [Google Scholar] [CrossRef]
- Krafts, K.P. Tissue Repair. Organogenesis 2010, 6, 225–233. [Google Scholar] [CrossRef]
- Arora, A.; Kothari, A.; Katti, D.S. Pore Orientation Mediated Control of Mechanical Behavior of Scaffolds and Its Application in Cartilage-Mimetic Scaffold Design. J. Mech. Behav. Biomed. Mater. 2015, 51, 169–183. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extraction | % (EchA + SpA) | % SpB | % SpD | % SpE | % Echinamine | EC50 (mg/mL) |
|---|---|---|---|---|---|---|
| P. l_ 28.03.2023 | 50 | 11 | 0 | 37 | 1.5 | 0.0037 |
| P. l_ 11.01.2023 | 23.6 | 73 | 0 | 1.2 | 2.8 | 0.03 |
| P. l_ 06.07.2022 | 26 | 70 | 0 | 1.7 | 0.2 | 0.033 |
| P. l_12.04.2022 | 22 | 72 | 0 | 3.9 | 1.2 | 0.019 |
| P. l_ 10.01.2022 | 92.5 | 7.5 | 0 | 0 | 0 | 0.027 |
| S. g_ 08.06.2023_P | 90.7 | 0 | 4.1 | 4.6 | 0.6 | 0.0034 |
| S. g_ 08.06.2023_W | 86 | 0 | 2.8 | 11.2 | 0 | 0.0041 |
| S. g_ 17.05.2023_P | 86.4 | 0 | 5.1 | 8 | 0.5 | 0.0053 |
| S. g_ 17.05.2023_W | 75.8 | 0 | 1.9 | 21.8 | 0.5 | 0.0068 |
| S. g_ 13.02.2023_P | 66.4 | 0 | 18.9 | 14.7 | 0 | 0.0071 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roncoroni, M.; Martinelli, G.; Farris, S.; Marzorati, S.; Sugni, M. Sea Urchin Food Waste into Bioactives: Collagen and Polyhydroxynaphtoquinones from P. lividus and S. granularis. Mar. Drugs 2024, 22, 163. https://doi.org/10.3390/md22040163
Roncoroni M, Martinelli G, Farris S, Marzorati S, Sugni M. Sea Urchin Food Waste into Bioactives: Collagen and Polyhydroxynaphtoquinones from P. lividus and S. granularis. Marine Drugs. 2024; 22(4):163. https://doi.org/10.3390/md22040163
Chicago/Turabian StyleRoncoroni, Margherita, Giordana Martinelli, Stefano Farris, Stefania Marzorati, and Michela Sugni. 2024. "Sea Urchin Food Waste into Bioactives: Collagen and Polyhydroxynaphtoquinones from P. lividus and S. granularis" Marine Drugs 22, no. 4: 163. https://doi.org/10.3390/md22040163
APA StyleRoncoroni, M., Martinelli, G., Farris, S., Marzorati, S., & Sugni, M. (2024). Sea Urchin Food Waste into Bioactives: Collagen and Polyhydroxynaphtoquinones from P. lividus and S. granularis. Marine Drugs, 22(4), 163. https://doi.org/10.3390/md22040163