
Marine Pharmacology in 2019–2021: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action †
 , ,
, ,  , and
, and 
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug Class | Compound/ Organism a | Chemistry | Pharmacologic Activity | IC50 b | MMOA c | Country/ Territory d | References |
|---|---|---|---|---|---|---|---|
| Antibacterial | adipostatin E (1)/ bacterium | Polyketide h | B. subtilis and L. monocytogenes inhibition | 3.4, 5.9 µM | PPCS inhibition | CRI, USA | [14] |
| Antibacterial | arenicin-3 (2)/worm | Peptide f | E. coli and K. pneumoniae inhibition | 0.38–0.76 μM + | Cell membrane disruption and ATP release | AUS, CHE, CHN, DNK, DEU, GBR, IRL | [15] |
| Antibacterial | bisanhydroaklavinone (3)/bacterium | Polyketide h | S. aureus inhibition | 16.6 μM + | Cell membrane damage and DNA leakage | PHL, SGP | [16] |
| Antibacterial | cladodionen (4)/ fungus | Polyketide h | P. aeruginosa quorum sensing inhibition | <400 µM | Downregulation of quorum sensing genes | CHN | [17] |
| Antibacterial | cyclo(l-leucyl-l-prolyl) (5)/bacterium | Peptide f | S. marcescens inhibition | 952 μM + | Biofilm formation inhibition | IND | [18] |
| Antibacterial | C. cervicornis diterpene (6)/alga | Terpenoid e | MR S. aureus inhibition | 22 μM + | Inhibition of efflux pump | BRA | [19] |
| Antibacterial | chrysophaentin I (7)/alga | Polyketide h | S. aureus inhibition | 15.5 μM + | Cytoskeletal protein FtsZ inhibition | USA | [20] |
| Antibacterial | crustin (8)/shrimp | Peptide f | M. luteus inhibition | 2.5 μM + | Membrane disruption and depolarization | CHN | [21] |
| Antibacterial | D. candidum alkaloid (9)/ascidian | Alkaloid f | S. aureus, E. coli, K. pneumoniae inhibition | 18.5 μM + | Biofilm formation inhibition | ITA | [22] |
| Antibacterial | doscadenamide A (10)/cyanobacterium | Peptide f/ Polyketide h | P. aeruginosa quorum sensing activation | <10 µM | AHL-binding site | USA | [23] |
| Antibacterial | kalafungin (11)/ bacterium | Polyketide h | S. aureus inhibition | 27, 53 μM + | Non-competitive β-lactamase inhibition | IND | [24] |
| Antibacterial | korormicin A (12)/bacterium | Polyketide h | V. cholerae and P. aeruginosa inhibition | 10–30 μM + | Reactive oxygen species production | BRA, JPN, USA | [25] |
| Antibacterial | lactoquinomycin A (13)/bacterium | Polyketide h | MR S. aureus and S. enterica inhibition | 0.06–0.55 μM + | Induction of DNA damage | S. KOR | [26] |
| Antibacterial | octominin (14)/ octopus | Peptide f | S. parauberis inhibition | 18.8 μM + | Membrane disruption and chromosomal DNA binding | S. KOR | [27] |
| Antibacterial | P. chrysogenum dipeptide (15)/ fungus | Peptide f | C. violaceum and P. aeruginosa inhibition | 91.4 mM + | Anti-quorum sensing activity | CHN | [28] |
| Antibacterial | piscidin 5 (16)/fish | Peptide f | V. parahaemolyticus and P. damselae inhibition | 1.5–6.2 µM | Membrane disruption and DNA binding | CHN | [29] |
| Antibacterial | phorbaketal B and C (17, 18)/sponge | Terpenoid e | S. aureus biofilm inhibition | <125 μM | Downregulation of hemolysin-related genes | S. KOR | [30] |
| Antibacterial | S. algae polyketide (19)/bacterium | Polyketide h | E. coli and MR S. aureus inhibition | 9.3 μM + | MRSA penicillin-binding protein active site docking | IND | [31] |
| Antibacterial | S. algae polyketide (20)/bacterium | Polyketide h | VR E. faecalis and MR S. aureus inhibition | 2–6 μM + | Siderophore mechanism of action | IND | [32] |
| Antibacterial | securamine H (21)/bryozoan | Alkaloid f | S. aureus inhibition | 3.13 μM + | Reduction in metabolic activity | NOR | [33] |
| Antibacterial | turgencin A (22)/ ascidian | Peptide f | C. glutamicum and B. subtilis inhibition | 0.4 μM + | Cell membrane disruption | AUS, NOR | [34] |
| Antibacterial | tyramine (23)/ bacterium | Alkaloid f | P. aeruginosa quorum sensing inhibition | 7.3 mM + | Pyoverdine production inhibition | ESP | [35] |
| Antifungal | amantelide A (24)/ cyanobacterium | Polyketide h | S. cervisiae inhibition | 12.5, 50 μM | Ergosterol binding and actin polymerization promotion | JPN, PHL, USA | [36] |
| Antifungal | atranone Q (25)/ fungus | Terpenoid e | C. albicans growth inhibition | 20.5 μM | Cytoplasm agglutination and cell membrane alterations | CHN | [37] |
| Antifungal | fusarilactone A (26)/fungus | Polyketide h | P. theae growth inhibition | 118.2 μM | HMG-CoA inhibition | CHN | [38] |
| Antifungal | 2-n-heptyl-4- hydroxyquinoline (27)/bacterium | Alkaloid f | C. albicans hyphal growth inhibition | 46.9 μM | cAMP-Efg1 pathway inhibition | S. KOR | [39] |
| Antifungal | oceanapiside (28)/sponge | Polyketide h | C. glabrata inhibition | 15.4 μM | Sphingolipid synthesis inhibition | PHL, USA | [40] |
| Antifungal | puupehenone (29)/sponge | Terpenoid e | CAS-insensitive C. neoformans inhibition | 7.6–15.2 μM + | CWI integrity pathway disruption | USA | [41] |
| Antifungal | S. olivaceus butyrylamide (30)/bacterium | Shikimate g | C. albicans hyphal growth inhibition and adhesion | 487.4 μM + | Downregulation of hyphal formation genes | CHN | [42] |
| Antimalarial | capillasterquinone B (31)/bacterium | Polyketide h | P. falciparum 3D7 inhibition | 29.3 µM | Lysyl-tRNA synthetase binding | DEU, EGY, GBR, SAU | [43] |
| Antimalarial | kakeromamide B (32)/cyanobacterium | Peptide f | Blood-stage P. falciparum inhibition | 8.9 μM | Binding to Plasmodium actin and sortilin | USA | [44] |
| Antimalarial | friomaramide (33)/sponge | Peptide f | P. falciparum sporozoites liver infection inhibition | <6.1 μM * | Hepatocyte nuclei viability confirmed | AUS, USA | [45] |
| Antimalarial | nitenin (34)/sponge | Terpenoid e | P. falciparum inhibition | 0.29 μM | Ring to trophozoite transition | USA | [46] |
| Antiprotozoal | 4-epi-arbusculin A (35)/zoanthid | Terpenoid e | A. castellanii inhibition | 26 μM | Programmed cell death induction | ESP | [47] |
| Antiprotozoal | epinecidin-1 (36)/fish | Peptide f | Trichomonas vaginalis inhibition | <26.7 µM | Membrane disruption | TWN | [48] |
| Antiprotozoal | isololiolide (37)/ hydroid | Terpenoid e | T. cruzi trypomastigotes and amastigotes inhibition | 32, 40 μM | Disruption of membrane integrity | BRA, USA | [49] |
| Antiprotozoal | dehydrothyrsiferol (38)/alga | Terpenoid e | A. castellanii growth inhibition | 5.3 μM | Mitochondrial malfunction | MEX, ESP | [50] |
| Antiprotozoal | gallinamide A (39)/cyanobacterium | Peptide f | T. cruzi amastigote inhibition | 14.7 nM | Recombinant cruzain inhibition | USA | [51] |
| Antiprotozoal | 7-oxostaurosporine (40)/bacterium | Alkaloid f | A. castellanii growth inhibition | 0.8, 0.9, 5.5 μM | Mitochondrial malfunction | ECU, ESP | [52] |
| Antiprotozoal | polyaurine A (41)/ ascidian | Alkaloid f | S. mansoni inhibition | >100 μM | Egg-production impairment in vitro | IDN, ITA | [53] |
| Antituberculosis | fiscpropionate A (42)/fungus | Polyketide h | M. tuberculosis MptpB inhibition | 5.1 μM | Noncompetitive inhibition | CHN | [54] |
| Antituberculosis | fucoxanthin (43)/alga | Terpenoid e | M. tuberculosis strains inhibition | 2.8–4.1 μM + | TBNAT inhibition | CHL, CZE, IRN, ROU | [55] |
| Antiviral | chartarlactam T (44)/fungus | Alkaloid f | Zika virus inhibition | 10 μM * | Protein E inhibition | CHN | [56] |
| Antiviral | harzianoic acids A and B (45, 46)/fungus | Terpenoid e | HCV inhibition | 35,43 μM | Virus replication and entry inhibition | CHN, DEU | [57] |
| Antiviral | homoseongomycin (47)/bacterium | Polyketide h | VEEV and EEEV inhibition | 8.6 μM | Viral replication inhibition | TWN, USA | [58] |
| Antiviral | penicillixanthone A (48)/fungus | Polyketide h | HIV-1 replication inhibition | 0.36 μM | CCR5/CXCR4 receptor antagonist | CHN | [59] |
| Antiviral | portimine (49)/ dinoflagellate | Polyketide h | HIV-1 replication inhibition | 4.1 nM | Reverse-transcriptase inhibition | JPN | [60] |



2. Marine Compounds with Antibacterial, Antifungal, Antiprotozoal, Antituberculosis and Antiviral Activities
2.1. Antibacterial Activity
2.2. Antifungal Activity
2.3. Antiprotozoal and Antituberculosis Activity
2.4. Antiviral Activity
3. Marine Compounds with Antidiabetic and Anti-Inflammatory Activity, and Affecting the Immune and Nervous System
| Drug Class | Compound/ Organism a | Chemistry | Pharmacological Activity | IC50 b | MMOA c | Country/ Territory d | References |
|---|---|---|---|---|---|---|---|
| Antidiabetic | xyloccensin-1 (50)/ mangrove | Terpenoid f | α-glucosidase inhibition | 0.25 mM | Docking studies completed | IND | [61] |
| Antidiabetic | CYC27 (51)/alga | Shikimate h | Reduction in blood glucose | 50 mg/kg/day ** | Insulin signaling pathways enhanced | CHN | [62] |
| Antidiabetic | fucoxanthin (43)/alga | Terpenoid f | α-amylase and α-glucosidase inhibition | 121 µM | Mixed-type inhibition kinetics | DNK, MYS, S. KOR, THA | [63,64] |
| Antidiabetic | fucoxanthin (43)/alga | Terpenoid f | Decrease ROS production in kidney mensangial cell line | 0.5 µM * | Epigenomic and transcriptomic effects | USA | [65] |
| Antidiabetic | abeo-oleanene (52)/alga | Terpenoid f | α-amylase and α-glucosidase inhibition | 0.29 mM | Docking studies completed | IND | [66] |
| Antidiabetic | isophloroglucin A (53)/alga | Polyketide e | Glucose homeostasis improvement | 1.35 mg/kg/day ** | GLUT4 levels increased | S. KOR | [67] |
| Antidiabetic | S. latiuscula bromophenol (54)/alga | Shikimate h | α-glucosidase inhibition | 1.92 µM | PTP1B competitive inhibition | S. KOR | [68] |
| Antidiabetic | H. fusiformis fatty acid (55)/alga | Fatty Acids | α-glucosidase inhibition | 48 µM | PTP1B inhibition | S. KOR | [69] |
| Antidiabetic | tripalmitin (56)/fungus | Fatty Acids | α-glucosidase inhibition | 3.75 µM | Mixed-type inhibition kinetics | PAN | [70] |
| Anti-inflammatory | A. depilans EnP(5,8) (57)/sea hare | Terpenoid f | Macrophage NO, COX-2, IL-6 and TNF-α | 18.4 μM | Nos2 and COX-2 expression decrease | ESP, PRT | [71] |
| Anti-inflammatory | Aspergillus sp. aglycone (58)/fungus | Polyketide e | Macrophage NO release inhibition | 6 μM | NF-κB inhibition | CHN | [72] |
| Anti-inflammatory | brevenal (59)/ dinoflagellate | Polyketide e | Macrophage TNF-α inhibition | 0.1 nM | Macrophage activation inhibition | USA | [73] |
| Anti-inflammatory | caniferolide A (60)/ bacterium | Polyketide e | Microglia NO, IL-1β, IL-6 release inhibition | 0.01 μM * | iNOS, ERK, JNK expression inhibition | ESP | [74] |
| Anti-inflammatory | C. inophyllum terpenoids (61, 62)/mangrove | Terpenoid f/ Shikimate h | Macrophage NO and IL-1β release inhibition | 2.4, 7 μM | iNOS induction and NF-κB inhibition | VNM, S. KOR | [75] |
| Anti-inflammatory | curdepsidone C (63)/fungus | Polyketide e/ Shikimate h | Human macrophage IL-1β release inhibition | 7.5 μM | JNK and ERK inhibition | CHN | [76] |
| Anti-inflammatory | collismycin C (64)/ bacterium | Alkaloid g | Murine sepsis inhibition and survival | 4 mg/kg ** | NF-κB and p38 inhibition | S. KOR | [77] |
| Anti-inflammatory | dieckol (65)/alga | Polyketide e | Decreased murine liver NLRP3 synthesis | 2.5 mg/kg/day ** | NF-κB and NLRP3 inhibition | S. KOR | [78] |
| Anti-inflammatory | dysiarenone (66)/sponge | Terpenoid f | Macrophage IL-6, TNF-α and LTB4 release inhibition | 2–8 μM * | NF-κB, p38, ERK, Akt inhibition | CHN | [79] |
| Anti-inflammatory | epiloliolide (67)/alga | Terpenoid f | Human periodontal ligament cell iNOS, IL-1, IL-6, and TNF-α inhibition | >10 μM * | NLRP3 decrease and PKA/CREB increase | S. KOR | [80] |
| Anti-inflammatory | fucoxanthin (43)/ diatom | Terpenoid f | Murine sepsis inhibition and survival | 1 mg/kg ** | NF-κB inhibition | CHN, TWN, USA | [81] |
| Anti-inflammatory | fucoxanthin (43)/ diatom | Terpenoid f | Murine liver inflammation inhibition | 10–40 mg/kg ** | NF-κB inhibition and NRF2 increase | CHN | [82] |
| Anti-inflammatory | fucoxanthin (43)/alga | Terpenoid f | Macrophage osteoclastogenesis inhibition | <5 μM * | ERK, p38 inhibition and NRF2 increase | S. KOR | [83] |
| Anti-inflammatory | fucoxanthin (43)/alga | Terpenoid f | Macrophage iNOS and COX-2 expression inhibition | 5, 10 μM * | NF-κB inhibition | CHN, USA | [84] |
| Anti-inflammatory | fucoxanthinol (68)/ diatom | Terpenoid f | Microglia NO and PGE2 expression inhibition | 20 μM * | NF-κB, Akt, MAPK inhibition and NRF2 increase | CHN | [85] |
| Anti-inflammatory | hirsutanol A (69)/ fungus | Terpenoid f | LPS-induced MMP-9 release and lung injury attenuation | 30 mg/kg ** | NF-κB, STAT3, ERK inhibition | RUS, TWN | [86] |
| Anti-inflammatory | 2-epi-jaspine B (70)/sponge | Alkaloid g | Rat arthritis inhibition | 30 mg/kg ** | SphK1 inhibition | CHN | [87] |
| Anti-inflammatory | L. glandulifera diterpenes (71, 72)/alga | Terpenoid f | Macrophage NO release inhibition | 2.3, 2.9 μM | iNOS induction inhibition | GRC | [88] |
| Anti-inflammatory | mojabanchromanol (73)/alga | Terpenoid f | Murine alveolar epithelial cell line lipid peroxidation inhibition | 147.4 µM * | ERK, JNK inhibition | S. KOR | [89] |
| Anti-inflammatory | neuchromenin (74)/ fungus | Polyketide e | Microglia NO and PGE2 inhibition | 2.7, 3.2 μM | NF-κB and p38 inhibition | S. KOR | [90] |
| Anti-inflammatory | O-demethylrenierone (75)/sponge | Alkaloid g | Human macrophage NO and PGE2, inhibition | 10 µM * | NF-κB inhibition and increase | S. KOR, VNM | [91] |
| Anti-inflammatory | penicitrinone A (76)/fungus | Polyketide e | Human neutrophil superoxide anion inhibition | 2.7 µM | caspase-3- dependent apoptosis induction | TWN | [92] |
| Anti-inflammatory | phyllohemiketal A (77)/sponge | Terpenoid f | Human macrophage NO and PGE2 inhibition | 5 µM * | NF-κB, p38, ERK and JNK inhibition and NRF2 increase | S. KOR | [93] |
| Anti-inflammatory | sclerketide C (78)/ fungus | Alkaloid g | Macrophage NO release inhibition | 2.7 µM | iNOS and COX-2 mRNA expression decrease | CHN | [94] |
| Anti-inflammatory | grasshopper ketone (79)/alga | Terpenoid f | Macrophage NO, IL-1β, IL-6 release inhibition | 4.5–45 µM * | NF-κB, p38, ERK, JNK inhibition | S. KOR | [95] |
| Anti-inflammatory | S. mastoidea prodigiosins (80, 81)/bacterium | Alkaloid g | Rat gastric inflammation inhibition | >100 mg/kg ** | NF-κB inhibition and HO-1 increase | EGY | [96] |
| Anti-inflammatory | topsentin (82)/sponge | Alkaloid g | Human keratinocyte COX-2 expression inhibition | 1.2 µM | AP-1, p38, JNK, and Erk inhibition | S. KOR | [97] |
| Anti-inflammatory | tuberatolide B (83)/alga | Polyketide e/ Terpenoid f | Macrophage NO, IL-1β, IL-6 release inhibition | 29.6 µM * | NF-κB, p38, ERK, JNK inhibition | S. KOR | [98] |
| Immune system | astaxanthin (84)/alga | Terpenoid f | Inhibition of LPS-induced dendritic cell dysfunction | 5–20 µM * | HO-1 and NRF-2 increase | CHN | [99] |
| Immune system | crassolide (85)/soft coral | Terpenoid f | Suppression of dendritic cell maturation and T cell responses | 2.5 µM * | DC maturation and pro-inflammatory cytokines inhibition | TWN | [100] |
| Immune system | C. sinensis peptide (86)/mollusk | Peptide g | Increased murine macrophage phagocytosis | 25 µM * | NF-κB and NLRP3 increase | CHN | [101] |
| Immune system | dieckol (65)/alga | Polyketide e | Decreased intestinal Th17 cells and increased Treg cells | 2.5 mg/kg/day ** | NF-κB and IL-6 decrease | S. KOR | [102] |
| Immune system | echinochrome A (87)/sea urchin | Polyketide e | Expansion of PBMC-derived CD34+ cells | 10 µM * | ROS and p38MAPK/JNK phosphorylation decrease | S. KOR, RUS | [103] |
| Immune system | echinochrome A (87)/sea urchin | Polyketide e | Protection against murine inflammatory bowel disease | 10 mg/kg ** | Regulatory T cell production increase | S. KOR, RUS | [104] |
| Immune system | echinochrome A (87)/sea urchin | Polyketide e | Inhibition of murine bleomycin-induced scleroderma | 1 µM * | STAT3 phosphorylation decrease | S. KOR, RUS | [105] |
| Immune system | eckol (88)/alga | Polyketide e | Inhibition murine IgE-mediated PCA reaction | 50 µg/mouse ** | FCεR and NF-κB activation decrease | S. KOR | [106] |
| Immune system | phomaketide A (89)/fungus | Polyketide e/ Terpenoid f | Lymphangiogenesis inhibition | 3.7 µM | VEGFR-3 phosphorylation and PKCδ activation decrease | TWN | [107] |
| Immune system | S. scabra cembranoid (90)/soft coral | Terpenoid f | LPS-induced B lymphocyte proliferation | 4.4 µM | B cell proliferation decrease and IL-10 increase | CHN | [108] |
| Immune system | sticholysins I and II (proteins of about 20KD)/sea anemone | Peptide g | Maturation of dendritic cells | 0.05 µM * | TLR4 and MYD88 activation decrease | BRA, CUB, USA | [109] |
| Immune system | T. weissflogii phosphoglycolipid (91)/diatom | Polyketide e | Immune stimulation of human monocyte-derived dendritic cells | 6.8 µM * | TLR4 and NF-κB activation decrease | ITA | [110] |
| Nervous system | alternarin A (92)/fungus | Terpenoid f | Neuronal spontaneous Ca2+ oscillations (SCO) inhibition | 3.2 µM | SCO frequency and amplitude decreased | CHN, HU | [111] |
| Nervous system | anabaseine (93)/worm | Alkaloid g | α7 nAChR inhibition | 1.85–3.85 µM | Membrane depolarization | USA | [112] |
| Nervous system | A. insuetus TMC-120Ac and TMC-120B (94, 95)/fungus | Alkaloid g | Mouse focal seizure duration reduction | 10 mg/kg ** | Undetermined | BEL, DNK, NOR | [113] |
| Nervous system | Ara and ETrA (96, 97)/alga | Fatty Acids | AChE inhibition | 1.6–2.4 mM | Non-competitive inhibition | CHN | [114] |
| Nervous system | astaxanthin (84)/shrimp | Terpenoid f | Reduction in LPS-induced memory impairment | 30 or 50 mg/kg ** | Inhibits STAT3 phosphorylation | S. KOR, USA | [115] |
| Nervous system | astaxanthin (84)/shrimp | Terpenoid f | Cognitive dysfunction protection | 10 mg/kg ** | ROS reduction and decreased Ab | THA | [116] |
| Nervous system | 8,8′-bieckol (98)/alga | Polyketide e | BACE1 and AChE inhibition | 1.6–4.6 µM | Non-competitive or competitive inhibition | S. KOR | [117] |
| Nervous system | brevetoxin (99)/ dinoflagellate | Polyketide e | VGSC activator | 2.4 nM | Shifts voltage dependence, slows inactivation | JPN, USA | [118] |
| Nervous system | C. austini conorfamides (100, 101)/cone snail | Peptide g | α7 nAChR inhibition | 0.68–0.76 µM | Inhibition of Ca2+ ion flow | AUS, MEX | [119] |
| Nervous system | C. geographus conosteroid (102)/ cone snail | Terpenoid f | Hot plate murine pain model inhibition | 2–10 mg/kg ** | GABAAR negative allosteric modulator | USA | [120] |
| Nervous system | C. lividus conotoxin Lv1F (103)/cone snail | Peptide g | α3β2 nAChR inhibition; hotplate and formalin murine pain inhibition | 0.0089 µM; 25–100 µg/kg ** | Competitive binding; unknown | CHN | [121,122] |
| Nervous system | Con-T[M8Q] (104)/cone snail | Peptide g | Inhibition of murine morphine dependence | 15 nmol/kg ** | NMDAR GluN2B antagonist | CHN, USA | [123] |
| Nervous system | dictyol C (105)/alga | Terpenoid f | Neuroprotection of rat CIRI | 80 µg/kg ** | Increased Nrf2/ARE signaling pathway | CHN | [124] |
| Nervous system | echinochrome A (87)/sea urchin | Polyketide e | Mitigation of cerebral ischemic injury | 10 µM ** | Decreases pro-apoptotic factors; increased survival factors | S. KOR, RUS | [125] |
| Nervous system | eckol (88)/alga | Polyketide e | Dopamine D3/D4 agonist | 42, 43 µM | GPCR signaling | S. KOR | [126] |
| Nervous system | eleganolone (106)/alga | Terpenoid f | Human neuroblastoma cells neurotoxicity inhibition | 0.1–1 µM * | Decreases ROS levels and apoptotic factors | BRA, ESP, PRT | [127] |
| Nervous system | frondoside A (107)/sea cucumber | Terpenoid f | Dopaminergic degeneration inhibition | 0.1, 0.5 µM * | Increase in protein degradation pathway, decrease apoptotic factors | THA | [128] |
| Nervous system | fucosterol (108)/alga | Terpenoid f | Aβ-induced neuronal apoptosis | 10 µM * | Decreased pro-apoptotic factors; decreased APP mRNA | MYS | [129] |
| Nervous system | fucosterol (108)/alga | Terpenoid f | Neurodegenerative disorders system pharmacology | NA | Neuronal survival pathways | S.KOR, | [130] |
| Nervous system | fucoxanthin (43)/alga | Terpenoid f | Reduced corneal denervation | 10 mg/kg ** | Increased Nrf2 expression | TWN | [131] |
| Nervous system | fucoxanthin (43)/alga | Terpenoid f | Reduction in PC12 neurons intracellular ROS | 1 µM * | Binds to Keap1 | CHN | [132] |
| Nervous system | H. crispa peptides (109–111)/sea anemone | Peptide g | Inhibition of ASIC ion channels | 1.25–4.95 µM | rASIC1a ion channel inhibition | RUS | [133] |
| Nervous system | H. scabra 2-BTHF (112)/sea cucumber | Polyketide e | Aβ-induced C. elegans paralysis inhibition | 6.9 µM * | Decreased the formation of Ab oligomers and fibrils | THA | [134] |
| Nervous system | neo-debromoaplysiatoxins E and F (113, 114)/cyanobacterium | Terpenoid f/ Shikimate h | Kv1.5 inhibition | 1.22–2.85 µM | Binding to Kv1.5 S6 domain | CHN | [135] |
| Nervous system | okadaic acid (115)/ dinoflagellate | Polyketide e | Chick embryo neural tube defects | 0.5 µM * | Increased ROS, decreased Nrf2-signaling pathway | CHN | [136] |
| Nervous system | pinnatoxins A and G (116, 117)/dinoflagellate | Polyketide e | Synaptic transmission block at neuromuscular junction | 2.8–3.1 nmol/kg ** | AChE inhibition | FRA, USA | [137] |
| Nervous system | PFF-A (118)/alga | Polyketide e | hMAO-A inhibition | 9.2 µM | Noncompetitive inhibition | S. KOR | [138] |
| Nervous system | sargachromanol (119)/alga | Terpenoid f | AChE inhibition | 0.79 µM | Mixed reversible inhibition | S. KOR | [139] |
| Nervous system | santacruzamate A (120)/cyanobacterium | Alkaloid g | Amelioration of AD-like pathology | 10 mg/kg ** | Increased KDELR, decreased ER stress | CHN | [140] |
| Nervous system | Sinularia sp. cembranoid (121)/soft coral | Terpenoid f | Aβ42 inhibition | >10 µM | Binds to c-terminal of Ab monomer | CHN | [141] |
| Nervous system | S. latiuscula bromophenol (54)/alga | Shikimate h | HD3R inhibition | 18.7 µM | Binding to HD3R orthosteric site | S. KOR | [142] |
| Nervous system | S. japonica GM2 (122)/alga | Sugar | PC12 neurons increased viability | 270–540 µM | Increased autophagy factors; decreased pro-apoptotic factors | CHN | [143] |
| Nervous system | S. latiuscula bromophenol (54)/alga | Shikimate h | BACE1, AChE and BChe inhibition | 2.3–4.03 µM | Non-competitive or competitive inhibition | S. KOR | [144] |
| Nervous system | stelletin B (123)/sponge | Terpenoid f | Reversal of zebrafish locomotor deficiency | 1 nM * | Increased Nrf2/ARE signaling; decreased pro-apoptotic factors | TWN | [145] |
| Nervous system | androstatriol (124)/ soft coral | Terpenoid f | Retinal ganglion cells protection | 80 µg/eye ** | Negative regulation of Keap1 | CHN | [146] |





3.1. Antidiabetic Activity
3.2. Anti-Inflammatory Activity
3.3. Marine Compounds with Activity on the Immune System
3.4. Marine Compounds Affecting the Nervous System
4. Marine Compounds with Miscellaneous Mechanisms of Action
| Compound/Organism a | Chemistry | Pharmacological Activity | IC50 b | MMOA c | Country/ Territory d | References |
|---|---|---|---|---|---|---|
| amantamide (125)/ cyanobacterium | Peptide g | CXCR7 stimulation | 2.5 µM | Erk1/2 phosphorylation increase | CHN, PHL, USA | [148] |
| A.neglectus macrocyclic lactone (126)/octopus | Polyketide e | DPPH radical scavenging | 0.9 mM | ACE-1 non-competitive inhibition | IND | [149] |
| A. subcrenata peptides (127, 128)/shellfish | Peptide g | DPPH radical scavenging | 1 mM | Insulin/IGF-1 signaling modulation | CHN | [150] |
| aspermytin A (129)/fungus | Polyketide e | S. aureus-derived SrtA inhibition | 0.146 mM | Reversible mixed inhibition | S. KOR | [151] |
| avarol (130)/sponge | Terpenoid f | Cholesteryl ester synthesis inhibition | 5.7 µM | SOAT inhibition | JPN | [152] |
| bieckol (131)/alga | Polyketide e | Murine cholesterol, LDL and triglyceride decrease | 2.5 mg/kg/day ** | Aortic LOX-1 and PKC-α expression decreased | S. KOR | [153] |
| 3-BDB (132)/alga | Shikimate h | HO-1 antioxidant enzyme upregulation | 10 µM * | Nrf2/HO-1 pathway activation | S. KOR | [154] |
| C. gigas peptide (133)/oyster | Peptide g | Osteogenesis induction | 0.1 µM * | Integrin α5β1 binding | CHN | [155] |
| C. gigas peptide (134)/oyster | Peptide g | Thrombin inhibition | 3.6 mM * | Competitive inhibition | CHN | [156] |
| D. herbacea diphenyl ether (135)/sponge | Polyketide e | Bacterial α-d-galactosidase inhibition | 4.26 µM | Irreversible active-site inactivation | RUS | [157] |
| dieckol (65)/alga | Shikimate h | ROS inhibition | 0.5 µM * | Enhanced NFE2L and SOD1 gene expression | S. KOR | [158] |
| dieckol (65)/alga | Shikimate h | UVB-induced skin damage reduction | 25 µM * | Enhanced collagen synthesis and pro-inflammatory cytokines reduction | S. KOR | [159] |
| DPHC (136)/alga | Polyketide e | High-fat diet-induced adiposity inhibition | 25, 50 mg/kg/day ** | Lipogenesis enzymes inhibition | S. KOR | [160,161] |
| DHPC (136)/alga | Polyketide e | NO stimulation | 20 µM * | AchR and VEGFR2 expression activation | S. KOR | [162] |
| eckol (88)/alga | Polyketide e | ROS inhibition | 30 µM * | MAPK signaling inhibition | S. KOR | [164] |
| E. stolonifera phlorotannin (137)/alga | Polyketide e | Tyrosinase inhibition | 1.6 µM | Competitive inhibition | S. KOR | [163] |
| farnesylquinone (138)/fungus | Polyketide e | Lipid-lowering activity | 0.5 mM | Mitochondrial β-oxidation enhancement | CHN, DEU | [165] |
| fucofuroeckol-A (139)/alga | Polyketide e | Melanogenesis inhibition | 25 µM * | Tyrosinase-related protein-activity inhibition | JPN | [166] |
| fucoxanthin (43)/alga | Terpenoid f | ACE inhibition | 0.8 mM | Non-competitive inhibition | IND | [167] |
| fucoxanthin (43)/alga | Terpenoid f | Reduction in GMC’s collagen IV and fibronectin | 2 μM * | Akt/Sirt1/FoxO3α signaling regulation | CHN | [168] |
| funalenone (140)/fungus | Polyketide e | PTP1B inhibition | 6.1 μM | Non-competitive inhibition | S. KOR | [169] |
| GQQ-792 (141)/fungus | Alkaloid g | PGK1 inhibition | 1.2 μM | Non-competitive inhibition | CHN | [171] |
| grincamycin B (142)/fungus | Polyketide e | IDH1 inhibition | 1.25 μM * | Increased CHOP and GADD34 gene expression | CHN, USA | [170] |
| H. abdominalis peptides (143, 144)/seahorse | Peptide g | ROS inhibition in HUVEC | 0.23 and 0.17 mM * | Nrf2 signaling activation | S. KOR | [172] |
| (−)-loliolide (145)/alga | Terpenoid g | Lipid accumulation suppresion | 62 μM * | Decreased adipogenic protein expression | S. KOR | [173] |
| monanchomycalin B (146)/sponge | Alkaloid g | α-PsGal inhibition | Not shown | Slow-biding irreversible inhibition | RUS | [174] |
| monacolin X (147)/fungus | Polyketide e | HUVEC tube formation inhibition | 30 μM * | VEGFR2 signaling modulation | IND, SGP | [175] |
| (−)-muqubilin A (148)/sponge | Terpenoid f | RXRα and PPARα agonist | 10 μM * | Positive RARα allosteric modulation | CAN, ITA, USA | [176] |
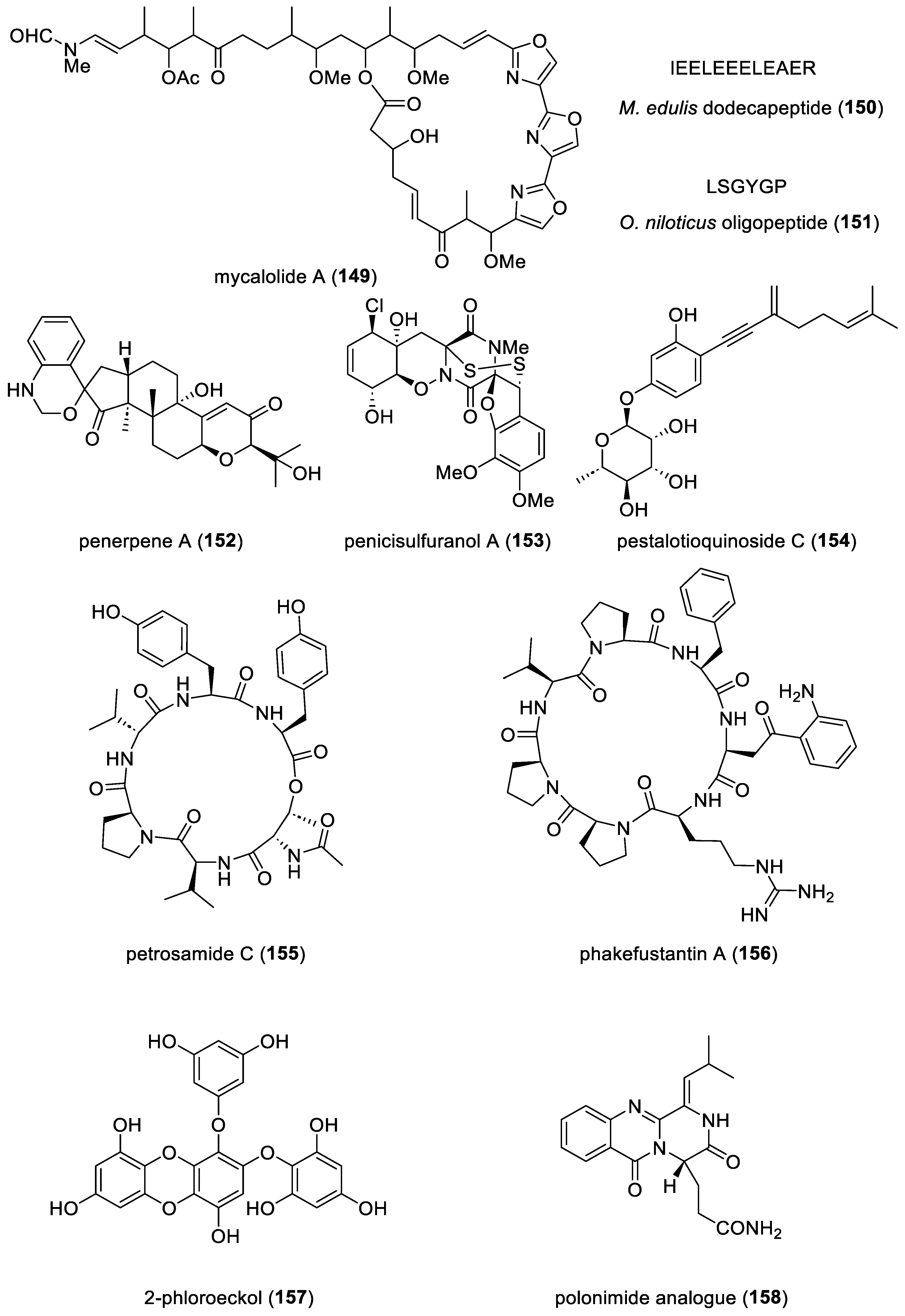
| mycalolide A (149)/sponge | Polyketide e | Cytokinesis inhibition | 11 µM | F actin inhibition and binucleation induction | JPN | [177] |
| M. edulis dodecapeptide (150)/mussel | Peptide g | Osteoblast growth stimulation | 67.2 µM | Binding to cellular 1L5G and 3V14 integrins | CHN | [178] |
| O. niloticus oligopeptide (151)/fish | Peptide g | NO and ROS inhibition | 10 μM * | NF-κB pathway suppression | CHN | [179] |
| penerpene A (152))/fungus | Terpenoid f | PTP inhibition | 1.7 μM | Docking studies completed | CHN | [180] |
| penicisulfuranol A (153)/fungus | Alkaloid g | Hsp90 inhibition | 0.5 μM | Binding to Hsp90α C-terminus | CHN | [181] |
| pestalotioquinoside C (154)/ fungus | Polyketide e | ABCA1 mRNA upregulation | 50 μM | LXRα receptor binding | CHN | [182] |
| petrosamide C (155)/fungus | Peptide g | Pancreatic lipase inhibition | 0.5 μM | Competitive inhibition | CHN | [183] |
| phakefustantin A (156) sponge | Peptide g | Akt expression inhibition | 10 µM * | RXR-α binding | CHN | [184] |
| 2-phloroeckol (157)/alga | Polyketide e | Tyrosinase inhibition | 7 µM | Slow-binding competitive inhibition | S. KOR | [185] |
| phlorofucofuroeckol A (118)/alga | Polyketide e | Collagen type 1 expression inhibition | 25 µM * | MAPK and SMAD 2/3 pathway downregulation | S. KOR | [186] |
| phlorofucofuroeckol A (118)/alga | Polyketide e | Osteoblastogenesis stimulation | 5 µM * | BMP and Wnt/β catenin- signaling activation | S. KOR | [187] |
| polonimide analogue (158)/ fungus | Alkaloid g | Insect GH18 chitinase OfChi-h inhibition | <1 µM * | Docking studies completed | CHN | [188] |
| P. morrowii bromophenol (132)/alga | Shikimate h | Adipogenesis inhibition | 25 µM * | PPAR-γ, C/EBPα, leptin inhibition and AMPK enhancement | S. KOR | [189] |
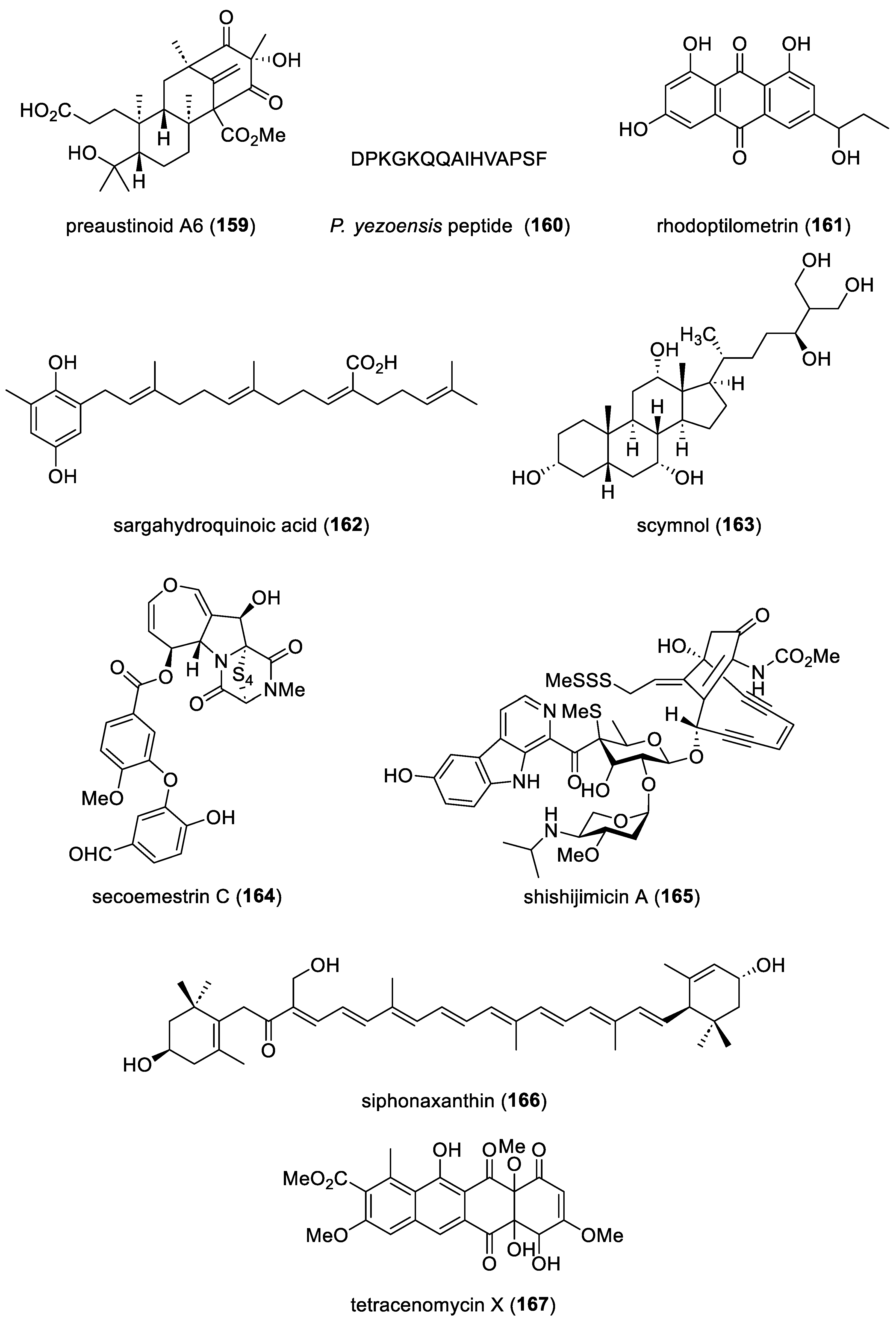
| preaustinoid A6 (159)/fungus | Terpenoid f | PTP inhibition | 17.6 µM | Non-competitive inhibition | S. KOR, VNM | [190] |
| P. yezoensis peptide (160)/alga | Peptide g | Dexamethasone-induced atrophy protection | 0.31 µM | IFG-1 signaling activation | S. KOR | [191] |
| rhodoptilometrin (161)/crinoid | Polyketide e | Wound healing and cell migration stimulation | 1 µM * | FAK, fibronectin and type 1 collagen increased | TWN | [192] |
| sargahydroquinoic acid (162)/alga | Terpenoid f | Activation of lipid catabolism | 2.5 µM * | PPAR-γ and AMPKα activation | S. KOR | [193] |
| scymnol (163)/shark | Terpenoid f | Activation of TGR5 receptor | 0.5 mM * | Sustained intracellular Ca2+ release | AUS | [194] |
| secoemestrin C (164)/fungus | Alkaloid g | ICL inhibition | 4.77 µM | ICL mRNA expression inhibition | S. KOR | [195] |
| shishijimicin A (165)/ascidian | Alkaloid g | DNA cleavage | 0.014 µM | Binding to double-stranded DNA minor groove | GRC, SGP, USA, | [196] |
| siphonaxanthin (166)/alga | Terpenoid f | Cellular Nrf2 protein expression activation | 1 μM * | Nrf2 signaling activation | JPN | [197] |
| S. latiuscula bromophenol (54)/alga | Polyketide e | Tyrosinase inhibition | 2.9 µM | Competitive inhibition | S. KOR | [198] |
| tetracenomycin X (167)/bacterium | Polyketide e | Cyclin D1 downregulation | 2.5 µM * | Cyclin D1 proteosomal degradation | CHN | [199] |
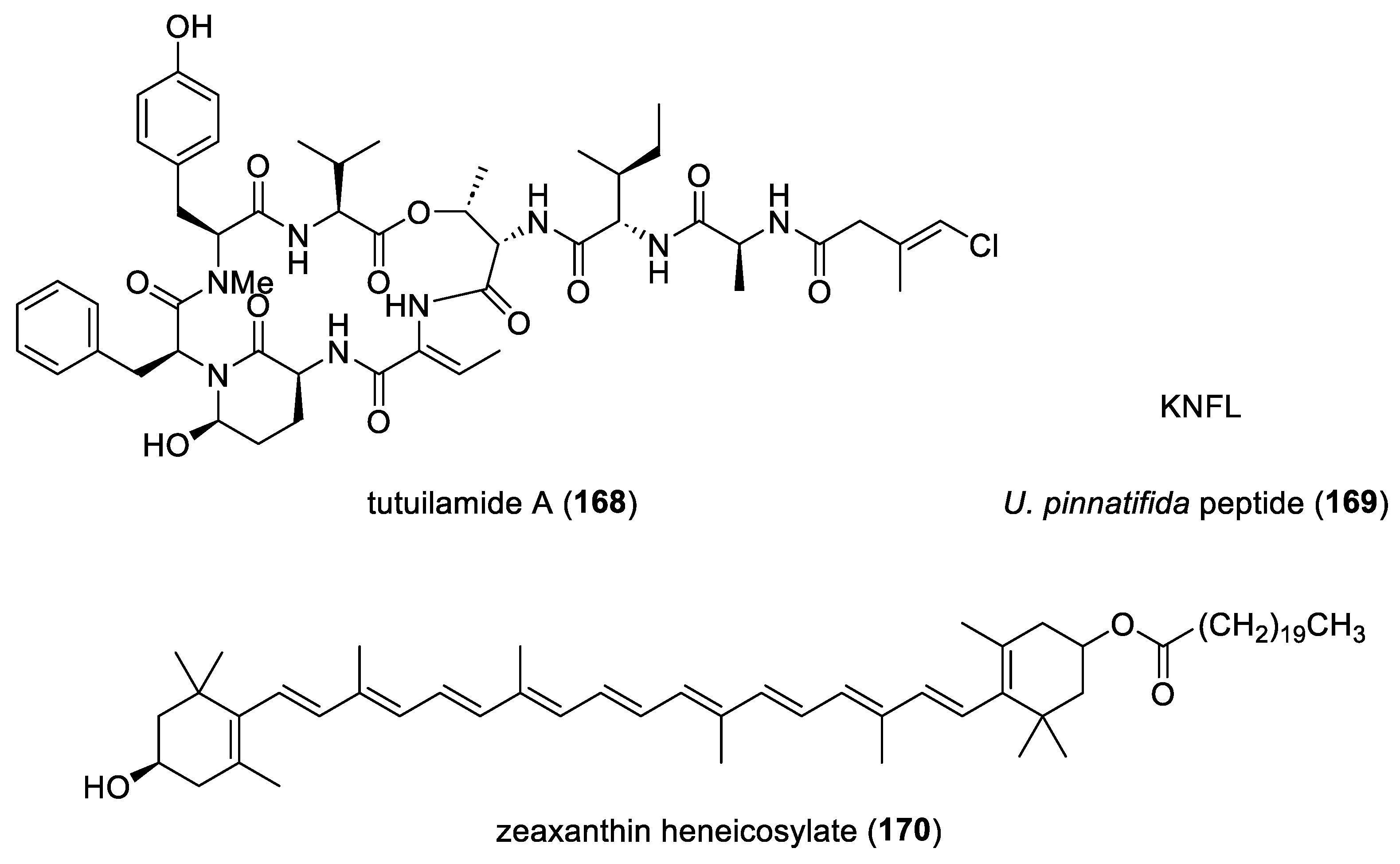
| tutuilamide A (168)/ cyanobacterium | Peptide g | Elastase inhibition | 0.001 µM | Docking studies completed | BRA, CHN, DEU, USA | [200] |
| U. pinnatifida peptide (169)/alga | Peptide g | ACE inhibition | 225 µM | Mixed-type inhibition | CHN | [201] |
| zeaxanthin heneicosylate (170)/alga | Terpenoid f | In vivo inhibition of age-associated cardiac dysfunction | 250 µg/kg ** | RXR-α activation | EGY | [202] |


5. Reviews on Marine Pharmacology and Pharmaceuticals
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mayer, A.M.S.; Lehmann, V.K.B. Marine pharmacology in 1998: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, anthelmintic, antiplatelet, antiprotozoal, and antiviral activities; with actions on the cardiovascular, endocrine, immune, and nervous systems; and other miscellaneous mechanisms of action. Pharmacologist 2000, 42, 62–69. [Google Scholar]
- Mayer, A.M.S.; Hamann, M.T. Marine pharmacology in 1999: Compounds with antibacterial, anticoagulant, antifungal, anthelmintic, anti-inflammatory, antiplatelet, antiprotozoal and antiviral activities affecting the cardiovascular, endocrine, immune and nervous systems, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2002, 132, 315–339. [Google Scholar] [PubMed]
- Mayer, A.M.S.; Hamann, M.T. Marine pharmacology in 2000: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antituberculosis, and antiviral activities; affecting the cardiovascular, immune, and nervous systems and other miscellaneous mechanisms of action. Mar. Biotechnol. 2004, 6, 37–52. [Google Scholar]
- Mayer, A.M.S.; Hamann, M.T. Marine pharmacology in 2001–2002: Marine compounds with anthelmintic, antibacterial, anticoagulant, antidiabetic, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 140, 265–286. [Google Scholar] [PubMed]
- Mayer, A.M.S.; Rodriguez, A.D.; Berlinck, R.G.; Hamann, M.T. Marine pharmacology in 2003–2004: Marine compounds with anthelmintic antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 145, 553–581. [Google Scholar] [PubMed]
- Mayer, A.M.S.; Rodriguez, A.D.; Berlinck, R.G.; Hamann, M.T. Marine pharmacology in 2005–2006: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim. Biophys. Acta 2009, 1790, 283–308. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Rodriguez, A.D.; Berlinck, R.G.; Fusetani, N. Marine pharmacology in 2007–2008: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous system, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 191–222. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine Pharmacology in 2009–2011: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Immune and Nervous Systems, and other Miscellaneous Mechanisms of Action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine Pharmacology in 2012–2013: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2017, 15, 273. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Guerrero, A.J.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine Pharmacology in 2014–2015: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, Antiviral, and Anthelmintic Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2019, 18, 5. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Guerrero, A.J.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine Pharmacology in 2016–2017: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2021, 19, 49. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Pierce, M.L.; Howe, K.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine pharmacology in 2018: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Pharmacol. Res. 2022, 183, 106391. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, F.J.; Bowden, B.F.; Toth, S.I. Antitumor and Cytotoxic Compounds from Marine Organisms. Pharm. Bioact. Nat. Prod. 1993, 1, 197–308. [Google Scholar]
- Gomez-Rodriguez, L.; Schultz, P.J.; Tamayo-Castillo, G.; Dotson, G.D.; Sherman, D.H.; Tripathi, A. Adipostatins E-J, New Potent Antimicrobials Identified as Inhibitors of Coenzyme—A Biosynthesis. Tetrahedron Lett. 2020, 61, 151469. [Google Scholar] [CrossRef] [PubMed]
- Elliott, A.G.; Huang, J.X.; Neve, S.; Zuegg, J.; Edwards, I.A.; Cain, A.K.; Boinett, C.J.; Barquist, L.; Lundberg, C.V.; Steen, J.; et al. An amphipathic peptide with antibiotic activity against multidrug-resistant Gram-negative bacteria. Nat. Commun. 2020, 11, 3184. [Google Scholar] [CrossRef] [PubMed]
- Paderog, M.J.V.; Suarez, A.F.L.; Sabido, E.M.; Low, Z.J.; Saludes, J.P.; Dalisay, D.S. Anthracycline Shunt Metabolites from Philippine Marine Sediment-Derived Streptomyces Destroy Cell Membrane Integrity of Multidrug-Resistant Staphylococcus aureus. Front. Microbiol. 2020, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhao, L.; Wu, H.; Zhao, C.; Gong, Q.; Yu, W. Cladodionen Is a Potential Quorum Sensing Inhibitor against Pseudomonas aeruginosa. Mar. Drugs 2020, 18, 205. [Google Scholar] [CrossRef] [PubMed]
- Gowrishankar, S.; Pandian, S.K.; Balasubramaniam, B.; Balamurugan, K. Quorum quelling efficacy of marine cyclic dipeptide -cyclo(l-leucyl-l-prolyl) against the uropathogen Serratia marcescens. Food Chem. Toxicol. 2019, 123, 326–336. [Google Scholar] [CrossRef] [PubMed]
- de Figueiredo, C.S.; Menezes Silva, S.M.P.; Abreu, L.S.; da Silva, E.F.; da Silva, M.S.; Cavalcanti de Miranda, G.E.; Costa, V.C.O.; Le Hyaric, M.; Siqueira Junior, J.P.; Barbosa Filho, J.M.; et al. Dolastane diterpenes from Canistrocarpus cervicornis and their effects in modulation of drug resistance in Staphylococcus aureus. Nat. Prod. Res. 2019, 33, 3231–3239. [Google Scholar] [CrossRef]
- Davison, J.R.; Bewley, C.A. Antimicrobial Chrysophaentin Analogs Identified from Laboratory Cultures of the Marine Microalga Chrysophaeum taylorii. J. Nat. Prod. 2019, 82, 148–153. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Sun, Y.; Sun, L. A Crustin from Hydrothermal Vent Shrimp: Antimicrobial Activity and Mechanism. Mar. Drugs 2021, 19, 176. [Google Scholar] [CrossRef]
- Campana, R.; Favi, G.; Baffone, W.; Lucarini, S. Marine Alkaloid 2,2-Bis(6-bromo-3-indolyl) Ethylamine and Its Synthetic Derivatives Inhibit Microbial Biofilms Formation and Disaggregate Developed Biofilms. Microorganisms 2019, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Matthew, S.; Chen, Q.Y.; Kwan, J.C.; Paul, V.J.; Luesch, H. Discovery and Total Synthesis of Doscadenamide A: A Quorum Sensing Signaling Molecule from a Marine Cyanobacterium. Org. Lett. 2019, 21, 7274–7278. [Google Scholar] [CrossRef] [PubMed]
- Jabila Mary, T.R.; Kannan, R.R.; Muthamil Iniyan, A.; Carlton Ranjith, W.A.; Nandhagopal, S.; Vishwakarma, V.; Prakash Vincent, S.G. β-lactamase inhibitory potential of kalafungin from marine Streptomyces in Staphylococcus aureus infected zebrafish. Microbiol. Res. 2021, 244, 126666. [Google Scholar] [CrossRef] [PubMed]
- Maynard, A.; Butler, N.L.; Ito, T.; da Silva, A.J.; Murai, M.; Chen, T.; Koffas, M.A.G.; Miyoshi, H.; Barquera, B. Antibiotic Korormicin A Kills Bacteria by Producing Reactive Oxygen Species. J. Bacteriol. 2019, 201, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.; Kwon, O.S.; Shin, J.; Oh, K.B. Antibacterial Activity and Mode of Action of Lactoquinomycin A from Streptomyces bacillaris. Mar. Drugs 2020, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Thulshan Jayathilaka, E.H.T.; Liyanage, T.D.; Rajapaksha, D.C.; Dananjaya, S.H.S.; Nikapitiya, C.; Whang, I.; De Zoysa, M. Octominin: An antibacterial and anti-biofilm peptide for controlling the multidrug resistance and pathogenic Streptococcus parauberis. Fish Shellfish Immunol. 2021, 110, 23–34. [Google Scholar] [CrossRef]
- Yu, X.; Li, L.; Sun, S.; Chang, A.; Dai, X.; Li, H.; Wang, Y.; Zhu, H. A Cyclic Dipeptide from Marine Fungus Penicillium chrysogenum DXY-1 Exhibits Anti-quorum Sensing Activity. ACS Omega 2021, 6, 7693–7700. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zheng, L.B.; Mao, Y.; Wang, J.; Lin, L.S.; Su, Y.Q.; Li, Y. The antibacterial activity and mechanism analysis of piscidin 5 like from Larimichthys crocea. Dev. Comp. Immunol. 2019, 92, 43–49. [Google Scholar] [CrossRef]
- Kim, Y.G.; Lee, J.H.; Lee, S.; Lee, Y.K.; Hwang, B.S.; Lee, J. Antibiofilm Activity of Phorbaketals from the Marine Sponge Phorbas sp. against Staphylococcus aureus. Mar. Drugs 2021, 19, 301. [Google Scholar] [CrossRef]
- Kizhakkekalam, V.K.; Chakraborty, K.; Joy, M. Antibacterial and antioxidant aryl-enclosed macrocyclic polyketide from intertidal macroalgae associated heterotrophic bacterium Shewanella algae. Med. Chem. Res. 2020, 29, 145–155. [Google Scholar] [CrossRef]
- Chakraborty, K.; Kizhakkekalam, V.K.; Joy, M. Macrocyclic polyketides with siderophore mode of action from marine heterotrophic Shewanella algae: Prospective anti-infective leads attenuate drug-resistant pathogens. J. Appl. Microbiol. 2021, 130, 1552–1570. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.Ø.; Hansen, I.K.Ø.; Richard, C.S.; Jenssen, M.; Andersen, J.H.; Hansen, E.H. Antimicrobial activity of securamines from the bryozoan Securiflustra securifrons. Nat. Prod. Commun. 2021, 16, 1934578X21996180. [Google Scholar] [CrossRef]
- Hansen, I.K.O.; Isaksson, J.; Poth, A.G.; Hansen, K.O.; Andersen, A.J.C.; Richard, C.S.M.; Blencke, H.M.; Stensvåg, K.; Craik, D.J.; Haug, T. Isolation and Characterization of Antimicrobial Peptides with Unusual Disulfide Connectivity from the Colonial Ascidian Synoicum turgens. Mar. Drugs 2020, 18, 51. [Google Scholar] [CrossRef] [PubMed]
- Reina, J.C.; Perez-Victoria, I.; Martin, J.; Llamas, I. A Quorum-Sensing Inhibitor Strain of Vibrio alginolyticus Blocks Qs-Controlled Phenotypes in Chromobacterium violaceum and Pseudomonas aeruginosa. Mar. Drugs 2019, 17, 494. [Google Scholar] [CrossRef]
- Elsadek, L.A.; Matthews, J.H.; Nishimura, S.; Nakatani, T.; Ito, A.; Gu, T.; Luo, D.; Salvador-Reyes, L.A.; Paul, V.J.; Kakeya, H.; et al. Genomic and Targeted Approaches Unveil the Cell Membrane as a Major Target of the Antifungal Cytotoxin Amantelide A. Chembiochem 2021, 22, 1790–1799. [Google Scholar] [CrossRef]
- Yang, B.; He, Y.; Lin, S.; Zhang, J.; Li, H.; Wang, J.; Hu, Z.; Zhang, Y. Antimicrobial Dolabellanes and Atranones from a Marine-Derived Strain of the Toxigenic Fungus Stachybotrys chartarum. J. Nat. Prod. 2019, 82, 1923–1929. [Google Scholar] [CrossRef]
- Tang, X.X.; Yan, X.; Fu, W.H.; Yi, L.Q.; Tang, B.W.; Yu, L.B.; Fang, M.J.; Wu, Z.; Qiu, Y.K. New β-Lactone with Tea Pathogenic Fungus Inhibitory Effect from Marine-Derived Fungus MCCC3A00957. J. Agric. Food Chem. 2019, 67, 2877–2885. [Google Scholar] [CrossRef]
- Kim, H.; Hwang, J.Y.; Chung, B.; Cho, E.; Bae, S.; Shin, J.; Oh, K.B. 2-Alkyl-4-hydroxyquinolines from a Marine-Derived Streptomyces sp. Inhibit Hyphal Growth Induction in Candida albicans. Mar. Drugs 2019, 17, 133. [Google Scholar] [CrossRef]
- Dalisay, D.S.; Rogers, E.W.; Molinski, T.F. Oceanapiside, a Marine Natural Product, Targets the Sphingolipid Pathway of Fluconazole-Resistant Candida glabrata. Mar. Drugs 2021, 19, 126. [Google Scholar] [CrossRef]
- Tripathi, S.K.; Feng, Q.; Liu, L.; Levin, D.E.; Roy, K.K.; Doerksen, R.J.; Baerson, S.R.; Shi, X.; Pan, X.; Xu, W.H.; et al. Puupehenone, a Marine-Sponge-Derived Sesquiterpene Quinone, Potentiates the Antifungal Drug Caspofungin by Disrupting Hsp90 Activity and the Cell Wall Integrity Pathway. mSphere 2020, 5, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Sun, C.; Zhang, C.; Song, S.; Sun, X.; Ju, J.; Deng, Y. Efficacy of Compounds Isolated from Streptomyces olivaceus against the Morphogenesis and Virulence of Candida albicans. Mar. Drugs 2019, 17, 442. [Google Scholar] [CrossRef] [PubMed]
- Alhadrami, H.A.; Thissera, B.; Hassan, M.H.A.; Behery, F.A.; Ngwa, C.J.; Hassan, H.M.; Pradel, G.; Abdelmohsen, U.R.; Rateb, M.E. Bio-Guided Isolation of Antimalarial Metabolites from the Coculture of Two Red Sea Sponge-Derived Actinokineospora and Rhodococcus spp. Mar. Drugs 2021, 19, 109. [Google Scholar] [CrossRef] [PubMed]
- Sweeney-Jones, A.M.; Gagaring, K.; Antonova-Koch, J.; Zhou, H.; Mojib, N.; Soapi, K.; Skolnick, J.; McNamara, C.W.; Kubanek, J. Antimalarial Peptide and Polyketide Natural Products from the Fijian Marine Cyanobacterium Moorea producens. Mar. Drugs 2020, 18, 167. [Google Scholar] [CrossRef] [PubMed]
- Knestrick, M.A.; Wilson, N.G.; Roth, A.; Adams, J.H.; Baker, B.J. Friomaramide, a Highly Modified Linear Hexapeptide from an Antarctic Sponge, Inhibits Plasmodium falciparum Liver-Stage Development. J. Nat. Prod. 2019, 82, 2354–2358. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.E.; Collins, J.E.; Roberts, B.; Roberts, J.C.; Winder, P.L.; Reed, J.K.; Diaz, M.C.; Pomponi, S.A.; Chakrabarti, D. Antiplasmodial Compounds from Deep-Water Marine Invertebrates. Mar. Drugs 2021, 19, 179. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Expósito, R.L.; Nocchi, N.; Reyes-Batlle, M.; Sifaoui, I.; Suárez-Gómez, B.; Díaz-Marrero, A.R.; Souto, M.L.; Piñero, J.E.; Fernández, J.J.; Lorenzo-Morales, J. Antiamoebic effects of sesquiterpene lactones isolated from the zoanthid Palythoa aff. clavata. Bioorg. Chem. 2021, 108, 104682. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.N.; Chuang, C.M.; Chen, J.Y.; Chieh-Yu, P. Epinecidin-1: A marine fish antimicrobial peptide with therapeutic potential against Trichomonas vaginalis infection in mice. Peptides 2019, 112, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.L.; Romanelli, M.M.; Borborema, S.E.T.; Johns, D.M.; Migotto, A.E.; Lago, J.H.G.; Tempone, A.G. Antitrypanosomal activity of isololiolide isolated from the marine hydroid Macrorhynchia philippina (Cnidaria, Hydrozoa). Bioorg. Chem. 2019, 89, 103002. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Diaz-Marrero, A.R.; Cen-Pacheco, F.; Sifaoui, I.; Reyes-Batlle, M.; Souto, M.L.; Hernandez Daranas, A.; Pinero, J.E.; Fernandez, J.J. Evaluation of Oxasqualenoids from the Red Alga Laurencia viridis against Acanthamoeba. Mar. Drugs 2019, 17, 420. [Google Scholar] [CrossRef]
- Boudreau, P.D.; Miller, B.W.; McCall, L.I.; Almaliti, J.; Reher, R.; Hirata, K.; Le, T.; Siqueira-Neto, J.L.; Hook, V.; Gerwick, W.H. Design of Gallinamide A Analogs as Potent Inhibitors of the Cysteine Proteases Human Cathepsin L and Trypanosoma cruzi Cruzain. J. Med. Chem. 2019, 62, 9026–9044. [Google Scholar] [CrossRef] [PubMed]
- Cartuche, L.; Reyes-Batlle, M.; Sifaoui, I.; Arberas-Jimenez, I.; Pinero, J.E.; Fernandez, J.J.; Lorenzo-Morales, J.; Diaz-Marrero, A.R. Antiamoebic Activities of Indolocarbazole Metabolites Isolated from Streptomyces sanyensis Cultures. Mar. Drugs 2019, 17, 588. [Google Scholar] [CrossRef] [PubMed]
- Casertano, M.; Imperatore, C.; Luciano, P.; Aiello, A.; Putra, M.Y.; Gimmelli, R.; Ruberti, G.; Menna, M. Chemical Investigation of the Indonesian Tunicate Polycarpa aurata and Evaluation of the Effects against Schistosoma mansoni of the Novel Alkaloids Polyaurines A and B. Mar. Drugs 2019, 17, 278. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, Q.; Li, S.; Cui, H.; Sun, Z.; Chen, D.; Lu, Y.; Liu, H.; Zhang, W. Polypropionate Derivatives with Mycobacterium tuberculosis Protein Tyrosine Phosphatase B Inhibitory Activities from the Deep-Sea-Derived Fungus Aspergillus fischeri FS452. J. Nat. Prod. 2019, 82, 3440–3449. [Google Scholar] [CrossRef] [PubMed]
- Sudomova, M.; Shariati, M.A.; Echeverria, J.; Berindan-Neagoe, I.; Nabavi, S.M.; Hassan, S.T.S. A Microbiological, Toxicological, and Biochemical Study of the Effects of Fucoxanthin, a Marine Carotenoid, on Mycobacterium tuberculosis and the Enzymes Implicated in Its Cell Wall: A Link between Mycobacterial Infection and Autoimmune Diseases. Mar. Drugs 2019, 17, 641. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Li, Y.; Guo, X.; Ji, W.; Lin, W. Chartarlactams Q-T, Dimeric Phenylspirodrimanes with Antibacterial and Antiviral Activities. Chem. Biodivers. 2020, 17, e2000170. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, L.; Peng, Z.; Liu, D.; Si, L.; Wang, J.; Yuan, B.; Huang, J.; Proksch, P.; Lin, W. Harzianoic acids A and B, new natural scaffolds with inhibitory effects against hepatitis C virus. Bioorg. Med. Chem. 2019, 27, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.C.; Lehman, C.W.; Stewart, A.K.; Panny, L.; Bracci, N.; Wright, J.L.C.; Paige, M.; Strangman, W.K.; Kehn-Hall, K. Homoseongomycin, a compound isolated from marine actinomycete bacteria K3-1, is a potent inhibitor of encephalitic alphaviruses. Antivir. Res. 2021, 191, 105087. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Yang, B.; Liu, J.; Xun, T.; Liu, Y.; Zhou, X. Penicillixanthone A, a marine-derived dual-coreceptor antagonist as anti-HIV-1 agent. Nat. Prod. Res. 2019, 33, 1467–1471. [Google Scholar] [CrossRef]
- Izumida, M.; Suga, K.; Ishibashi, F.; Kubo, Y. The Spirocyclic Imine from a Marine Benthic Dinoflagellate, Portimine, Is a Potent Anti-Human Immunodeficiency Virus Type 1 Therapeutic Lead Compound. Mar. Drugs 2019, 17, 495. [Google Scholar] [CrossRef]
- Das, S.K.; Samantaray, D.; Sahoo, S.K.; Pradhan, S.K.; Samanta, L.; Thatoi, H. Bioactivity guided isolation of antidiabetic and antioxidant compound from Xylocarpus granatum J. Koenig bark. 3 Biotech 2019, 9, 198. [Google Scholar] [CrossRef]
- Luo, J.; Jiang, B.; Li, C.; Jia, X.; Shi, D. CYC27 Synthetic Derivative of Bromophenol from Red Alga Rhodomela confervoides: Anti-Diabetic Effects of Sensitizing Insulin Signaling Pathways and Modulating RNA Splicing-Associated RBPs. Mar. Drugs 2019, 17, 49. [Google Scholar] [CrossRef] [PubMed]
- Zaharudin, N.; Staerk, D.; Dragsted, L.O. Inhibition of α-glucosidase activity by selected edible seaweeds and fucoxanthin. Food Chem. 2019, 270, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Kawee-Ai, A.; Kim, A.; Kim, S. Inhibitory activities of microalgal fucoxanthin against α-amylase, α-glucosidase, and glucose oxidase in 3T3-L1 cells linked to type 2 diabetes. J. Oceanol. Limnol. 2019, 37, 928–937. [Google Scholar] [CrossRef]
- Hudlikar, R.R.; Sargsyan, D.; Li, W.; Wu, R.; Zheng, M.; Kong, A.N. Epigenomic, Transcriptomic, and Protective Effect of Carotenoid Fucoxanthin in High Glucose-Induced Oxidative Stress in Mes13 Kidney Mesangial Cells. Chem. Res. Toxicol. 2021, 34, 713–722. [Google Scholar] [CrossRef]
- Chakraborty, K.; Antony, T. First report of antioxidative abeo-oleanenes from red seaweed Gracilaria salicornia as dual inhibitors of starch digestive enzymes. Med. Chem. Res. 2019, 28, 696–710. [Google Scholar] [CrossRef]
- Yang, H.W.; Son, M.; Choi, J.; Oh, S.; Jeon, Y.J.; Byun, K.; Ryu, B. Effect of Ishophloroglucin A, A Component of Ishige okamurae, on Glucose Homeostasis in the Pancreas and Muscle of High Fat Diet-Fed Mice. Mar. Drugs 2019, 17, 608. [Google Scholar] [CrossRef] [PubMed]
- Paudel, P.; Seong, S.H.; Park, H.J.; Jung, H.A.; Choi, J.S. Anti-Diabetic Activity of 2,3,6-Tribromo-4,5-Dihydroxybenzyl Derivatives from Symphyocladia latiuscula through PTP1B Downregulation and α-Glucosidase Inhibition. Mar. Drugs 2019, 17, 166. [Google Scholar] [CrossRef]
- Seong, S.H.; Nguyen, D.H.; Wagle, A.; Woo, M.H.; Jung, H.A.; Choi, J.S. Experimental and Computational Study to Reveal the Potential of Non-Polar Constituents from Hizikia fusiformis as Dual Protein Tyrosine Phosphatase 1B and α-Glucosidase Inhibitors. Mar. Drugs 2019, 17, 302. [Google Scholar] [CrossRef]
- Lopez, D.; Cherigo, L.; Mejia, L.C.; Loza-Mejia, M.A.; Martinez-Luis, S. α-Glucosidase inhibitors from a mangrove associated fungus, Zasmidium sp. strain EM5-10. BMC Chem. 2019, 13, 22. [Google Scholar] [CrossRef]
- Pereira, R.B.; Pereira, D.M.; Jimenez, C.; Rodriguez, J.; Nieto, R.M.; Videira, R.A.; Silva, O.; Andrade, P.B.; Valentao, P. Anti-Inflammatory Effects of 5α,8α-Epidioxycholest-6-en-3β-ol, a Steroidal Endoperoxide Isolated from Aplysia depilans, Based on Bioguided Fractionation and NMR Analysis. Mar. Drugs 2019, 17, 330. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Chen, C.; Sun, W.; Zang, Y.; Li, Q.; Wang, W.; Zeng, F.; Liu, J.; Zhou, Y.; Zhou, Q.; et al. Phenolic C-Glycosides and Aglycones from Marine-Derived Aspergillus sp. and Their Anti-Inflammatory Activities. J. Nat. Prod. 2019, 82, 1098–1106. [Google Scholar] [CrossRef] [PubMed]
- Keeler, D.M.; Grandal, M.K.; McCall, J.R. Brevenal, a Marine Natural Product, is Anti-Inflammatory and an Immunomodulator of Macrophage and Lung Epithelial Cells. Mar. Drugs 2019, 17, 184. [Google Scholar] [CrossRef] [PubMed]
- Alvarino, R.; Alonso, E.; Lacret, R.; Oves-Costales, D.; Genilloud, O.; Reyes, F.; Alfonso, A.; Botana, L.M. Caniferolide A, a Macrolide from Streptomyces caniferus, Attenuates Neuroinflammation, Oxidative Stress, Amyloid-Beta, and Tau Pathology In Vitro. Mol. Pharm. 2019, 16, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Van Thanh, N.; Jang, H.J.; Vinh, L.B.; Linh, K.T.P.; Huong, P.T.T.; Cuong, N.X.; Nam, N.H.; Van Minh, C.; Kim, Y.H.; Yang, S.Y. Chemical constituents from Vietnamese mangrove Calophyllum inophyllum and their anti-inflammatory effects. Bioorg. Chem. 2019, 88, 102921. [Google Scholar] [CrossRef]
- Ding, Y.; An, F.; Zhu, X.; Yu, H.; Hao, L.; Lu, Y. Curdepsidones B–G, Six Depsidones with Anti-Inflammatory Activities from the Marine-Derived Fungus Curvularia sp. IFB-Z10. Mar. Drugs 2019, 17, 266. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.K.; Jeong, S.Y.; Yang, S.; Kim, K.M.; Choi, H.; Bae, J.S. Suppressive effects of collismycin C on polyphosphate-mediated vascular inflammatory responses. Fitoterapia 2019, 134, 447–453. [Google Scholar] [CrossRef]
- Oh, S.; Son, M.; Byun, K.A.; Jang, J.T.; Choi, C.H.; Son, K.H.; Byun, K. Attenuating Effects of Dieckol on High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease by Decreasing the NLRP3 Inflammasome and Pyroptosis. Mar. Drugs 2021, 19, 318. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.Y.; Zhang, H.; Chen, Y.Y.; Jiao, W.H.; Fan, J.T.; Liu, Z.Q.; Lin, H.W.; Cheng, B.H. Dysiarenone from Marine Sponge Dysidea arenaria Attenuates ROS and Inflammation via Inhibition of 5-LOX/NF-κB/MAPKs and Upregulation of Nrf-2/OH-1 in RAW 264.7 Macrophages. J. Inflamm. Res. 2021, 14, 587–597. [Google Scholar] [CrossRef]
- Kim, E.N.; Nabende, W.Y.; Jeong, H.; Hahn, D.; Jeong, G.S. The marine-derived natural product epiloliolide isolated from Sargassum horneri regulates NLRP3 via PKA/CREB, promoting proliferation and anti-inflammatory effects of human periodontal ligament cells. Mar. Drugs 2021, 19, 388. [Google Scholar] [CrossRef]
- Su, J.; Guo, K.; Huang, M.; Liu, Y.; Zhang, J.; Sun, L.; Li, D.; Pang, K.L.; Wang, G.; Chen, L.; et al. Fucoxanthin, a Marine Xanthophyll Isolated from Conticribra weissflogii ND-8: Preventive Anti-Inflammatory Effect in a Mouse Model of Sepsis. Front. Pharmacol. 2019, 10, 906. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Tian, X.; Zhang, W.; Zheng, P.; Huang, F.; Ding, G.; Yang, Z. Protective Effects of Fucoxanthin against Alcoholic Liver Injury by Activation of Nrf2-Mediated Antioxidant Defense and Inhibition of TLR4-Mediated Inflammation. Mar. Drugs 2019, 17, 552. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.J.; Choi, Y.S.; Oh, Y.R.; Kang, E.H.; Khang, G.; Park, Y.B.; Lee, Y.J. Fucoxanthin Suppresses Osteoclastogenesis via Modulation of MAP Kinase and Nrf2 Signaling. Mar. Drugs 2021, 19, 132. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, R.; Liu, K.; Li, M.; Luo, H.; Cui, L.; Huang, L.; Luo, L. Fucoxanthin attenuates LPS-induced acute lung injury via inhibition of the TLR4/MyD88 signaling axis. Aging 2020, 13, 2655–2667. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, L.; Sun, P.; Zhang, Y.; Wu, T.; Sun, H.; Cheng, K.W.; Chen, F. Fucoxanthinol from the Diatom Nitzschia Laevis Ameliorates Neuroinflammatory Responses in Lipopolysaccharide-Stimulated BV-2 Microglia. Mar. Drugs 2020, 18, 116. [Google Scholar] [CrossRef] [PubMed]
- Jan, J.S.; Yang, C.H.; Wang, M.H.; Lin, F.L.; Yen, J.L.; Hsieh, I.; Khotimchenko, M.; Lee, T.H.; Hsiao, G. Hirsutanol A Attenuates Lipopolysaccharide-Mediated Matrix Metalloproteinase 9 Expression and Cytokines Production and Improves Endotoxemia-Induced Acute Sickness Behavior and Acute Lung Injury. Mar. Drugs 2019, 17, 360. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yang, H.; Zhi, Y.; Yao, Q.; Liu, B. Evaluation of pyrrolidine-based analog of jaspine B as potential SphK1 inhibitors against rheumatoid arthritis. Bioorg. Med. Chem. Lett. 2021, 34, 127754. [Google Scholar] [CrossRef]
- Daskalaki, M.G.; Vyrla, D.; Harizani, M.; Doxaki, C.; Eliopoulos, A.G.; Roussis, V.; Ioannou, E.; Tsatsanis, C.; Kampranis, S.C. Neorogioltriol and Related Diterpenes from the Red Alga Laurencia Inhibit Inflammatory Bowel Disease in Mice by Suppressing M1 and Promoting M2-Like Macrophage Responses. Mar. Drugs 2019, 17, 97. [Google Scholar] [CrossRef] [PubMed]
- Herath, K.; Kim, H.J.; Jang, J.H.; Kim, H.S.; Kim, H.J.; Jeon, Y.J.; Jee, Y. Mojabanchromanol Isolated from Sargassum horneri Attenuates Particulate Matter Induced Inflammatory Responses via Suppressing TLR2/4/7-MAPK Signaling in MLE-12 Cells. Mar. Drugs 2020, 18, 355. [Google Scholar] [CrossRef]
- Ha, T.M.; Kim, D.C.; Sohn, J.H.; Yim, J.H.; Oh, H. Anti-Inflammatory and Protein Tyrosine Phosphatase 1B Inhibitory Metabolites from the Antarctic Marine-Derived Fungal Strain Penicillium glabrum SF-7123. Mar. Drugs 2020, 18, 247. [Google Scholar] [CrossRef]
- Kim, Y.N.; Ji, Y.K.; Kim, N.H.; Van Tu, N.; Rho, J.R.; Jeong, E.J. Isoquinolinequinone Derivatives from a Marine Sponge (Haliclona sp.) Regulate Inflammation in In Vitro System of Intestine. Mar. Drugs 2021, 19, 90. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.C.; Chang, C.H.; Liao, H.R.; Cheng, M.J.; Wu, M.D.; Fu, S.L.; Chen, J.J. Rare Chromone Derivatives from the Marine-Derived Penicillium citrinum with Anti-Cancer and Anti-Inflammatory Activities. Mar. Drugs 2021, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kim, N.H.; Lee, S.; Kim, Y.N.; Heo, J.D.; Jeong, E.J.; Rho, J.R. Deacetylphylloketal, a New Phylloketal Derivative from a Marine Sponge, Genus Phyllospongia, with Potent Anti-Inflammatory Activity in In Vitro Co-Culture Model of Intestine. Mar. Drugs 2019, 17, 634. [Google Scholar] [CrossRef]
- Liu, Z.; Qiu, P.; Liu, H.; Li, J.; Shao, C.; Yan, T.; Cao, W.; She, Z. Identification of anti-inflammatory polyketides from the coral-derived fungus Penicillium sclerotiorin: In vitro approaches and molecular-modeling. Bioorg. Chem. 2019, 88, 102973. [Google Scholar] [CrossRef]
- Kim, M.J.; Jeong, S.M.; Kang, B.K.; Kim, K.B.; Ahn, D.H. Anti-Inflammatory Effects of Grasshopper Ketone from Sargassum fulvellum Ethanol Extract on Lipopolysaccharide-Induced Inflammatory Responses in RAW 264.7 Cells. J. Microbiol. Biotechnol. 2019, 29, 820–826. [Google Scholar] [CrossRef]
- Abdelfattah, M.S.; Elmallah, M.I.Y.; Ebrahim, H.Y.; Almeer, R.S.; Eltanany, R.M.A.; Abdel Moneim, A.E. Prodigiosins from a marine sponge-associated actinomycete attenuate HCl/ethanol-induced gastric lesion via antioxidant and anti-inflammatory mechanisms. PLoS ONE 2019, 14, e0216737. [Google Scholar] [CrossRef]
- Hwang, J.; Kim, D.; Park, J.S.; Park, H.J.; Shin, J.; Lee, S.K. Photoprotective Activity of Topsentin, A Bis(Indole) Alkaloid from the Marine Sponge Spongosorites genitrix, by Regulation of COX-2 and Mir-4485 Expression in UVB-Irradiated Human Keratinocyte Cells. Mar. Drugs 2020, 18, 87. [Google Scholar] [CrossRef]
- Kim, E.A.; Kim, S.Y.; Kim, J.; Oh, J.Y.; Kim, H.S.; Yoon, W.J.; Kang, D.H.; Heo, S.J. Tuberatolide B isolated from Sargassum macrocarpum inhibited LPS-stimulated inflammatory response via MAPKs and NF-κB signaling pathway in RAW264.7 cells and zebrafish model. J. Funct. Foods 2019, 52, 109–115. [Google Scholar] [CrossRef]
- Yin, Y.; Xu, N.; Shi, Y.; Zhou, B.; Sun, D.; Ma, B.; Xu, Z.; Yang, J.; Li, C. Astaxanthin Protects Dendritic Cells from Lipopolysaccharide-Induced Immune Dysfunction. Mar. Drugs 2021, 19, 346. [Google Scholar] [CrossRef]
- Lin, C.C.; Chang, Y.K.; Lin, S.C.; Su, J.H.; Chao, Y.H.; Tang, K.T. Crassolide Suppresses Dendritic Cell Maturation and Attenuates Experimental Antiphospholipid Syndrome. Molecules 2021, 26, 2492. [Google Scholar] [CrossRef]
- Li, W.; Ye, S.; Zhang, Z.; Tang, J.; Jin, H.; Huang, F.; Yang, Z.; Tang, Y.; Chen, Y.; Ding, G.; et al. Purification and Characterization of a Novel Pentadecapeptide from Protein Hydrolysates of Cyclina sinensis and Its Immunomodulatory Effects on RAW264.7 Cells. Mar. Drugs 2019, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Shim, M.; Son, M.; Jang, J.T.; Son, K.H.; Byun, K. Attenuating Effects of Dieckol on Endothelial Cell Dysfunction via Modulation of Th17/Treg Balance in the Intestine and Aorta of Spontaneously Hypertensive Rats. Antioxidants 2021, 10, 298. [Google Scholar] [CrossRef]
- Park, G.B.; Kim, M.J.; Vasileva, E.A.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; Han, J.; Lee, H.S.; Kim, D.; Jeong, J.Y. Echinochrome A Promotes Ex Vivo Expansion of Peripheral Blood-Derived CD34+ Cells, Potentially through Downregulation of ROS Production and Activation of the Src-Lyn-p110δ Pathway. Mar. Drugs 2019, 17, 526. [Google Scholar] [CrossRef]
- Oh, S.J.; Seo, Y.; Ahn, J.S.; Shin, Y.Y.; Yang, J.W.; Kim, H.K.; Han, J.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; et al. Echinochrome A Reduces Colitis in Mice and Induces In Vitro Generation of Regulatory Immune Cells. Mar. Drugs 2019, 17, 622. [Google Scholar] [CrossRef]
- Park, G.T.; Yoon, J.W.; Yoo, S.B.; Song, Y.C.; Song, P.; Kim, H.K.; Han, J.; Bae, S.J.; Ha, K.T.; Mishchenko, N.P.; et al. Echinochrome A Treatment Alleviates Fibrosis and Inflammation in Bleomycin-Induced Scleroderma. Mar. Drugs 2021, 19, 237. [Google Scholar] [CrossRef]
- Han, E.J.; Kim, H.S.; Sanjeewa, K.K.A.; Herath, K.; Jeon, Y.J.; Jee, Y.; Lee, J.; Kim, T.; Shim, S.Y.; Ahn, G. Eckol from Ecklonia cava Suppresses Immunoglobulin E-mediated Mast Cell Activation and Passive Cutaneous Anaphylaxis in Mice. Nutrients 2020, 12, 1361. [Google Scholar] [CrossRef]
- Tai, H.C.; Lee, T.H.; Tang, C.H.; Chen, L.P.; Chen, W.C.; Lee, M.S.; Chen, P.C.; Lin, C.Y.; Chi, C.W.; Chen, Y.J.; et al. Phomaketide A Inhibits Lymphangiogenesis in Human Lymphatic Endothelial Cells. Mar. Drugs 2019, 17, 215. [Google Scholar] [CrossRef]
- Yang, M.; Li, H.; Zhang, Q.; Wu, Q.H.; Li, G.; Chen, K.X.; Guo, Y.W.; Tang, W.; Li, X.W. Highly diverse cembranoids from the South China Sea soft coral Sinularia scabra as a new class of potential immunosuppressive agents. Bioorg. Med. Chem. 2019, 27, 3469–3476. [Google Scholar] [CrossRef]
- Laborde, R.J.; Ishimura, M.E.; Abreu-Butin, L.; Nogueira, C.V.; Grubaugh, D.; Cruz-Leal, Y.; Luzardo, M.C.; Fernández, A.; Mesa, C.; Pazos, F.; et al. Sticholysins, pore-forming proteins from a marine anemone can induce maturation of dendritic cells through a TLR4 dependent-pathway. Mol. Immunol. 2021, 131, 144–154. [Google Scholar] [CrossRef]
- Manzo, E.; Gallo, C.; Sartorius, R.; Nuzzo, G.; Sardo, A.; De Berardinis, P.; Fontana, A.; Cutignano, A. Immunostimulatory Phosphatidylmonogalactosyldiacylglycerols (PGDG) from the Marine Diatom Thalassiosira weissflogii: Inspiration for a Novel Synthetic Toll-Like Receptor 4 Agonist. Mar. Drugs 2019, 17, 103. [Google Scholar] [CrossRef]
- Wang, H.L.; Li, R.; Li, J.; He, J.; Cao, Z.Y.; Kurtan, T.; Mandi, A.; Zheng, G.L.; Zhang, W. Alternarin A, a Drimane Meroterpenoid, Suppresses Neuronal Excitability from the Coral-Associated Fungi Alternaria sp. ZH-15. Org. Lett. 2020, 22, 2995–2998. [Google Scholar] [CrossRef] [PubMed]
- Andrud, K.; Xing, H.; Gabrielsen, B.; Bloom, L.; Mahnir, V.; Lee, S.; Green, B.T.; Lindstrom, J.; Kem, W. Investigation of the Possible Pharmacologically Active Forms of the Nicotinic Acetylcholine Receptor Agonist Anabaseine. Mar. Drugs 2019, 17, 614. [Google Scholar] [CrossRef]
- Copmans, D.; Kildgaard, S.; Rasmussen, S.A.; Slezak, M.; Dirkx, N.; Partoens, M.; Esguerra, C.V.; Crawford, A.D.; Larsen, T.O.; de Witte, P.A.M. Zebrafish-Based Discovery of Antiseizure Compounds from the North Sea: Isoquinoline Alkaloids TMC-120A and TMC-120B. Mar. Drugs 2019, 17, 607. [Google Scholar] [CrossRef]
- Yang, W.C.; Zhang, Y.Y.; Li, Y.J.; Nie, Y.Y.; Liang, J.Y.; Liu, Y.Y.; Liu, J.S.; Zhang, Y.P.; Song, C.; Qian, Z.J.; et al. Chemical Composition and Anti-Alzheimer’s Disease-Related Activities of a Functional Oil from the Edible Seaweed Hizikia fusiforme. Chem. Biodivers. 2020, 17, e2000055. [Google Scholar] [CrossRef]
- Han, J.H.; Lee, Y.S.; Im, J.H.; Ham, Y.W.; Lee, H.P.; Han, S.B.; Hong, J.T. Astaxanthin Ameliorates Lipopolysaccharide-Induced Neuroinflammation, Oxidative Stress and Memory Dysfunction through Inactivation of the Signal Transducer and Activator of Transcription 3 Pathway. Mar. Drugs 2019, 17, 123. [Google Scholar] [CrossRef]
- Taksima, T.; Chonpathompikunlert, P.; Sroyraya, M.; Hutamekalin, P.; Limpawattana, M.; Klaypradit, W. Effects of Astaxanthin from Shrimp Shell on Oxidative Stress and Behavior in Animal Model of Alzheimer’s Disease. Mar. Drugs 2019, 17, 628. [Google Scholar] [CrossRef]
- Lee, J.; Jun, M. Dual BACE1 and Cholinesterase Inhibitory Effects of Phlorotannins from Ecklonia cava—An In Vitro and in Silico Study. Mar. Drugs 2019, 17, 91. [Google Scholar] [CrossRef]
- Konoki, K.; Baden, D.G.; Scheuer, T.; Catterall, W.A. Molecular Determinants of Brevetoxin Binding to Voltage-Gated Sodium Channels. Toxins 2019, 11, 513. [Google Scholar] [CrossRef]
- Jin, A.H.; Cristofori-Armstrong, B.; Rash, L.D.; Roman-Gonzalez, S.A.; Espinosa, R.A.; Lewis, R.J.; Alewood, P.F.; Vetter, I. Novel conorfamides from Conus austini venom modulate both nicotinic acetylcholine receptors and acid-sensing ion channels. Biochem. Pharmacol. 2019, 164, 342–348. [Google Scholar] [CrossRef]
- Niu, C.; Leavitt, L.S.; Lin, Z.; Paguigan, N.D.; Sun, L.; Zhang, J.; Torres, J.P.; Raghuraman, S.; Chase, K.; Cadeddu, R.; et al. Neuroactive Type-A γ-Aminobutyric Acid Receptor Allosteric Modulator Steroids from the Hypobranchial Gland of Marine Mollusk, Conus geographus. J. Med. Chem. 2021, 64, 7033–7043. [Google Scholar] [CrossRef]
- Guo, M.; Yu, J.; Zhu, X.; Zhangsun, D.; Luo, S. Characterization of an α 4/7-Conotoxin LvIF from Conus lividus That Selectively Blocks α3β2 Nicotinic Acetylcholine Receptor. Mar. Drugs 2021, 19, 398. [Google Scholar] [CrossRef]
- Qiang, Y.; Wu, Y.; Zhao, D.; Zhao, B.; Wang, F.; Ren, S.; Wen, Y.; Gu, J.; Zhang, L.; Liu, K.; et al. Discovery and characterization of the novel conotoxin Lv1d from Conus lividus that presents analgesic activity. Toxicon 2021, 194, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yu, Z.; Yu, S.; Zhu, C.; Dong, M.; Mao, W.; Hu, J.; Prorok, M.; Su, R.; Dai, Q. A Conantokin Peptide Con-T[M8Q] Inhibits Morphine Dependence with High Potency and Low Side Effects. Mar. Drugs 2021, 19, 44. [Google Scholar] [CrossRef]
- Wu, J.; Xi, Y.; Li, G.; Zheng, Y.; Wang, Z.; Wang, J.; Fang, C.; Sun, Z.; Hu, L.; Jiang, W.; et al. Hydroazulene Diterpenes from a Dictyota Brown Alga and Their Antioxidant and Neuroprotective Effects against Cerebral Ischemia-Reperfusion Injury. J. Nat. Prod. 2021, 84, 1306–1315. [Google Scholar] [CrossRef]
- Kim, R.; Hur, D.; Kim, H.K.; Han, J.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; Chang, W. Echinochrome A Attenuates Cerebral Ischemic Injury through Regulation of Cell Survival after Middle Cerebral Artery Occlusion in Rat. Mar. Drugs 2019, 17, 501. [Google Scholar] [CrossRef]
- Paudel, P.; Seong, S.H.; Wu, S.; Park, S.; Jung, H.A.; Choi, J.S. Eckol as a Potential Therapeutic against Neurodegenerative Diseases Targeting Dopamine D3/D4 Receptors. Mar. Drugs 2019, 17, 108. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Alves, C.; Pinteus, S.; Susano, P.; Simoes, M.; Guedes, M.; Martins, A.; Rehfeldt, S.; Gaspar, H.; Goettert, M.; et al. Disclosing the potential of eleganolone for Parkinson’s disease therapeutics: Neuroprotective and anti-inflammatory activities. Pharmacol. Res. 2021, 168, 105589. [Google Scholar] [CrossRef] [PubMed]
- Chalorak, P.; Sanguanphun, T.; Limboonreung, T.; Meemon, K. Neurorescue Effects of Frondoside A and Ginsenoside Rg3 in C. elegans Model of Parkinson’s Disease. Molecules 2021, 26, 4843. [Google Scholar] [CrossRef] [PubMed]
- Gan, S.Y.; Wong, L.Z.; Wong, J.W.; Tan, E.L. Fucosterol exerts protection against amyloid β-induced neurotoxicity, reduces intracellular levels of amyloid beta and enhances the mRNA expression of neuroglobin in amyloid β-induced SH-SY5Y cells. Int. J. Biol. Macromol. 2019, 121, 207–213. [Google Scholar] [CrossRef]
- Hannan, M.A.; Dash, R.; Sohag, A.A.M.; Moon, I.S. Deciphering Molecular Mechanism of the Neuropharmacological Action of Fucosterol through Integrated System Pharmacology and In Silico Analysis. Mar. Drugs 2019, 17, 639. [Google Scholar] [CrossRef]
- Chen, S.J.; Lee, C.J.; Lin, T.B.; Peng, H.Y.; Liu, H.J.; Chen, Y.S.; Tseng, K.W. Protective Effects of Fucoxanthin on Ultraviolet B-Induced Corneal Denervation and Inflammatory Pain in a Rat Model. Mar. Drugs 2019, 17, 152. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Han, H.; Liu, J.; Tang, M.; Wu, X.; Cao, X.; Zhao, T.; Lu, Y.; Niu, T.; Chen, J.; et al. Fucoxanthin Prevents 6-OHDA-Induced Neurotoxicity by Targeting Keap1. Oxid. Med. Cell. Longev. 2021, 2021, 6688708. [Google Scholar] [CrossRef] [PubMed]
- Kalina, R.S.; Koshelev, S.G.; Zelepuga, E.A.; Kim, N.Y.; Kozlov, S.A.; Kozlovskaya, E.P.; Monastyrnaya, M.M.; Gladkikh, I.N. APETx-Like Peptides from the Sea Anemone Heteractis crispa, Diverse in Their Effect on ASIC1a and ASIC3 Ion Channels. Toxins 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- Tangrodchanapong, T.; Sornkaew, N.; Yurasakpong, L.; Niamnont, N.; Nantasenamat, C.; Sobhon, P.; Meemon, K. Beneficial Effects of Cyclic Ether 2-Butoxytetrahydrofuran from Sea Cucumber Holothuria scabra against Aβ Aggregate Toxicity in Transgenic Caenorhabditis elegans and Potential Chemical Interaction. Molecules 2021, 26, 2195. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.T.; Zhang, H.H.; Tang, Y.H.; Zhang, F.Z.; Han, B.N. Two New Neo-debromoaplysiatoxins—A Pair of Stereoisomers Exhibiting Potent Kv1.5 Ion Channel Inhibition Activities. Mar. Drugs 2019, 17, 652. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Wang, G.; Li, D.; Li, H.; Liu, J.; Yang, X.; Yang, W. Okadaic Acid Exposure Induced Neural Tube Defects in Chicken (Gallus gallus) Embryos. Mar. Drugs 2021, 19, 322. [Google Scholar] [CrossRef] [PubMed]
- Benoit, E.; Couesnon, A.; Lindovsky, J.; Iorga, B.I.; Araoz, R.; Servent, D.; Zakarian, A.; Molgo, J. Synthetic Pinnatoxins A and G Reversibly Block Mouse Skeletal Neuromuscular Transmission In Vivo and In Vitro. Mar. Drugs 2019, 17, 306. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.H.; Paudel, P.; Choi, J.W.; Ahn, D.H.; Nam, T.J.; Jung, H.A.; Choi, J.S. Probing Multi-Target Action of Phlorotannins as New Monoamine Oxidase Inhibitors and Dopaminergic Receptor Modulators with the Potential for Treatment of Neuronal Disorders. Mar. Drugs 2019, 17, 377. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.P.; Kang, M.G.; Lee, J.Y.; Oh, J.M.; Baek, S.C.; Leem, H.H.; Park, D.; Cho, M.L.; Kim, H. Potent inhibition of acetylcholinesterase by sargachromanol I from Sargassum siliquastrum and by selected natural compounds. Bioorg. Chem. 2019, 89, 103043. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.C.; Tan, H.; Zhang, Y.; Xu, J.; Liu, W.L.; Li, Z.Y.; Li, W.P. Santacruzamate A Ameliorates AD-Like Pathology by Enhancing ER Stress Tolerance through Regulating the Functions of KDELR and Mia40-ALR in vivo and in vitro. Front. Cell. Neurosci. 2019, 13, 61. [Google Scholar] [CrossRef]
- Jiang, C.S.; Ru, T.; Yao, L.G.; Miao, Z.H.; Guo, Y.W. Four new cembranoids from the Chinese soft coral Sinularia sp. and their anti-Aβ aggregation activities. Fitoterapia 2019, 136, 104176. [Google Scholar] [CrossRef]
- Paudel, P.; Park, S.E.; Seong, S.H.; Jung, H.A.; Choi, J.S. Bromophenols from Symphyocladia latiuscula Target Human Monoamine Oxidase and Dopaminergic Receptors for the Management of Neurodegenerative Diseases. J. Agric. Food Chem. 2020, 68, 2426–2436. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, W.; Deng, Z.; Zhang, Q.; Wang, J. Glucuronomannan GM2 from Saccharina japonica Enhanced Mitochondrial Function and Autophagy in a Parkinson’s Model. Mar. Drugs 2021, 19, 58. [Google Scholar] [CrossRef]
- Paudel, P.; Seong, S.H.; Zhou, Y.; Park, H.J.; Jung, H.A.; Choi, J.S. Anti-Alzheimer’s Disease Activity of Bromophenols from a Red Alga, Symphyocladia latiuscula (Harvey) Yamada. ACS Omega 2019, 4, 12259–12270. [Google Scholar] [CrossRef]
- Feng, C.W.; Chen, N.F.; Wen, Z.H.; Yang, W.Y.; Kuo, H.M.; Sung, P.J.; Su, J.H.; Cheng, S.Y.; Chen, W.F. In Vitro and In Vivo Neuroprotective Effects of Stellettin B through Anti-Apoptosis and the Nrf2/HO-1 Pathway. Mar. Drugs 2019, 17, 315. [Google Scholar] [CrossRef]
- Sheng, L.; Lu, B.; Chen, H.; Du, Y.; Chen, C.; Cai, W.; Yang, Y.; Tian, X.; Huang, Z.; Chi, W.; et al. Marine-Steroid Derivative 5α-Androst-3β, 5α, 6β-triol Protects Retinal Ganglion Cells from Ischemia–Reperfusion Injury by Activating Nrf2 Pathway. Mar. Drugs 2019, 17, 267. [Google Scholar] [CrossRef]
- Nakamura, Y.; Matsumoto, H.; Wu, C.H.; Fukaya, D.; Uni, R.; Hirakawa, Y.; Katagiri, M.; Yamada, S.; Ko, T.; Nomura, S.; et al. Alpha 7 nicotinic acetylcholine receptors signaling boosts cell-cell interactions in macrophages effecting anti-inflammatory and organ protection. Commun. Biol. 2023, 6, 666. [Google Scholar] [CrossRef]
- Liang, X.; Luo, D.; Yan, J.L.; Rezaei, M.A.; Salvador-Reyes, L.A.; Gunasekera, S.P.; Li, C.; Ye, T.; Paul, V.J.; Luesch, H. Discovery of Amantamide, a Selective CXCR7 Agonist from Marine Cyanobacteria. Org. Lett. 2019, 21, 1622–1626. [Google Scholar] [CrossRef]
- Chakraborty, K.; Krishnan, S.; Joy, M. Macrocyclic lactones from seafood Amphioctopus neglectus: Newly described natural leads to attenuate angiotensin-II induced cardiac hypertrophy. Biomed. Pharmacother. 2019, 110, 155–167. [Google Scholar] [CrossRef]
- Shi, H.; Hu, X.; Zheng, H.; Li, C.; Sun, L.; Guo, Z.; Huang, W.; Yu, R.; Song, L.; Zhu, J. Two novel antioxidant peptides derived from Arca subcrenata against oxidative stress and extend lifespan in Caenorhabditis elegans. J. Funct. Foods 2021, 81, 104462. [Google Scholar] [CrossRef]
- Park, S.C.; Chung, B.; Lee, J.; Cho, E.; Hwang, J.Y.; Oh, D.C.; Shin, J.; Oh, K.B. Sortase A-Inhibitory Metabolites from a Marine-Derived Fungus Aspergillus sp. Mar. Drugs 2020, 18, 359. [Google Scholar] [CrossRef] [PubMed]
- Ohshiro, T.; Kobayashi, K.; Suzuki, A.; Yamazaki, H.; Uchida, R.; Namikoshi, M.; Tomoda, H. Inhibition of neutral lipid synthesis by avarols from a marine sponge. Bioorg. Med. Chem. Lett. 2019, 29, 2283–2285. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Oh, S.; Jang, J.T.; Son, K.H.; Byun, K. Pyrogallol-Phloroglucinol-6 6-Bieckol on Attenuates High-Fat Diet-Induced Hypertension by Modulating Endothelial-to-Mesenchymal Transition in the Aorta of Mice. Oxid. Med. Cell. Longev. 2021, 2021, 8869085. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.S.; Fernando, P.; Kang, K.A.; Piao, M.J.; Zhen, A.X.; Kang, H.K.; Koh, Y.S.; Hyun, J.W. Marine Compound 3-bromo-4,5-dihydroxybenzaldehyde Protects Skin Cells against Oxidative Damage via the Nrf2/HO-1 Pathway. Mar. Drugs 2019, 17, 234. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xu, Z.; Fan, F.; Shi, P.; Tu, M.; Wang, Z.; Du, M. Identification and mechanism evaluation of a novel osteogenesis promoting peptide from Tubulin Alpha-1C chain in Crassostrea gigas. Food Chem. 2019, 272, 751–757. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Cheng, S.; Fan, F.; Tu, M.; Xu, Z.; Du, M. Identification and molecular mechanism of antithrombotic peptides from oyster proteins released in simulated gastro-intestinal digestion. Food Funct. 2019, 10, 5426–5435. [Google Scholar] [CrossRef] [PubMed]
- Utkina, N.K.; Likhatskaya, G.N.; Balabanova, L.A.; Bakunina, I.Y. Sponge-derived polybrominated diphenyl ethers and dibenzo-p-dioxins, irreversible inhibitors of the bacterial alpha-d-galactosidase. Environ. Sci. Process. Impacts 2019, 21, 1754–1763. [Google Scholar] [CrossRef] [PubMed]
- Pyeon, D.B.; Lee, S.E.; Yoon, J.W.; Park, H.J.; Park, C.O.; Kim, S.H.; Oh, S.H.; Lee, D.G.; Kim, E.Y.; Park, S.P. The antioxidant dieckol reduces damage of oxidative stress-exposed porcine oocytes and enhances subsequent parthenotes embryo development. Mol. Reprod. Dev. 2021, 88, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Je, J.G.; Yang, H.W.; Jeon, Y.J.; Lee, S. Dieckol, an Algae-Derived Phenolic Compound, Suppresses UVB-Induced Skin Damage in Human Dermal Fibroblasts and Its Underlying Mechanisms. Antioxidants 2021, 10, 352. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, L.; Im, S.; Hwang, O.; Kim, H.S.; Kang, M.C.; Lee, S.H. Anti-Obesity Effect of Diphlorethohydroxycarmalol Isolated from Brown Alga Ishige okamurae in High-Fat Diet-Induced Obese Mice. Mar. Drugs 2019, 17, 637. [Google Scholar] [CrossRef]
- Kang, M.C.; Ding, Y.; Kim, H.S.; Jeon, Y.J.; Lee, S.H. Inhibition of Adipogenesis by Diphlorethohydroxycarmalol (DPHC) through AMPK Activation in Adipocytes. Mar. Drugs 2019, 17, 44. [Google Scholar] [CrossRef]
- Lu, Y.A.; Jiang, Y.; Yang, H.W.; Hwang, J.; Jeon, Y.J.; Ryu, B. Diphlorethohydroxycarmalol Isolated from Ishige okamurae Exerts Vasodilatory Effects via Calcium Signaling and PI3K/Akt/eNOS Pathway. Int. J. Mol. Sci. 2021, 22, 1610. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, B.; Wagle, A.; Seong, S.H.; Paudel, P.; Kim, H.R.; Jung, H.A.; Choi, J.S. Phlorotannins with Potential Anti-tyrosinase and Antioxidant Activity Isolated from the Marine Seaweed Ecklonia stolonifera. Antioxidants 2019, 8, 240. [Google Scholar] [CrossRef]
- Zhen, A.X.; Hyun, Y.J.; Piao, M.J.; Fernando, P.; Kang, K.A.; Ahn, M.J.; Yi, J.M.; Kang, H.K.; Koh, Y.S.; Lee, N.H.; et al. Eckol Inhibits Particulate Matter 2.5-Induced Skin Keratinocyte Damage via MAPK Signaling Pathway. Mar. Drugs 2019, 17, 444. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Xu, M.; Yang, A.; Zhao, Y.; Liu, D.; Huang, J.; Proksch, P.; Lin, W. Reducing Effect of Farnesylquinone on Lipid Mass in C. elegans by Modulating Lipid Metabolism. Mar. Drugs 2019, 17, 336. [Google Scholar] [CrossRef] [PubMed]
- Ohno, Y.; Kondo, S.; Tajima, K.; Shibata, T.; Itoh, T. Effect of phlorotannins isolated from Eisenia bicyclis on melanogenesis in mouse B16 melanoma cells. Nat. Prod. Commun. 2021, 16, 1934578X211019264. [Google Scholar] [CrossRef]
- Raji, V.; Loganathan, C.; Sadhasivam, G.; Kandasamy, S.; Poomani, K.; Thayumanavan, P. Purification of fucoxanthin from Sargassum wightii Greville and understanding the inhibition of angiotensin 1-converting enzyme: An in vitro and in silico studies. Int. J. Biol. Macromol. 2020, 148, 696–703. [Google Scholar] [CrossRef]
- Yang, G.; Jin, L.; Zheng, D.; Tang, X.; Yang, J.; Fan, L.; Xie, X. Fucoxanthin Alleviates Oxidative Stress through Akt/Sirt1/FoxO3α Signaling to Inhibit HG-Induced Renal Fibrosis in GMCs. Mar. Drugs 2019, 17, 702. [Google Scholar] [CrossRef]
- Kim, D.C.; Minh Ha, T.; Sohn, J.H.; Yim, J.H.; Oh, H. Protein tyrosine phosphatase 1B inhibitors from a marine-derived fungal strain Aspergillus sp. SF-5929. Nat. Prod. Res. 2020, 34, 675–682. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Z.X.; Zhao, W.C.; Huang, H.B.; Wang, J.Q.; Zhang, H.; Lu, J.Y.; Wang, R.N.; Li, W.; Cheng, Z.; et al. Identification and characterization of isocitrate dehydrogenase 1 (IDH1) as a functional target of marine natural product grincamycin B. Acta Pharmacol. Sin. 2021, 42, 801–813. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, L.; Yu, G.; Qi, X.; Zhang, A.; Lu, Z.; Li, D.; Li, J. Identification of a novel non-ATP-competitive protein kinase inhibitor of PGK1 from marine nature products. Biochem. Pharmacol. 2021, 183, 114343. [Google Scholar] [CrossRef]
- Oh, Y.; Ahn, C.B.; Je, J.Y. Cytoprotective Role of Edible Seahorse (Hippocampus abdominalis)-Derived Peptides in H2O2-Induced Oxidative Stress in Human Umbilical Vein Endothelial Cells. Mar. Drugs 2021, 19, 86. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.G.; Kim, H.S.; Je, J.G.; Hwang, J.; Sanjeewa, K.K.A.; Lee, D.S.; Song, K.M.; Choi, Y.S.; Kang, M.C.; Jeon, Y.J. Lipid Inhibitory Effect of (−)-loliolide Isolated from Sargassum horneri in 3T3-L1 Adipocytes: Inhibitory Mechanism of Adipose-Specific Proteins. Mar. Drugs 2021, 19, 96. [Google Scholar] [CrossRef]
- Bakunina, I.; Likhatskaya, G.; Slepchenko, L.; Balabanova, L.; Tekutyeva, L.; Son, O.; Shubina, L.; Makarieva, T. Effect of Pentacyclic Guanidine Alkaloids from the Sponge Monanchora pulchra on Activity of α-Glycosidases from Marine Bacteria. Mar. Drugs 2018, 17, 22. [Google Scholar] [CrossRef]
- Nagabhishek, S.N.K.; Kumar, A.M.; Sambhavi, B.; Balakrishnan, A.; Katakia, Y.T.; Chatterjee, S.; Nagasundaram, N. A marine sponge associated fungal metabolite monacolin X suppresses angiogenesis by down regulating VEGFR2 signaling. RSC Adv. 2019, 9, 26646–26667. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, E.; Iannotti, F.A.; Falkenberg, L.G.; Martella, A.; Gentile, A.; De Maio, F.; Ciavatta, M.L.; Gavagnin, M.; Waxman, J.S.; Di Marzo, V.; et al. In Silico Identification and Experimental Validation of (−)-Muqubilin A, a Marine Norterpene Peroxide, as PPARα/γ-RXRα Agonist and RARα Positive Allosteric Modulator. Mar. Drugs 2019, 17, 110. [Google Scholar] [CrossRef] [PubMed]
- Hayashi-Takanaka, Y.; Kina, Y.; Nakamura, F.; Yamazaki, S.; Harata, M.; Soest, R.; Kimura, H.; Nakao, Y. Effect of mycalolides isolated from a marine sponge Mycale aff. nullarosette on actin in living cells. Sci. Rep. 2019, 9, 7540. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, H.; Fan, F.; Shi, P.; Tu, M.; Cheng, S.; Wang, Z.; Du, M. Bone formation activity of an osteogenic dodecapeptide from blue mussels (Mytilus edulis). Food Funct. 2019, 10, 5616–5625. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gong, F.; Chen, M.F.; Li, C.; Hong, P.; Sun, S.; Zhou, C.; Qian, Z.J. In Vitro Vascular-Protective Effects of a Tilapia By-Product Oligopeptide on Angiotensin II-Induced Hypertensive Endothelial Injury in HUVEC by Nrf2/NF-κB Pathways. Mar. Drugs 2019, 17, 431. [Google Scholar] [CrossRef]
- Kong, F.D.; Fan, P.; Zhou, L.M.; Ma, Q.Y.; Xie, Q.Y.; Zheng, H.Z.; Zheng, Z.H.; Zhang, R.S.; Yuan, J.Z.; Dai, H.F.; et al. Penerpenes A–D, Four Indole Terpenoids with Potent Protein Tyrosine Phosphatase Inhibitory Activity from the Marine-Derived Fungus Penicillium sp. KFD28. Org. Lett. 2019, 21, 4864–4867. [Google Scholar] [CrossRef]
- Dai, J.; Chen, A.; Zhu, M.; Qi, X.; Tang, W.; Liu, M.; Li, D.; Gu, Q.; Li, J. Penicisulfuranol A, a novel C-terminal inhibitor disrupting molecular chaperone function of Hsp90 independent of ATP binding domain. Biochem. Pharmacol. 2019, 163, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liang, Z.; Li, K.; Yang, B.; Liu, Y.; Fang, W.; Tang, L.; Zhou, X. Ene-yne Hydroquinones from a Marine-derived Strain of the Fungus Pestalotiopsis neglecta with Effects on Liver X Receptor Alpha. J. Nat. Prod. 2020, 83, 1258–1264. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.Z.; Liu, J.T.; Hu, Q.; He, R.J.; Guan, X.Q.; Ge, G.B.; Han, H.; Yang, F.; Lin, H.W. Pancreatic Lipase Inhibitory Cyclohexapeptides from the Marine Sponge-Derived Fungus Aspergillus sp. 151304. J. Nat. Prod. 2020, 83, 2287–2293. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liao, H.; Liu, L.Y.; Sun, F.; Chen, H.F.; Jiao, W.H.; Zhu, H.R.; Yang, F.; Huang, G.; Zeng, D.Q.; et al. Phakefustatins A-C: Kynurenine-Bearing Cycloheptapeptides as RXRα Modulators from the Marine Sponge Phakellia fusca. Org. Lett. 2020, 22, 6703–6708. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, S.; Park, S.; Park, J.S.; Kim, Y.H.; Yang, S.Y. Slow-Binding Inhibition of Tyrosinase by Ecklonia cava Phlorotannins. Mar. Drugs 2019, 17, 359. [Google Scholar] [CrossRef] [PubMed]
- Heo, S.Y.; Jeong, M.S.; Lee, H.S.; Kim, Y.J.; Park, S.H.; Jung, W.K. Phlorofucofuroeckol A from Ecklonia cava ameliorates TGF-β1-induced fibrotic response of human tracheal fibroblasts via the downregulation of MAPKs and SMAD 2/3 pathways inactivated TGF-β receptor. Biochem. Biophys. Res. Commun. 2020, 522, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Ahn, B.N.; Karadeniz, F.; Kim, J.A.; Lee, J.I.; Seo, Y.; Kong, C.S. Phlorofucofuroeckol A from Edible Brown Alga Ecklonia Cava Enhances Osteoblastogenesis in Bone Marrow-Derived Human Mesenchymal Stem Cells. Mar. Drugs 2019, 17, 543. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.C.; Zhang, Y.H.; Gao, W.B.; Pan, L.; Zhu, H.J.; Cao, F. Absolute Configurations and Chitinase Inhibitions of Quinazoline-Containing Diketopiperazines from the Marine-Derived Fungus Penicillium polonicum. Mar. Drugs 2020, 18, 479. [Google Scholar] [CrossRef]
- Ko, S.C.; Ding, Y.; Kim, J.; Ye, B.R.; Kim, E.A.; Jung, W.K.; Heo, S.J.; Lee, S.H. Bromophenol (5-bromo-3,4-dihydroxybenzaldehyde) isolated from red alga Polysiphonia morrowii inhibits adipogenesis by regulating expression of adipogenic transcription factors and AMP-activated protein kinase activation in 3T3-L1 adipocytes. Phytother. Res. 2019, 33, 737–744. [Google Scholar] [CrossRef]
- Park, J.S.; Quang, T.H.; Thi Thanh Ngan, N.; Sohn, J.H.; Oh, H. New preaustinoids from a marine-derived fungal strain Penicillium sp. SF-5497 and their inhibitory effects against PTP1B activity. J. Antibiot. 2019, 72, 629–633. [Google Scholar] [CrossRef]
- Lee, M.K.; Choi, J.W.; Choi, Y.H.; Nam, T.J. Protective Effect of Pyropia yezoensis Peptide on Dexamethasone-Induced Myotube Atrophy in C2C12 Myotubes. Mar. Drugs 2019, 17, 284. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.C.; Lai, Y.C.; Kuo, T.J.; Su, J.H.; Sung, P.J.; Feng, C.W.; Lin, Y.Y.; Chen, P.C.; Tai, M.H.; Cheng, S.Y.; et al. Rhodoptilometrin, a Crinoid-Derived Anthraquinone, Induces Cell Regeneration by Promoting Wound Healing and Oxidative Phosphorylation in Human Gingival Fibroblast Cells. Mar. Drugs 2019, 17, 138. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Lee, B.; Lim, S.-J.; Choi, J.S.; Kim, H.-R. Sargahydroquinoic acid, a major compound in Sargassum serratifolium (C. Agardh) C. Agardh, widely activates lipid catabolic pathways, contributing to the formation of beige-like adipocytes. J. Funct. Foods 2019, 58, 355–366. [Google Scholar] [CrossRef]
- Halkias, C.; Darby, W.G.; Feltis, B.N.; McIntyre, P.; Macrides, T.A.; Wright, P.F.A. Marine Bile Natural Products as Agonists of the TGR5 Receptor. J. Nat. Prod. 2021, 84, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-Y.; Chung, B.; Kwon, O.-S.; Park, S.C.; Cho, E.; Oh, D.-C.; Shin, J.; Oh, K.-B. Inhibitory effects of epipolythiodioxopiperazine fungal metabolites on isocitrate lyase in the glyoxylate cycle of Candida albicans. Mar. Drugs 2021, 19, 295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, R.; Ba, S.; Lu, Z.; Pitsinos, E.N.; Li, T.; Nicolaou, K.C. DNA Binding and Cleavage Modes of Shishijimicin A. J. Am. Chem. Soc. 2019, 141, 7842–7852. [Google Scholar] [CrossRef]
- Zheng, J.; Manabe, Y.; Sugawara, T. Siphonaxanthin, a carotenoid from green algae Codium cylindricum, protects Ob/Ob mice fed on a high-fat diet against lipotoxicity by ameliorating somatic stresses and restoring anti-oxidative capacity. Nutr. Res. 2020, 77, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Paudel, P.; Wagle, A.; Seong, S.H.; Park, H.J.; Jung, H.A.; Choi, J.S. A New Tyrosinase Inhibitor from the Red Alga Symphyocladia latiuscula (Harvey) Yamada (Rhodomelaceae). Mar. Drugs 2019, 17, 295. [Google Scholar] [CrossRef]
- Qiao, X.; Gan, M.; Wang, C.; Liu, B.; Shang, Y.; Li, Y.; Chen, S. Tetracenomycin X Exerts Antitumour Activity in Lung Cancer Cells through the Downregulation of Cyclin D1. Mar. Drugs 2019, 17, 63. [Google Scholar] [CrossRef]
- Keller, L.; Canuto, K.M.; Liu, C.; Suzuki, B.M.; Almaliti, J.; Sikandar, A.; Naman, C.B.; Glukhov, E.; Luo, D.; Duggan, B.M.; et al. Tutuilamides A–C: Vinyl-Chloride-Containing Cyclodepsipeptides from Marine Cyanobacteria with Potent Elastase Inhibitory Properties. ACS Chem. Biol. 2020, 15, 751–757. [Google Scholar] [CrossRef]
- Feng, X.; Liao, D.; Sun, L.; Wu, S.; Lan, P.; Wang, Z.; Li, C.; Zhou, Q.; Lu, Y.; Lan, X. Affinity Purification of Angiotensin Converting Enzyme Inhibitory Peptides from Wakame (Undaria pinnatifida) Using Immobilized ACE on Magnetic Metal Organic Frameworks. Mar. Drugs 2021, 19, 177. [Google Scholar] [CrossRef] [PubMed]
- El-Baz, F.K.; Hussein, R.A.; Saleh, D.O.; Abdel Jaleel, G.A.R. Zeaxanthin Isolated from Dunaliella salina Microalgae Ameliorates Age Associated Cardiac Dysfunction in Rats through Stimulation of Retinoid Receptors. Mar. Drugs 2019, 17, 290. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2021, 38, 362–413. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2022, 39, 1122–1171. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2023, 40, 275–325. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Luo, D.; Luesch, H. Advances in exploring the therapeutic potential of marine natural products. Pharmacol. Res. 2019, 147, 104373. [Google Scholar] [CrossRef]
- Nunez-Pons, L.; Shilling, A.; Verde, C.; Baker, B.J.; Giordano, D. Marine Terpenoids from Polar Latitudes and Their Potential Applications in Biotechnology. Mar. Drugs 2020, 18, 401. [Google Scholar] [CrossRef]
- Mateos, R.; Perez-Correa, J.R.; Dominguez, H. Bioactive Properties of Marine Phenolics. Mar. Drugs 2020, 18, 501. [Google Scholar] [CrossRef] [PubMed]
- Martins, B.T.; Correia da Silva, M.; Pinto, M.; Cidade, H.; Kijjoa, A. Marine natural flavonoids: Chemistry and biological activities. Nat. Prod. Res. 2019, 33, 3260–3272. [Google Scholar] [CrossRef]
- Braddock, A.A.; Theodorakis, E.A. Marine Spirotetronates: Biosynthetic Edifices That Inspire Drug Discovery. Mar. Drugs 2019, 17, 232. [Google Scholar] [CrossRef]
- Vil, V.A.; Gloriozova, T.A.; Terent’ev, A.O.; Savidov, N.; Dembitsky, V.M. Hydroperoxides derived from marine sources: Origin and biological activities. Appl. Microbiol. Biotechnol. 2019, 103, 1627–1642. [Google Scholar] [CrossRef] [PubMed]
- Althagbi, H.I.; Alarif, W.M.; Al-Footy, K.O.; Abdel-Lateff, A. Marine-Derived Macrocyclic Alkaloids (MDMAs): Chemical and Biological Diversity. Mar. Drugs 2020, 18, 368. [Google Scholar] [CrossRef]
- Dahiya, R.; Dahiya, S.; Fuloria, N.K.; Kumar, S.; Mourya, R.; Chennupati, S.V.; Jankie, S.; Gautam, H.; Singh, S.; Karan, S.K.; et al. Natural Bioactive Thiazole-Based Peptides from Marine Resources: Structural and Pharmacological Aspects. Mar. Drugs 2020, 18, 329. [Google Scholar] [CrossRef]
- El-Demerdash, A.; Kumla, D.; Kijjoa, A. Chemical Diversity and Biological Activities of Meroterpenoids from Marine Derived-Fungi: A Comprehensive Update. Mar. Drugs 2020, 18, 317. [Google Scholar] [CrossRef]
- Li, G.; Dickschat, J.S.; Guo, Y.W. Diving into the world of marine 2,11-cyclized cembranoids: A summary of new compounds and their biological activities. Nat. Prod. Rep. 2020, 37, 1367–1383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zou, J.; Yan, X.; Chen, J.; Cao, X.; Wu, J.; Liu, Y.; Wang, T. Marine-Derived Macrolides 1990–2020: An Overview of Chemical and Biological Diversity. Mar. Drugs 2021, 19, 180. [Google Scholar] [CrossRef] [PubMed]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural Products from Cyanobacteria: Focus on Beneficial Activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef]
- Fidor, A.; Konkel, R.; Mazur-Marzec, H. Bioactive Peptides Produced by Cyanobacteria of the Genus Nostoc: A Review. Mar. Drugs 2019, 17, 561. [Google Scholar] [CrossRef]
- Huang, I.S.; Zimba, P.V. Cyanobacterial bioactive metabolites—A review of their chemistry and biology. Harmful Algae 2019, 86, 139–209. [Google Scholar] [CrossRef]
- Li, Y.; Naman, C.B.; Alexander, K.L.; Guan, H.; Gerwick, W.H. The Chemistry, Biochemistry and Pharmacology of Marine Natural Products from Leptolyngbya, a Chemically Endowed Genus of Cyanobacteria. Mar. Drugs 2020, 18, 508. [Google Scholar] [CrossRef]
- Tan, L.T.; Phyo, M.Y. Marine Cyanobacteria: A Source of Lead Compounds and their Clinically-Relevant Molecular Targets. Molecules 2020, 25, 2197. [Google Scholar] [CrossRef] [PubMed]
- Qamar, H.; Hussain, K.; Soni, A.; Khan, A.; Hussain, T.; Chénais, B. Cyanobacteria as Natural Therapeutics and Pharmaceutical Potential: Role in Antitumor Activity and as Nanovectors. Molecules 2021, 26, 247. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Ferrante, M.I.; Rogato, A. Marine Natural Products from Microalgae: An Omics Overview. Mar. Drugs 2019, 17, 269. [Google Scholar] [CrossRef] [PubMed]
- Sathasivam, R.; Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Microalgae metabolites: A rich source for food and medicine. Saudi J. Biol. Sci. 2019, 26, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N. Current Trends on Seaweeds: Looking at Chemical Composition, Phytopharmacology, and Cosmetic Applications. Molecules 2019, 24, 4182. [Google Scholar] [CrossRef] [PubMed]
- Cikos, A.M.; Jurin, M.; Coz-Rakovac, R.; Jokic, S.; Jerkovic, I. Update on Monoterpenes from Red Macroalgae: Isolation, Analysis, and Bioactivity. Mar. Drugs 2019, 17, 537. [Google Scholar] [CrossRef] [PubMed]
- Freile-Pelegrín, Y.; Tasdemir, D. Seaweeds to the rescue of forgotten diseases: A review. Bot. Mar. 2019, 62, 211–226. [Google Scholar] [CrossRef]
- Hannan, M.A.; Sohag, A.A.M.; Dash, R.; Haque, M.N.; Mohibbullah, M.; Oktaviani, D.F.; Hossain, M.T.; Choi, H.J.; Moon, I.S. Phytosterols of marine algae: Insights into the potential health benefits and molecular pharmacology. Phytomedicine 2020, 69, 153201. [Google Scholar] [CrossRef]
- Rosa, G.P.; Tavares, W.R.; Sousa, P.M.C.; Pages, A.K.; Seca, A.M.L.; Pinto, D. Seaweed Secondary Metabolites with Beneficial Health Effects: An Overview of Successes in In Vivo Studies and Clinical Trials. Mar. Drugs 2019, 18, 8. [Google Scholar] [CrossRef]
- Rushdi, I.; Abdel-Rahman, I.; Saber, H.; Attia, E.; Abdelraheem, W.; Madkour, H.; Hassan, H.; Elmaidomv, A.; Abdelmohsen, U. Pharmacological and natural products diversity of the brown algae genus Sargassum. RSC Adv. 2020, 10, 24951–24972. [Google Scholar] [CrossRef]
- Purcell-Meyerink, D.; Packer, M.A.; Wheeler, T.T.; Hayes, M. Aquaculture Production of the Brown Seaweeds Laminaria digitata and Macrocystis pyrifera: Applications in Food and Pharmaceuticals. Molecules 2021, 26, 1306. [Google Scholar] [CrossRef] [PubMed]
- Shannon, E.; Conlon, M.; Hayes, M. Seaweed Components as Potential Modulators of the Gut Microbiota. Mar. Drugs 2021, 19, 358. [Google Scholar] [CrossRef] [PubMed]
- Thangaraj, S.; Bragadeeswaran, S.; Gokula, V. Bioactive compounds of sea anemones: A review. Int. J. Pept. Res. Ther. 2019, 25, 1405–1416. [Google Scholar] [CrossRef]
- Tinta, T.; Kogovsek, T.; Klun, K.; Malej, A.; Herndl, G.J.; Turk, V. Jellyfish-Associated Microbiome in the Marine Environment: Exploring Its Biotechnological Potential. Mar. Drugs 2019, 17, 94. [Google Scholar] [CrossRef] [PubMed]
- Ciavatta, M.L.; Lefranc, F.; Vieira, L.M.; Kiss, R.; Carbone, M.; van Otterlo, W.A.L.; Lopanik, N.B.; Waeschenbach, A. The Phylum Bryozoa: From Biology to Biomedical Potential. Mar. Drugs 2020, 18, 200. [Google Scholar] [CrossRef] [PubMed]
- Guillen, P.O.; Jaramillo, K.B.; Genta-Jouve, G.; Thomas, O.P. Marine natural products from zoantharians: Bioactivity, biosynthesis, systematics, and ecological roles. Nat. Prod. Rep. 2020, 37, 515–540. [Google Scholar] [CrossRef]
- Youssef, D.T.A.; Almagthali, H.; Shaala, L.A.; Schmidt, E.W. Secondary Metabolites of the Genus Didemnum: A Comprehensive Review of Chemical Diversity and Pharmacological Properties. Mar. Drugs 2020, 18, 307. [Google Scholar] [CrossRef] [PubMed]
- Barone, G.; Varrella, S.; Tangherlini, M.; Rastelli, E.; Dell’Anno, A.; Danovaro, R.; Corinaldesi, C. Marine fungi: Biotechnological perspectives from deep-hypersaline anoxic basins. Diversity 2019, 11, 113. [Google Scholar] [CrossRef]
- Youssef, F.S.; Ashour, M.L.; Singab, A.N.B.; Wink, M. A Comprehensive Review of Bioactive Peptides from Marine Fungi and Their Biological Significance. Mar. Drugs 2019, 17, 559. [Google Scholar] [CrossRef]
- Youssef, F.S.; Simal-Gandara, J. Comprehensive Overview on the Chemistry and Biological Activities of Selected Alkaloid Producing Marine-Derived Fungi as a Valuable Reservoir of Drug Entities. Biomedicines 2021, 9, 485. [Google Scholar] [CrossRef]
- Pandey, A. Pharmacological significance of marine microbial bioactive compounds. Environ. Chem. Lett. 2019, 17, 1741–1751. [Google Scholar] [CrossRef]
- Sang, V.T.; Dat, T.T.H.; Vinh, L.B.; Cuong, L.C.V.; Oanh, P.T.T.; Ha, H.; Kim, Y.H.; Anh, H.L.T.; Yang, S.Y. Coral and Coral-Associated Microorganisms: A Prolific Source of Potential Bioactive Natural Products. Mar. Drugs 2019, 17, 468. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.D.; Vitorino, I.; Reyes, F.; Vicente, F.; Lage, O.M. From Ocean to Medicine: Pharmaceutical Applications of Metabolites from Marine Bacteria. Antibiotics 2020, 9, 455. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Abdel-Wahab, N.M.; Hassan, H.M.; Abdelmohsen, U.R. Saccharopolyspora: An underexplored source for bioactive natural products. J. Appl. Microbiol. 2020, 128, 314–329. [Google Scholar] [CrossRef]
- Xu, J.L.; Liu, Z.F.; Zhang, X.W.; Liu, H.L.; Wang, Y. Microbial Oligosaccharides with Biomedical Applications. Mar. Drugs 2021, 19, 350. [Google Scholar] [CrossRef] [PubMed]
- Shady, N.H.; Fouad, M.A.; Salah Kamel, M.; Schirmeister, T.; Abdelmohsen, U.R. Natural Product Repertoire of the Genus Amphimedon. Mar. Drugs 2018, 17, 19. [Google Scholar] [CrossRef] [PubMed]
- Varijakzhan, D.; Loh, J.Y.; Yap, W.S.; Yusoff, K.; Seboussi, R.; Lim, S.E.; Lai, K.S.; Chong, C.M. Bioactive Compounds from Marine Sponges: Fundamentals and Applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef]
- Nabeelah Bibi, S.; Fawzi, M.M.; Gokhan, Z.; Rajesh, J.; Nadeem, N.; Rengasamy Kannan, R.R.; Albuquerque, R.D.D.G.; Pandian, S.K. Ethnopharmacology, Phytochemistry, and Global Distribution of Mangroves—A Comprehensive Review. Mar Drugs 2019, 17, 231. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Sharifi-Rad, J.; Martorell, M.; Ali, E.S.; Asghar, M.N.; Deeba, F.; Firoz, C.K.; Mubarak, M.S. Chemical profile and therapeutic potentials of Xylocarpus moluccensis (Lam.) M. Roem.: A literature-based review. J. Ethnopharmacol. 2020, 259, 112958. [Google Scholar] [CrossRef]
- Jia, S.L.; Chi, Z.; Liu, G.L.; Hu, Z.; Chi, Z.M. Fungi in mangrove ecosystems and their potential applications. Crit. Rev. Biotechnol. 2020, 40, 852–864. [Google Scholar] [CrossRef]
- Hanif, N.; Murni, A.; Tanaka, C.; Tanaka, J. Marine Natural Products from Indonesian Waters. Mar. Drugs 2019, 17, 364. [Google Scholar] [CrossRef]
- El-Hossary, E.M.; Abdel-Halim, M.; Ibrahim, E.S.; Pimentel-Elardo, S.M.; Nodwell, J.R.; Handoussa, H.; Abdelwahab, M.F.; Holzgrabe, U.; Abdelmohsen, U.R. Natural Products Repertoire of the Red Sea. Mar. Drugs 2020, 18, 457. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wu, W.; Liu, X.; Zaleta-Pinet, D.A.; Clark, B.R. Bioactive Compounds Isolated from Marine-Derived Microbes in China: 2009–2018. Mar. Drugs 2019, 17, 339. [Google Scholar] [CrossRef] [PubMed]
- Pech-Puch, D.; Perez-Povedano, M.; Gomez, P.; Martinez-Guitian, M.; Lasarte-Monterrubio, C.; Vazquez-Ucha, J.C.; Novoa-Olmedo, M.L.; Guillen-Hernandez, S.; Villegas-Hernandez, H.; Bou, G.; et al. Marine Organisms from the Yucatan Peninsula (Mexico) as a Potential Natural Source of Antibacterial Compounds. Mar. Drugs 2020, 18, 369. [Google Scholar] [CrossRef] [PubMed]
- Shinde, P.; Banerjee, P.; Mandhare, A. Marine natural products as source of new drugs: A patent review (2015–2018). Expert Opin. Ther. Pat. 2019, 29, 283–309. [Google Scholar] [CrossRef] [PubMed]
- Pereira, F. Have marine natural product drug discovery efforts been productive and how can we improve their efficiency? Expert Opin. Drug Discov. 2019, 14, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J. Natural Product Based Antibody Drug Conjugates: Clinical Status as of November 9, 2020. J. Nat. Prod. 2021, 84, 917–931. [Google Scholar] [CrossRef] [PubMed]
- Andryukov, B.; Mikhailov, V.; Besednova, N. The biotechnological potential of secondary metabolites from marine bacteria. J. Mar. Sci. Eng 2019, 7, 176. [Google Scholar] [CrossRef]
- Stincone, P.; Brandelli, A. Marine bacteria as source of antimicrobial compounds. Crit. Rev. Biotechnol. 2020, 40, 306–319. [Google Scholar] [CrossRef]
- Datta, S.R.; Roy, A. Antimicrobial peptides as potential therapeutic agents: A review. Int. J. Pept. Res. Ther. 2021, 27, 555–577. [Google Scholar] [CrossRef]
- Zhang, S.; Liang, X.; Gadd, G.M.; Zhao, Q. Marine Microbial-Derived Antibiotics and Biosurfactants as Potential New Agents against Catheter-Associated Urinary Tract Infections. Mar. Drugs 2021, 19, 255. [Google Scholar] [CrossRef]
- Borges, A.; Simoes, M. Quorum Sensing Inhibition by Marine Bacteria. Mar. Drugs 2019, 17, 427. [Google Scholar] [CrossRef]
- Chen, J.; Wang, B.; Lu, Y.; Guo, Y.; Sun, J.; Wei, B.; Zhang, H.; Wang, H. Quorum Sensing Inhibitors from Marine Microorganisms and Their Synthetic Derivatives. Mar. Drugs 2019, 17, 80. [Google Scholar] [CrossRef]
- Torres, M.; Dessaux, Y.; Llamas, I. Saline Environments as a Source of Potential Quorum Sensing Disruptors to Control Bacterial Infections: A Review. Mar. Drugs 2019, 17, 191. [Google Scholar] [CrossRef]
- Zhao, J.; Li, X.; Hou, X.; Quan, C.; Chen, M. Widespread Existence of Quorum Sensing Inhibitors in Marine Bacteria: Potential Drugs to Combat Pathogens with Novel Strategies. Mar. Drugs 2019, 17, 275. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, S.; Shen, J.; Zhu, K. Nonribosomal antibacterial peptides that target multidrug-resistant bacteria. Nat. Prod. Rep. 2019, 36, 573–592. [Google Scholar] [CrossRef]
- Barbosa, F.; Pinto, E.; Kijjoa, A.; Pinto, M.; Sousa, E. Targeting antimicrobial drug resistance with marine natural products. Int. J. Antimicrob. Agents 2020, 56, 106005. [Google Scholar] [CrossRef]
- Liu, M.; El-Hossary, E.M.; Oelschlaeger, T.A.; Donia, M.S.; Quinn, R.J.; Abdelmohsen, U.R. Potential of marine natural products against drug-resistant bacterial infections. Lancet Infect. Dis. 2019, 19, e237–e245. [Google Scholar] [CrossRef]
- Casertano, M.; Menna, M.; Imperatore, C. The Ascidian-Derived Metabolites with Antimicrobial Properties. Antibiotics 2020, 9, 510. [Google Scholar] [CrossRef]
- Neshani, A.; Zare, H.; Akbari Eidgahi, M.R.; Khaledi, A.; Ghazvini, K. Epinecidin-1, a highly potent marine antimicrobial peptide with anticancer and immunomodulatory activities. BMC Pharmacol. Toxicol. 2019, 20, 33. [Google Scholar] [CrossRef]
- Portelinha, J.; Duay, S.S.; Yu, S.I.; Heilemann, K.; Libardo, M.D.J.; Juliano, S.A.; Klassen, J.L.; Angeles-Boza, A.M. Antimicrobial Peptides and Copper(II) Ions: Novel Therapeutic Opportunities. Chem. Rev. 2021, 121, 2648–2712. [Google Scholar] [CrossRef] [PubMed]
- Willems, T.; De Mol, M.L.; De Bruycker, A.; De Maeseneire, S.L.; Soetaert, W.K. Alkaloids from Marine Fungi: Promising Antimicrobials. Antibiotics 2020, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Tang, S.; Cao, S. Antimicrobial compounds from marine fungi. Phytochem. Rev. 2021, 20, 85–117. [Google Scholar] [CrossRef]
- Karpinski, T.M. Marine Macrolides with Antibacterial and/or Antifungal Activity. Mar. Drugs 2019, 17, 241. [Google Scholar] [CrossRef] [PubMed]
- Alves, E.; Dias, M.; Lopes, D.; Almeida, A.; Domingues, M.D.R.; Rey, F. Antimicrobial Lipids from Plants and Marine Organisms: An Overview of the Current State-of-the-Art and Future Prospects. Antibiotics 2020, 9, 441. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.C.; Resende, D.; da Costa, P.M.; Pinto, M.M.M.; Sousa, E. Tryptophan derived natural marine alkaloids and synthetic derivatives as promising antimicrobial agents. Eur. J. Med. Chem. 2021, 209, 112945. [Google Scholar] [CrossRef] [PubMed]
- Arockianathan, P.M.; Mishra, M.; Niranjan, R. Recent Status and Advancements in the Development of Antifungal Agents: Highlights on Plant and Marine Based Antifungals. Curr. Top. Med. Chem. 2019, 19, 812–830. [Google Scholar] [CrossRef] [PubMed]
- Christy, M.P.; Uekusa, Y.; Gerwick, L.; Gerwick, W.H. Natural Products with Potential to Treat RNA Virus Pathogens Including SARS-CoV-2. J. Nat. Prod. 2021, 84, 161–182. [Google Scholar] [CrossRef] [PubMed]
- Hamoda, A.M.; Fayed, B.; Ashmawy, N.S.; El-Shorbagi, A.A.; Hamdy, R.; Soliman, S.S.M. Marine Sponge is a Promising Natural Source of Anti-SARS-CoV-2 Scaffold. Front. Pharmacol. 2021, 12, 666664. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Ali, A.; Wang, Q.; Irfan, M.; Khan, A.; Zeb, M.T.; Zhang, Y.J.; Chinnasamy, S.; Wei, D.Q. Marine natural compounds as potents inhibitors against the main protease of SARS-CoV-2—A molecular dynamic study. J. Biomol. Struct. Dyn. 2021, 39, 3627–3637. [Google Scholar] [CrossRef]
- El-Tantawy, W.H.; Temraz, A. Natural products for the management of the hepatitis C virus: A biochemical review. Arch. Physiol. Biochem. 2020, 126, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.T.; Zhu, H.J.; Cao, F. Marine Natural Products as a Source of Drug Leads against Respiratory Viruses: Structural and Bioactive Diversity. Curr. Med. Chem. 2021, 28, 3568–3594. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.; Brunet, C.; Noonan, D.M.; Albini, A. Marine Algal Antioxidants as Potential Vectors for Controlling Viral Diseases. Antioxidants 2020, 9, 392. [Google Scholar] [CrossRef] [PubMed]
- Pagarete, A.; Ramos, A.S.; Puntervoll, P.; Allen, M.J.; Verdelho, V. Antiviral Potential of Algal Metabolites—A Comprehensive Review. Mar. Drugs 2021, 19, 94. [Google Scholar] [CrossRef] [PubMed]
- Besednova, N.N.; Andryukov, B.G.; Zaporozhets, T.S.; Kryzhanovsky, S.P.; Fedyanina, L.N.; Kuznetsova, T.A.; Zvyagintseva, T.N.; Shchelkanov, M.Y. Antiviral Effects of Polyphenols from Marine Algae. Biomedicines 2021, 9, 200. [Google Scholar] [CrossRef]
- Diaz-Marrero, A.R.; Lopez-Arencibia, A.; Bethencout-Estrella, C.J.; Cen-Pacheco, F.; Sifaoui, I.; Hernandez Creus, A.; Duque-Ramirez, M.C.; Souto, M.L.; Hernandez Daranas, A.; Lorenzo-Morales, J.; et al. Antiprotozoal activities of marine polyether triterpenoids. Bioorg. Chem. 2019, 92, 103276. [Google Scholar] [CrossRef]
- Lee, S.M.; Kim, M.S.; Hayat, F.; Shin, D. Recent Advances in the Discovery of Novel Antiprotozoal Agents. Molecules 2019, 24, 3886. [Google Scholar] [CrossRef]
- Zhang, S.; Kavianinia, I.; Brimble, M. Naturally occurring antitubercular cyclic peptides. Tetrahedron Lett. 2019, 60, 151339. [Google Scholar] [CrossRef]
- Alvarez-Bardon, M.; Perez-Pertejo, Y.; Ordonez, C.; Sepulveda-Crespo, D.; Carballeira, N.M.; Tekwani, B.L.; Murugesan, S.; Martinez-Valladares, M.; Garcia-Estrada, C.; Reguera, R.M.; et al. Screening Marine Natural Products for New Drug Leads against Trypanosomatids and Malaria. Mar. Drugs 2020, 18, 187. [Google Scholar] [CrossRef]
- Nweze, J.A.; Mbaoji, F.N.; Li, Y.M.; Yang, L.Y.; Huang, S.S.; Chigor, V.N.; Eze, E.A.; Pan, L.X.; Zhang, T.; Yang, D.F. Potentials of marine natural products against malaria, leishmaniasis, and trypanosomiasis parasites: A review of recent articles. Infect. Dis. Poverty 2021, 10, 9. [Google Scholar] [CrossRef]
- Aguiar, A.C.C.; Parisi, J.R.; Granito, R.N.; de Sousa, L.R.F.; Renno, A.C.M.; Gazarini, M.L. Metabolites from Marine Sponges and Their Potential to Treat Malarial Protozoan Parasites Infection: A Systematic Review. Mar. Drugs 2021, 19, 134. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.M.; Wang, C.Y.; Gerwick, W.H.; Shao, C.L. Marine natural products as potential anti-tubercular agents. Eur. J. Med. Chem. 2019, 165, 273–292. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Kaushik, A.C.; Bhatti, A.I.; Zhang, Y.J.; Zhang, S.; Wei, A.J.; Malik, S.I.; Wei, D.Q. Marine Natural Products and Drug Resistance in Latent Tuberculosis. Mar. Drugs 2019, 17, 549. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.M.; Zheng, K.W. Marine natural products with anti-inflammation effects. Tradit. Med. Res. 2020, 5, 252–260. [Google Scholar] [CrossRef]
- Bilal, M.; Qindeel, M.; Nunes, L.V.; Duarte, M.T.S.; Ferreira, L.F.R.; Soriano, R.N.; Iqbal, H.M.N. Marine-Derived Biologically Active Compounds for the Potential Treatment of Rheumatoid Arthritis. Mar. Drugs 2020, 19, 10. [Google Scholar] [CrossRef]
- Souza, C.R.M.; Bezerra, W.P.; Souto, J.T. Marine Alkaloids with Anti-Inflammatory Activity: Current Knowledge and Future Perspectives. Mar. Drugs 2020, 18, 147. [Google Scholar] [CrossRef]
- Xu, J.; Yi, M.; Ding, L.; He, S. A Review of Anti-Inflammatory Compounds from Marine Fungi, 2000–2018. Mar. Drugs 2019, 17, 636. [Google Scholar] [CrossRef]
- Di Costanzo, F.; Di Dato, V.; Ianora, A.; Romano, G. Prostaglandins in Marine Organisms: A Review. Mar. Drugs 2019, 17, 428. [Google Scholar] [CrossRef]
- Kemp, D.C.; Kwon, J.Y. Fish and Shellfish-Derived Anti-Inflammatory Protein Products: Properties and Mechanisms. Molecules 2021, 26, 3225. [Google Scholar] [CrossRef]
- Ahmad, S.; Saleem, M.; Riaz, N.; Lee, Y.S.; Diri, R.; Noor, A.; Almasri, D.; Bagalagel, A.; Elsebai, M.F. The Natural Polypeptides as Significant Elastase Inhibitors. Front. Pharmacol. 2020, 11, 688. [Google Scholar] [CrossRef]
- Zhang, Y.; Min, J.; Zhang, L. Anti-inflammatory and immunomodulatory effects of marine n-3 polyunsaturated fatty acids on human health and diseases. J. Ocean Univ. China 2019, 18, 481–492. [Google Scholar] [CrossRef]
- Rausch, J.; Gillespie, S.; Orchard, T.; Tan, A.; McDaniel, J.C. Systematic review of marine-derived omega-3 fatty acid supplementation effects on leptin, adiponectin, and the leptin-to-adiponectin ratio. Nutr. Res. 2021, 85, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Meng, Y.; Li, N.; Wang, Q.; Chen, L. The effects of low-ratio n-6/n-3 PUFA on biomarkers of inflammation: A systematic review and meta-analysis. Food Funct. 2021, 12, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, W.; Chen, Y.; Wan, X.; Wang, J. Fucoxanthin: A promising compound for human inflammation-related diseases. Life Sci. 2020, 255, 117850. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, R. Antioxidant properties of marine algae: An overview. Biosci. Res. 2019, 16, 986–996. [Google Scholar]
- Saraswati; Giriwono, P.E.; Iskandriati, D.; Tan, C.P.; Andarwulan, N. Sargassum Seaweed as a Source of Anti-Inflammatory Substances and the Potential Insight of the Tropical Species: A Review. Mar. Drugs 2019, 17, 590. [Google Scholar] [CrossRef]
- Riccio, G.; Lauritano, C. Microalgae with Immunomodulatory Activities. Mar. Drugs 2019, 18, 2. [Google Scholar] [CrossRef]
- Dolmatova, L.S.; Dolmatov, I.Y. Different Macrophage Type Triggering as Target of the Action of Biologically Active Substances from Marine Invertebrates. Mar. Drugs 2020, 18, 37. [Google Scholar] [CrossRef]
- Rubilar, T.; Barbieri, E.S.; Gazquez, A.; Avaro, M. Sea Urchin Pigments: Echinochrome A and Its Potential Implication in the Cytokine Storm Syndrome. Mar. Drugs 2021, 19, 267. [Google Scholar] [CrossRef]
- Sanina, N. Vaccine Adjuvants Derived from Marine Organisms. Biomolecules 2019, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Zhao, J.; Xing, M.; Xiao, H.; Zhang, Q.; Liang, H.; Ji, A.; Song, S. Current Research Landscape of Marine-Derived Anti-Atherosclerotic Substances. Mar. Drugs 2020, 18, 440. [Google Scholar] [CrossRef]
- Zhao, J.; Cao, Q.; Xing, M.; Xiao, H.; Cheng, Z.; Song, S.; Ji, A. Advances in the Study of Marine Products with Lipid-Lowering Properties. Mar. Drugs 2020, 18, 390. [Google Scholar] [CrossRef]
- Dwivedi, R.; Pomin, V.H. Marine Antithrombotics. Mar. Drugs 2020, 18, 514. [Google Scholar] [CrossRef] [PubMed]
- Doshi, G.; Nailwal, N. A Review on Molecular Mechanisms and Patents of Marine-derived Anti-thrombotic Agents. Curr. Drug Targets 2021, 22, 318–335. [Google Scholar] [CrossRef]
- Carvalhal, F.; Cristelo, R.R.; Resende, D.; Pinto, M.M.M.; Sousa, E.; Correia-da-Silva, M. Antithrombotics from the Sea: Polysaccharides and Beyond. Mar. Drugs 2019, 17, 170. [Google Scholar] [CrossRef] [PubMed]
- Barkia, I.; Saari, N.; Manning, S.R. Microalgae for High-Value Products Towards Human Health and Nutrition. Mar. Drugs 2019, 17, 304. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.W.; Fernando, K.H.N.; Oh, J.Y.; Li, X.; Jeon, Y.J.; Ryu, B. Anti-Obesity and Anti-Diabetic Effects of Ishige okamurae. Mar. Drugs 2019, 17, 202. [Google Scholar] [CrossRef]
- Chen, L.; Liu, R.; He, X.; Pei, S.; Li, D. Effects of brown seaweed polyphenols, a class of phlorotannins, on metabolic disorders via regulation of fat function. Food Funct. 2021, 12, 2378–2388. [Google Scholar] [CrossRef]
- Pradhan, B.; Nayak, R.; Patra, S.; Jit, B.P.; Ragusa, A.; Jena, M. Bioactive Metabolites from Marine Algae as Potent Pharmacophores against Oxidative Stress-Associated Human Diseases: A Comprehensive Review. Molecules 2020, 26, 37. [Google Scholar] [CrossRef]
- Rayapu, L.; Chakraborty, K.; Valluru, L. Marine Algae as a Potential Source for Anti-diabetic Compounds—A Brief Review. Curr. Pharm. Des. 2021, 27, 789–801. [Google Scholar] [CrossRef]
- Sachithanandam, V.; Lalitha, P.; Parthiban, A.; Mageswaran, T.; Manmadhan, K.; Sridhar, R. A Review on Antidiabetic Properties of Indian Mangrove Plants with Reference to Island Ecosystem. Evid. Based Complement. Altern. Med. 2019, 2019, 4305148. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.; Kim, M.B.; Park, Y.K.; Lee, J.Y. Health benefits of fucoxanthin in the prevention of chronic diseases. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158618. [Google Scholar] [CrossRef] [PubMed]
- Landon, R.; Gueguen, V.; Petite, H.; Letourneur, D.; Pavon-Djavid, G.; Anagnostou, F. Impact of Astaxanthin on Diabetes Pathogenesis and Chronic Complications. Mar. Drugs 2020, 18, 357. [Google Scholar] [CrossRef] [PubMed]
- Gabbia, D.; De Martin, S. Brown Seaweeds for the Management of Metabolic Syndrome and Associated Diseases. Molecules 2020, 25, 4182. [Google Scholar] [CrossRef] [PubMed]
- Gunathilaka, T.L.; Samarakoon, K.; Ranasinghe, P.; Peiris, L.D.C. Antidiabetic Potential of Marine Brown Algae-a Mini Review. J. Diabetes Res. 2020, 2020, 1230218. [Google Scholar] [CrossRef] [PubMed]
- Balasa, A.F.; Chircov, C.; Grumezescu, A.M. Marine Biocompounds for Neuroprotection—A Review. Mar. Drugs 2020, 18, 290. [Google Scholar] [CrossRef] [PubMed]
- Catanesi, M.; Caioni, G.; Castelli, V.; Benedetti, E.; d’Angelo, M.; Cimini, A. Benefits under the Sea: The Role of Marine Compounds in Neurodegenerative Disorders. Mar. Drugs 2021, 19, 24. [Google Scholar] [CrossRef] [PubMed]
- Ahmmed, M.K.; Ahmmed, F.; Tian, H.; Carne, A.; Bekhit, A.E. Marine omega-3 (n-3) phospholipids: A comprehensive review of their properties, sources, bioavailability, and relation to brain health. Compr. Rev. Food Sci. Food Saf. 2019, 19, 64–123. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.H.; Muttenthaler, M.; Dutertre, S.; Himaya, S.W.A.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Conotoxins: Chemistry and Biology. Chem. Rev. 2019, 119, 11510–11549. [Google Scholar] [CrossRef]
- Jimenez, E.C. Post-translationally modified conopeptides: Biological activities and pharmacological applications. Peptides 2021, 139, 170525. [Google Scholar] [CrossRef]
- Baj, A.; Bistoletti, M.; Bosi, A.; Moro, E.; Giaroni, C.; Crema, F. Marine Toxins and Nociception: Potential Therapeutic Use in the Treatment of Visceral Pain Associated with Gastrointestinal Disorders. Toxins 2019, 11, 449. [Google Scholar] [CrossRef]
- Barbalace, M.C.; Malaguti, M.; Giusti, L.; Lucacchini, A.; Hrelia, S.; Angeloni, C. Anti-Inflammatory Activities of Marine Algae in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 3061. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease. Mar. Drugs 2019, 17, 609. [Google Scholar] [CrossRef]
- Hannan, M.A.; Dash, R.; Haque, M.N.; Mohibbullah, M.; Sohag, A.A.M.; Rahman, M.A.; Uddin, M.J.; Alam, M.; Moon, I.S. Neuroprotective Potentials of Marine Algae and Their Bioactive Metabolites: Pharmacological Insights and Therapeutic Advances. Mar. Drugs 2020, 18, 347. [Google Scholar] [CrossRef]
- Rathnayake, A.U.; Abuine, R.; Kim, Y.J.; Byun, H.G. Anti-Alzheimer’s Materials Isolated from Marine Bio-resources: A Review. Curr. Alzheimer. Res. 2019, 16, 895–906. [Google Scholar] [CrossRef]
- Ciccone, L.; Vandooren, J.; Nencetti, S.; Orlandini, E. Natural Marine and Terrestrial Compounds as Modulators of Matrix Metalloproteinases-2 (MMP-2) and MMP-9 in Alzheimer’s Disease. Pharmaceuticals 2021, 14, 86. [Google Scholar] [CrossRef]
- Kabir, M.T.; Uddin, M.S.; Jeandet, P.; Emran, T.B.; Mitra, S.; Albadrani, G.M.; Sayed, A.A.; Abdel-Daim, M.M.; Simal-Gandara, J. Anti-Alzheimer’s Molecules Derived from Marine Life: Understanding Molecular Mechanisms and Therapeutic Potential. Mar. Drugs 2021, 19, 251. [Google Scholar] [CrossRef]
- Rahman, M.A.; Dash, R.; Sohag, A.A.M.; Alam, M.; Rhim, H.; Ha, H.; Moon, I.S.; Uddin, M.J.; Hannan, M.A. Prospects of Marine Sterols against Pathobiology of Alzheimer’s Disease: Pharmacological Insights and Technological Advances. Mar. Drugs 2021, 19, 167. [Google Scholar] [CrossRef]
- Silva, M.; Seijas, P.; Otero, P. Exploitation of Marine Molecules to Manage Alzheimer’s Disease. Mar. Drugs 2021, 19, 373. [Google Scholar] [CrossRef]
- Castaneda, A.; Ferraz, R.; Vieira, M.; Cardoso, I.; Vasconcelos, V.; Martins, R. Bridging Cyanobacteria to Neurodegenerative Diseases: A New Potential Source of Bioactive Compounds against Alzheimer’s Disease. Mar. Drugs 2021, 19, 343. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, Z.; Cui, W. Marine-Derived Natural Compounds for the Treatment of Parkinson’s Disease. Mar. Drugs 2019, 17, 221. [Google Scholar] [CrossRef]
- Fakhri, S.; Aneva, I.Y.; Farzaei, M.H.; Sobarzo-Sanchez, E. The Neuroprotective Effects of Astaxanthin: Therapeutic Targets and Clinical Perspective. Molecules 2019, 24, 2640. [Google Scholar] [CrossRef]
- Sorrenti, V.; Davinelli, S.; Scapagnini, G.; Willcox, B.J.; Allsopp, R.C.; Willcox, D.C. Astaxanthin as a Putative Geroprotector: Molecular Basis and Focus on Brain Aging. Mar. Drugs 2020, 18, 351. [Google Scholar] [CrossRef]
- Bahbah, E.I.; Ghozy, S.; Attia, M.S.; Negida, A.; Emran, T.B.; Mitra, S.; Albadrani, G.M.; Abdel-Daim, M.M.; Uddin, M.S.; Simal-Gandara, J. Molecular Mechanisms of Astaxanthin as a Potential Neurotherapeutic Agent. Mar Drugs 2021, 19, 201. [Google Scholar] [CrossRef]
- Liao, Q.; Feng, Y.; Yang, B.; Lee, S.M. Cnidarian peptide neurotoxins: A new source of various ion channel modulators or blockers against central nervous systems disease. Drug Discov. Today 2019, 24, 189–197. [Google Scholar] [CrossRef]
- Finol-Urdaneta, R.K.; Belovanovic, A.; Micic-Vicovac, M.; Kinsella, G.K.; McArthur, J.R.; Al-Sabi, A. Marine Toxins Targeting Kv1 Channels: Pharmacological Tools and Therapeutic Scaffolds. Mar. Drugs 2020, 18, 173. [Google Scholar] [CrossRef]
- Stonik, V.A.; Stonik, I.V. Marine Excitatory Amino Acids: Structure, Properties, Biosynthesis and Recent Approaches to Their Syntheses. Molecules 2020, 25, 3049. [Google Scholar] [CrossRef]
- Hong, A.; Tu, L.C.; Yang, I.; Lim, K.M.; Nam, S.J. Marine natural products with monoamine oxidase (MAO) inhibitory activity. Pharm. Biol. 2020, 58, 716–720. [Google Scholar] [CrossRef]
- Barbosa, A.J.M.; Roque, A.C.A. Free Marine Natural Products Databases for Biotechnology and Bioengineering. Biotechnol. J. 2019, 14, e1800607. [Google Scholar] [CrossRef]
- van Santen, J.A.; Kautsar, S.A.; Medema, M.H.; Linington, R.G. Microbial natural product databases: Moving forward in the multi-omics era. Nat. Prod. Rep. 2021, 38, 264–278. [Google Scholar] [CrossRef]
- Stuart, K.A.; Welsh, K.; Walker, M.C.; Edrada-Ebel, R. Metabolomic tools used in marine natural product drug discovery. Expert Opin. Drug Discov. 2020, 15, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E.; Andersen, R.J. Biologically active marine natural products and their molecular targets discovered using a chemical genetics approach. Nat. Prod. Rep. 2020, 37, 617–633. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Umezawa, K. Cellular Signal Transductions and Their Inhibitors Derived from Deep-Sea Organisms. Mar. Drugs 2021, 19, 205. [Google Scholar] [CrossRef] [PubMed]
- Juarez-Portilla, C.; Olivares-Banuelos, T.; Molina-Jimenez, T.; Sanchez-Salcedo, J.A.; Moral, D.I.D.; Meza-Menchaca, T.; Flores-Munoz, M.; Lopez-Franco, O.; Roldan-Roldan, G.; Ortega, A.; et al. Seaweeds-derived compounds modulating effects on signal transduction pathways: A systematic review. Phytomedicine 2019, 63, 153016. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kim, H. Astaxanthin Modulation of Signaling Pathways That Regulate Autophagy. Mar. Drugs 2019, 17, 546. [Google Scholar] [CrossRef] [PubMed]
- Giannaccare, G.; Pellegrini, M.; Senni, C.; Bernabei, F.; Scorcia, V.; Cicero, A.F.G. Clinical Applications of Astaxanthin in the Treatment of Ocular Diseases: Emerging Insights. Mar. Drugs 2020, 18, 239. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, N.; Zhang, T.; Zhang, B.; Sajeevan, T.P.; Joseph, V.; Armstrong, L.; He, S.; Yan, X.; Naman, C.B. A Systematic Review of Recently Reported Marine Derived Natural Product Kinase Inhibitors. Mar. Drugs 2019, 17, 493. [Google Scholar] [CrossRef] [PubMed]
- Parate, S.; Kumar, V.; Lee, G.; Rampogu, S.; Hong, J.C.; Lee, K.W. Marine-Derived Natural Products as ATP-Competitive mTOR Kinase Inhibitors for Cancer Therapeutics. Pharmaceuticals 2021, 14, 282. [Google Scholar] [CrossRef] [PubMed]
- Raghuvanshi, R.; Bharate, S.B. Preclinical and Clinical Studies on Bryostatins, A Class of Marine-Derived Protein Kinase C Modulators: A Mini-Review. Curr. Top. Med. Chem. 2020, 20, 1124–1135. [Google Scholar] [CrossRef]
- Wu, R.; Chen, H.; Chang, N.; Xu, Y.; Jiao, J.; Zhang, H. Unlocking the Drug Potential of the Bryostatin Family: Recent Advances in Product Synthesis and Biomedical Applications. Chemistry 2020, 26, 1166–1195. [Google Scholar] [CrossRef]
- Luesch, H.; Paavilainen, V.O. Natural products as modulators of eukaryotic protein secretion. Nat. Prod. Rep. 2020, 37, 717–736. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Matsumori, N. Chemical diversity and mode of action of natural products targeting lipids in the eukaryotic cell membrane. Nat. Prod. Rep. 2020, 37, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Risinger, A.L.; Du, L. Targeting and extending the eukaryotic druggable genome with natural products: Cytoskeletal targets of natural products. Nat. Prod. Rep. 2020, 37, 634–652. [Google Scholar] [CrossRef]
- Carazo, A.; Mladenka, P.; Pavek, P. Marine Ligands of the Pregnane X Receptor (PXR): An Overview. Mar. Drugs 2019, 17, 554. [Google Scholar] [CrossRef] [PubMed]
- Vasilopoulou, M.; Ioannou, E.; Roussis, V.; Chondrogianni, N. Modulation of the ubiquitin-proteasome system by marine natural products. Redox Biol. 2021, 41, 101897. [Google Scholar] [CrossRef] [PubMed]
- González-Andrés, P.; Fernández-Peña, L.; Díez-Poza, C.; Villalobos, C.; Nuñez, L.; Barbero, A. Marine Heterocyclic Compounds That Modulate Intracellular Calcium Signals: Chemistry and Synthesis Approaches. Mar. Drugs 2021, 19, 78. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, H.; Waditee-Sirisattha, R. Antioxidative, Anti-Inflammatory, and Anti-Aging Properties of Mycosporine-Like Amino Acids: Molecular and Cellular Mechanisms in the Protection of Skin-Aging. Mar. Drugs 2019, 17, 222. [Google Scholar] [CrossRef]
- Rosic, N.N. Mycosporine-Like Amino Acids: Making the Foundation for Organic Personalised Sunscreens. Mar. Drugs 2019, 17, 638. [Google Scholar] [CrossRef]
- Nowruzi, B.; Sarvari, G.; Blanco, S. The cosmetic application of cyanobacterial secondary metabolites. Algal Res. 2020, 49, 101959. [Google Scholar] [CrossRef]
- Jesumani, V.; Du, H.; Aslam, M.; Pei, P.; Huang, N. Potential Use of Seaweed Bioactive Compounds in Skincare—A Review. Mar. Drugs 2019, 17, 688. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Seaweeds as nutraceuticals for health and nutrition. Phycologia 2019, 58, 563–577. [Google Scholar] [CrossRef]
- Thiyagarasaiyar, K.; Goh, B.H.; Jeon, Y.J.; Yow, Y.Y. Algae Metabolites in Cosmeceutical: An Overview of Current Applications and Challenges. Mar. Drugs 2020, 18, 323. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayer, A.M.S.; Mayer, V.A.; Swanson-Mungerson, M.; Pierce, M.L.; Rodríguez, A.D.; Nakamura, F.; Taglialatela-Scafati, O. Marine Pharmacology in 2019–2021: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2024, 22, 309. https://doi.org/10.3390/md22070309
Mayer AMS, Mayer VA, Swanson-Mungerson M, Pierce ML, Rodríguez AD, Nakamura F, Taglialatela-Scafati O. Marine Pharmacology in 2019–2021: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Marine Drugs. 2024; 22(7):309. https://doi.org/10.3390/md22070309
Chicago/Turabian StyleMayer, Alejandro M. S., Veronica A. Mayer, Michelle Swanson-Mungerson, Marsha L. Pierce, Abimael D. Rodríguez, Fumiaki Nakamura, and Orazio Taglialatela-Scafati. 2024. "Marine Pharmacology in 2019–2021: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action" Marine Drugs 22, no. 7: 309. https://doi.org/10.3390/md22070309