
Antibacterial and Immunosuppressive Effects of a Novel Marine Brown Alga-Derived Ester in Atopic Dermatitis

 , , , ,
, , , ,
Abstract

1. Introduction
2. Results and Discussion
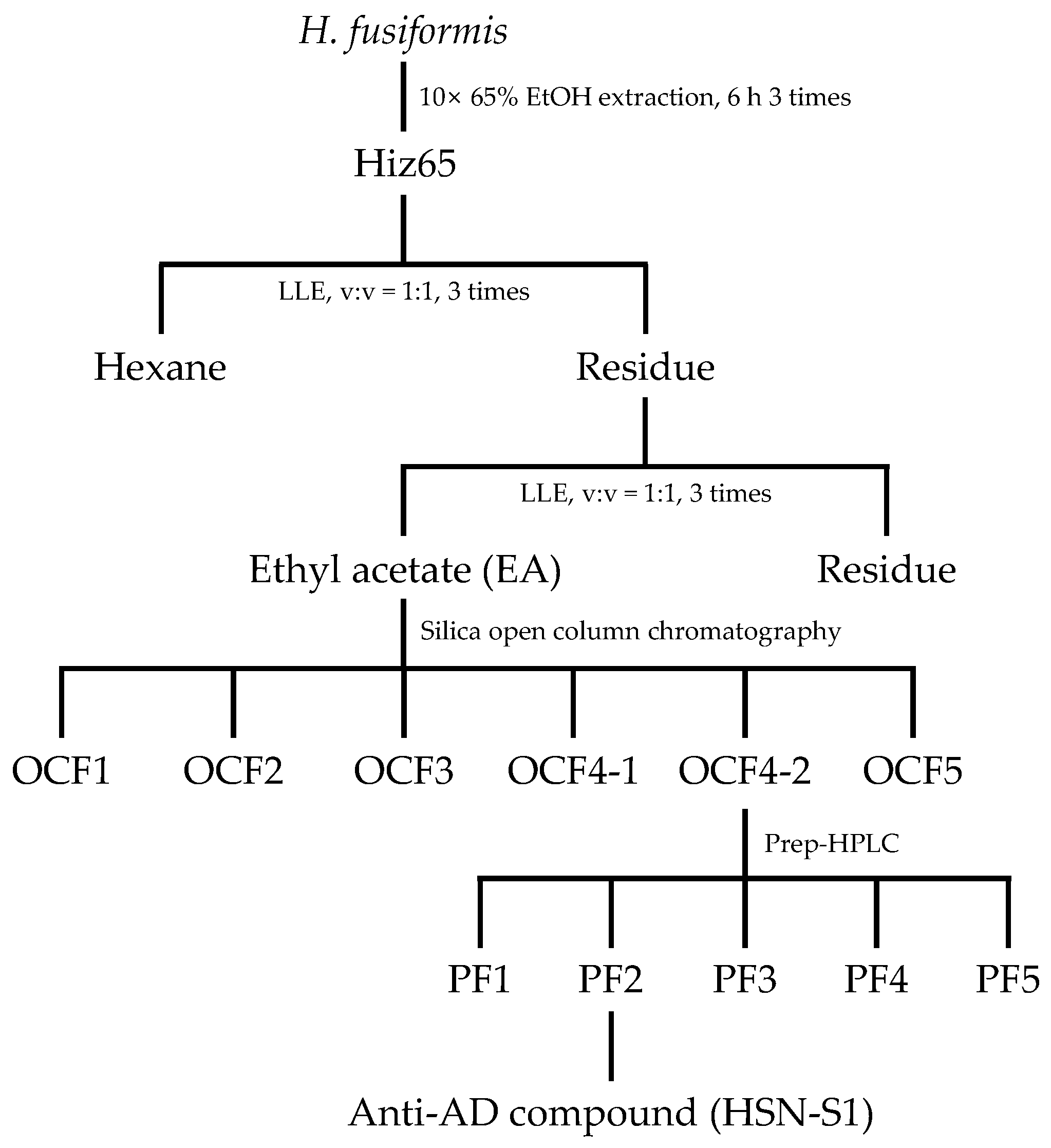
2.1. Isolation of HSN-S1 via Bioactivity-Guided Approach
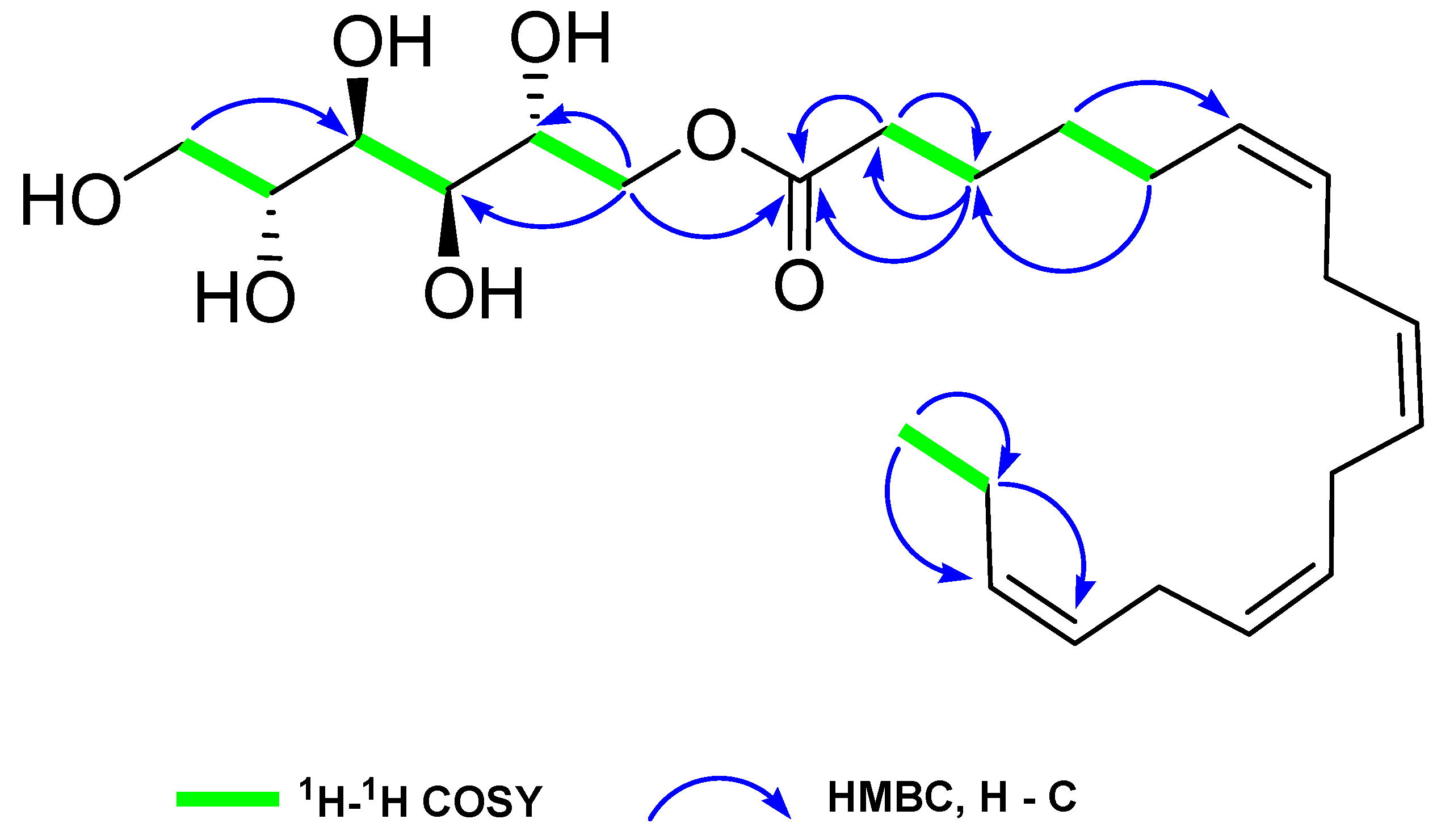
2.2. Structural Characterization of HSN-S1
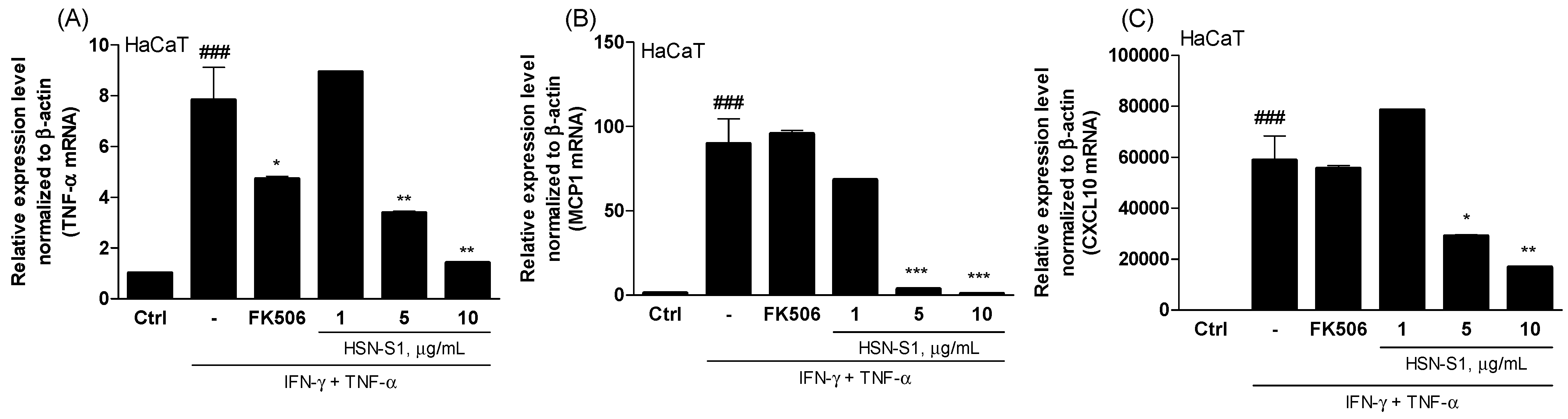
2.3. Effectiveness of HSN-S1 on Keratinocytes
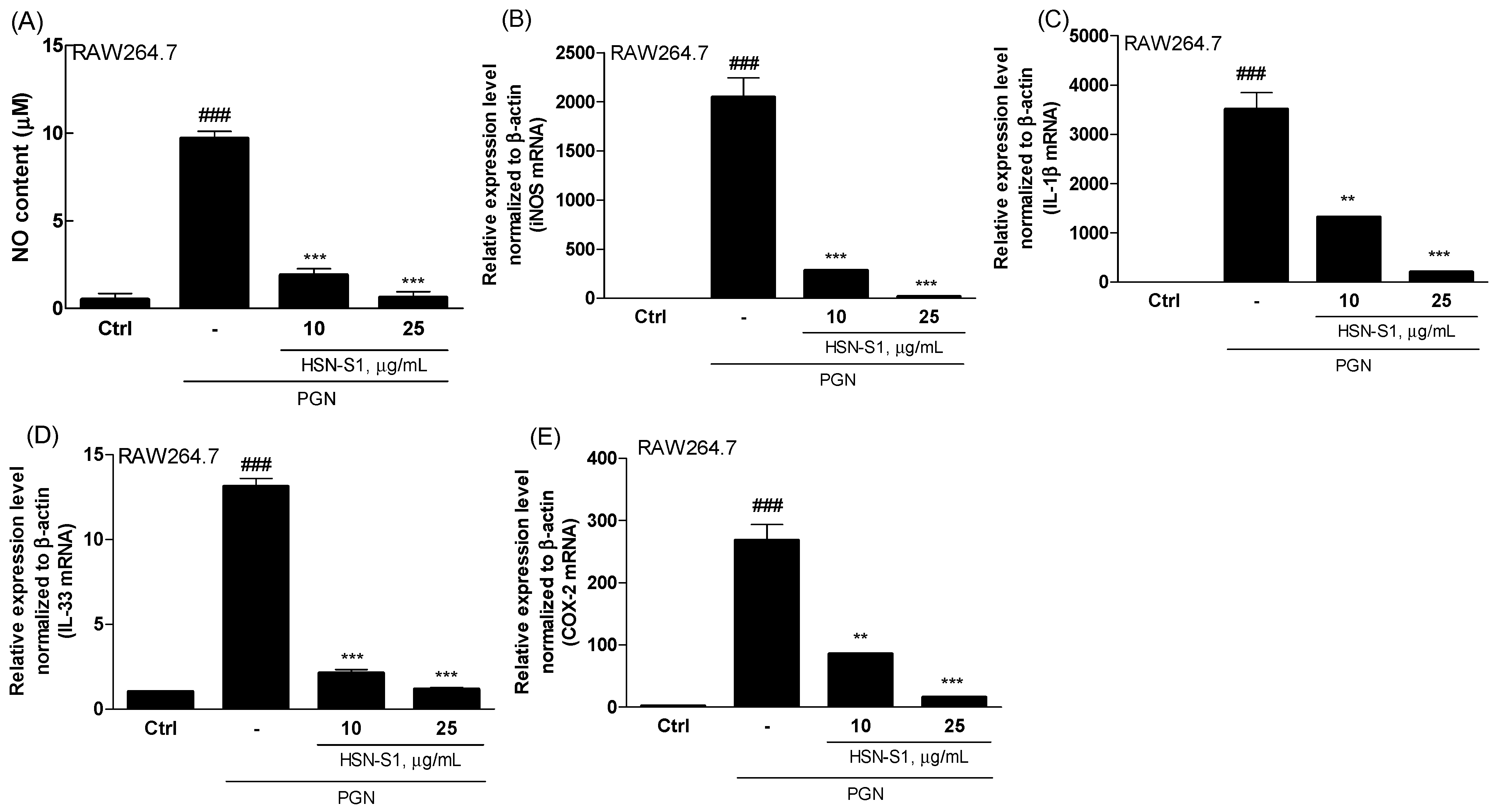
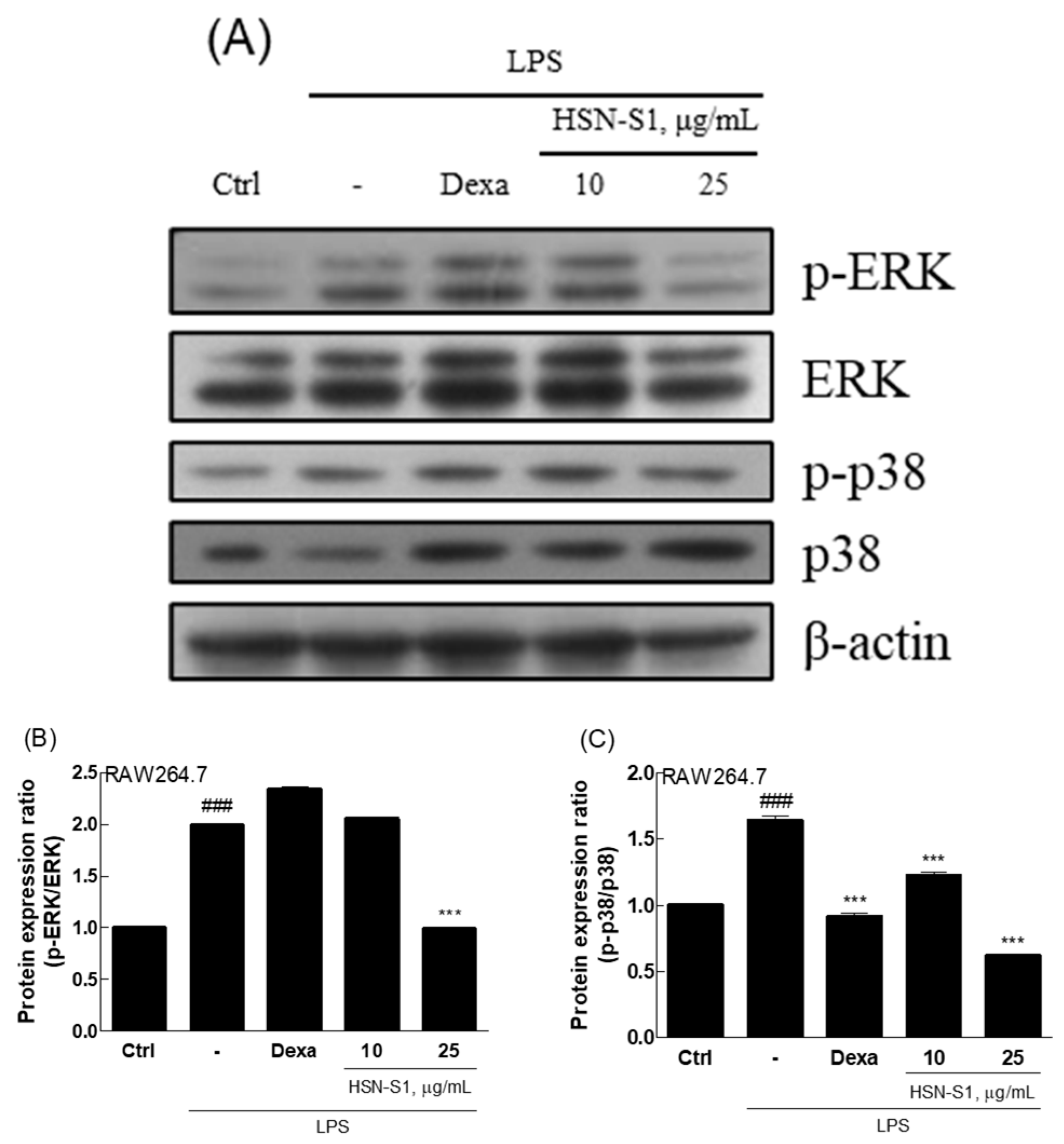
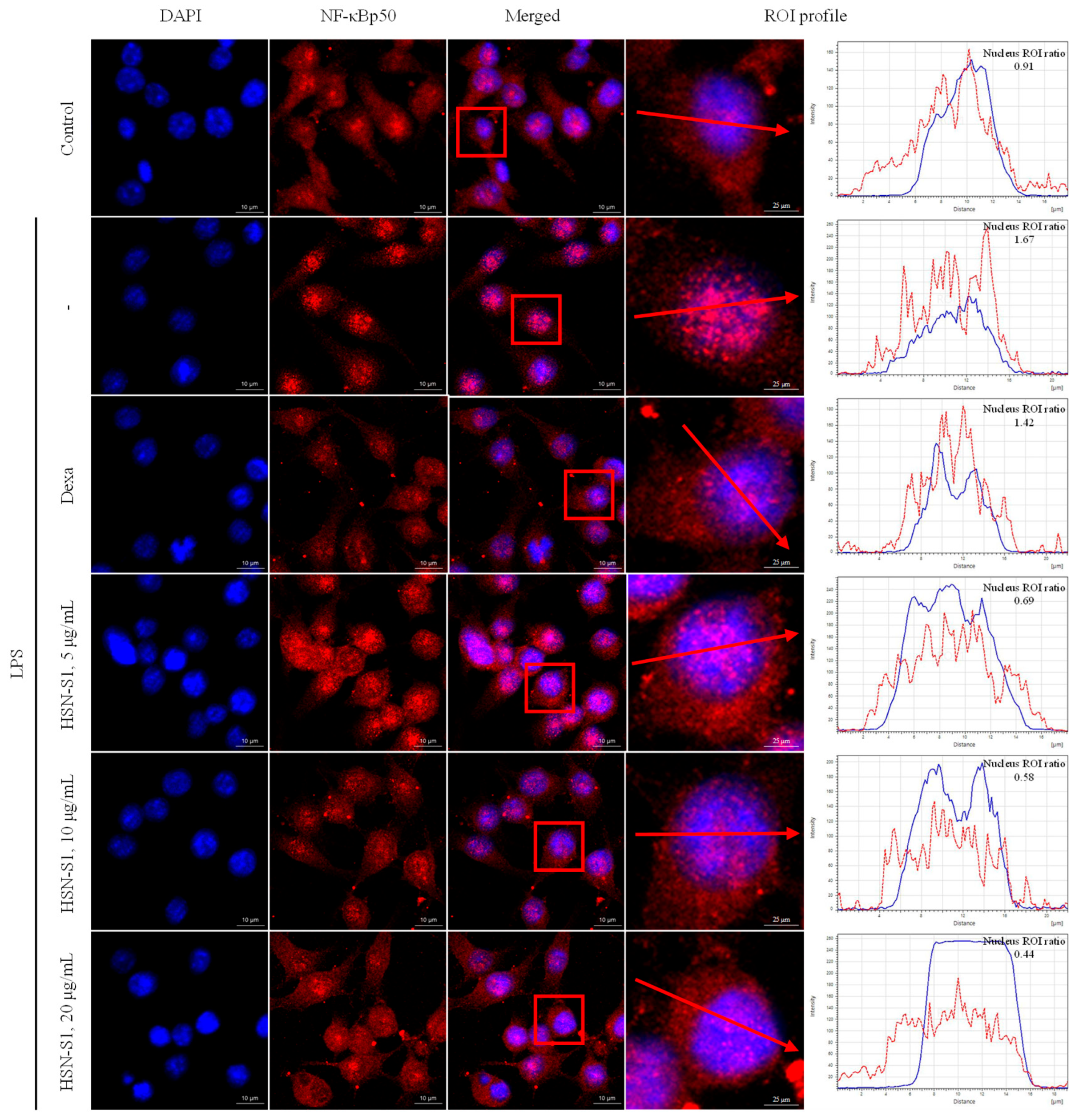
2.4. Effect of HSN-S1 on Macrophage-Mediated Inflammation
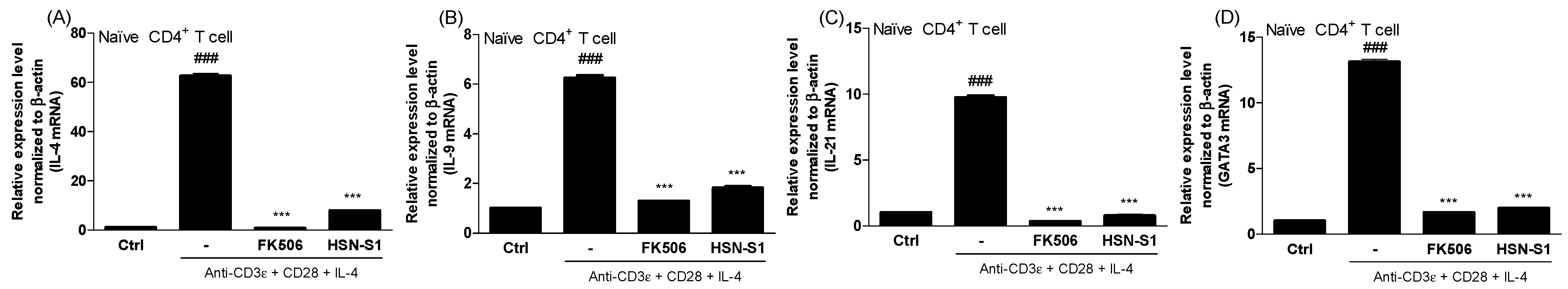
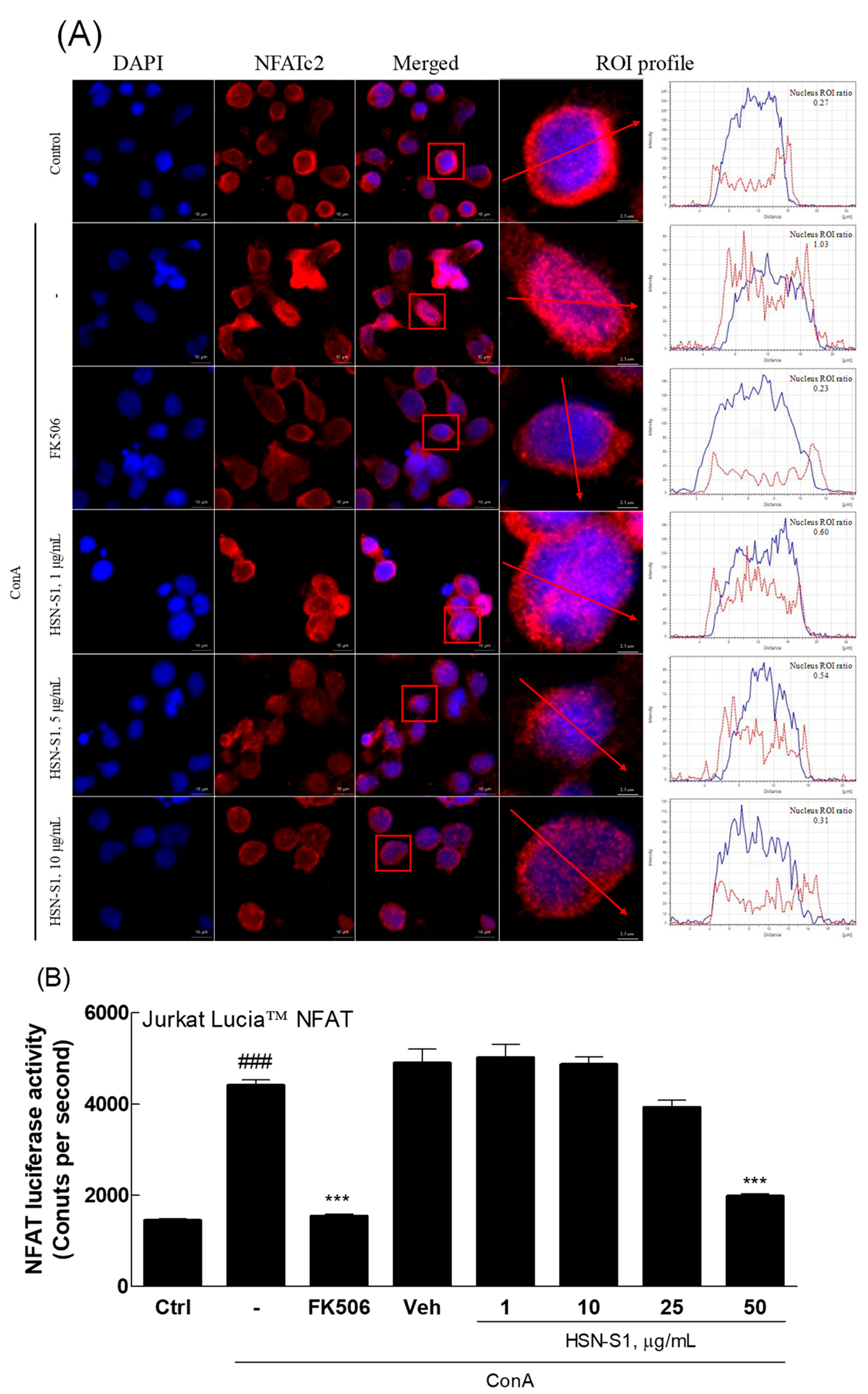
2.5. Immunomodulatory Actions of HSN-S1 in Th Cells
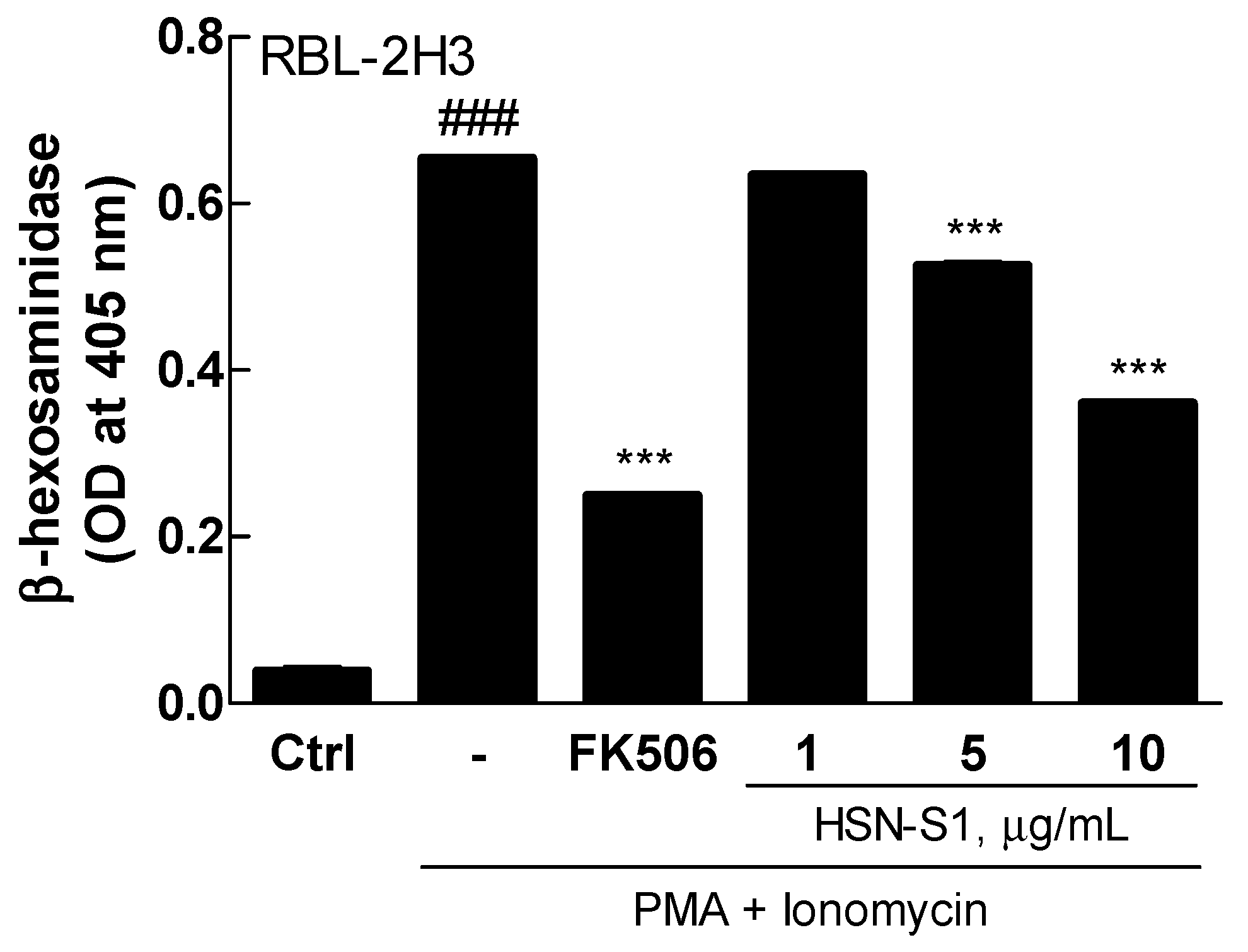
2.6. HSN-S1 and Mast Cell Degranulation: A β-Hexosaminidase Release Assay
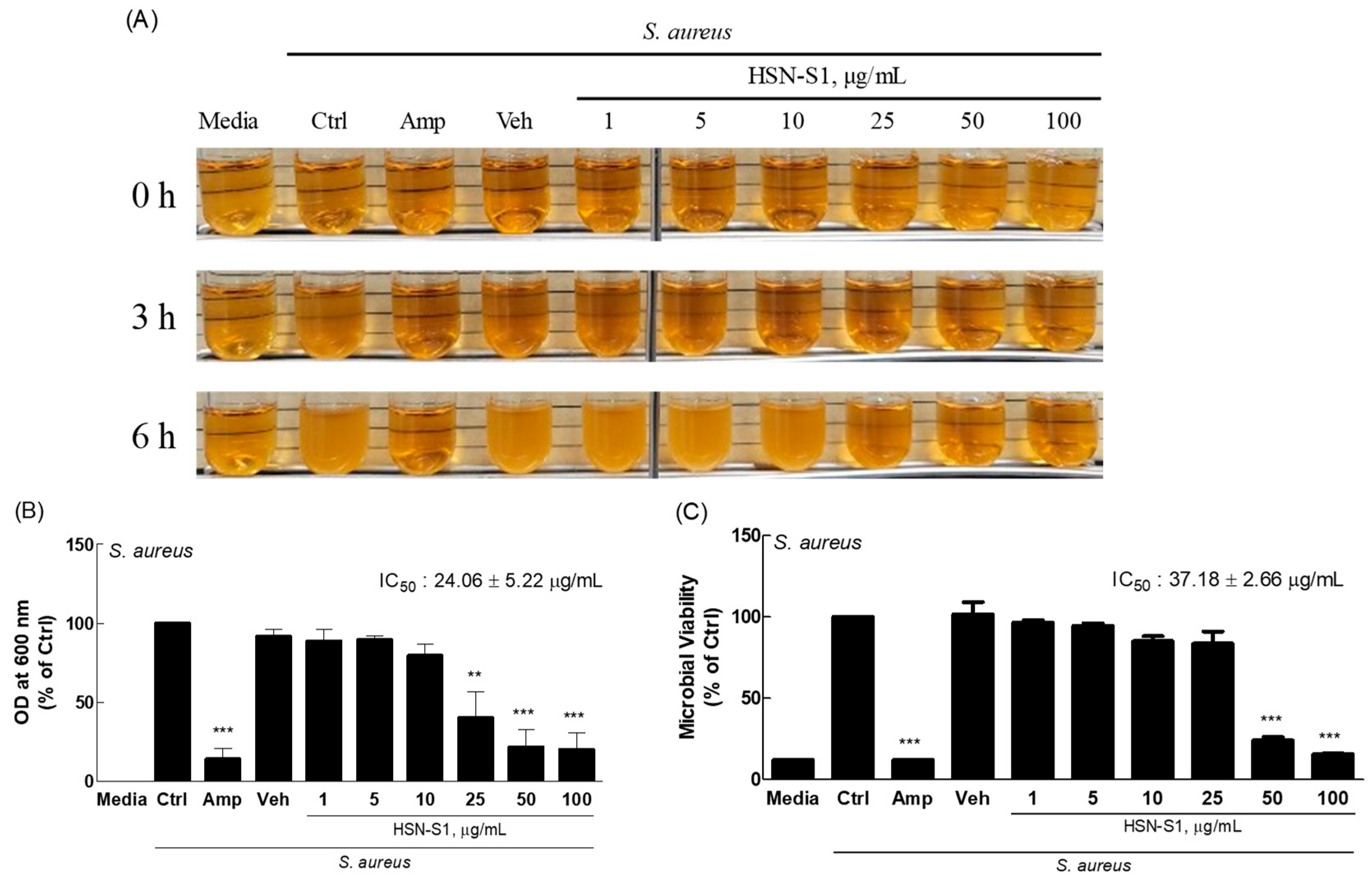
2.7. Antimicrobial Activity of HSN-S1 Against S. aureus
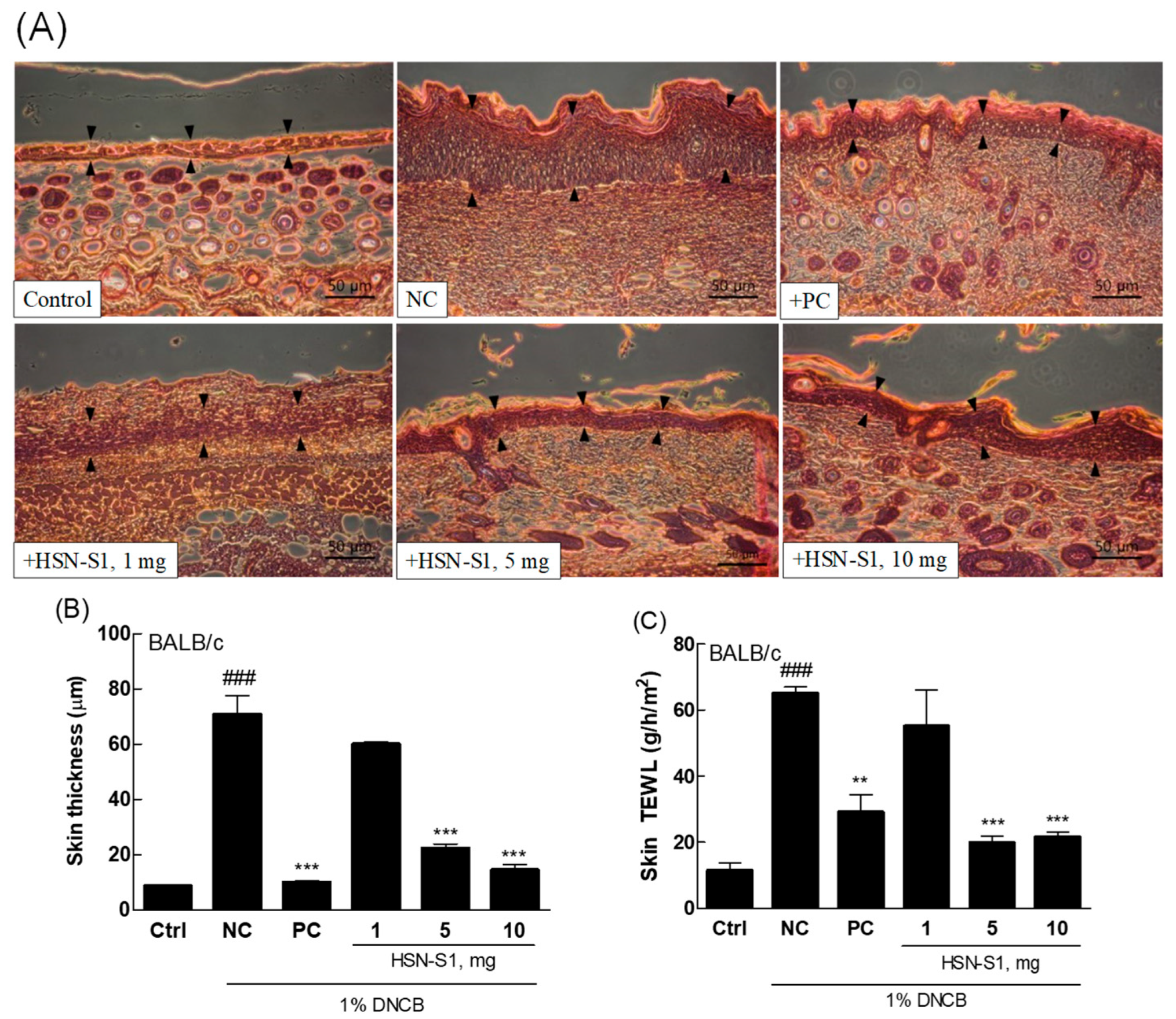
2.8. HSN-S1’s Role in Reducing Epidermal Lichenification
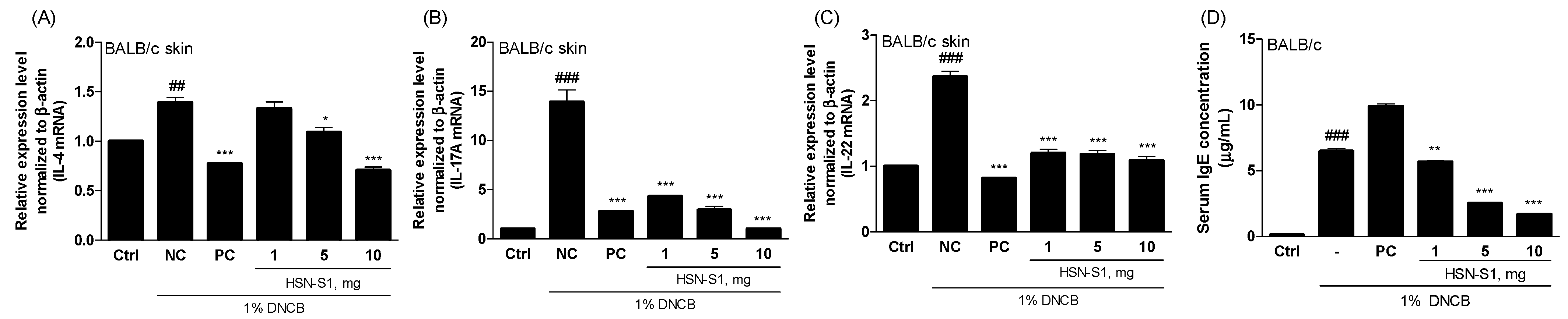
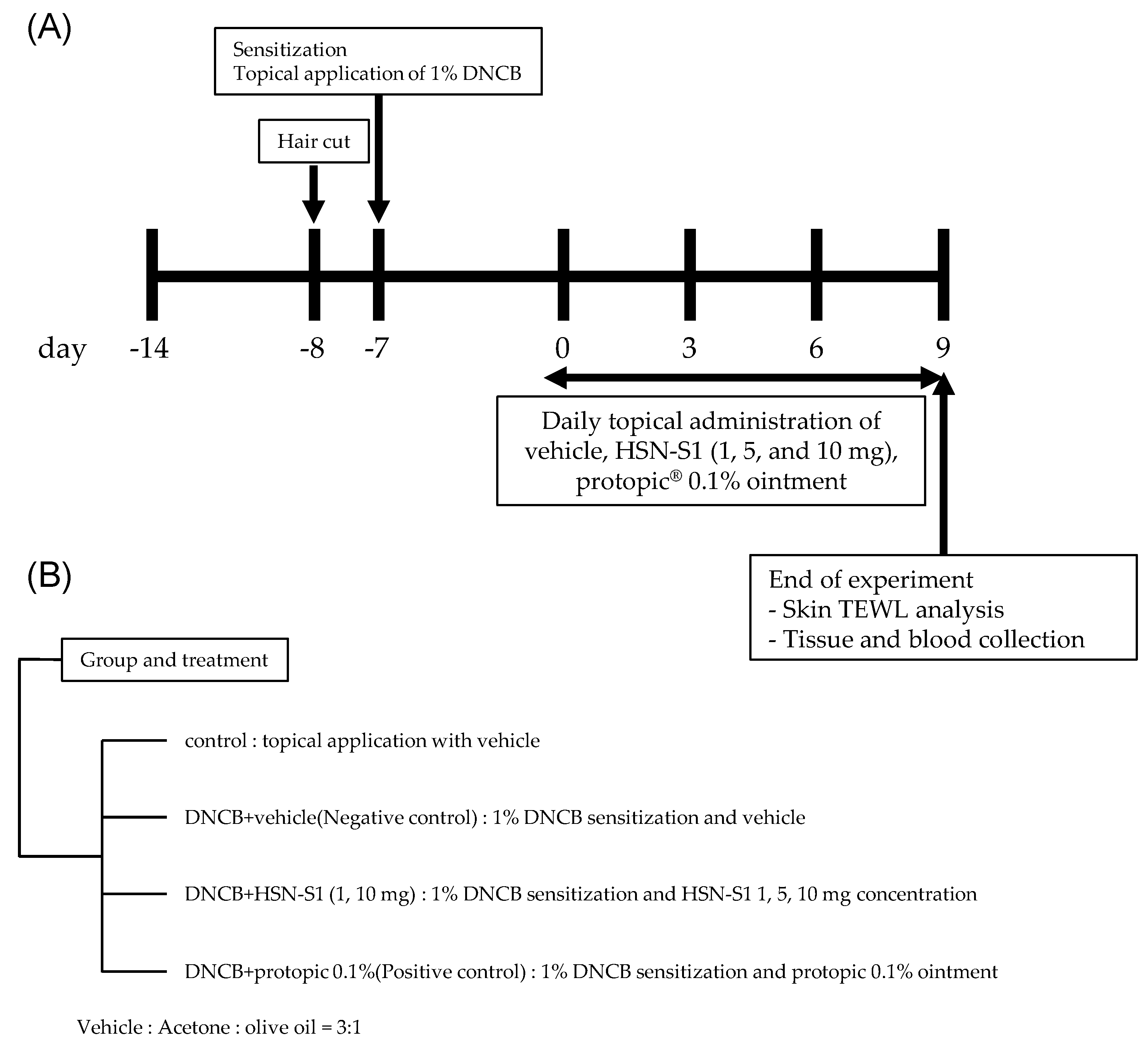
2.9. In Vivo Efficacy of HSN-S1: Th Cell-Mediated Cytokine Modulation and Serum Immunoglobulin (IgE) Suppression by HSN-S1 in AD Models
3. Materials and Methods
3.1. Compound Extraction and Characterization
3.1.1. Anti-AD Bioactivity-Guided Isolation of H. fusiformis Compounds
3.1.2. Chromatographic Analysis and Sample Preparation
3.1.3. NMR and Mass Spectrometry Analysis
3.2. In Vitro Assays for Immunomodulatory and Anti-Bacterial Activity
3.2.1. Culture of AD-Related Cell Lines
3.2.2. Splenocyte Culture
3.2.3. THP-1 Lucia™ NF-κB and Jurkat Lucia™ NFAT Luciferase Assay
3.2.4. NO and Antimicrobial Susceptibility Assay
3.2.5. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
3.2.6. Western Blot Analysis
3.2.7. Confocal Microscopy Analysis
3.3. In Vivo Efficacy Evaluation
3.3.1. Animal Care and AD-like Skin Lesion Induction
3.3.2. Histological Analysis
3.3.3. TEWL Analysis
3.3.4. Serum IgE Measurement in Blood
3.4. Statistical Analysis
4. Conclusion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fiset, P.-O.; Leung, D.Y.M.; Hamid, Q. Immunopathology of atopic dermatitis. J. Allergy Clin. Immunol. 2006, 118, 287–290. [Google Scholar] [CrossRef]
- Czarnowicki, T.; He, H.; Canter, T.; Han, J.; Lefferdink, R.; Erickson, T.; Rangel, S.; Kameyama, N.; Kim, H.J.; Pavel, A.B.; et al. Evolution of pathologic T-cell subsets in patients with atopic dermatitis from infancy to adulthood. J. Allergy Clin. Immunol. 2020, 145, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Wang, V.; Boguniewicz, J.; Boguniewicz, M.; Ong, P.Y. The infectious complications of atopic dermatitis. Ann. Allergy Asthma Immunol. 2021, 126, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Cardona, I.D.; Cho, S.H.; Leung, D.Y.M. Role of bacterial superantigens in atopic dermatitis: Implications for future therapeutic strategies. Am. J. Clin. Dermatol. 2006, 7, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci. Transl. Med. 2017, 9, eaah4680. [Google Scholar] [CrossRef]
- Lyons, J.J.; Milner, J.D.; Stone, K.D. Atopic dermatitis in children: Clinical features, pathophysiology, and treatment. Immunol. Allergy Clin. North Am. 2015, 35, 161–183. [Google Scholar] [CrossRef] [PubMed]
- Carr, W.W. Topical calcineurin inhibitors for atopic dermatitis: Review and treatment recommendations. Paediatr. Drugs 2013, 15, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, A.; Barbarot, S.; Bieber, T.; Christen-Zaech, S.; Deleuran, M.; Fink-Wagner, A.; Gieler, U.; Girolomoni, G.; Lau, S.; Muraro, A.; et al. Consensus-based European guidelines for treatment of atopic eczema (atopic dermatitis) in adults and children: Part II. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 850–878. [Google Scholar] [CrossRef]
- Clowry, J.; Irvine, A.D.; McLoughlin, R.M. Next-generation anti-Staphylococcus aureus vaccines: A potential new therapeutic option for atopic dermatitis? J. Allergy Clin. Immunol. 2019, 143, 78–81. [Google Scholar] [CrossRef]
- Harbige, L.S. Fatty acids, the immune response, and autoimmunity: A question of n-6 essentiality and the balance between n-6 and n-3. Lipids 2003, 38, 323–341. [Google Scholar] [CrossRef]
- Clarke, S.R.; Mohamed, R.; Bian, L.; Routh, A.F.; Kokai-Kun, J.F.; Mond, J.J.; Tarkowski, A.; Foster, S.J. The Staphylococcus aureus surface protein IsdA mediates resistance to innate defenses of human skin. Cell Host Microbe 2007, 1, 199–212. [Google Scholar] [CrossRef]
- Domenico, E.G.D.; Cavallo, I.; Bordignon, V.; Prignano, G.; Sperduti, I.; Gurtner, A.; Trento, E.; Toma, L.; Pimpinelli, F.; Capitanio, B.; et al. Inflammatory cytokines and biofilm production sustain Staphylococcus aureus outgrowth and persistence: A pivotal interplay in the pathogenesis of Atopic Dermatitis. Sci. Rep. 2018, 8, 9573. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.B.; Ebersole, J.L. A novel bioactivity of omega-3 polyunsaturated fatty acids and their ester derivatives. Mol. Oral. Microbiol. 2010, 25, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Neumann, Y.; Ohlsen, K.; Donat, S.; Engelmann, S.; Kusch, H.; Albrecht, D.; Cartron, M.; Hurd, A.; Foster, S.J. The effect of skin fatty acids on Staphylococcus aureus. Arch. Microbiol. 2015, 197, 245–267. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Lawlor, K.C. Antibacterial activity of long-chain polyunsaturated fatty acids against Propionibacterium acnes and Staphylococcus aureus. Mar. Drugs 2013, 11, 4544–4557. [Google Scholar] [CrossRef]
- Plichta, J.K.; Radek, K.A. Sugar-coating wound repair: A review of FGF-10 and dermatan sulfate in wound healing and their potential application in burn wounds. J. Burn. Care Res. 2012, 33, 299–310. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-kappaB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef]
- Das, P.; Mounika, P.; Yellurkar, M.L.; Prasanna, V.S.; Sarkar, S.; Velayutham, R.; Arumugam, S. Keratinocytes: An Enigmatic Factor in Atopic Dermatitis. Cells 2022, 11, 1683. [Google Scholar] [CrossRef] [PubMed]
- Piipponen, M.; Li, D.; Landén, N.X. The Immune Functions of Keratinocytes in Skin Wound Healing. Int. J. Mol. Sci. 2020, 21, 8790. [Google Scholar] [CrossRef]
- Mehta, N.N.; Teague, H.L.; Swindell, W.R.; Baumer, Y.; Ward, N.L.; Xing, X.; Baugous, B.; Johnston, A.; Joshi, A.A.; Silverman, J.; et al. IFN-gamma and TNF-alpha synergism may provide a link between psoriasis and inflammatory atherogenesis. Sci. Rep. 2017, 7, 13831. [Google Scholar] [CrossRef]
- Kim, H.J.; Baek, J.; Lee, J.R.; Roh, J.Y.; Jung, Y. Optimization of Cytokine Milieu to Reproduce Atopic Dermatitis-related Gene Expression in HaCaT Keratinocyte Cell Line. Immune Netw. 2018, 18, e9. [Google Scholar] [CrossRef]
- Kasraie, S.; Werfel, T. Role of macrophages in the pathogenesis of atopic dermatitis. Mediators Inflamm. 2013, 2013, 942375. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Yang, Y.; Li, Y.; Xu, L.; Wang, Y.; Guo, Z.; Song, H.; Yang, M.; Luo, B.; Zheng, A.; et al. Ephedrine hydrochloride inhibits PGN-induced inflammatory responses by promoting IL-10 production and decreasing proinflammatory cytokine secretion via the PI3K/Akt/GSK3beta pathway. Cell. Mol. Immunol. 2013, 10, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Sroka-Tomaszewska, J.; Trzeciak, M. Molecular Mechanisms of Atopic Dermatitis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 4130. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Bi, D.; Zheng, R.; Cai, N.; Xu, H.; Zhou, R.; Lu, J.; Wan, M.; Xu, X. Identification and activation of TLR4-mediated signalling pathways by alginate-derived guluronate oligosaccharide in RAW264.7 macrophages. Sci. Rep. 2017, 7, 1663. [Google Scholar] [CrossRef] [PubMed]
- Olona, A.; Hateley, C.; Muralidharan, S.; Wenk, M.R.; Torta, F.; Behmoaras, J. Sphingolipid metabolism during Toll-like receptor 4 (TLR4)-mediated macrophage activation. Br. J. Pharmacol. 2021, 178, 4575–4587. [Google Scholar] [CrossRef] [PubMed]
- Boyman, O.; Werfel, T.; Akdis, C.A. The suppressive role of IL-10 in contact and atopic dermatitis. J. Allergy Clin. Immunol. 2012, 129, 160–161. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, R.; Kuchen, S.; Lipsky, P.E. The role of IL-21 in regulating B-cell function in health and disease. Immunol. Rev. 2008, 223, 60–86. [Google Scholar] [CrossRef] [PubMed]
- Piruzyan, M.; Shitanda, I.; Shimauchi, Y.; Okita, G.; Tsurekawa, Y.; Moriuchi, M.; Nakano, Y.; Teramoto, K.; Suico, M.A.; Shuto, T.; et al. A novel condition of mild electrical stimulation exerts immunosuppression via hydrogen peroxide production that controls multiple signaling pathway. PLoS ONE 2020, 15, e0234867. [Google Scholar] [CrossRef]
- Huang, L.; Pi, J.; Wu, J.; Zhou, H.; Cai, J.; Li, T.; Liu, L. A rapid and sensitive assay based on particle analysis for cell degranulation detection in basophils and mast cells. Pharmacol. Res. 2016, 111, 374–383. [Google Scholar] [CrossRef]
- Geoghegan, J.A.; Irvine, A.D.; Foster, T.J. Staphylococcus aureus and Atopic Dermatitis: A Complex and Evolving Relationship. Trends Microbiol. 2018, 26, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Lacey, K.A.; Mulcahy, M.E.; Towell, A.M.; Geoghegan, J.A.; McLoughlin, R.M. Clumping factor B is an important virulence factor during Staphylococcus aureus skin infection and a promising vaccine target. PLoS Pathog. 2019, 15, e1007713. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, L.M.; Walsh, E.J.; Massey, R.C.; Peacock, S.J.; Foster, T.J. Staphylococcus aureus clumping factor B (ClfB) promotes adherence to human type I cytokeratin 10: Implications for nasal colonization. Cell Microbiol. 2002, 4, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, D. Topical Antibacterials in Dermatology. Indian. J. Dermatol. 2021, 66, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Ogonowska, P.; Gilaberte, Y.; Barańska-Rybak, W.; Nakonieczna, J. Colonization With Staphylococcus aureus in Atopic Dermatitis Patients: Attempts to Reveal the Unknown. Front. Microbiol. 2020, 11, 567090. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, M. The Role of Th17-Related Cytokines in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 1314. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.; Lu, J.; Choi, E.B.; Oh, M.H.; Jeong, M.; Barmettler, S.; Zhu, Z.; Zheng, T. Expression of IL-22 in the Skin Causes Th2-Biased Immunity, Epidermal Barrier Dysfunction, and Pruritus via Stimulating Epithelial Th2 Cytokines and the GRP Pathway. J. Immunol. 2017, 198, 2543–2555. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.J.; Luo, C.-H.; Thio, C.L.-P.; Chang, Y.-J. Immunomodulatory Role of Staphylococcus aureus in Atopic Dermatitis. Pathogens 2022, 11, 422. [Google Scholar] [CrossRef]
- Yang, G.; Seok, J.K.; Kang, H.C.; Cho, Y.-Y.; Lee, H.S.; Lee, J.Y. Skin Barrier Abnormalities and Immune Dysfunction in Atopic Dermatitis. Int. J. Mol. Sci. 2020, 21, 2867. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Atom Position | 1H NMR δH (ppm), Multiplicity (J in Hz) | 13C NMR δC (ppm) |
|---|---|---|
| 1 | - | 173.0 |
| 2 | 2.30, t (7.0) | 33.4 |
| 3 | 1.54, quint (7.0) | 26.4 |
| 4 | 1.34, quint (7.0) | 28.5 |
| 5 | 2.05, t (7.0) | 24.1 |
| 6 | 5.35–5.34, m | 126.9 |
| 7 | 5.34–5.30, m | 127.7 |
| 8 | 2.82–2.79, m | 25.2 |
| 9 | 5.34–5.30, m | 127.7 |
| 10 | 5.34–5.30, m | 127.8 |
| 11 | 2.82–2.79, m | 25.2 |
| 12 | 5.34–5.30, m | 128.0 |
| 13 | 5.34–5.30, m | 128.1 |
| 14 | 2.82–2.79, m | 25.1 |
| 15 | 5.30–5.28, m | 129.6 |
| 16 | 5.37–5.35, m | 131.5 |
| 17 | 2.03, t (7.0) | 20.0 |
| 18 | 0.92, t (7.7) | 14.1 |
| 1’ | 4.28, dd (11.2, 2.8); 3.94, dd (11.2, 7.0) | 66.9 |
| 2’ | 3.66, m | 68.3 |
| 3’ | 3.55, d (9.1) | 69.5 |
| 4’ | 3.55, d (9.1) | 71.2 |
| 5’ | 3.45, m | 71.2 |
| 6’ | 3.39, dd (11.2, 6.3); 3.61, dd (11.2, 3.5) | 63.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.S.; Ahn, J.W.; Ha, N.R.; Damodar, K.; Jang, S.K.; Yoo, Y.-M.; Gyoung, Y.S.; Joo, S.S. Antibacterial and Immunosuppressive Effects of a Novel Marine Brown Alga-Derived Ester in Atopic Dermatitis. Mar. Drugs 2024, 22, 354. https://doi.org/10.3390/md22080354
Kim HS, Ahn JW, Ha NR, Damodar K, Jang SK, Yoo Y-M, Gyoung YS, Joo SS. Antibacterial and Immunosuppressive Effects of a Novel Marine Brown Alga-Derived Ester in Atopic Dermatitis. Marine Drugs. 2024; 22(8):354. https://doi.org/10.3390/md22080354
Chicago/Turabian StyleKim, Hyun Soo, Jeong Won Ahn, Na Reum Ha, Kongara Damodar, Su Kil Jang, Yeong-Min Yoo, Young Soo Gyoung, and Seong Soo Joo. 2024. "Antibacterial and Immunosuppressive Effects of a Novel Marine Brown Alga-Derived Ester in Atopic Dermatitis" Marine Drugs 22, no. 8: 354. https://doi.org/10.3390/md22080354
APA StyleKim, H. S., Ahn, J. W., Ha, N. R., Damodar, K., Jang, S. K., Yoo, Y.-M., Gyoung, Y. S., & Joo, S. S. (2024). Antibacterial and Immunosuppressive Effects of a Novel Marine Brown Alga-Derived Ester in Atopic Dermatitis. Marine Drugs, 22(8), 354. https://doi.org/10.3390/md22080354